
Construction of a Novel Chimeric Dextransucrase Fused to the Carbohydrate-Binding Module CBM2a

 , and
, and
Abstract
:
1. Introduction
2. Results and Discussion
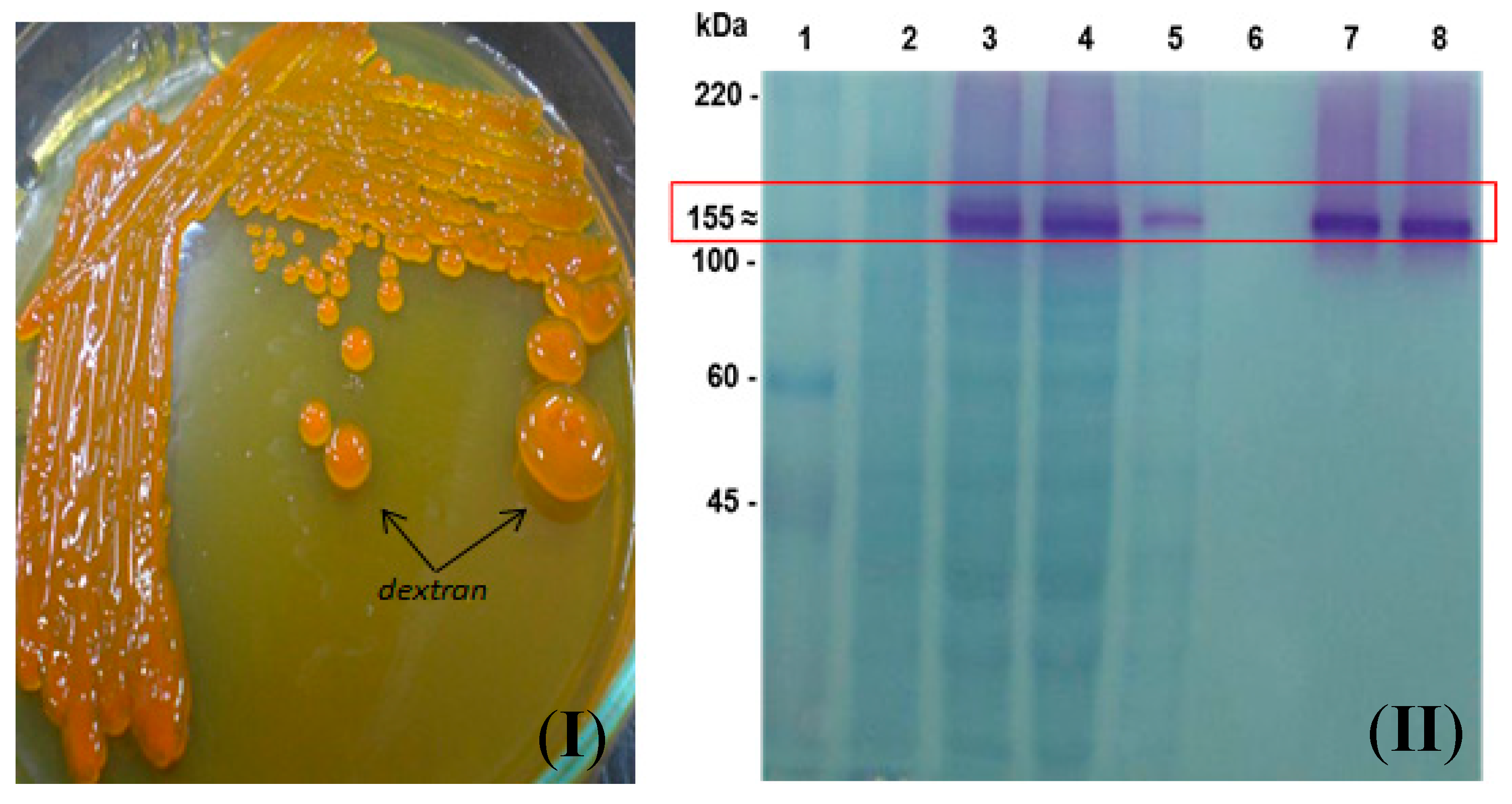
2.1. Design of a Chimeric Dextransucrase (DSR-F-∆SP-∆GBD-CBM2a) Fused to the Carbohydrate-Binding Module CBM2a
2.2. Subcloning of DSR-F-∆SP-∆GBD to Obtain DSR-F-∆SP-∆GBD-CBM2a in the Expression Vector pdsrF-CBM2a and Purification of the Fusion Protein DSR-F-∆SP-∆GBD-CBM2a
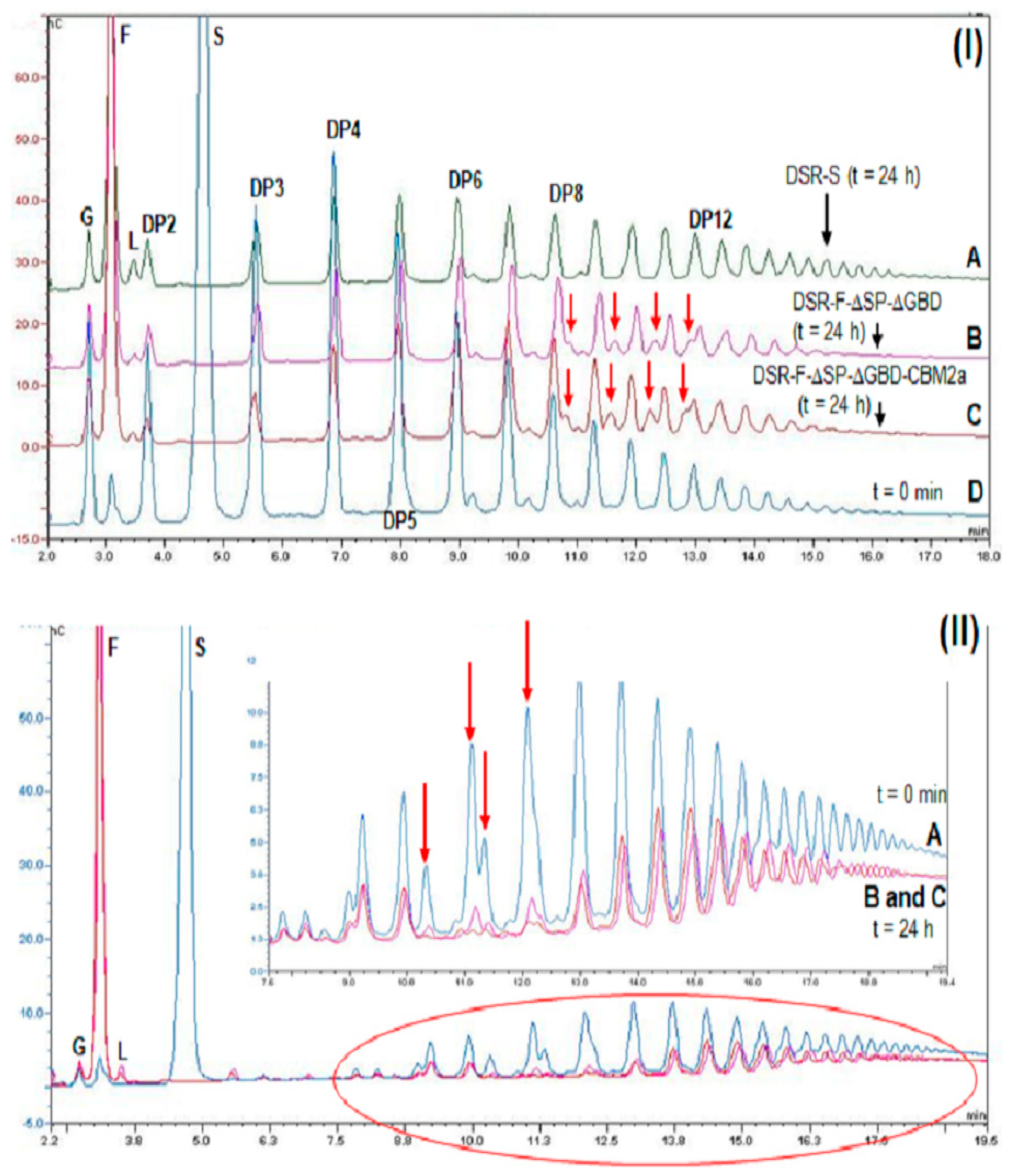
2.3. Action of DSR-F-ΔSP-ΔGBD-CBM2a in Polymerization and Acceptor Reactions
2.4. Action of DSR-F-ΔSP-ΔGBD-CBM2a from Sucrose and Linear Oligodextrans (1500 and 6000 g·mol−1)
3. Materials and Methods
3.1. Bacterial Strains and Culture Media
3.2. Subcloning to Express the dsrF-ΔSP-ΔGBD-CBM2a Gene in E. coli
3.3. Inducible Production of DSR-F-ΔSP-ΔGBD-CBM2a in E. coli
3.4. Cellulose Affinity Chromatography (CAC)
3.5. Determination of Enzymatic Activity Dextransucrase
3.6. SDS-PAGE and Zymograms
3.7. Production of High-Molecular Weight Dextrans, Maltooligosaccharides, and Linear Dextran Modification
3.8. High-Performance Anion-Exchange Chromatography with Pulsed Amperometric Detection (HPAEC-PAD)
3.9. High-Performance Size Exclusion Chromatography (HPSEC)
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pu, Y.; Zou, Q.; Hou, D.; Zhang, Y.; Chen, S. Molecular weight kinetics and chain scission models for dextran polymers during ultrasonic degradation. Carbohydr. Polym. 2017, 156, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Zannini, E.; Waters, D.M.; Coffey, A.; Arendt, E.K. Production, properties, and industrial food application of lactic acid bacteria-derived exopolysaccharides. Appl. Microbiol. Biotechnol. 2016, 100, 1121–1135. [Google Scholar] [CrossRef]
- Vettori, M.H.P.B.; Blanco, K.C.; Cortezi, M.; De Lima, C.J.B.; Contiero, J. Dextran: Effect of process parameters on production, purification and molecular weight and recent applications. Diálogos Cienc. 2012, 171–186. [Google Scholar] [CrossRef]
