The Pattern of Fatty Acids Displaced by EPA and DHA Following 12 Months Supplementation Varies between Blood Cell and Plasma Fractions

Abstract
:1. Introduction
2. Experimental Section
2.1. Original Trial
2.2. Data Analysis
3. Results
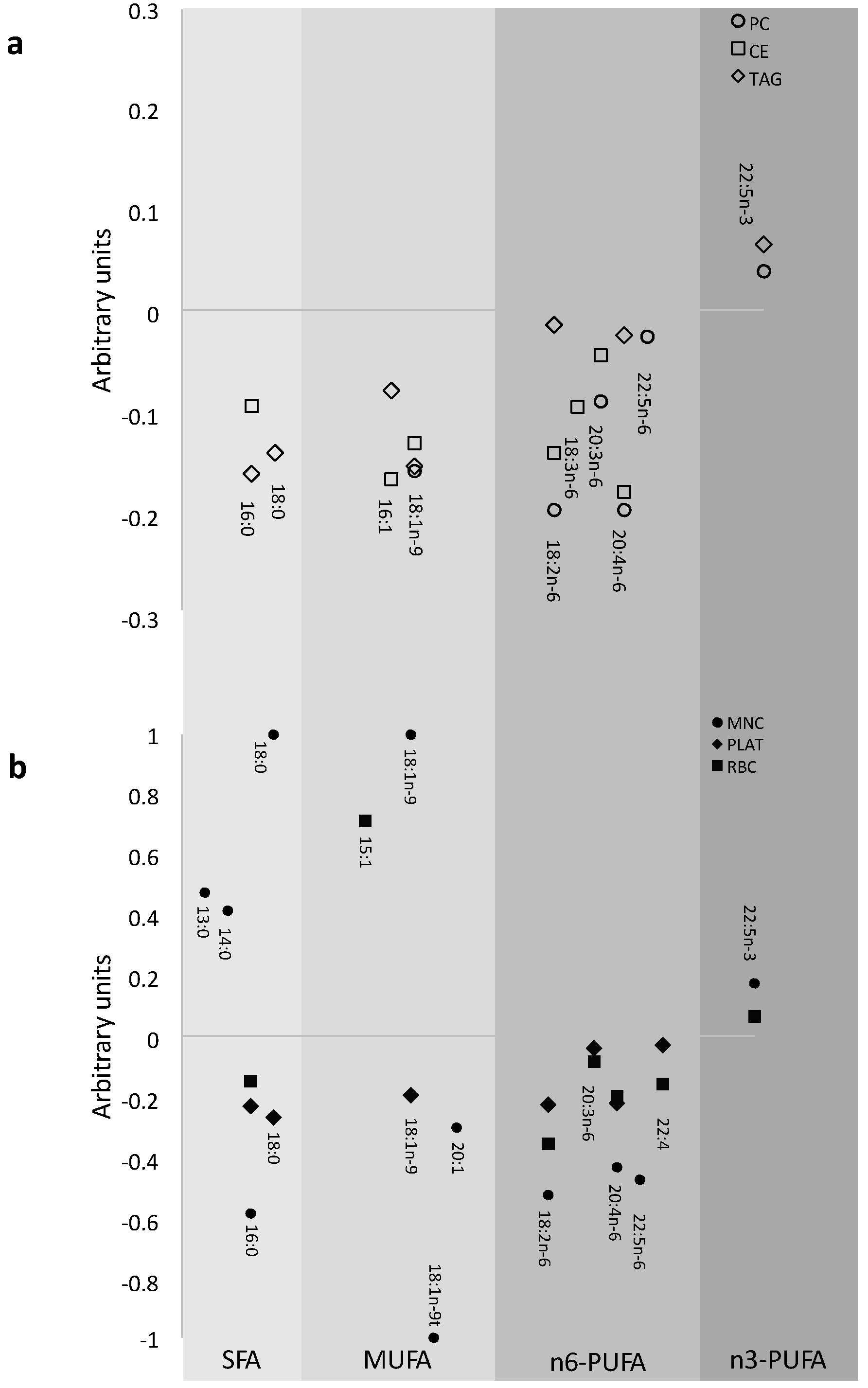
3.1. Prevalence of FA in the Different Pools at Baseline

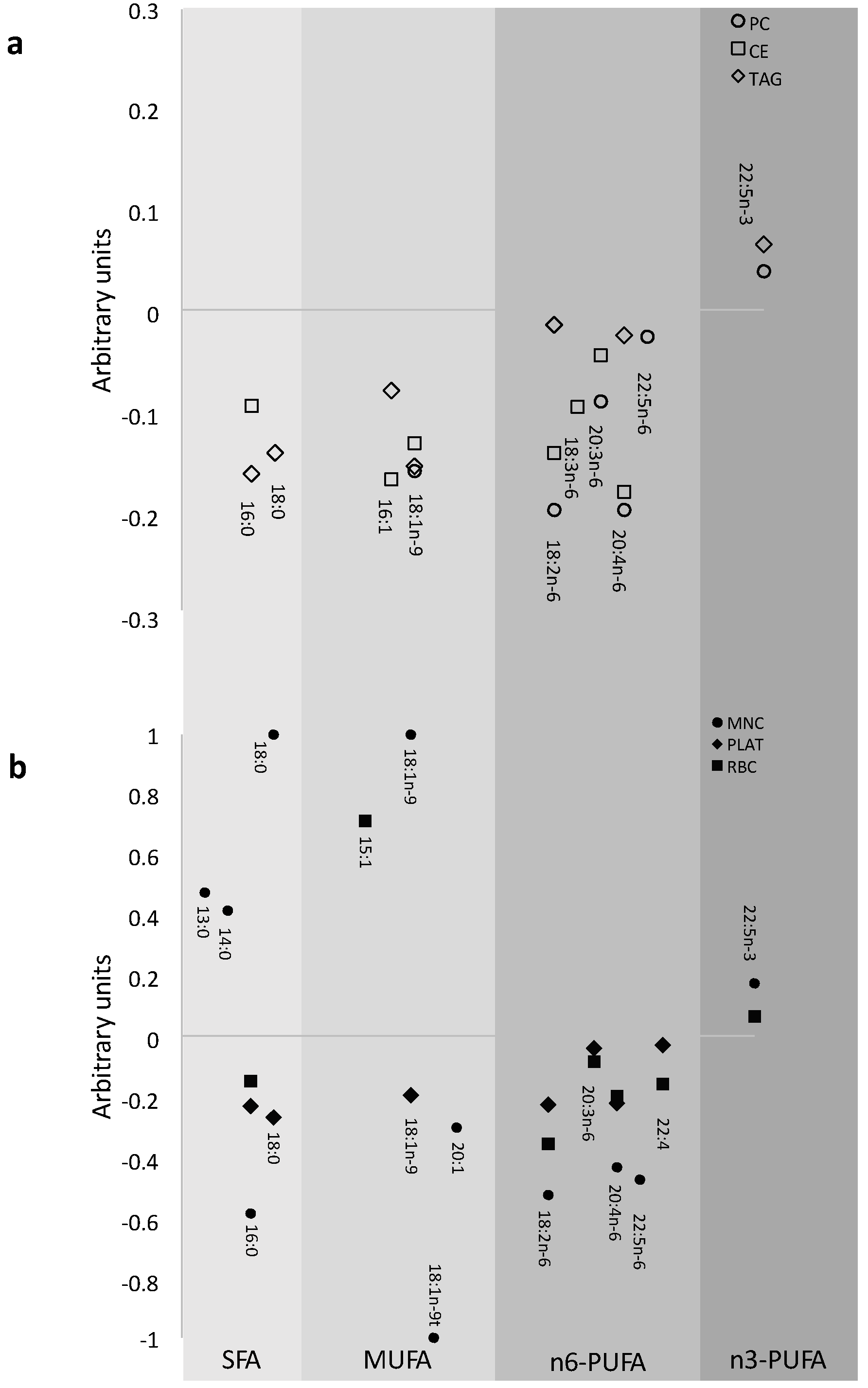{kind=link}
{kind=link}
{kind=link}
| Fatty Acid | MNC | PLAT | RBC | CE | PC | TAG |
|---|---|---|---|---|---|---|
| 10:0 | 0.14 (0.25) | 0.03 (0.05) | 0.00 (0.07) | 0.06 (0.04) | 0.00 (0.00) | 0.00 (0.00) |
| 12:0 | 0.05 (0.16) | 0.08 (0.08) | 0.00 (0.00) | 0.06 (0.04) | 0.00 (0.00) | 0.06 (0.15) |
| 13:0 | 0.03 (0.11) | 0.00 (0.00) | 0.00 (0.00) | 0.13 (0.14) | 0.00 (0.00) | 0.00 (0.13) |
| 14:0 | 0.41 (0.35) | 0.74 (0.40) | 0.25 (0.12) | 0.49 (0.38) | 0.27 (0.14) | 1.49 (1.09) |
| 14:1n-9 | 0.16 (0.21) | 0.06 (0.09) | 0.00 (0.00) | 0.04 (0.02) | 0.00 (0.00) | 0.08 (0.10) |
| 15:0 | 0.19 (0.14) | 0.18 (0.07) | 0.09 (0.14) | 0.15 (0.07) | 0.15 (0.21) | 0.27 (0.08) |
| 15:1 | 0.14 (0.36) | 0.07 (0.21) | 2.21 (1.99) | 0.05 (0.03) | 0.05 (0.03) | 0.05 (0.04) |
