Influence of Cooking (Microwaving and Broiling) on Cylindrospermopsin Concentration in Muscle of Nile Tilapia (Oreochromis niloticus) and Characterization of Decomposition Products

 , and
, and
Abstract
:1. Introduction
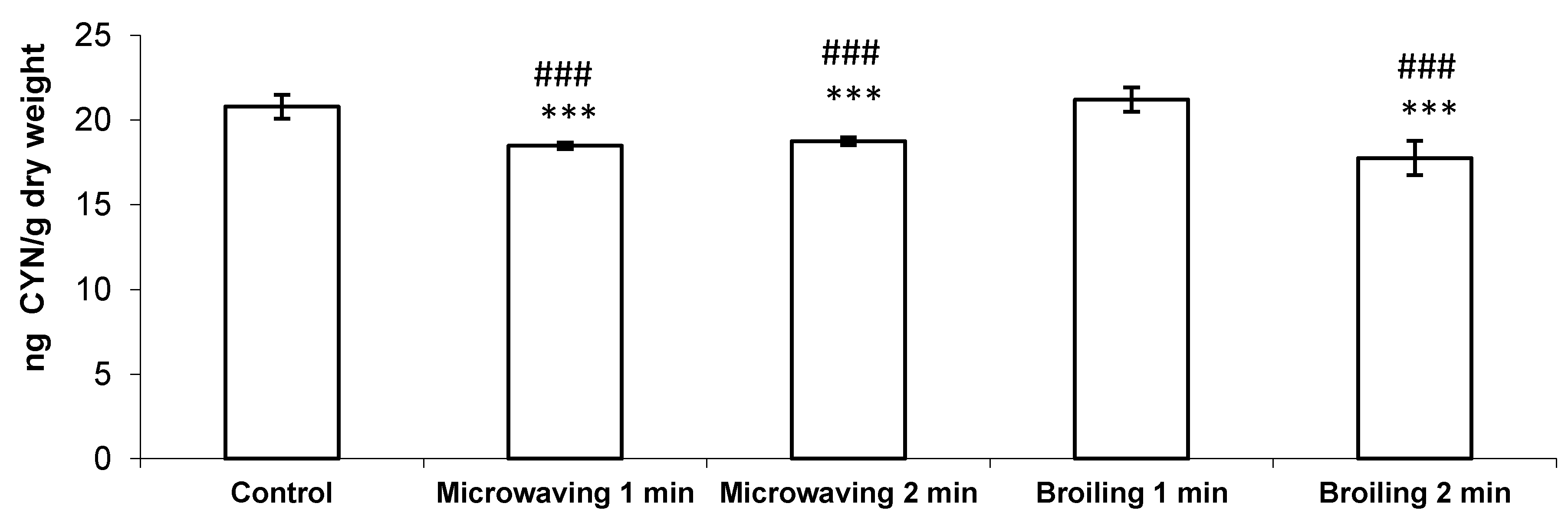
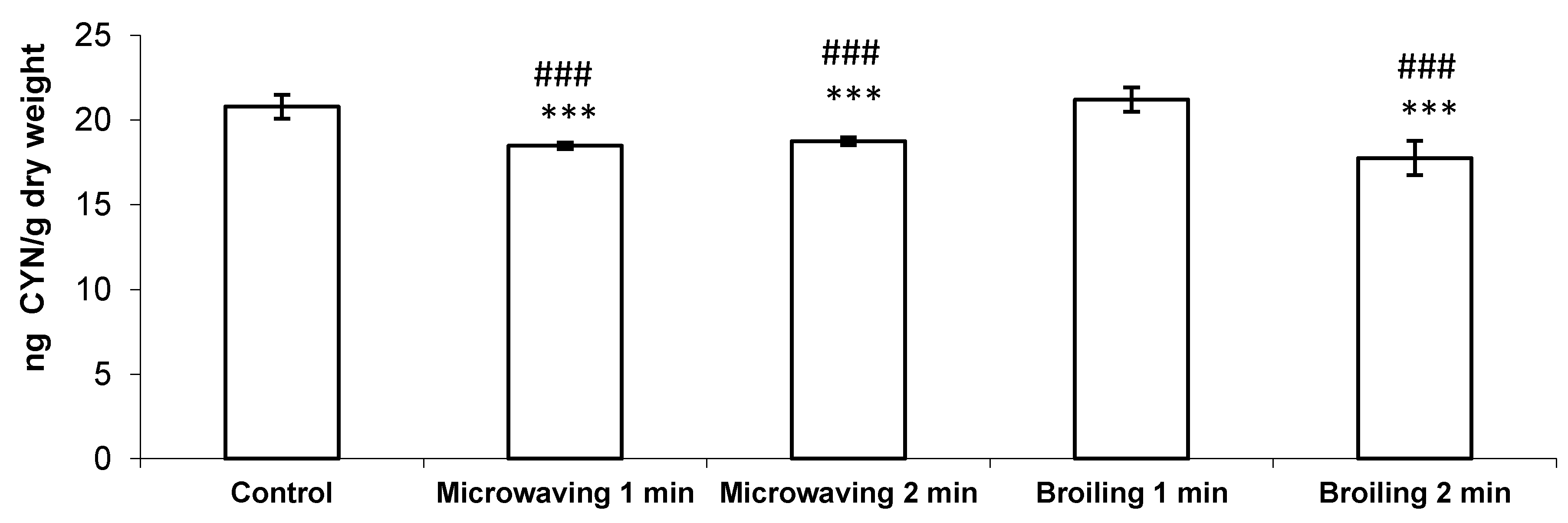
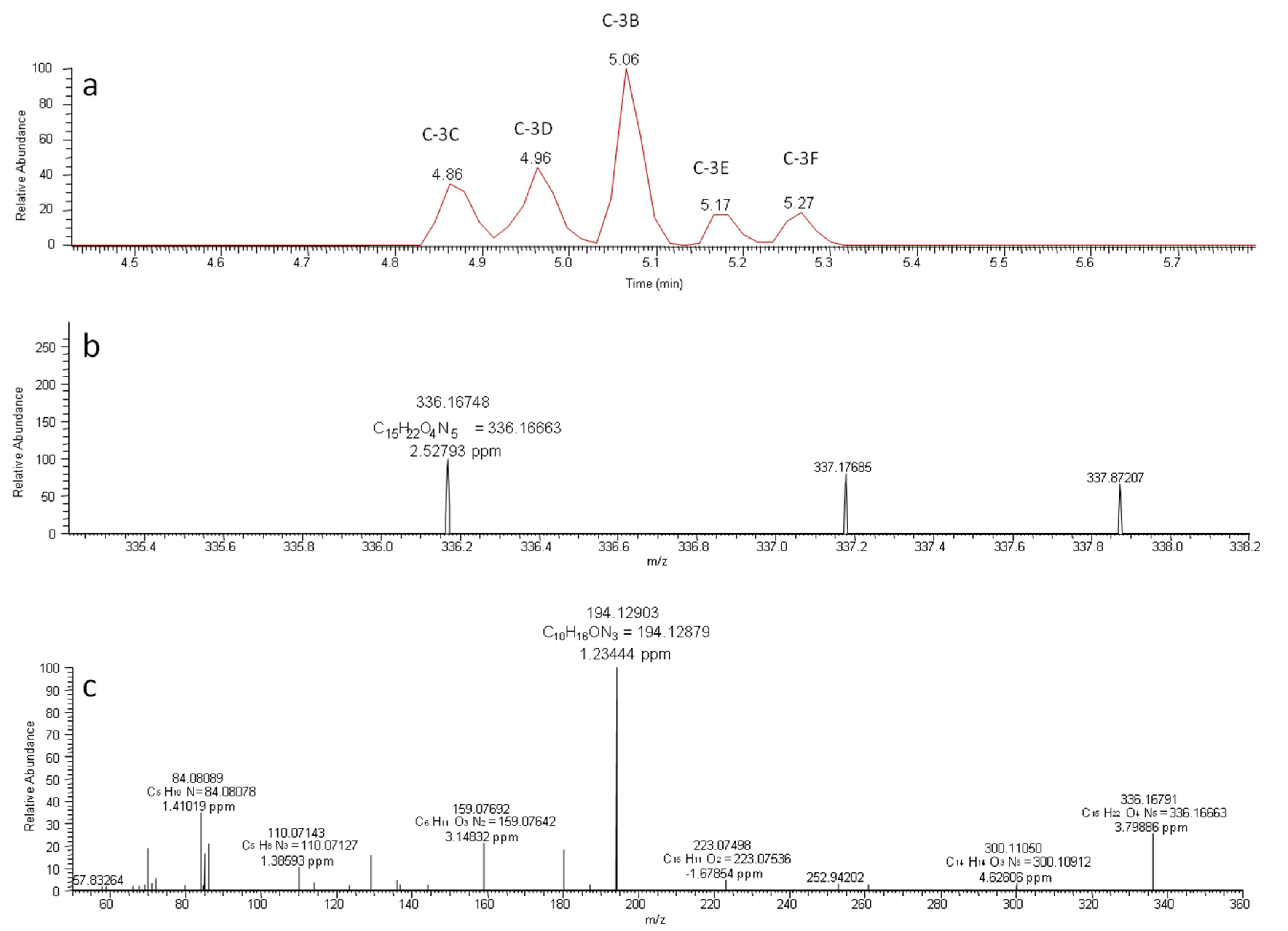
2. Results and Discussion
3. Conclusions
4. Materials and Methods
4.1. Reagents
4.2. Sample Preparation
4.3. Simulation of Food Processing Practices
4.4. Extraction and Purification of CYN from Fish Fillets
4.5. Chromatographic Conditions
4.6. Analytical Detection of CYN Decomposition Products after Cooking by UHPLC-MS/MS Orbitrap
4.7. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- De la Cruz, A.A.; Hiskia, A.; Kaloudis, T.; Chernoff, N.; Hill, D.; Antoniou, M.G.; He, X.; Loftin, K.; O’Shea, K.; Zhao, C.; et al. A review on cylindrospermopsin: The global occurrence, detection, toxicity and degradation of a potent cyanotoxin. Environ. Sci. Process. Impacts 2013, 15, 1979–2003. [Google Scholar] [CrossRef] [PubMed]
- Wimmer, K.M.; Strangman, W.K.; Wright, J.L.C. 7-Deoxydesulfocylindrospermopsin and 7-deoxy-desulfo-12-acetylcylindrospermopsin: Two new cylindrospermopsin analogs isolated from a Thai strain of Cylindrospermopsis raciborskii. Harmful Algae 2014, 37, 203–206. [Google Scholar] [CrossRef]