- Ryan, P.M.; Ross, R.P.; Fitzgerald, G.F.; Caplice, N.M.; Stanton, C. Sugar-coated: Exopolysaccharide producing lactic acid bacteria for food and human health applications. Food Funct. 2015, 6, 679–693. [Google Scholar] [CrossRef] [PubMed]
- Salazar, N.; Gueimonde, M.; De Los Reyes-Gavilán, C.G.; Ruas-Madiedo, P. Exopolysaccharides Produced by Lactic Acid Bacteria and Bifidobacteria as Fermentable Substrates by the Intestinal Microbiota. Crit. Rev. Food Sci. Nutr. 2016, 56, 1440–1453. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Zhang, J. Bacterial glucans: Production, properties, and applications. Appl. Microbiol. Biotechnol. 2016, 100, 9023–9036. [Google Scholar] [CrossRef] [PubMed]
- Naessens, M.; Cerdobbek, A.; Soetaert, W.; Vandamme, E.J. Leuconostoc dextransucrase and dextran: Production, properties and applications. J. Chem. Technol. Biotechnol. 2005, 80, 845–860. [Google Scholar] [CrossRef]
- Chen, Z.; Ni, D.; Zhang, W.; Stressler, T.; Mu, W. Lactic acid bacteria-derived α-glucans: From enzymatic synthesis to miscellaneous applications. Biotechnol. Adv. 2021, 47, 107708. [Google Scholar] [CrossRef]
- Badel, S.; Bernardi, T.; Michaud, P. New perspectives for Lactobacilli exopolysaccharides. Biotechnol. Adv. 2011, 29, 54–66. [Google Scholar] [CrossRef]
- Zdolsek, H.J.; Vegfors, M.; Lindahl, T.L.; Tornquist, T.; Bortnik, P.; Hahn, R.G. Hydroxyethyl starches and dextran during hip replacement surgery: Effects on blood volume and coagulation. Acta Anaesthesiol. Scand. 2011, 55, 677–685. [Google Scholar] [CrossRef]
- Monchois, V.; Remaud-Simeon, M.; Russel, R.R.; Monsan, P.; Willemot, R.M. Characterization of Leuconostoc mesenteroides NRRL B-512F dextransucrase (DSRS) and identification of amino-acid residues playing a key role in enzyme activity. Appl. Microbiol. Biotechnol. 1997, 48, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Lombard, V.; Golaconda Ramulu, H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The Carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 2014, 42, D490–D495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passerini, D.; Vuillemin, M.; Ufarté, L.; Morel, S.; Loux, V.; Fontagné-Faucher, C.; Monsan, P.; Remaud-Siméon, M.; Moulis, C. Inventory of the GH70 enzymes encoded by Leuconostoc citreum NRRL B-1299–identification of three novel-transglucosylases. FEBS J. 2015, 282, 2115–2130. [Google Scholar] [CrossRef] [PubMed]