| 16:0 | 18.2 (2.94) | 20.7 (2.54) | 19.3 (1.83) | 11.3 (1.14) | 28.9 (1.80) | 25.6 (3.89) |
| 16:1n-7 | 1.13 (1.44) | 1.97 (1.14) | 0.38 (0.19) | 2.80 (1.74) | 0.70 (0.35) | 3.49 (1.50) |
| 17:0 | 0.32 (0.20) | 0.26 (0.06) | 0.27 (0.08) | 0.07 (0.08) | 0.35 (0.10) | 0.30 (0.13) |
| 17:1n-8 | 0.30 (0.37) | 0.91 (0.90) | 4.05 (0.63) | 0.05 (0.05) | 0.19 (0.11) | 0.19 (0.10) |
| 18:0 | 17.4 (4.95) | 8.67 (3.07) | 16.2 (1.27) | 0.76 (0.25) | 12.8 (1.39) | 3.07 (1.02) |
| 18:1n-9t | 1.94 (1.33) | 0.17 (0.07) | 0.16 (0.22) | 1.31 (0.43) | 1.55 (0.32) | 2.44 (0.62) |
| 18:1n-9c | 16.2 (2.75) | 19.9 (3.55) | 13.1 (1.45) | 19.1 (2.66) | 11.2 (1.69) | 39.8 (3.96) |
| 18:2n-6t | 0.00 (0.19) | 0.14 (0.12) | 0.21 (0.26) | 0.00 (0.00) | 0.00 (0.00) | 0.00 (0.00) |
| 18:2n-6c | 11.4 (9.04) | 25.1 (7.42) | 9.93 (1.94) | 51.7 (6.42) | 22.4 (3.76) | 16.6 (5.63) |
| 18:3n-6 | 0.20 (0.33) | 0.43 (0.25) | 0.09 (0.16) | 0.95 (0.58) | 0.11 (0.08) | 0.34 (0.23) |
| 18:3n-3 | 0.22 (0.30) | 0.62 (0.31) | 0.32 (0.33) | 0.56 (0.25) | 0.22 (0.15) | 1.00 (0.49) |