- Loftin, K.A.; Graham, J.L.; Hilborn, E.D.; Lehmann, S.C.; Meyer, M.T.; Dietze, J.E.; Griffith, C.B. Cyanotoxins in inland lakes of the United States: Occurrence and potential recreational health risks in the EPA National Lakes Assessment 2007. Harmful Algae 2016, 56, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Sivonen, K.; Jones, G. Cyanobacterial Toxins. In Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; Chorus, I., Bartram, J., Eds.; E & FN Spon: London, UK, 1999; pp. 41–111. [Google Scholar]
- Shaw, G.R.; Seawright, A.A.; Moore, M.R.; Lam, P.K.S. Cylindrospermopsin, a cyanobacterial alkaloid: Evaluation of its toxicologic activity. Ther. Drug Monit. 2000, 22, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Chiswell, R.K.; Shaw, G.R.; Eaglesham, G.; Smith, M.J.; Norris, R.L.; Seawright, A.A.; Moore, M.R. Stability of cylindrospermopsin, the toxin from the cyanobacterium, Cylindrospermopsis raciborskii: Effect of pH, temperature, and sunlight on decomposition. Environ. Toxicol. 1999, 14, 155–161. [Google Scholar] [CrossRef]
- Falconer, I.R.; Humpage, A.R. Cyanobacterial (blue-green algal) toxins in water supplies: Cylindrospermopsins. Environ. Toxicol. 2006, 21, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Guzmán-Guillén, R.; Lomares, I.; Moreno, I.M.; Prieto Ortega, A.I.; Moyano, R.; Blanco, A.; Cameán, A.M. Cylindrospermopsin induces neurotoxicity in tilapia fish (Oreochromis niloticus) exposed to Aphanizomenon ovalisporum. Aquat. Toxicol. 