- Gangoiti, J.; van Leeuwen, S.; Gerwig, G.; Duboux, S.; Vafiadi, C.; Pijning, T.; Dijkhuizen, L. 4,3-α-Glucanotransferase, a novel reaction specificity in glycoside hydrolase family 70 and clan GH-H. Sci. Rep. 2017, 7, 39761. [Google Scholar] [CrossRef] [Green Version]
- Macgregor, E.A.; Jespersen, H.M.; Svensson, B. A circularly permuted alpha-amylase-type alpha/beta-barrel structure in glucan-synthesizing glucosyltransferases. FEBS Lett. 1996, 378, 263–266. [Google Scholar] [CrossRef] [Green Version]
- Vujicić-Zagar, A.; Dijkstra, B.W. Monoclinic crystal form of Aspergillus niger alpha-amylase in complex with maltose at 1.8 angstroms resolution. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2006, 62, 716–721. [Google Scholar] [CrossRef] [Green Version]
- Pijning, T.; Vujicic-Zagar, A.; Kralj, S.; Eeuwema, W.; Dijkhuizen, L.; Dijkstra, B.W. Biochemical and crystallographic characterization of a glucansucrase from Lactobacillus reuteri 180. Biocat. Biotrans. 2008, 26, 12–17. [Google Scholar] [CrossRef] [Green Version]
- Molina, M.; Cioci, G.; Moulis, C.; Séverac, E.; Remaud-Simeon, M. Bacterial α-Glucan and branching sucrases from GH70 Family: Discovery, structure-function relationship studies and engineering. Microorganisms 2021, 9, 1607. [Google Scholar] [CrossRef]
- Fraga, R.; Moulis, C.; Escalier, P.; Remaud-Simeón, M.; Monsan, P. Isolation of a Gene from Leuconostoc citreum B/110-1-2 Encoding a Novel Dextransucrase Enzyme. Curr. Microbiol. 2011, 62, 1260–1266. [Google Scholar] [CrossRef]
- Fraga, R.; Martínez, A.; Moulis, C.; Escalier, P.; Morel, S.; Remaud-Siméon, M.; Monsan, P. A novel dextransucrase is produced by Leuconostoc citreum strain B/110-1-2: An isolate used for the industrial production of dextran and dextran-derivatives. J. Ind. Microbiol. Biotechnol. 2011, 38, 1499–1503. [Google Scholar] [CrossRef]