| 20:0 | 0.53 (0.62) | 0.26 (0.19) | 0.10 (0.16) | 0.13 (0.05) | 0.15 (0.07) | 0.28 (0.16) |
| 20:1n-9 | 0.60 (1.00) | 0.35 (0.33) | 0.36 (0.22) | 0.06 (0.09) | 0.20 (0.10) | 0.29 (0.14) |
| 20:2n-6 | 0.46 (0.60) | 0.20 (0.07) | 0.22 (0.18) | 0.07 (0.05) | 0.34 (0.11) | 0.15 (0.09) |
| 20:3n-3 | 0.04 (0.19) | 0.10 (0.06) | 0.15 (0.27) | 0.00 (0.00) | 0.00 (0.22) | 0.00 (0.00) |
| 20:3n-6 | 1.88 (1.42) | 1.63 (0.50) | 1.75 (0.56) | 0.76 (0.26) | 3.30 (1.11) | 0.29 (0.14) |
| 20:4n-6 | 16.0 (9.19) | 9.54 (4.18) | 15.5 (2.14) | 6.59 (2.10) | 9.55 (2.39) | 1.55 (0.82) |
| 20:4n-4 | 0.22 (0.50) | 0.00 (0.00) | 0.00 (0.00) | 0.08 (0.04) | 0.18 (0.11) | 0.07 (0.11) |
| 20:5n-3 | 0.57 (0.46) | 1.00 (0.52) | 1.42 (1.12) | 0.91 (0.64) | 1.05 (0.57) | 0.22 (0.41) |
| 21:0 | 0.30 (0.50) | 0.13 (0.17) | 0.37 (0.46) | 0.03 (0.04) | 0.06 (0.09) | 0.00 (0.00) |
| 22:0 | 0.18 (0.32) | 0.23 (0.10) | 0.20 (0.24) | 0.00 (0.03) | 0.03 (0.14) | 0.00 (0.00) |
| 22:2n-6 | 0.07 (0.21) | 0.00 (0.00) | 0.00 (0.00) | 0.00 (0.00) | 0.00 (0.00) | 0.00 (0.08) |
| 22:4n-6 | 0.76 (1.00) | 0.46 (0.40) | 2.80 (0.89) | 0.03 (0.04) | 0.34 (0.19) | 0.14 (0.07) |
