Immobilization of Old Yellow Enzymes via Covalent or Coordination Bonds
by
 , , ,
, , ,
Francesca Tentori
1, 
Teodora Bavaro
2 ,
,
Elisabetta Brenna
1,3,*,
Danilo Colombo
1,
Daniela Monti
3,
Riccardo Semproli
2 and
Daniela Ubiali
2,* 1
Department of Chemistry, Materials and Chemical Engineering “Giulio Natta”, Politecnico di Milano, via Mancinelli 7, 20131 Milano, Italy
2
Department of Drug Sciences, Università degli Studi di Pavia, viale Taramelli 12, 27100 Pavia, Italy
3
Istituto di Scienze e Tecnologie Chimiche “Giulio Natta”, CNR, via Bianco 9, 20131 Milano, Italy
*
Authors to whom correspondence should be addressed.
Catalysts 2020, 10(2), 260; https://doi.org/10.3390/catal10020260
Submission received: 30 January 2020
/
Revised: 15 February 2020
/
Accepted: 17 February 2020
/
Published: 20 February 2020
(This article belongs to the Special Issue Immobilization of Enzymes)
Abstract
:Ene-reductases (ERs) belonging to the old yellow enzyme (OYE) family have been thoroughly investigated for the stereospecific reduction of activated prochiral C=C double bonds. In this work, OYE3 was immobilized both by covalent binding on glyoxyl-agarose (OYE3-GA), and by affinity-based adsorption on EziGTM particles (OYE3-EziG). The immobilized OYE3-GA was demonstrated to be active (activity recovery = 52%) and to retain almost 100% of its activity under the enzymatic assay conditions (50 mM phosphate buffer pH 7, 28 °C) for six days, whereas the activity of the non-immobilized enzyme dropped to 50% after two days. In the case of EziGTM, the highest activity recovery (54%) was achieved by using the most hydrophilic carrier (EziGTM Opal) that was selected for the full characterization of this type of enzyme preparation (stability, recycling, re-use, enzyme leakage). OYE3-EziG was slightly less stable than OYE3-GA under the same experimental conditions. OYE3-GA could be recycled and re-used for up to 12 reaction cycles in the bioreduction of α-methyl-trans-cinnamaldehyde; after 12 runs, the highest conversion achieved was 40%. In the case of the co-immobilized OYE3/GDH-EziG, the conversion dropped to 56% after two reaction cycles. No enzyme leakage was detected over 48 h for both OYE3-GA and OYE3/GDH-EziG (50 mM phosphate buffer pH 7, 28 °C). These seed results pave the way for a true optimization of the immobilization of OYE3, as well as for the use of immobilized OYE3 for preparative applications both in batch and continuous flow conditions.
1. Introduction
Ene-reductases (ERs) belonging to the old yellow enzyme (OYE) family (EC 1.6.99.1) are flavin mononucleotide (FMN)-containing oxidoreductases, which are able to catalyze the stereoselective reduction of C=C double bonds activated by the presence of a suitable electron-withdrawing group (EWG) as a substituent of the alkene moiety [1,2]. The reaction occurs through a stereospecific anti hydrogen addition to the C=C double bond, according to the following steps [3]. The enzyme-bound flavin (reduced by NAD(P)H cofactor to FMNH2) transfers a hydride to the olefin carbon atom in β position with respect to the EWG. A proton is delivered by a tyrosine side-chain (Tyr196 for OYE1-3) of the enzyme active site to the α-carbon atom (with respect to EWG) on the opposite face of the alkene. For the application of OYE-mediated reductions in synthetic sequences of practical interest, it is advantageous to promote the in situ regeneration of the reduced nicotinamide cofactor by using a NAD(P)H-dependent glucose dehydrogenase (GDH) with glucose as a sacrificial co-substrate.
The OYE-mediated bioreduction has been widely investigated in the last decade to establish substrate scope and stereoselectivity of these enzymes [4,5,6], and to study the combination of this reaction with other chemo- and/or biocatalyzed transformations, both in concomitant and subsequent cascade procedures [7,8]. Several works showed the advantages of using OYE-catalyzed hydrogenation to prepare chiral building blocks for the synthesis of APIs (Active Pharmaceutical Ingredients) [9,10,11,12], flavors and fragrances [13,14,15].
The synthetic potential of the OYE-mediated bioreduction can be further expanded by OYE immobilization on solid carriers. If OYEs are converted into stable, readily-recoverable and recyclable heterogeneous catalysts, then their suitability for large-scale applications can be enhanced, in agreement with one of the key green chemistry research areas established by the ACS Green Chemistry Institute® Pharmaceutical Roundtable (catalyst immobilization without significant loss in kinetics) [16].
Studies on the immobilization of oxidoreductases are gradually increasing, but still only a few examples of ER immobilization have been described to date. Recently, a recombinant ER from the OYE family and its cofactor-recycling partner enzyme GDH (commercially available from Amano Enzyme Inc.) were co-immobilized by using two immobilization methods (i.e., cross-linked enzyme aggregates (CLEAs) and a so-called “biomimetic” immobilization (BI)) by entrapment within a network of fused silica particles. Activity recovery of both enzyme preparations were about 45%. Immobilization enhanced the thermal stability (50 °C) of both enzymes and markedly increased their resistance to acidic pH (5-6) in comparison with the non-immobilized enzyme. Moreover, both immobilized enzymes were successfully recycled and re-used for up to 14 reaction cycles. The catalytic “performance” of the CLEA preparation in the reduction of 4-(4-methoxyphenyl)-3-buten-2-one resulted to be superior, both to the non-immobilized enzymes and the entrapped enzymes by BI [17].
In a very recent study, an ER from Thermus scotoductus SA-01 (TsOYE) was encapsulated with a light-harvesting dye in an alginate hydrogel for NADH-free, photobiocatalytic asymmetric hydrogenation of 2-methylcyclohexenone. The TsOYE encapsulated in alginate hydrogel exhibited enhanced stability against external stresses (e.g., heat, organic solvents), also on a repeated use. However, the long-term reusability of TsOYE alginate hydrogel was hampered by photobleaching and leakage of the dye used [18].
OYE immobilization is still under-investigated and, to the best of our knowledge, no attempt of immobilization has ever been reported for OYE3, one of the most performing members of this enzyme class. The impressive results obtained in OYE3-catalyzed biotransformations in the past years and the benefits envisaged for ER immobilization urged us to undergo a “seed-study” in this frame.
In this work, OYE3 was immobilized both by covalent binding on glyoxyl-agarose (OYE3-GA), and by metal affinity interaction on EziGTM particles (OYE3-EziG). The GA carrier [19,20], widely explored for the irreversible immobilization of enzymes by multipoint attachment, has proven its efficiency and versatility as immobilization support by affording robust biocatalysts, both for batch and flow applications [21,22]. On the other hand, EziGTM is a novel carrier made of controlled porosity glass (CPG) particles containing chelated Fe(III) for His-tag binding. This carrier can specifically immobilize tagged enzymes from a crude mixture by affinity interaction and is, thus, frequently used to combine immobilization and purification steps [23,24]. In this context, EziGTM was studied to explore a milder immobilization technique based on non-covalent interaction.