- Fraga, R.; Pacios, S.; Arísticas, R.C.; Martínez, L.; Lafargue, M.; Montes, A.; Remaud-Simeon, M.; Monsan, P. Cloning and Partial Characterization of an Extracellular Dextransucrase Coding Region (DSR-V) from Leuconostoc citreum M-3. In Microbial Exopolysaccharides: Current Research and Developments, 1st ed.; Duru, Ö.A., Ed.; Caister Academic Press: Norfolk, UK, 2019; Chapter 11; pp. 295–314. [Google Scholar] [CrossRef]
- Joucla, G.; Pizzut, S.; Monsan, P.; Remaud-Simeon, M. Construction of a fully active truncated alternansucrase partially deleted of its carboxy-terminal domain. FEBS Lett. 2006, 580, 763–768. [Google Scholar] [CrossRef] [Green Version]
- Olivares-Illana, V.; López-Munguía, A.; Olvera, C. Molecular characterization of inulosucrase from Leuconostoc citreum: A fructosyltransferase within a glucosyltransferase. J. Bacteriol. 2003, 185, 3606–3612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vuillemin, M.; Claverie, M.; Brison, Y.; Séverac, E.; Bondy, P.; Morel, S.; Monsan, P.; Moulis, C.; Remaud-Siméon, M. Characterization of the first α-(1→3) branching sucrases of the GH70 family. J. Biol. Chem. 2016, 291, 7687–7702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parlak, M.; Ustek, D.; Tanriseven, A. Designing of a novel dextransucrase efficient in acceptor reactions. Carbohydr. Res. 2014, 386, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Wang, Y.; Ye, X.; Percival Zhang, Y.H. Simple protein purification through affinity adsorption on regenerated amorphous cellulose followed by intein self-cleavage. J. Chromatogr. A 2008, 1194, 150–154. [Google Scholar] [CrossRef]
- Meng, X.; Dobruchowska, J.M.; Pijning, T.; Gerwig, G.J.; Kamerling, J.P.; Dijkhuizen, L. Truncation of domain V of the multidomain glucansucrase GTF180 of Lactobacillus reuteri 180 heavily impairs its polysaccharide-synthesizing ability. Appl. Microbiol. Biotechnol. 2015, 99, 5885–5894. [Google Scholar] [CrossRef]