| 22:5n-3 | 1.48 (0.84) | 0.88 (0.26) | 3.11 (0.60) | 0.07 (0.08) | 0.95 (0.28) | 0.30 (0.43) |
| 22:5n-6 | 0.37 (0.74) | 0.13 (0.05) | 0.47 (0.23) | 0.04 (0.04) | 0.23 (0.18) | 0.17 (0.22) |
| 22:6n-3 | 1.86 (0.76) | 1.97 (0.71) | 5.34 (1.88) | 0.63 (0.34) | 3.47 (1.51) | 0.72 (0.46) |
| 23:0 | 0.04 (0.17) | 0.05 (0.07) | 0.00 (0.00) | 0.00 (0.00) | 0.00 (0.00) | 0.00 (0.26) |
| 24:0 | 0.60 (1.16) | 0.10 (0.05) | 0.38 (0.13) | 0.00 (0.00) | 0.00 (0.00) | 0.00 (0.00) |
| 24:1n-9 | 0.20 (0.28) | 0.00 (0.00) | 0.00 (0.00) | 0.04 (0.04) | 0.25 (0.14) | 0.00 (0.13) |
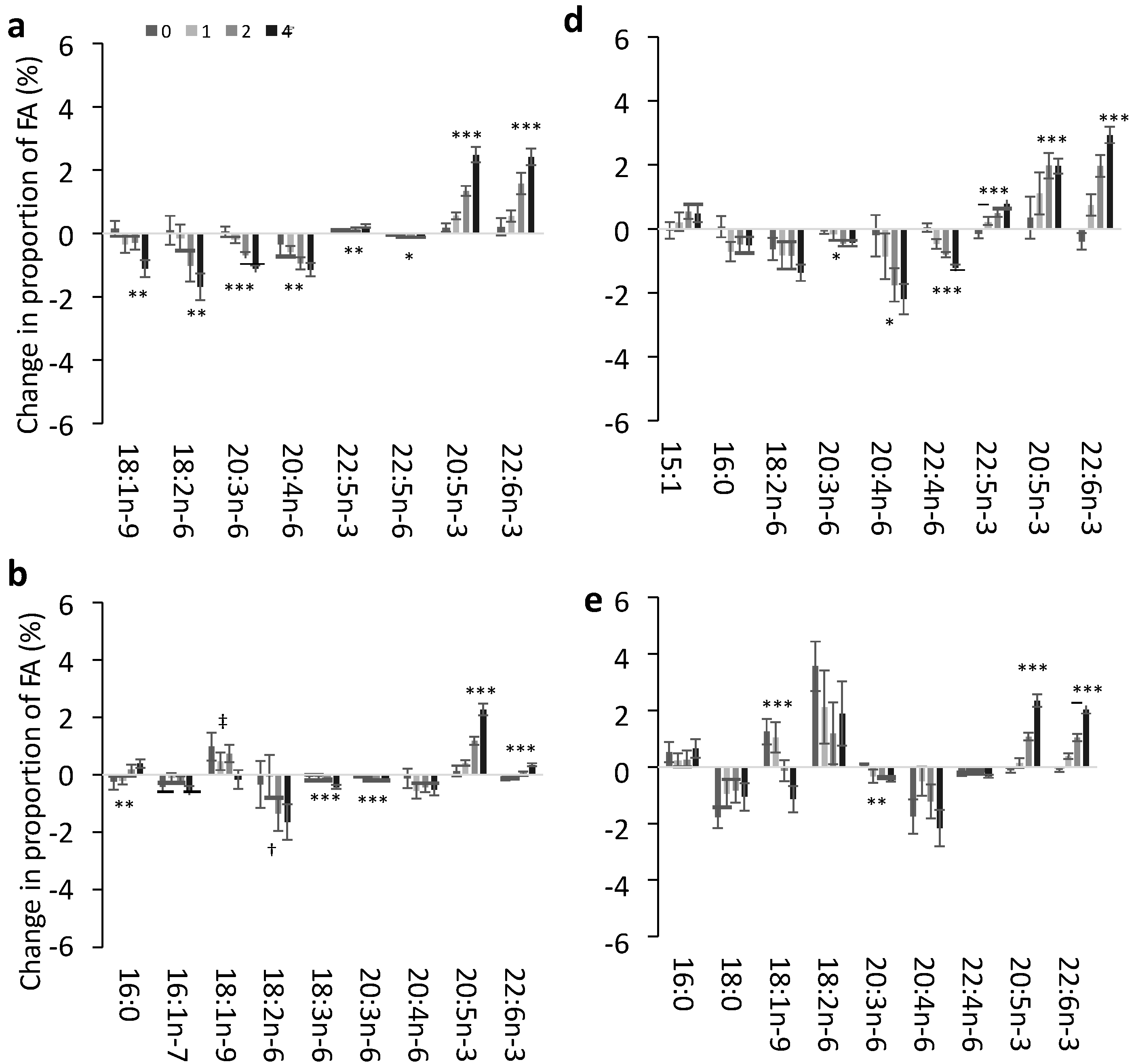
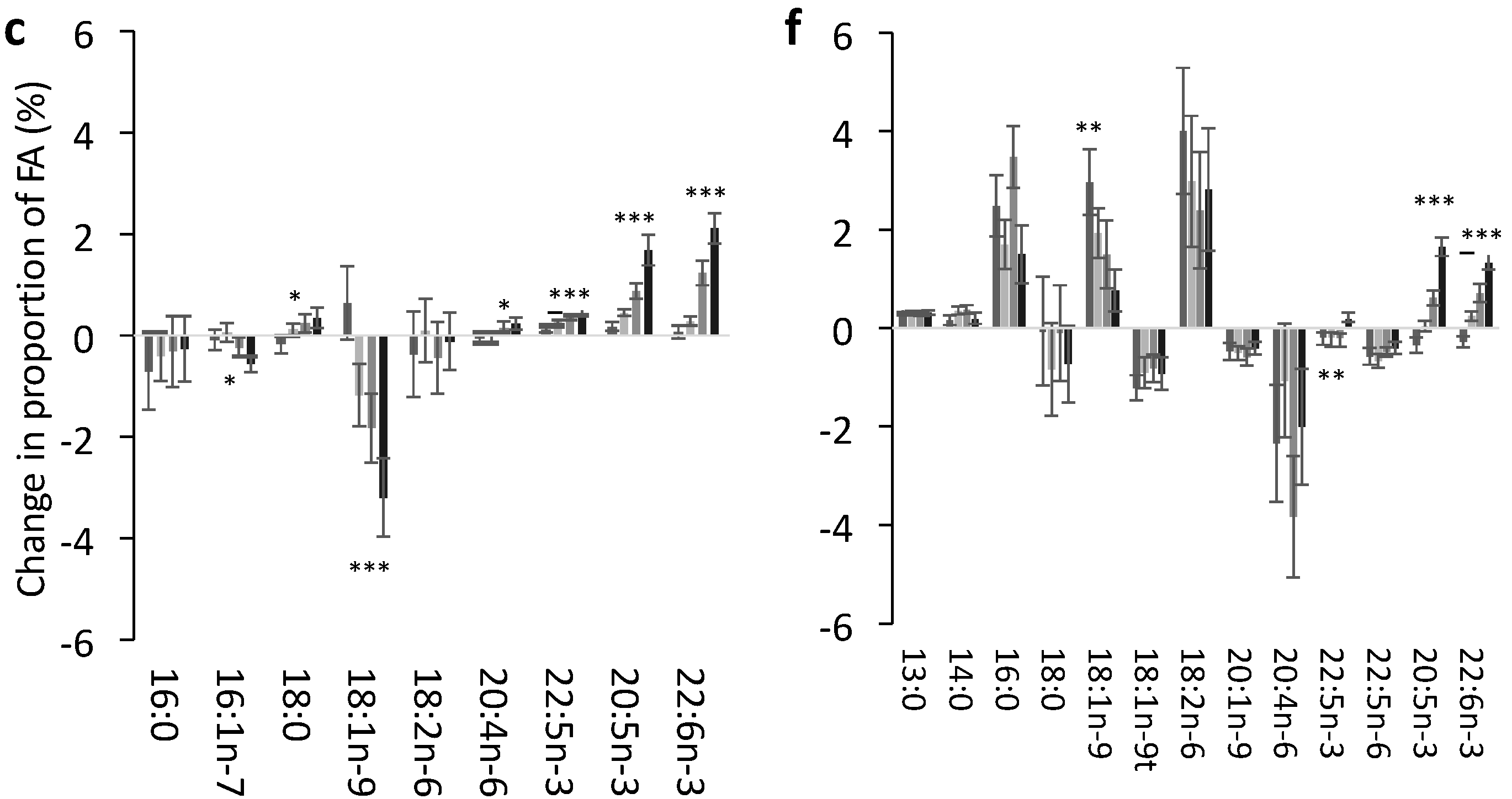
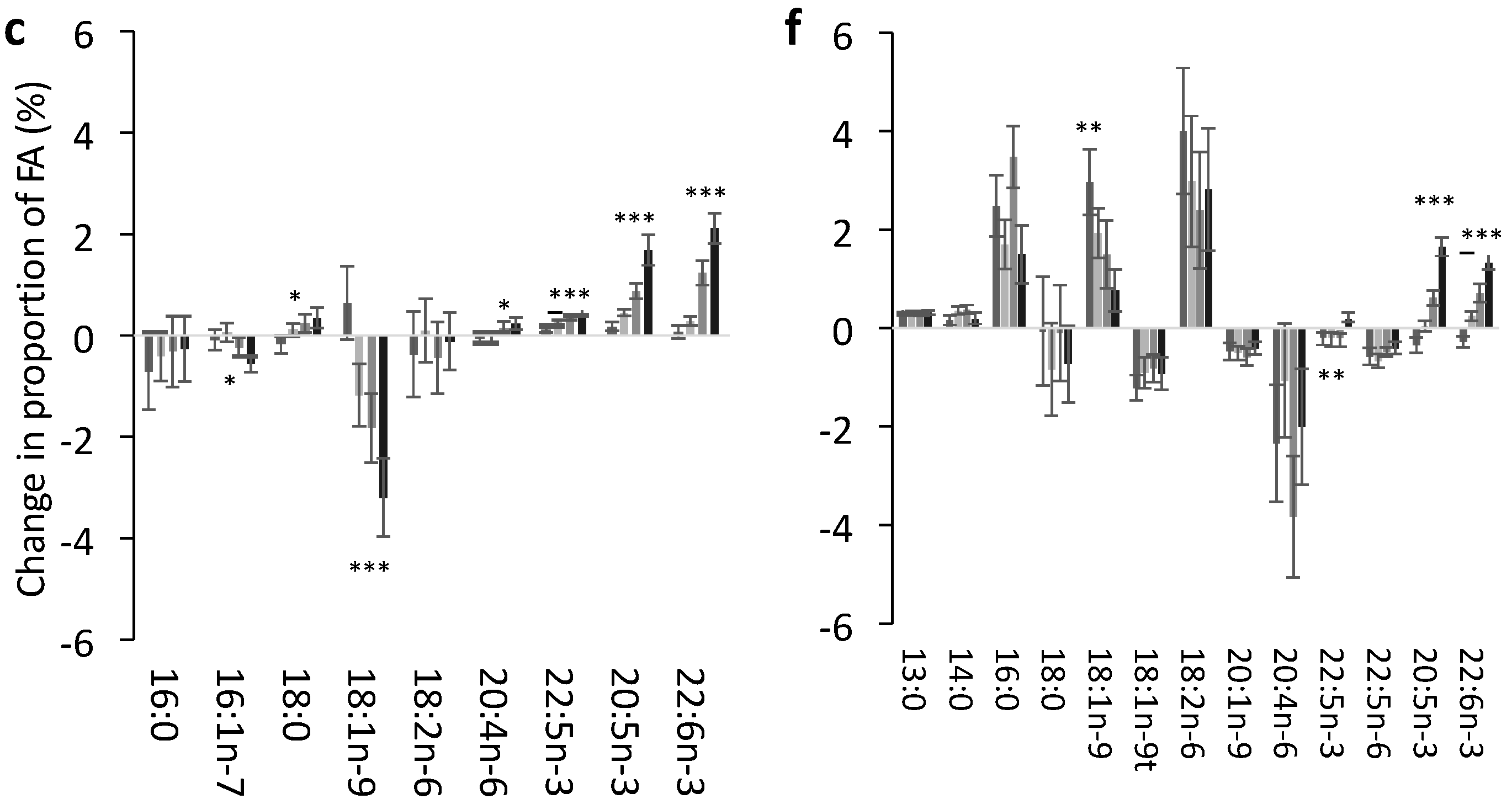
3.2. Change in FA Profile in Each Lipid Pool
3.3. Effect of Dose of EPA and DHA on the Change in Identified FA in Each Pool
3.4. Impact of EPA and DHA Supplementation on the FA Profile
| n-6:n-3 PUFA Ratio | Change in n-6:n 3 PUFA Ratio ‡ | Overall Effect of Dose at 12 Months (p) | |||
|---|---|---|---|---|---|
| 0 Portion Value † | 1 Portion | 2 Portion | 4 Portions | ||
| Plasma PC | 14.4 ± 0.59 | −3.32 | −5.41 | −7.48 | <0.0001 |