2015, 161, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Berry, J.P.; Jaja-Chimedza, A.; Dávalos-Lind, L.; Lind, O. Apparent bioaccumulation of cylindrospermopsin and paralytic shellfish toxins by finfish in Lake Catemaco (Veracruz, Mexico). Food Addit. Contam. Part A 2012, 29, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Ibelings, B.W.; Chorus, I. Accumulation of cyanobacterial toxins in freshwater “seafood” and its consequences for public health: A review. Environ. Pollut. 2007, 150, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Praena, D.; Jos, A.; Pichardo, S.; Moreno, I.M.; Cameán, A.M. Presence and bioaccumulation of microcystins and cylindrospermopsin in food and the effectiveness of some cooking techniques at decreasing their concentrations: A review. Food Chem. Toxicol. 2013, 53, 139–152. [Google Scholar] [CrossRef] [PubMed]
- Buratti, F.M.; Manganelli, M.; Vichi, S.; Stefanelli, M.; Scardala, S.; Testai, E.; Funari, E. Cyanotoxins: Producing organisms, occurrence, toxicity, mechanism of action and human health toxicological risk evaluation. Arch. Toxicol. 2017, 91, 1049–1130. [Google Scholar] [CrossRef] [PubMed]
- Freitas, M.; Azevedo, J.; Carvalho, A.P.; Mendes, V.M.; Manadas, B.; Campos, A.; Vasconcelos, V. Bioaccessibility and changes on cylindrospermopsin concentration in edible mussels with storage and processing time. Food Control 2016, 59, 567–574. [Google Scholar] [CrossRef]