- Molina, M.; Moulis, C.; Monties, N.; Pizzut-Serin, S.; Guieysse, D.; Morel, S.; Cioci, G.; Remaud-Simeon, M. Deciphering an undecided enzyme: Investigations of the structural determinants involved in the linkage specificity of alternansucrase. ACS Catal. 2019, 9, 2222–2237. [Google Scholar] [CrossRef]
- Moulis, C.; Joucla, G.; Harrison, D.; Fabre, E.; Potocki-Veronese, G.; Monsan, P.; Remaud-Simeon, M. Understanding the polymerization mechanism of glycoside-hydrolase family 70 glucansucrases. J. Biol. Chem. 2006, 281, 31254–31267. [Google Scholar] [CrossRef] [Green Version]
- Molina, M.; Moulis, C.; Monties, N.; David, G.; Morel, S.; Cioci, G.; Remaud-Simeon, M. A specific oligosaccharide-binding site in the alternansucrase catalytic domain mediates alternan elongation. J. Biol. Chem. 2020, 295, 9474–9489. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, H.M.; Hellman, N.N.; Koepsell, H.J. Factors affecting the molecular weight of enzymatically synthesized dextran. J. Am. Chem. Soc. 1953, 75, 757–758. [Google Scholar] [CrossRef]
- Bechtner, J.; Hassler, V.; Wefers, D.; Vogel, R.F.; Jakob, F. Insights into extracellular dextran formation by Liquorilactobacillus nagelii TMW 1.1827 using secretomes obtained in the presence or absence of sucrose. Enzyme Microb. Technol. 2021, 143, 109724. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-H.P.; Cui, J.; Lynd, L.R.; Kuang, L.R. A transition from cellulose swelling to cellulose dissolution by o-phosphoric acid: Evidence from enzymatic hydrolysis and supramolecular structure. Biomacromolecules 2006, 7, 644–648. [Google Scholar] [CrossRef]
- Sumner, J.; Howell, S. A method for determination of invertase activity. J. Biol. Chem. 1935, 108, 51–54. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Miller, A.W.; Robyt, J.F. Detection of dextransucrase and levansucrase on polyacrylamide gels by the periodic acid-Schiff stain: Staining artifacts and their prevention. Anal. Biochem. 1986, 156, 357–363. [Google Scholar] [CrossRef]