2. Results and Discussion
2.1. Enzyme Activity Assays
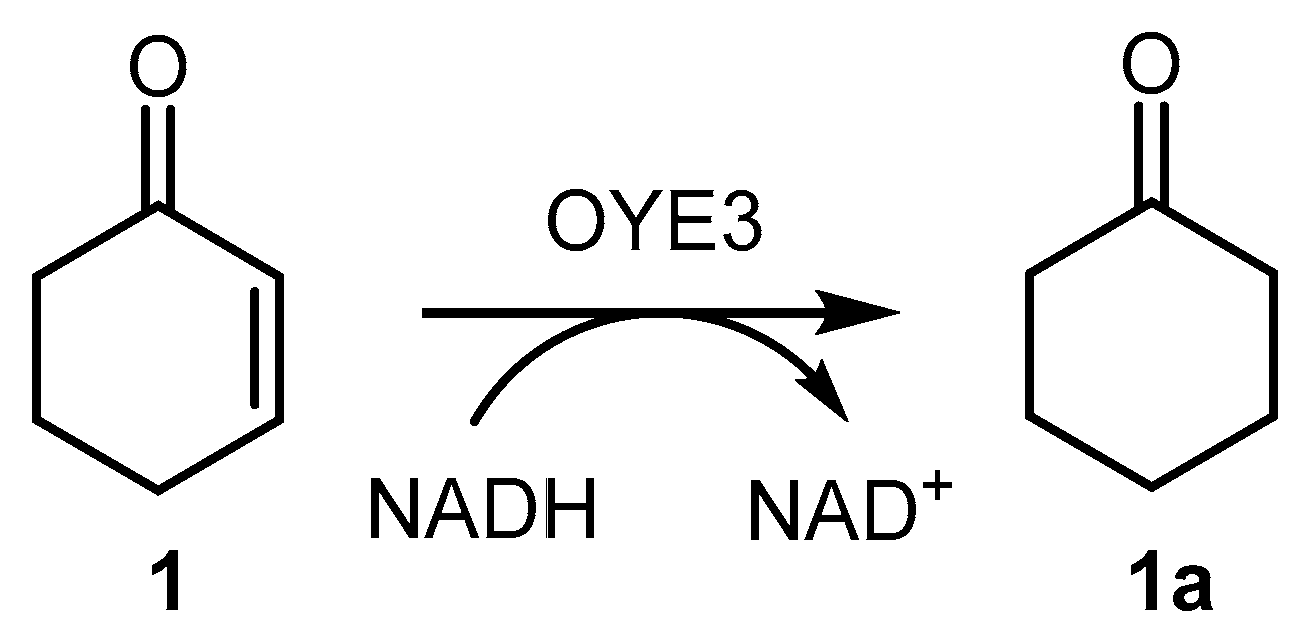
In order to characterize OYE3, the enzyme activity was assessed spectrophotometrically, as described for other ERs [25,26,27], by measuring the consumption of NADH at 340 nm in the reduction of cyclohex-2-enone (1) (see Scheme 1 and Materials and Methods, Section 3.3.1). This assay was found to be suitable for the soluble enzyme, but not for assessing the activity of the immobilized enzyme (and, indeed, to calculate the activity recovery after immobilization) because of the interference of the solid carrier with the spectrophotometric readouts. Therefore, a GC-MS assay (see Scheme 2 and Materials and Methods, Section 3.3.2) based on the reduction of (R)-carvone (2) in the presence of stoichiometric NADH was used. Unless otherwise stated, this latter assay was used to determine the activity of both soluble and immobilized OYE3.
A spectrophotometric assay was also used to assess the activity of soluble GDH by measuring the formation of NADH over time at 340 nm as a result of glucose oxidation (experimental conditions: 50 mM phosphate buffer pH 7, [substrate] = 20 mM, [NAD+] = 0.2 mM, 28 °C, reaction time = 2 min, volume = 1 mL). Glucose dehydrogenase was coupled to OYE3 for cofactor regeneration in the reduction of α-methyl-trans-cinnamaldehyde (3) (see Scheme 3 and Materials and Methods, Section 3.4).
2.2. Preliminary Study of OYE3 Stability at Alkaline pH
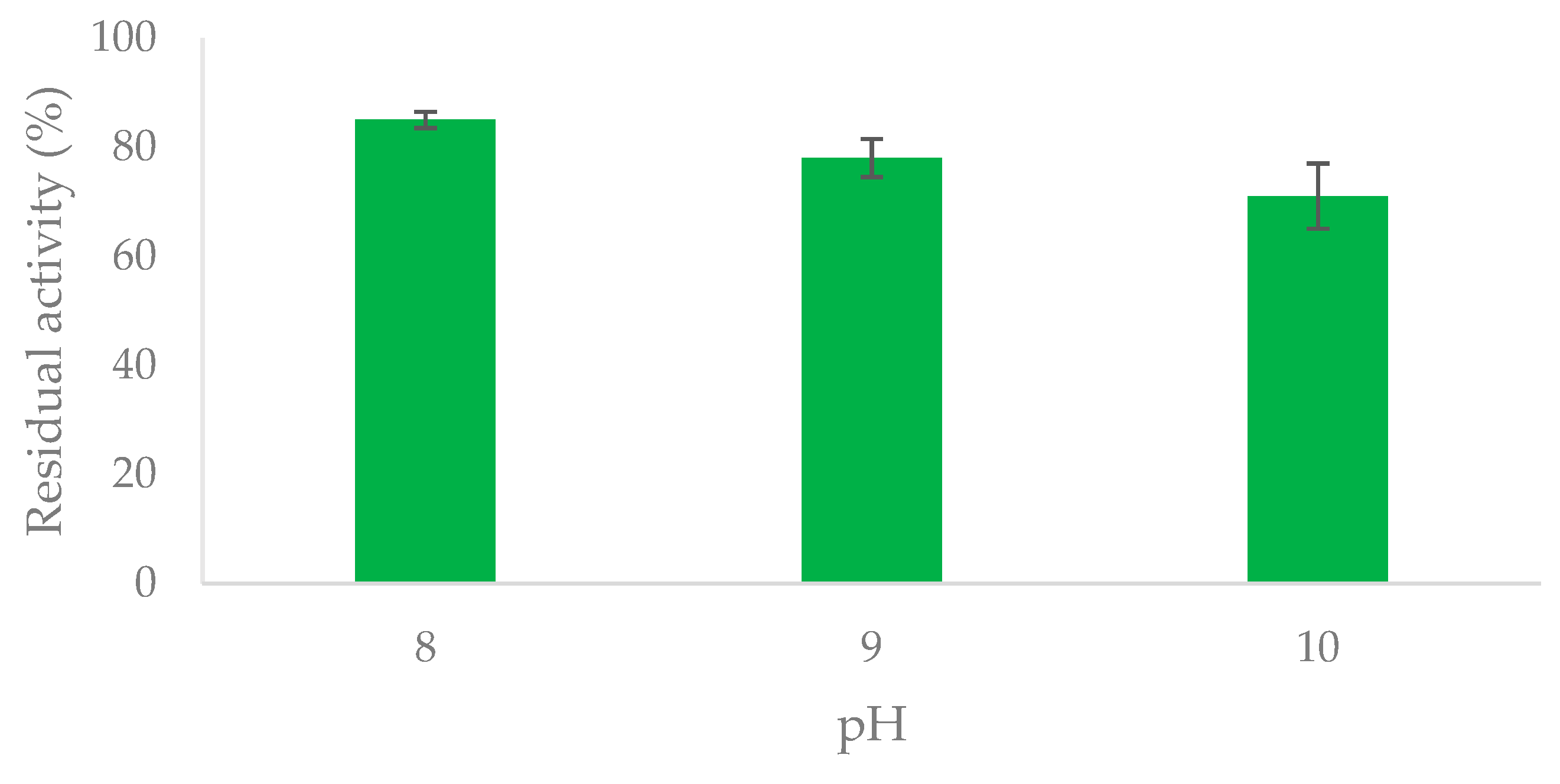
Covalent immobilization of enzymes on glyoxyl-agarose (GA) relies on the formation of imino bonds between the non-protonated ε-amino groups of enzyme Lys residues and the aldehyde groups of the carrier [19,20]. This immobilization is required to be carried out at alkaline pH (i.e., pH ≥ 10) and a final chemical reduction step is necessary for the generation of irreversible C-N bonds. Whenever compatible with the enzyme stability, sodium borohydride is used to this aim, since it transforms Schiff’s bases into secondary amino bonds and the unreacted aldehyde groups of the carrier into inert hydroxyl moieties. Alternatively, the milder sodium cyanoborohydride or 2-picoline borane can be used for those enzymes which are sensitive to sodium borohydride reduction (but unreacted aldehyde groups of the carrier remain unchanged in this case) [28]. Thus, the first step before immobilization on GA was to assay the stability of OYE3 at alkaline pH. A pH “window” of 8–10 was considered and 3 h was selected as the endpoint (according to the general protocol for this type of immobilization) [19]. Enzyme activity before and after incubation at pH 8–10 (see Section 3.5, Materials and Methods) was assessed spectrophotometrically using the cyclohex-2-enone assay, Scheme 1). Data of the stability assay at pH 8–10 are reported in Figure 1. The residual activity of OYE3 after 3 h at pH 10 was about 70%. This result was considered acceptable for the immobilization on GA.