| (−4.90, −1.73) | (−7.06, −3.77) | (−9.09, −5.87) | |||
| Plasma CE | 35.5 ± 1.50 | −7.54 | −14.5 | −20.7 | <0.0001 |
| (−11.5, −3.51) | (−18.7, −10.4) | (−24.8, −16.6) | |||
| Plasma TAG | 10.0 ± 0.49 | −1.35 | −2.88 | −4.59 | <0.0001 |
| (−2.65, −0.05) | (−4.23, −1.54) | (−5.90, −3.27) | |||
| RBC | 4.90 ± 0.27 | −0.82 | −1.31 | −1.81 | <0.0001 |
| (−1.54, −1.04) | (−2.06, −0.55) | (−2.54, −1.07) | |||
| PLAT | 15.3 ± 0.70 | −4.17 | −6.56 | −9.76 | <0.0001 |
| (−5.69, −2.64) | (−8.13, −4.99) | (−11.3, −8.22) | |||
| MNC | 16.2 ± 0.69 | −3.20 | −6.24 | −8.71 | <0.0001 |
| (−5.04, −1.36) | (−8.15, −4.33) | (−10.6, −6.85) | |||

4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Deckelbaum, R.; Torrejon, C. The omega-3 fatty acid nutritional landscape: Health benefits and sources. J. Nutr. 2012, 142, 587S–591S. [Google Scholar] [CrossRef] [PubMed]
- Saravanan, P.; Davidson, N.C.; Schmidt, E.B.; Calder, P.C. Cardiovascular effects of marine omega-3 fatty acids. Lancet 2010, 376, 540–550. [Google Scholar] [CrossRef]
- Calder, P.C. Very long chain omega-3 (n-3) fatty acids and human health. Eur. J. Lipid Sci. Technol. 2014, 116, 1280–1300. [Google Scholar] [CrossRef]