- Guzmán-Guillén, R.; Maisanaba, S.; Prieto Ortega, A.I.; Valderrama-Fernández, R.; Jos, A.; Cameán, A.M. Changes on cylindrospermopsin concentration and characterization of decomposition products in fish muscle (Oreochromis niloticus) by boiling and steaming. Food Control 2017, 77, 210–220. [Google Scholar] [CrossRef]
- Van Buynder, P.G.; Oughtred, T.; Kirkby, B.; Phillips, S.; Eaglesham, G.; Thomas, K.; Burch, M. Nodularin uptake by seafood during a cyanobacterial bloom. Environ. Toxicol. 2001, 16, 468–471. [Google Scholar] [CrossRef] [PubMed]
- Morais, J.; Augusto, M.; Carvalho, A.P.; Vale, M.; Vasconcelos, V.M. Cyanobacteria hepatotoxins, microcystins: Bioavailability in contaminated mussels exposed to different environmental conditions. Eur. Food Res. Technol. 2008, 227, 949–952. [Google Scholar] [CrossRef]
- Bruno, M.; Melchiorre, S.; Messineo, V.; Volpi, F.; Di Corcia, A.; Aragona, I.; Guglielmone, G.; Di Paolo, C.; Cenni, M.; Ferranti, P.; et al. Microcystin Detection in Contaminated Fish from Italian Lakes Using ELISA Immunoassays and LC-MS/MS Analysis. In Handbook on Cyanobacteria; Gault, P.M., Marler, H.J., Eds.; Nova Science Publishers Inc.: New York, NY, USA, 2009; pp. 191–210. Available online: https://www.researchgate.net/profile/Milena_Bruno/publication/288081041_Microcystin_detection_in_contaminated_fish_from_Italian_lakes_using_elisa_immunoassays_and_Lc-MsMs_analysis/links/574701e608aef66a78b0708e.pdf (accessed on 25 May 2017).