2.3. Immobilization of OYE3
Immobilization of OYE3 on GA (OYE3-GA) was performed according to a standard protocol by using a 2 mg g−1 protein loading [19], as described in Section 3.6 (Materials and Methods). In order to extend the enzyme stability during the immobilization, the reaction was performed at 4 °C. After 3 h, almost all the protein was immobilized (immobilization yield = 95%), as revealed by protein measurements of the supernatant from the immobilization reaction. The activity recovery, after reduction of the immobilized enzyme with NaBH4, was 52% (Table 1).
For the immobilization by metal-coordination, three EziGTM resins were tested (namely Opal or EziG 1, Coral or EziG 2, and Amber or EziG 3). According to the indications of the supplier, Opal is a hydrophilic carrier (glass), whereas Coral and Amber are hydrophobic and semi-hydrophilic, respectively. The lower hydrophilicity of Coral and Amber in comparison with Opal is due to the presence of different polymeric coatings on the particle silica surface [30]. Specifically, EziGTM Coral is made from hybrid CPG with polyvinyl benzyl chloride, whereas EziGTM Amber is produced from hybrid CPG with a blended co-polymer. Pore size distribution of the carriers ranges from ~500 Å as standard (Opal) to ~300 Å (Coral and Amber, due to the polymer coating). According to the indications of the supplier, a higher protein loading (4.2 mg g−1) was applied to EziGTM carriers. In all cases, the immobilization was complete. The activity recovery for EziGTM Opal was 54%, thus resulting comparable to that previously obtained with GA. In the case of the hybrid CPG Coral and Amber, the activity recovery was less than half with respect to EziGTM Opal. Under the same binding chemistry, it is plausible that this result may be ascribed to the effect of the polymer coating in tuning the hydrophilicity–hydrophobicity balance of the carrier. However, a systematic study on these carriers was not performed at this stage. Due to the positive result achieved with EziGTM Opal, this carrier was selected for the full characterization of this type of enzyme preparation (stability, recycling, re-use, enzyme leakage).
2.4. Stability of Immobilized OYE3
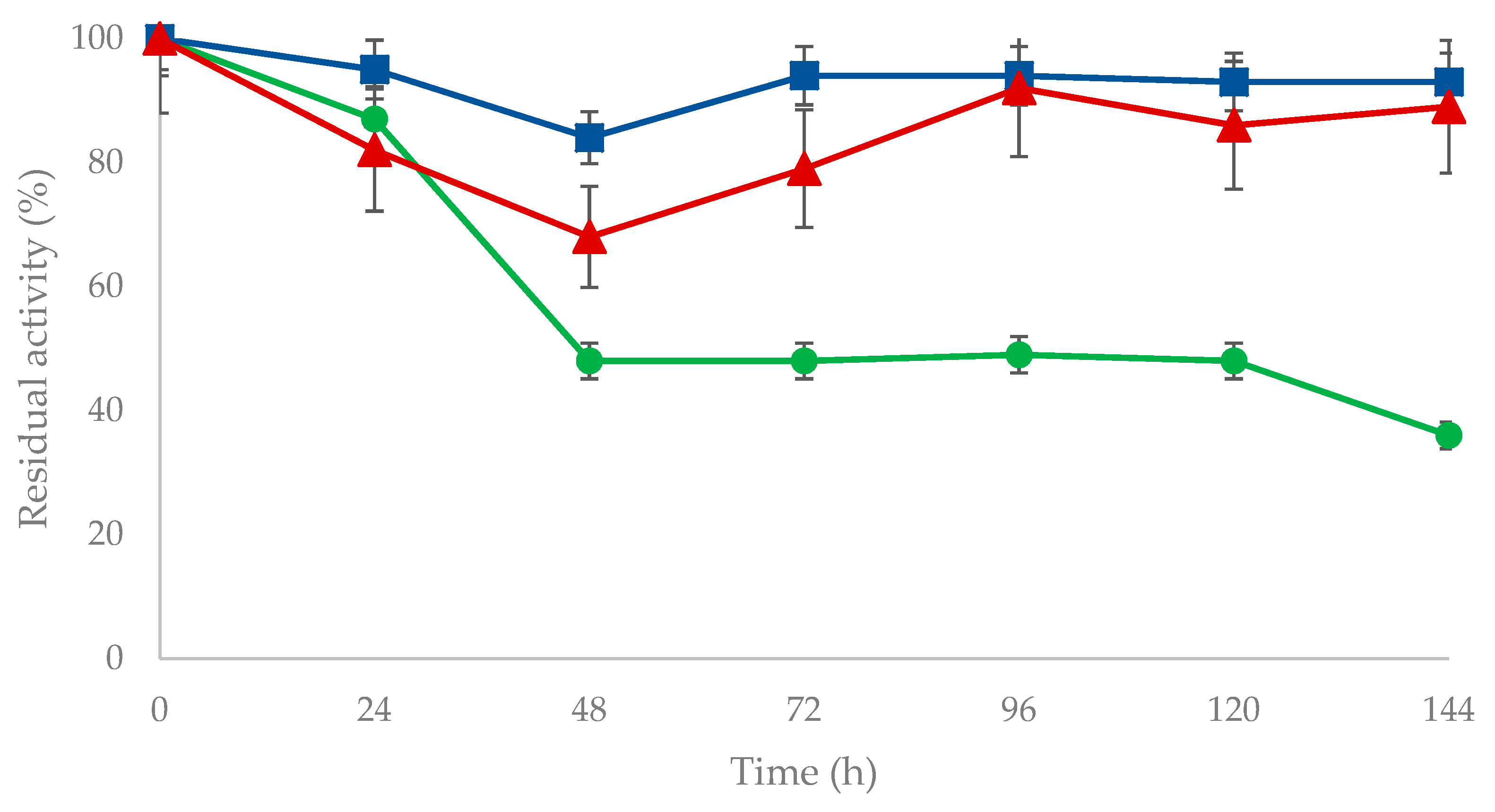
Increased stability is recognized as one of the most important advantages of immobilized enzymes [31,32]. To verify whether this “general rule” might be applied also to immobilized OYE3 preparations, the stability of both OYE3-GA and OYE3-EziG (Opal) was tested under the experimental conditions used for the reduction of (R)-carvone (2) (Scheme 2). The stability assay was carried on for six days (Figure 2). From the stability time course, it clearly emerged that immobilization exerted a stabilizing effect on the soluble enzyme. The OYE3-GA retained almost 100% of its activity, whereas the activity of the non-immobilized enzyme dropped to 50% after two days. On the other hand, OYE3-EziG was slightly less stable than OYE3-GA, but it still retained about 90% of its starting activity after 144 h.
2.5. Recycling of Immobilized OYE3
Biocatalyst recycling and re-use is a further strength point of immobilized enzymes [31,32,33]. Thus, both OYE3-GA and OYE3-EziG were used in the bioreduction of α-methyl-trans-cinnamaldehyde (3, Scheme 3), then recovered and added to a newly-prepared reaction mixture.