- Hoy, S.; Keating, G. Omega-3 ethylester concentrate. Drugs 2009, 69, 1077–1105. [Google Scholar] [CrossRef] [PubMed]
- Bradberry, J.C.; Hilleman, D.E. Overview of omega-3 fatty acid therapies. P T. 2013, 38, 681–691. [Google Scholar] [PubMed]
- Yates, C.M.; Calder, P.C.; Ed Rainger, G. Pharmacology and therapeutics of omega-3 polyunsaturated fatty acids in chronic inflammatory disease. P T. 2014, 141, 272–282. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Marine omega-3 fatty acids and inflammatory processes: Effects, mechanisms and clinical relevance. Biochim. Biophys. Acta 2015, 1851, 469–484. [Google Scholar] [CrossRef] [PubMed]
- Arterburn, L.M.; Hall, E.B.; Oken, H. Distribution, interconversion, and dose response of n-3 fatty acids in humans. Am. J. Clin. Nutr. 2006, 83, S1467–1476S. [Google Scholar]
- Lovegrove, J.A.; Lovegrove, S.S.; Lesauvage, S.V.; Brady, L.M.; Saini, N.; Minihane, A.M.; Williams, C.M. Moderate fish-oil supplementation reverses low-platelet, long-chain n-3 polyunsaturated fatty acid status and reduces plasma triacylglycerol concentrations in British Indo-Asians. Am. J. Clin. Nutr. 2004, 79, 974–982. [Google Scholar] [PubMed]