- Wong, C.-K; Hung, P.; Lee, K.L.H.; Mok, T.; Kam, K.-M. Effect of steam cooking on distribution of paralytic shellfish toxins in different tissue compartments of scallops Patinopecten yessoensis. Food Chem. 2009, 114, 72–80. [Google Scholar] [CrossRef]
- Zhang, D.; Xie, P.; Chen, J. Effects of temperature on the stability of microcystins in muscle of fish and its consequences for food safety. Bull. Environ. Contam. Toxicol. 2010, 84, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Guzmán-Guillén, R.; Prieto, A.I.; Moreno, I.; Soria, M.E.; Cameán, A.M. Effects of thermal treatments during cooking, microwave oven and boiling, on the unconjugated microcystin concentration in muscle of fish (Oreochromis niloticus). Food Chem. Toxicol. 2011, 49, 2060–2067. [Google Scholar] [CrossRef] [PubMed]
- Freitas, M.; Azevedo, J.; Carvalho, A.P.; Campos, A.; Vasconcelos, V. Effects of storage, processing and proteolytic digestion on microcystin-LR concentration in edible clams. Food Chem. Toxicol. 2014, 66, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Adamski, M.; Żmudzki, P.; Chrapusta, E.; Bober, B.; Kaminski, A.; Zabaglo, K.; Latkowska, E.; Bialczyk, J. Effect of pH and temperature on the stability of cylindrospermopsin. Characterization of decomposition products. Algal Res. 2016, 15, 129–134. [Google Scholar] [CrossRef]
- Adamski, M.; Żmudzki, P.; Chrapusta, E.; Kaminski, A.; Bober, B.; Zabaglo, K.; Bialczyk, J. Characterization of cylindrospermopsin decomposition products formed under irradiation conditions. Algal Res. 2016, 18, 1–6. [Google Scholar] [CrossRef]
- Testai, E.; Buratti, F.M.; Funari, E.; Manganelli, M.; Vichi, S.; Arnich, N.; Biré, R.; Fessard, V.; Sialehaamoa, A. Review and analysis of occurrence, exposure and toxicity of cyanobacteria toxins in food. Eur. Food Saf. Auth. 2016, 1–309. [Google Scholar] [CrossRef]
- Domingo, J.L. Influence of Cooking Processes on the Concentrations of Toxic Metals and Various Organic Environmental Pollutants in Food: A Review of the Published Literature. Crit. Rev. Food Sci. Nutr. 2011, 51, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Gaber, M.H.; El Halim, N.A.; Khali, W.A. Effect of microwave radiation on the biophysical properties of liposomes. Bioelectromagnetics 2005, 26, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Snider, B.B.; Xie, C. Chapter 2—Total Synthesis of (±)-Cylindrospermopsin. In Strategies and Tactics Organic Synthesis; Harmata, M., Ed.; Academic Press: London, UK, 2004; Volume 4, pp. 19–39. [Google Scholar]
- Søeborg, T.; Ingerslev, F.; Halling-Sørensen, B. Chemical stability of chlortetracycline and chlortetracycline degradation products and epimers in soil interstitial water. Chemosphere 2004, 57, 1515–1524. [Google Scholar] [CrossRef] [PubMed]
- Directive 2010/63/EU of the European Parliament and of the Council on the Protection of Animals Used for Scientific Purposes. Available online: http://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2010:276:0033:0079:en:PDF (accessed on 25 May 2017).