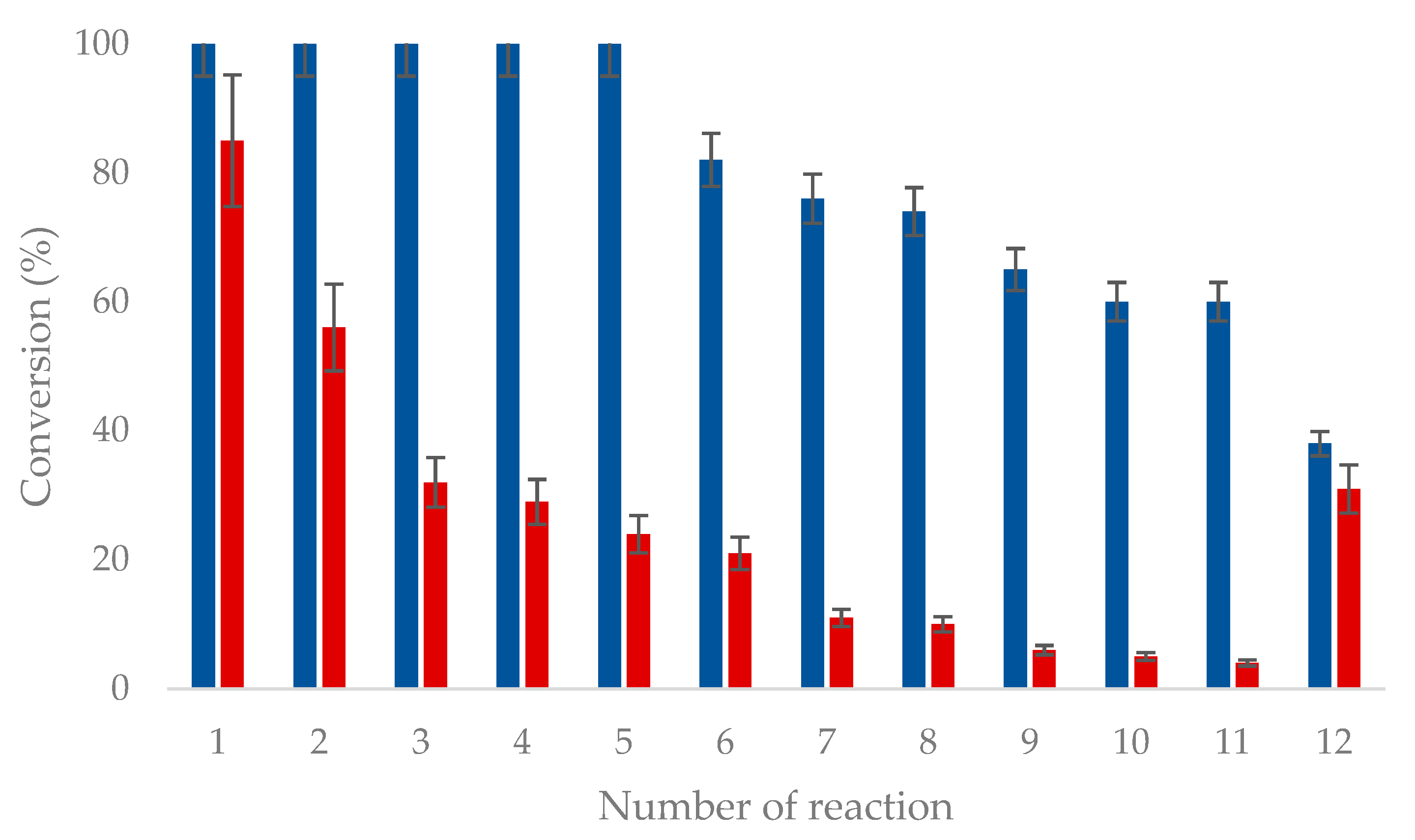
A catalytic amount of the cofactor (NADP+) was used since a GDH glucose regeneration system was coupled to the main biotransformation. In the case of OYE3-GA, GDH was added as a soluble enzyme to each reaction cycle. In the case of OYE3-EziG, a co-immobilized preparation OYE3/GDH was used, taking advantage of the His-tag displayed by both enzymes. The enzymes (OYE3 and GDH) were incubated with EziGTM. Activity recovery for OYE3 was 55%. OYE3-GA could be recycled and re-used for up to 12 reaction cycles. The highest conversion achieved after 12 runs was 40% (Figure 3). Moreover, it is worth noting that OYE3-GA retained completely its activity up to the fifth reaction cycle. In the case of OYE3/GDH-EziG, the initial conversion dropped to 56% after two reaction cycles. Interestingly, when the conversion % was almost negligible (reaction 11) and the biotransformation was supplemented with a fresh aliquot of soluble GDH, the reaction afforded a 30% conversion, thus suggesting that the gradual drop of the conversions registered for OYE3/GDH-EziG might depend either on the loss of activity of immobilized GDH or on its leakage from the carrier and/or deactivation, thereof.
2.6. Protein Leakage Assay of Immobilized OYE3
Protein leakage from the immobilization carrier is undesirable for several reasons: Loss of activity of the biocatalyst, lack of data reproducibility in the investigated biotransformation, costs and product contamination, to name a few [29]. Covalent immobilization of the enzyme to the carrier generally prevents the protein from leaching from the support surface. In this context, glyoxyl-agarose and other surface-functionalized carriers (i.e., epoxy carriers) are frequently preferred because of the “irreversible” nature of the bonds generated between the enzyme and the carrier. On the other hand, the high affinity and the specificity of the binding interaction based on the His-tag as in EziGTM-type carriers is plausible to generate a strong enzyme-support interaction which might prevent enzyme leakage. However, predictions about how an enzyme will respond to an immobilization process are often hard to make. Enzyme leakage is routinely verified by protein measurements [34] of the supernatant in which the immobilized preparation is incubated under specific conditions dictated by the type of information needed (e.g., reaction or storage conditions). Both OYE3-GA and OYE3/GDH-EziG were, thus, incubated in the reaction buffer (50 mM phosphate buffer pH 7) at 28 °C under stirring for 48 h. At scheduled times, samples of the incubation buffer were collected and submitted to SDS-PAGE analysis; silver staining, due to its high sensitivity in the low nanogram range, was used to detect protein traces after electrophoretic separation [35]. From the inspection of the SDS-PAGE gel (data not reported), no enzyme traces were detected for either OYE3-GA or OYE3/GDH-EziG. This suggests that the loss of activity of both immobilized enzymes registered during the recycling and re-use study, although to a different extent, cannot be ascribed to protein leakage phenomena.
3. Materials and Methods
3.1. General
Chemicals and solvents were purchased from Merck Sigma-Aldrich® Srl (Milano, Italy) and used without further purification. Agarose gel 6B-CL was purchased from Amersham Biosciences (Uppsala, Sweden) and activated to glyoxyl-agarose (GA), as previously reported [19]. EziGTM carriers were kindly provided by EnginZyme AB (Sweden) as free samples. GC-MS analyses were performed using a HP-5MS column (30 m × 0.25 mm × 0.25 μm, Agilent). The following temperature program was employed: 60 °C (1 min), 6 °C min−1/150 °C (1 min), 12 °C min−1/280 °C (5 min). Bradford assay [33] was performed by using a Shimadzu UV-1601 spectrophotometer and bovine serum albumin (BSA) was used as reference standard protein. All the enzymatic reactions were kept under stirring and controlled temperature by using a thermoshaker New Brunswick™ Excella® E24. Reaction extraction and phase separation were accomplished through a ZX3 Advanced Vortex Mixer from VELP Scientifica® and a Microcentrifuge ScanSpeed Mini (LaboGene™), respectively.
3.2. Enzyme Production
The enzymes OYE3 from Saccharomyces cerevisiae and GDH from Bacillus megaterium were prepared as His-tagged proteins and purified according to standard methods [36]. Concentration of the purified OYE3 was in the range 2–3 mg mL−1, whereas the concentration of GDH was 3–4 mg mL−1. Activity of OYE3 was measured both by the cyclohex-2-enone assay (see Section 3.3.1 and Scheme 1) and the (R)-carvone assay (see Section 3.3.2 and Scheme 2), resulting in 0.09 ± 0.008 U mg−1 and 0.13 ± 0.007 U mg−1, respectively. Activity of GDH was measured by a spectrophotometric assay (see Section 3.3.1), resulting in 6.7 ± 0.35 U mg−1.
3.3. Activity Assays
3.3.1. Spectrophotometric Enzymatic Assay
(a) For the OYE activity assay, in a 2 mL cuvette, a solution of cyclohex-2-enone (5 µL; stock solution = 10 mM in DMSO) and a solution of NADH (20 µL; stock solution = 10 mM in water) were added to 50 mM phosphate buffer pH 7 (916 µL). The reaction was carried out at 28 °C and started by adding to the blank solution 59 µL of OYE3 (177 µg, conc. = 3 mg mL−1).