- Fernandez-Patrón, C.; Hardy, E.; Seoane, J.; Castellanos, L. Double staining of Coomassie blue-stained polyacrylamide gels by imidazole-sodium dodecyl sulfate-zinc reverse staining: Sensitive detection of Coomassie blue-undetected proteins. Anal. Biochem. 1995, 224, 263–269. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plasmids | Description | Reference |
|---|---|---|
| pET38b(+) | Replicon colE1, vector used for protein production in E. coli, fused to the signal secretion Cex downstream the PT7lac promoter. It harbours the encoding region for the Carbohydrate-Binding Module. (CBM2a) and a His8 tag, Kmr, size: 5.8 kb. | Novagen |
| pSE380 | Replicon colE1, vector used for protein production in E. coli downstream the PTrc promoter, Apr, size: 4.4 kb. | Invitrogen |
| pSEdsrF | pSE380 NcoI-EcoRI, fused to the 3.7 kb amplicon NcoI-EcoRI obtained from pGEMdsrF. Harbours the truncated variant DsrF-∆SP-∆GBD, Apr, size: 8.1 kb | [19] |
| pETdsrF | pET38b(+) NcoI-EcoRI, fused to the 3.7 kb (DsrF-∆SP-∆GBD) NcoI-EcoRI from pSEdsrF, Kmr, size: 9.5 kb. | This work |
| pdsrF-CBM2a | pSE380 NcoI-AvrII, fused to the 4.2 kb DNA fragment (DsrF-∆SP-∆GBD-CBM2a) NcoI-AvrII from pETdsrF, Apr, size: 8.6 kb. | This work |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fraga Vidal, R.; Arísticas Ribalta, R.C.; Martínez Valdés, L.T.; Lafargue Gámez, M.; Montes Alvarez, A.; Rubio Sánchez, A.; Dubreucq, E.; Moreau, B. Construction of a Novel Chimeric Dextransucrase Fused to the Carbohydrate-Binding Module CBM2a. Catalysts 2021, 11, 1179. https://doi.org/10.3390/catal11101179
Fraga Vidal R, Arísticas Ribalta RC, Martínez Valdés LT, Lafargue Gámez M, Montes Alvarez A, Rubio Sánchez A, Dubreucq E, Moreau B. Construction of a Novel Chimeric Dextransucrase Fused to the Carbohydrate-Binding Module CBM2a. Catalysts. 2021; 11(10):1179. https://doi.org/10.3390/catal11101179
Chicago/Turabian StyleFraga Vidal, Reinaldo, Roberto Carlos Arísticas Ribalta, Lisandra Teresa Martínez Valdés, Meinardo Lafargue Gámez, Amanda Montes Alvarez, Arianne Rubio Sánchez, Eric Dubreucq, and Benoît Moreau. 2021. "Construction of a Novel Chimeric Dextransucrase Fused to the Carbohydrate-Binding Module CBM2a" Catalysts 11, no. 10: 1179. https://doi.org/10.3390/catal11101179