- Guzmán-Guillén, R.; Moreno, I.M.; Prieto, A.I.; Soria-Díaz, M.E.; Vasconcelos, V.M.; Cameán, A.M. CYN determination in tissues from fresh water fish by LC-MS/MS: Validation and application in tissues from subchronically exposed tilapia (Oreochromis niloticus). Talanta 2015, 131, 452–459. [Google Scholar] [CrossRef] [PubMed]
- Makarov, A.; Denisov, E.; Lange, O.; Horning, S. Dynamic range of mass accuracy in LTQ orbitrap hybrid mass spectrometer. J. Am. Soc. Mass Spectrom. 2006, 17, 977–982. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Decomposition Products | Retention Time (min) | Putative Chemical Formula | [M + H]+ Ion Full Scan (FS) | Fragments Parallel Reaction Monitoring (PRM) | |||||
|---|---|---|---|---|---|---|---|---|---|
| m/z obs. | m/z calc. | Mass Error (ppm) | Fragment Formula | m/z obs. | m/z calc. | Mass Error (ppm) | |||
| C-1 | 1.54 | C10H15N3O5S | 290.08090 | 290.08052 | 1.32 | C10H15N3O2 | 210.12402 | 210.12370 | 1.52 |
| C10H13N3O | 192.11319 | 192.11314 | 0.27 | ||||||
| C-2A | 2.45 | C10H17N3O5S | 292.09595 | 292.09617 | −0.76 | C10H17N3O2 | 212.13928 | 212.13935 | −0.34 |
| C10H15N3O | 194.12901 | 194.12879 | 1.16 | ||||||
| C10H13N3 | ND | 176.11822 | |||||||
| C-2B | 3.81 | C10H17N3O5S | 292.09634 | 292.09617 | 0.60 | C10H17N3O2 | 212.13954 | 212.13935 | 0.89 |
| C10H15N3O | 194.12900 | 194.12879 | 1.08 | ||||||
| C10H13N3 | ND | 176.11822 | |||||||
| 7-epi-CYN | 2.87 | C15H21N5O7S | 416.12320 | 416.12345 | −0.59 | C15H21N5O4 | 336.16690 | 336.16663 | 0.80 |
| C15H19N5O3 | 318.15652 | 318.15607 | 1.44 | ||||||
| C10H15N3O4S | 274.08606 | 274.08560 | 1.67 | ||||||
| C10H15N3O | 194.12900 | 194.12879 | 1.08 | ||||||
| C10H13N3 | 176.11835 | 176.11822 | 0.70 | ||||||
| CYN | 3.25 | C15H21N5O7S | 416.12350 | 416.12345 | 0.14 | C15H21N5O4 | 336.16693 | 336.16663 | 0.89 |
| C15H19N5O3 | 318.15637 | 318.15607 | 0.96 | ||||||
| C10H15N3O4S | 274.08600 | 274.08560 | 1.44 | ||||||
| C10H15N3O | 194.12901 | 194.12879 | 1.16 | ||||||
| C10H13N3 | 176.11833 | 176.11822 | 0.61 | ||||||
| C-3A | 3.44 | C15H21N5O4 | 336.16611 | 336.16663 | −1.56 | C10H15N3O | 194.12901 | 194.12879 | 1.16 |
| C10H13N3 | ND | 176.11822 | |||||||
| C-3B | 5.06 | C15H21N5O4 | 336.16687 | 336.16663 | 0.71 | C10H15N3O | 194.12895 | 194.12879 | 0.84 |
| C10H13N3 | ND | 176.11822 | |||||||
| C-3C | 4.86 | C15H21N5O4 | 336.16748 | 336.16663 | 2.53 | C10H15N3O | 194.12903 | 194.12879 | 1.23 |
| C10H13N3 | ND | 176.11822 | |||||||
| C-3D | 4.96 | C15H21N5O4 | 336.16660 | 336.16663 | −0.10 | C10H15N3O | 194.12891 | 194.12879 | 0.61 |
| C10H13N3 | ND | 176.11822 | |||||||
| C-3E | 5.17 | C15H21N5O4 | 336.16620 | 336.16663 | −1.28 | C10H15N3O | 194.12901 | 194.12879 | 1.16 |
| C10H13N3 | ND | 176.11822 | |||||||
| C-3F | 5.27 | C15H21N5O4 | 336.16693 | 336.16663 | 0.89 | C10H15N3O | 194.12929 | 194.12879 | 2.57 |
| C10H13N3 | ND | 176.11822 | |||||||
| C-4 | 4.94 | C15H19N5O7S | 414.10715 | 414.10780 | −1.56 | C15H19N5O4 | 334.15170 | 334.15098 | 2.16 |
| C10H13N3O4S | 272.07062 | 272.06995 | 2.44 | ||||||
| C10H13N3O | 192.11334 | 192.11314 | 1.06 | ||||||