(b) For the GDH activity assay, in a 2 mL cuvette, a solution of glucose (205 µL; stock solution = 1 M in water) and a solution of NAD+ (20 µL; stock solution = 10 mM in water) were added to 50 mM phosphate buffer pH 7 (960 µL). The reaction was carried out at 28 °C and started by adding to the blank solution 2 µL of GDH (6 µg, conc. = 3 mg mL−1).
The enzymatic activity (U mL−1) was calculated from the consumption formation of NADH, for (a) and (b), respectively, at 340 nm (εNAD(P)H = 6200 M−1 cm−1) according to the following equation:
ΔAbs/min: slope of Abs vs. time curve (mAU min−1), Vtot: total assay volume (mL), VE: volume of soluble enzyme used in the assay (mL), ε: NAD(P)H molar absorptivity (6200 M−1 cm−1).
One unit (U) is defined as the amount of enzyme that converts 1 µmol of substrate in the product in 1 min under specific temperature and pH conditions.
3.3.2. GC-MS Enzymatic Assay
In a 2 mL test-tube, a solution of (R)-carvone (10 µL; stock solution = 100 mM in DMSO) and 15 µL of soluble OYE3 (conc. = 3 mg mL−1) or immobilized OYE3 (ca. 20 mg for OYE3-GA or 45 mg for OYE3-EziG) were added to 50 mM phosphate buffer, pH 7 (final volume = 1 mL). The reaction was started by adding NADH (100 µL; stock solution = 10 mM in water) and the mixture was kept at 28 °C under stirring (thermoshaker) for 30 min, then vortexed for 30 s in the presence of CH2Cl2 (200 µL). The organic phase was separated by centrifugation (2 min, 13500 rpm), dried over Na2SO4 for 5 min, and analyzed by GC-MS (see Section 3.1 for analytical conditions). (R)-Carvone: GC-MS (EI) tR = 12.99 min: mz (%) = 150 (M+, 15), 135 (10), 108 (40), 93 (40), 82 (100). Product: GC-MS (EI) tR = 11.83 min: mz (%) = 152 (M+, 30), 137 (25), 123 (10), 109 (55), 95 (100). The enzymatic activity (U mL−1 for the non-immobilized enzyme, or U g−1 for the immobilized enzyme) was calculated from the percentage of substrate conversion after 30 min according to the following equation:
[S]: substrate concentration (mM), c: conversion (%), t: reaction time (30 min), E: amount of soluble enzyme (mL) or immobilized enzyme (g).
One unit (U) is defined as the amount of enzyme that converts 1 µmol of substrate in the product in 1 min under specific temperature and pH conditions.
3.4. OYE3-Mediated Reduction of α-Methyl-Trans-Cinnamaldehyde (3)
In a 2 mL test tube, a solution of α-methyl-trans-cinnamaldehyde (10 µL; stock solution = 500 mM in DMSO) was added to 50 mM phosphate buffer pH 7 (838 µL) followed by a solution of NADP+ (10 µL; stock solution = 10 mM in water), glucose (20 µL; stock solution = 1 M in water), and GDH (10 µL; 5 mg mL−1). The reaction (final volume = 1 mL) was started by adding 160 µg of OYE3 (non-immobilized enzyme = 53 µL; OYE3-GA = 80 mg; OYE3-EziG = 170 mg) and kept under stirring (thermoshaker) at 28 °C for 1 h. At the endpoint, in the case of the immobilized enzyme, the reaction was stopped by removing the biocatalyst by centrifugation (4 min, 13,200 rpm). The reaction mixture was vortexed for 30 s in the presence of CH2Cl2 (400 µL). The organic phase was separated by centrifugation (2 min, 13,200 rpm), dried over Na2SO4 for 5 min, and analyzed by GC-MS (see Section 3.1 for analytical conditions). α-Methyl-trans-cinnamaldehyde = GC-MS (EI) tR = 14.60 min = mz (%) = 146 (M+, 70), 145 (100), 117 (80), 115 (70), 91 (50). Product: GC-MS (EI) tR = 11.68 min = mz (%) = 148 (M+, 45), 133 (25), 105 (25), 91 (100). The conversion was evaluated from the ratio of the peak areas of product and substrate.
3.5. Stability of OYE3 in the pH Range 8-10
The stability of non-immobilized OYE3 was assayed in the pH range 8–10 by incubating 100 µL of the enzyme solution (stored in 20 mM phosphate buffer pH 8.5 containing 200 mM imidazole and 500 mM NaCl) in 100 µL of 50 mM phosphate buffer pH 7.5 (final pH = 8), 50 mM Tris-HCl buffer pH 9.5 (final pH = 9), or 50 mM NaOH-glycine buffer pH 11.5 (final pH = 10) at 20 °C. After 3 h, 100 µL were withdrawn from each sample and the enzymatic activity was measured spectrophotometrically in the reduction of cyclohex-2-enone (see Section 3.3.1). The residual activity was calculated according to the following equation:
3.6. Covalent Immobilization of OYE3 on Glyoxyl-Agarose (OYE3-GA)
Immobilization of OYE3 on glyoxyl-agarose was performed following the protocol previously reported [19]. Briefly, glyoxyl-agarose (1.8 g = 2.52 mL) was suspended in 50 mM Na2CO3 buffer (14.3 mL) at pH 10. After the addition of the desired amount of protein (1.2 mL, loading: 2 mg g−1), the suspension was kept under mechanical stirring for 3 h at 4 °C. Chemical reduction of Schiff bases and unreacted aldehyde groups was carried out over 30 min by adding NaBH4 to the mixture (18 mg; 1 mg mL−1 of suspension). The immobilized enzyme was then filtered and washed with 10 mM phosphate buffer pH 7 and deionized water.
The immobilization reaction was monitored by measuring the amount of protein in the supernatant before the addition of the carrier and after 1.5 h and 3 h [34]. The immobilization yield (%, endpoint = 3 h) was calculated according to the following equation [29]:
The activity of the immobilized OYE3 was measured by the GC-MS enzymatic assay reported in Section 3.3.2 (reduction of (R)-carvone, 2). The activity recovery (%) was calculated according to the following equation [29]:
3.7. Immobilization of OYE3 by Affinity-Based Adsorption (OYE3-EziG)
Immobilization of OYE3 by affinity-based adsorption on EziGTM (Opal, Coral, and Amber) was performed following the protocol of the supplier with minor modifications. EziGTM (100 mg) was added to a 2 mL test tube containing the enzyme (140 µL, loading = 4.2 mg·g−1 of dry carrier) diluted in 50 mM phosphate buffer pH 7 (1.5 mL) at r.t. After 30 min, the test tube was centrifugated (5 min, 13,200 rpm) and the supernatant was removed by pipetting. Following supplier’s instructions, the wet enzyme preparation was stored as such (when the immobilized enzyme was filtered under vacuum, it showed an almost complete loss of activity. The weight of the wet carrier was estimated to be 4.5-fold higher than the dry carrier. The activity recovery (%) was referred to the wet carrier weight).
The immobilization yield (%, endpoint = 30 min) was calculated as reported above (Section 3.6). The activity of the immobilized OYE3 was measured by the (R)-carvone assay (see Section 3.3.2.) and the activity recovery (%) was calculated as described above (Section 3.6).
Co-immobilization of OYE3 and GDH on EziG™ Opal was carried out by following the procedure applied to OYE3 immobilization. Briefly, EziG™ Opal (100 mg) was added to 50 mM phosphate buffer pH 7 (1.5 mL) containing OYE3 (140 µL, U = 0.06) and GDH (45 µL, U = 0.87). Monitoring and work-up of the immobilization mixture was performed as above reported.
3.8. Stability of Non-Immobilized OYE3, OYE3-GA, and OYE3-EziG
For each endpoint, non-immobilized or immobilized OYE3 (42 µg of protein) was incubated in a solution containing 50 mM phosphate buffer pH 7 (890 µL) and (R)-carvone (10 µL; stock solution = 100 mM in DMSO) at 28 °C. Each reaction was started by adding NADH (100 µL; stock solution: 10 mM in water). The final volume was 1 mL. The enzyme activity was assessed by the standard activity assay described in the Section 3.3.2 (reduction of (R)-carvone). The enzyme activity was compared with the starting activity which was considered as 100%.
3.9. Protein Leakage Assay
For each endpoint (1-3-6-24-48 h), 42 µg of protein (corresponding to 21 mg of OYE3-GA and 45 mg of OYE3/GDH-EziG) were incubated in 50 mM phosphate buffer pH 7 (200 µL). The suspensions were kept at 28 °C under stirring (thermoshaker). At each endpoint, a sample (15 µL) was withdrawn from the supernatant and analyzed by SDS-PAGE. GDH and OYE3 (450 ng, corresponding to 0.15 µL of each stock solution) were loaded in the gel as reference standards. Precision Plus Protein™ Unstained Protein Standards (10–250 kD) was used as molecular weight proteins size marker. PAGE analysis was performed in a Mini-Protean 3 system (Bio-Rad). SDS-PAGE was carried out on 12% polyacrylamide (running buffer = 50 mM MES, 50 mM Tris base, 0.1 % w/v SDS and 1 mM EDTA). Proteins were stained by means of a silver staining kit (SilverQuest™ kit, Thermofisher) according to supplier’s protocol.
3.10. Recycling of Immobilized Enzymes
Recycling of immobilized OYE3-GA and OYE3/GDH-EziG was performed by evaluating the conversion (%) of α-methyl-trans-cinnamaldehyde (3) into 3a (see Section 3.4). At the end of the reaction, the immobilized biocatalyst was washed twice with 1 mL of 50 mM phosphate buffer pH 7, separated through centrifugation, and then re-suspended under the conditions above described for the next reaction cycle.
4. Conclusions
Ene-reductases are a powerful tool for the stereoselective reduction of activated C=C double bonds under mild and environmentally friendly conditions. Due to the great interest in these enzymes for preparative applications, the access to stable and reusable heterogeneous biocatalysts is being sought. Immobilization of OYE3 both on the well-known glyoxyl-agarose (GA) by covalent binding and on controlled pore glass EziGTM by affinity-based adsorption was shown to produce active, stable and reusable biocatalysts. Although activity recovery for both enzyme preparations was similar (≈50%), OYE3-GA appeared to be superior about stability and recycling properties. In this latter regard, OYE3-GA could be re-used up to five reaction cycles without any loss of activity. Still, upon a prolonged re-use (up to 12 runs), this immobilized biocatalyst allowed to obtain a 40% conversion in the reduction of α-methyl-trans-cinnamaldehyde (3). On the other hand, in the case of the co-immobilized OYE3/GDH-EziG, a drop of conversion was observed after two reaction cycles, presumably due to deactivation of GDH. Interestingly, no evidence of enzyme leaching from both the supports was registered.
Author Contributions
Conceptualization, E.B. and D.U.; methodology, F.T., D.C. and R.S.; investigation, F.T., D.C., R.S. and T.B.; data curation, F.T.; writing (original draft preparation, review and editing) F.T., T.B., E.B., D.M. and D.U.; supervision, E.B., D.M. and D.U.; all authors critically reviewed the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
EnginZyme AB (Sweden) is gratefully acknowledged for the generous gift of EziGTM enzyme carriers. We thank Fabio Parmeggiani (Politecnico di Milano, Italy) for critical reading of the manuscript and assistance in the design of the graphical abstract.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Winkler, C.K.; Tasnádi, G.; Clay, D.; Hall, M.; Faber, K. Asymmetric bioreduction of activated alkenes to industrially relevant optically active compounds. J. Biotechnol. 2012, 162, 381–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toogood, H.S.; Scrutton, N.S. New developments in ‘ene’-reductase catalysed biological hydrogenations. Curr. Opin. Chem. Biol. 2014, 19, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Vaz, A.D.N.; Chakraborty, S.; Massey, V. Old yellow enzyme: Aromatization of cyclic enones and the mechanism of a novel dismutation reaction. Biochemistry 1995, 34, 4246–4256. [Google Scholar] [CrossRef] [PubMed]
- Gatti, F.G.; Parmeggiani, F.; Sacchetti, A. Synthetic strategies based on C=C bioreductions for the preparation of biologically active molecules. In Synthetic Methods for Biologically Active Molecules; Brenna, E., Ed.; WILEY-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2013; pp. 49–84. [Google Scholar]
- Toogood, H.S.; Scrutton, N.S. Discovery, characterization, engineering, and applications of ene-reductases for industrial biocatalysis. ACS Catal. 2018, 8, 3532–3549. [Google Scholar] [CrossRef] [PubMed]
- Winkler, C.K.; Faber, K.; Hall, M. Biocatalytic reduction of activated C=C-bonds and beyond: Emerging trends. Curr. Opin. Chem. Biol. 2018, 43, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Classen, T.; Korpak, M.; Schölzel, M.; Pietruszka, J. Stereoselective enzyme cascades: An efficient synthesis of chiral γ-butyrolactones. ACS Catal. 2014, 4, 1321–1331. [Google Scholar] [CrossRef]
- Brenna, E.; Crotti, M.; Gatti, F.G.; Monti, D.; Parmeggiani, F.; Pugliese, A.; Tentori, F. Biocatalytic synthesis of chiral cyclic γ-oxoesters by sequential C–H hydroxylation, alcohol oxidation and alkene reduction. Green Chem. 2017, 19, 5122–5130. [Google Scholar] [CrossRef]
- Winkler, C.K.; Clay, D.; Davies, S.; O’Neill, P.; McDaid, P.; Debarge, S.; Steflik, J.; Karmilowicz, M.; Wong, J.W.; Faber, K. Chemoenzymatic asymmetric synthesis of pregabalin precursors via asymmetric bioreduction of β-cyanoacrylate esters using ene-reductases. J. Org. Chem. 2013, 78, 1525–1533. [Google Scholar] [CrossRef]
- Brenna, E.; Gatti, F.G.; Manfredi, A.; Monti, D.; Parmeggiani, F. Biocatalyzed enantioselective reduction of activated C=C bonds: Synthesis of enantiomerically enriched α-halo-β-arylpropionic acids. Eur. J. Org. Chem. 2011, 4015–4022. [Google Scholar] [CrossRef]
- Brenna, E.; Crotti, M.; Gatti, F.G.; Monti, D.; Parmeggiani, F.; Powell, R.W.; Santangelo, S.; Stewart, J.D. Opposite enantioselectivity in the bioreduction of (Z)-β-aryl-β-cyanoacrylates mediated by the tryptophan 116 mutants of old yellow enzyme 1: Synthetic approach to (R)- and (S)-β-aryl-γ-lactams. Adv. Synth. Catal. 2015, 357, 1849–1860. [Google Scholar] [CrossRef]