| Decomposition Products | Retention Time (min) | Putative Chemical Formula | [M + H]+ Ion Full Scan (FS) | Fragments Parallel Reaction Monitoring (PRM) | |||||
|---|---|---|---|---|---|---|---|---|---|
| m/z obs. | m/z calc. | Mass Error (ppm) | Fragment Formula | m/z obs. | m/z calc. | Mass Error (ppm) | |||
| C-1 | 1.52 | C10H15N3O5S | 290.08063 | 290.08052 | 0.38 | C10H15N3O2 | 210.12390 | 210.12370 | 0.94 |
| C10H13N3O | 192.11357 | 192.11314 | 2.25 | ||||||
| C-2A | 2.44 | C10H17N3O5S | 292.09634 | 292.09617 | 0.60 | C10H17N3O2 | 212.13951 | 212.13935 | 0.74 |
| C10H15N3O | 194.12917 | 194.12879 | 1.94 | ||||||
| C10H13N3 | 176.11784 | 176.11822 | 2.16 | ||||||
| C-2B | 3.81 | C10H17N3O5S | 292.09631 | 292.09617 | 0.50 | C10H17N3O2 | 212.13954 | 212.13935 | 0.89 |
| C10H15N3O | 194.12903 | 194.12879 | 1.23 | ||||||
| C10H13N3 | 176.11868 | 176.11822 | 2.61 | ||||||
| 7-epi-CYN | 2.88 | C15H21N5O7S | 416.12357 | 416.12345 | 0.29 | C15H21N5O4 | 336.16693 | 336.16663 | 0.89 |
| C15H19N5O3 | 318.15631 | 318.15607 | 0.77 | ||||||
| C10H15N3O4S | 274.08621 | 274.08560 | 2.22 | ||||||
| C10H15N3O | 194.12901 | 194.12879 | 1.16 | ||||||
| C10H13N3 | 176.11847 | 176.11822 | 1.39 | ||||||
| CYN | 3.24 | C15H21N5O7S | 416.12369 | 416.12345 | 0.58 | C15H21N5O4 | 336.16696 | 336.16663 | 0.98 |
| C15H19N5O3 | 318.15643 | 318.15607 | 1.15 | ||||||
| C10H15N3O4S | 274.08603 | 274.08560 | 1.55 | ||||||
| C10H15N3O | 194.12904 | 194.12879 | 1.31 | ||||||
| C10H13N3 | 176.11835 | 176.11822 | 0.70 | ||||||
| C-3A | 3.44 | C15H21N5O4 | 336.16647 | 336.16663 | −0.47 | C10H15N3O | 194.12904 | 194.12879 | 1.31 |
| C10H13N3 | ND | 176.11822 | |||||||
| C-3E | 5.14 | C15H21N5O4 | 336.16721 | 336.16663 | 1.71 | C10H15N3O | 194.12903 | 194.12879 | 1.23 |
| C10H13N3 | ND | 176.11822 | |||||||
| C-4 | 5.03 | C15H19N5O7S | 414.10718 | 414.10780 | −1.49 | C15H19N5O4 | 334.15115 | 334.15098 | 0.52 |
| C10H13N3O4S | 272.07028 | 272.06995 | 1.21 | ||||||
| C10H13N3O | 192.11337 | 192.11314 | 1.22 | ||||||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prieto, A.I.; Guzmán-Guillén, R.; Valderrama-Fernández, R.; Jos, Á.; Cameán, A.M. Influence of Cooking (Microwaving and Broiling) on Cylindrospermopsin Concentration in Muscle of Nile Tilapia (Oreochromis niloticus) and Characterization of Decomposition Products. Toxins 2017, 9, 177. https://doi.org/10.3390/toxins9060177
Prieto AI, Guzmán-Guillén R, Valderrama-Fernández R, Jos Á, Cameán AM. Influence of Cooking (Microwaving and Broiling) on Cylindrospermopsin Concentration in Muscle of Nile Tilapia (Oreochromis niloticus) and Characterization of Decomposition Products. Toxins. 2017; 9(6):177. https://doi.org/10.3390/toxins9060177
Chicago/Turabian StylePrieto, Ana I., Remedios Guzmán-Guillén, Rocío Valderrama-Fernández, Ángeles Jos, and Ana M. Cameán. 2017. "Influence of Cooking (Microwaving and Broiling) on Cylindrospermopsin Concentration in Muscle of Nile Tilapia (Oreochromis niloticus) and Characterization of Decomposition Products" Toxins 9, no. 6: 177. https://doi.org/10.3390/toxins9060177