- Waller, J.; Toogood, H.S.; Karuppiah, V.; Rattray, N.J.W.; Mansell, D.J.; Leys, D.; Gardiner, J.M.; Fryszkowska, A.; Ahmed, S.T.; Bandichhor, R.; et al. Structural insights into the ene-reductase synthesis of profens. Org. Biomol. Chem. 2017, 15, 4440–4448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumru, C.; Classen, T.; Pietruszka, J. Enantioselective, catalytic one-pot synthesis of γ-butyrolactone-based fragrances. ChemCatChem 2018, 10, 4931–4940. [Google Scholar] [CrossRef]
- Guo, J.; Zhang, R.; Ouyang, J.; Zhang, F.; Qin, F.; Liu, G.; Zhang, W.; Li, H.; Ji, X.; Jia, X.; et al. Stereodivergent synthesis of carveol and dihydrocarveol through ketoreductasesene-reductases catalyzed asymmetric reduction. ChemCatChem 2018, 10, 5496–5504. [Google Scholar] [CrossRef]
- Stueckler, C.; Mueller, N.J.; Winkler, C.K.; Glueck, S.M.; Gruber, K.; Steinkellner, G.; Faber, K. Bioreduction of α-methylcinnamaldehyde derivatives: Chemo-enzymatic asymmetric synthesis of Lilial™ and Helional™. Dalton Trans. 2010, 39, 8472–8476. [Google Scholar] [CrossRef]
- Bryan, M.C.; Dunn, P.J.; Entwistle, D.; Gallou, F.; Koenig, S.G.; Hayler, J.D.; Hickey, M.R.; Hughes, S.; Kopach, M.E.; Moine, G.; et al. Key Green Chemistry research areas from a pharmaceutical manufacturers’ perspective revisited. Green Chem. 2018, 20, 5082–5103. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Xiao, W.; Xie, P.; Zheng, L. Co-immobilization of enoate reductase with a cofactor-recycling partner enzyme. Enzym. Microb. Technol. 2018, 109, 66–73. [Google Scholar] [CrossRef]
- Yoon, J.; Lee, S.H.; Tieves, F.; Rauch, M.; Hollmann, F.; Park, C.B. Light-harvesting dye−alginate hydrogel for solar-driven, sustainable biocatalysis of asymmetric hydrogenation. ACS Sustain. Chem. Eng. 2019, 7, 5632–5637. [Google Scholar] [CrossRef] [Green Version]
- Guisán, J.M. Aldehyde-agarose gels as activated supports for immobilization-stabilization of enzymes. Enzyme Microb. Technol. 1988, 10, 375–382. [Google Scholar] [CrossRef]
- Zucca, P.; Fernandez-Lafuente, R.; Sanjust, E. Agarose and its derivatives as supports for enzyme immobilization. Molecules 2016, 21, 1577. [Google Scholar] [CrossRef]
- Fernández-Lorente, G.; Lopez-Gallego, F.; Bolivar, J.M.; Rocha-Martin, J.; Moreno-Perez, S.; Guisán, J.M. Immobilization of proteins on highly activated glyoxyl supports: Dramatic increase of the enzyme stability via multipoint immobilization on pre-existing carriers. Curr. Org. Chem. 2015, 19, 1–13. [Google Scholar] [CrossRef]
- Dall’Oglio, F.; Contente, M.L.; Conti, P.; Molinari, F.; Monfredi, D.; Pinto, A.; Romano, D.; Ubiali, D.; Tamborini, L.; Serra, I. Flow-based stereoselective reduction of ketones using an immobilized ketoreductaseglucose dehydrogenase mixed bed system. Catal. Commun. 2017, 93, 29–32. [Google Scholar] [CrossRef]
- Cassimjee, K.E.; Kadow, M.; Wikmark, Y.; Svedendahl Humble, M.; Rothstein, M.L.; Rothstein, D.M.; Bäckvall, J.-E. A general protein purification and immobilization method on controlled porosity glass: Biocatalytic applications. Chem. Commun. 2014, 50, 9134–9137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, M.P.; Derrington, S.R.; Heath, R.S.; Porter, J.L.; Mangas-Sanchez, J.; Devine, P.N.; Truppo, M.D.; Turner, N.J. A generic platform for the immobilisation of engineered biocatalysts. Tetrahedron 2019, 75, 327–334. [Google Scholar] [CrossRef]
- Chaparro-Riggers, J.F.; Rogers, T.A.; Vazquez-Figueroa, E.; Polizzi, K.M.; Bommarius, A.S. Comparison of three enoate reductases and their potential use for biotransformations. Adv. Synth. Catal. 2007, 309, 1521–1531. [Google Scholar] [CrossRef]
- Kohli, R.M.; Massey, V. The oxidative half-reaction of Old Yellow Enzyme. J. Biol. Chem. 1998, 273, 32763–32770. [Google Scholar] [CrossRef] [Green Version]
- Hulley, M.E.; Toogood, H.S.; Fryszkowska, A.; Mansell, D.; Stephens, G.M.; Gardiner, J.M.; Scrutton, N.S. Focused directed evolution of pentaerythritol tetranitrate reductase by using automated anaerobic kinetic screening of site-saturated libraries. ChemBioChem 2010, 11, 2433–2447. [Google Scholar] [CrossRef]
- Orrego, A.H.; Romero-Fernández, M.; Millán-Linares, M.C.; Yust, M.M.; Guisán, J.M.; Rocha-Martin, J. Stabilization of enzymes by multipoint covalent attachment on aldehyde-supports: 2-picoline borane as an alternative reducing agent. Catalysts 2018, 8, 333. [Google Scholar] [CrossRef] [Green Version]
- Sheldon, R.A.; van Pelt, S. Enzyme immobilisation in biocatalysis: Why, what and how. Chem. Soc. Rev. 2013, 42, 6223–6235. [Google Scholar] [CrossRef] [Green Version]
- EnginZyme: Enzyme Carrier, Biocatalysis Research. Available online: http://enginzyme.com/ (accessed on 30 January 2020).
- Mateo, C.; Palomo, J.M.; Fernandez-Lorente, G.; Guisán, J.M.; Fernandez-Lafuente, R. Improvement of enzyme activity, stability and selectivity via immobilization techniques. Enzym. Microb. Technol. 2007, 40, 1451–1463. [Google Scholar] [CrossRef]
- Foley, A.M.; Maguire, A.R. The impact of recent developments in technologies which enable the increased use of biocatalysts. Eur. J. Org. Chem. 2019, 3713–3734. [Google Scholar] [CrossRef]
- Sheldon, R.A.; Brady, D. The limits to biocatalysis: Pushing the envelope. Chem. Commun. 2018, 54, 6088–6104. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 27, 248–254. [Google Scholar] [CrossRef]
- Chevallet, M.; Luche, S.; Rabilloud, T. Silver staining of proteins in polyacrylamide gels. Nat. Protoc. 2006, 1, 1852–1858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brenna, E.; Gatti, F.G.; Monti, D.; Parmeggiani, F.; Sacchetti, A. Cascade coupling of ene reductases with alcohol dehydrogenases: Enantioselective reduction of prochiral unsaturated aldehydes. ChemCatChem 2012, 4, 653–659. [Google Scholar] [CrossRef]
Scheme 1.
Reduction of cyclohex-2-enone (1) to cyclohexanone (1a) catalyzed by OYE3 (spectrophotometric activity assay). Experimental conditions: 50 mM phosphate buffer pH 7 (0.5% DMSO), [substrate] = 0.05 mM, [NADH] = 0.2 mM, 28 °C, reaction time = 80 s, volume = 1 mL.
Scheme 1.
Reduction of cyclohex-2-enone (1) to cyclohexanone (1a) catalyzed by OYE3 (spectrophotometric activity assay). Experimental conditions: 50 mM phosphate buffer pH 7 (0.5% DMSO), [substrate] = 0.05 mM, [NADH] = 0.2 mM, 28 °C, reaction time = 80 s, volume = 1 mL.

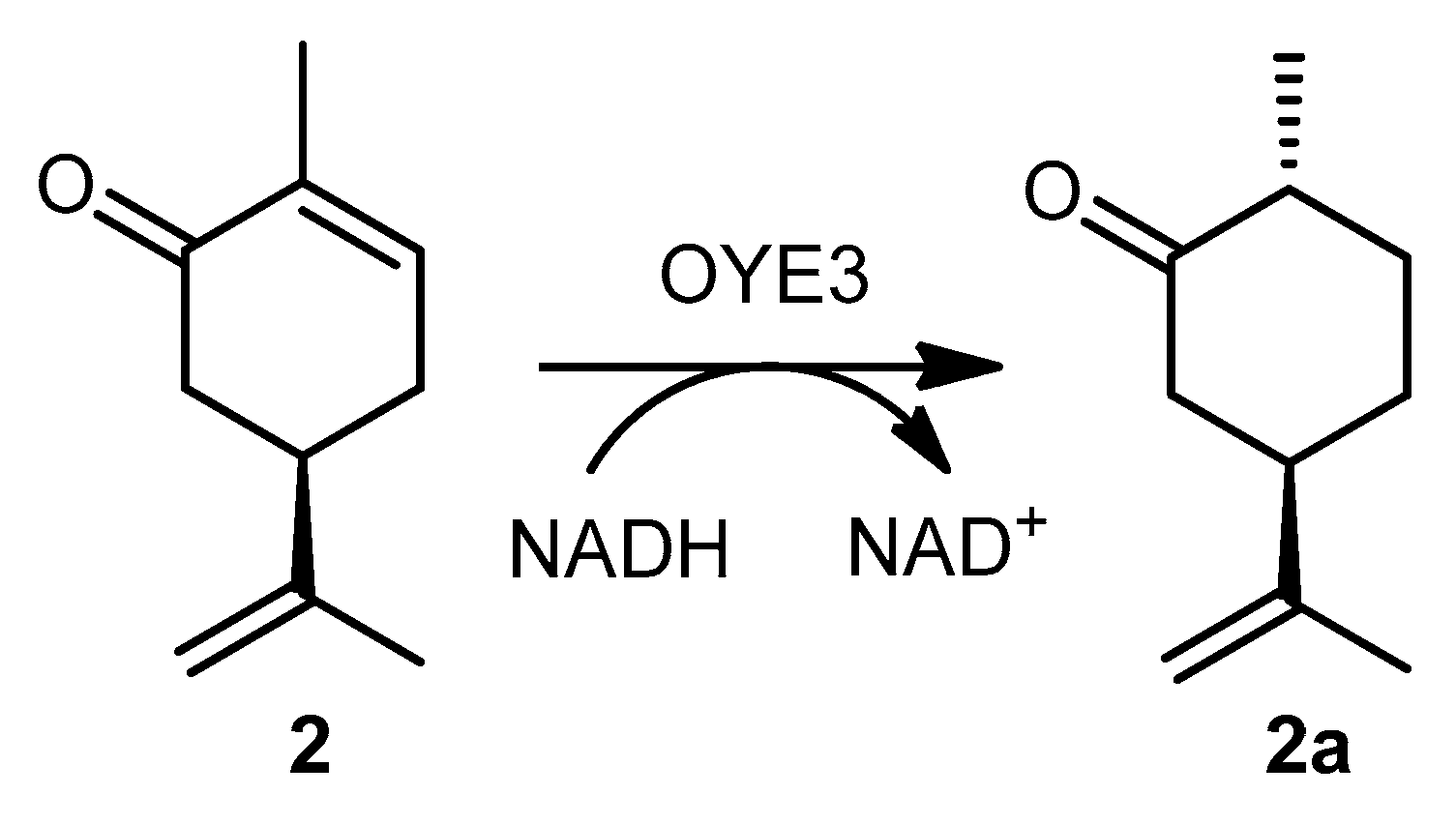
Scheme 2.
Reduction of (R)-carvone (2) to (2S,3R)-dihydrocarvone (2a) catalyzed by OYE3 (GC-MS activity assay). Experimental conditions: 50 mM phosphate buffer pH 7 (1% DMSO), [substrate] = 1 mM, [NADH] = 1 mM, 28 °C, reaction time = 30 min, volume = 1 mL.
Scheme 2.
Reduction of (R)-carvone (2) to (2S,3R)-dihydrocarvone (2a) catalyzed by OYE3 (GC-MS activity assay). Experimental conditions: 50 mM phosphate buffer pH 7 (1% DMSO), [substrate] = 1 mM, [NADH] = 1 mM, 28 °C, reaction time = 30 min, volume = 1 mL.

Figure 1.
Residual activity (%) of soluble OYE3 upon incubation at pH 8–10 (endpoint—3 h). Experimental conditions: 50 mM phosphate buffer, pH 8; 50 mM Tris-HCl buffer, pH 9; 50 mM NaOH-glycine buffer, pH 10; 20 °C. Enzyme activity was determined as reported in Scheme 1. Residual activity (%) was calculated as reported in Section 3.5 [29].
Figure 1.
Residual activity (%) of soluble OYE3 upon incubation at pH 8–10 (endpoint—3 h). Experimental conditions: 50 mM phosphate buffer, pH 8; 50 mM Tris-HCl buffer, pH 9; 50 mM NaOH-glycine buffer, pH 10; 20 °C. Enzyme activity was determined as reported in Scheme 1. Residual activity (%) was calculated as reported in Section 3.5 [29].

Figure 2.
Stability of soluble OYE3 (green circles), OYE3-GA (blue squares) and OYE3-EziG (red triangles). Experimental conditions: 50 mM phosphate buffer pH 7 (1% DMSO), (R)-carvone and NADH = 1 mM, 28 °C, volume = 1 mL.
Figure 2.
Stability of soluble OYE3 (green circles), OYE3-GA (blue squares) and OYE3-EziG (red triangles). Experimental conditions: 50 mM phosphate buffer pH 7 (1% DMSO), (R)-carvone and NADH = 1 mM, 28 °C, volume = 1 mL.

Scheme 3.
Reduction of α-methyl-trans-cinnamaldehyde (3) to (S)-α-methyl-β-phenylpropanal (3a) catalyzed by OYE3. The regeneration of NADP+ was ensured by the GDH glucose system. Experimental conditions: 50 mM phosphate buffer pH 7 (1% DMSO), [substrate] = 5 mM, [NADP+] = 0.1 mM, [glucose] = 20 mM, GDH = 10 µL; 5 mg mL−1, 28 °C, reaction time = 1 h, volume = 1 mL.
Scheme 3.
Reduction of α-methyl-trans-cinnamaldehyde (3) to (S)-α-methyl-β-phenylpropanal (3a) catalyzed by OYE3. The regeneration of NADP+ was ensured by the GDH glucose system. Experimental conditions: 50 mM phosphate buffer pH 7 (1% DMSO), [substrate] = 5 mM, [NADP+] = 0.1 mM, [glucose] = 20 mM, GDH = 10 µL; 5 mg mL−1, 28 °C, reaction time = 1 h, volume = 1 mL.

Figure 3.
Recycling of immobilized OYE3-GA (blue) and OYE3/GDH-EziG (red). Experimental conditions as reported in Scheme 3.
Figure 3.
Recycling of immobilized OYE3-GA (blue) and OYE3/GDH-EziG (red). Experimental conditions as reported in Scheme 3.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Results of OYE3 immobilization.
| Carrier | Enzyme Loading (mg g−1) | Immobilization Yield 1 (%) | Activity 2 (U g−1) | Activity Recovery 3 (%) |
|---|---|---|---|---|
| Glyoxyl-agarose | 2.0 | 95 | 0.023 | 52 |
| EziGTM Opal | 4.2 | 100 | 0.037 | 54 |
| EziGTM Coral | 4.2 | 100 | 0.015 | 23 |
| EziGTM Amber | 4.2 | 100 | 0.014 | 19 |
1 Calculated as reported in Section 3.6 [29]. 2 Determined by the (R)-carvone (2) assay (see Scheme 2). 3 Calculated as reported in Section 3.6 [29]. Activity measurements were performed at least in duplicate. Deviation from mean values was below 5%.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tentori, F.; Bavaro, T.; Brenna, E.; Colombo, D.; Monti, D.; Semproli, R.; Ubiali, D. Immobilization of Old Yellow Enzymes via Covalent or Coordination Bonds. Catalysts 2020, 10, 260. https://doi.org/10.3390/catal10020260
AMA Style
Tentori F, Bavaro T, Brenna E, Colombo D, Monti D, Semproli R, Ubiali D. Immobilization of Old Yellow Enzymes via Covalent or Coordination Bonds. Catalysts. 2020; 10(2):260. https://doi.org/10.3390/catal10020260
Chicago/Turabian StyleTentori, Francesca, Teodora Bavaro, Elisabetta Brenna, Danilo Colombo, Daniela Monti, Riccardo Semproli, and Daniela Ubiali. 2020. "Immobilization of Old Yellow Enzymes via Covalent or Coordination Bonds" Catalysts 10, no. 2: 260. https://doi.org/10.3390/catal10020260
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.