- Krul, E.S.; Lemke, S.L.; Mukherjea, R.; Taylor, M.L.; Goldstein, D.A.; Su, H.; Liu, P.; Lawless, A.; Harris, W.S.; Maki, K.C. Effects of duration of treatment and dosage of eicosapentaenoic acid and stearidonic acid on red blood cell eicosapentaenoic acid content. Prostaglandins Leukot. Essent. Fatty Acids 2012, 86, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Harris, W.S.; Sands, S.A.; Windsor, S.L.; Ali, H.A.; Stevens, T.L.; Magalski, A.; Porter, C.B.; Borkon, A.M. Omega-3 fatty acids in cardiac biopsies from heart transplantation patients. Circulation 2004, 110, 1645–1649. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Mechanisms of action of (n-3) fatty acids. J. Nutr. 2012, 142, 592S–599S. [Google Scholar] [CrossRef] [PubMed]
- Browning, L.M.; Walker, C.G.; Mander, A.P.; West, A.L.; Madden, J.; Gambell, J.M.; Young, S.; Wang, L.; Jebb, S.A.; Calder, P.C. Incorporation of eicosapentaenoic and docosahexaenoic acids into lipid pools when given as supplements providing doses equivalent to typical intakes of oily fish. Am. J. Clin. Nutr. 2012, 96, 748–758. [Google Scholar] [CrossRef] [PubMed]
- Walker, C.G.; Browning, L.M.; Mander, A.P.; Madden, J.; West, A.L.; Calder, P.C.; Jebb, S.A. Age and sex differences in the incorporation of EPA and DHA into plasma fractions, cells and adipose tissue in humans. Br. J. Nutr. 2014, 111, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Fitt, E.; Cole, D.; Ziauddeen, N.; Pell, D.; Stickley, E.; Harvey, A.; Stephen, A.M. DINO (Diet In Nutrients Out) – an integrated dietary assessment system. Public Health Nutr. 2014, 18, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Leaf, D.A.; Connor, W.E.; Barstad, L.; Sexton, G. Incorporation of dietary n-3 fatty acids into the fatty acids of human adipose tissue and plasma lipid classes. Am. J. Clin. Nutr. 1995, 62, 68–73. [Google Scholar] [PubMed]
- Dawczynski, C.; Massey, K.A.; Ness, C.; Kiehntopf, M.; Stepanow, S.; Platzer, M.; Grün, M.; Nicolaou, A.; Jahreis, G. Randomized placebo-controlled intervention with n-3 LC-PUFA-supplemented yoghurt: Effects on circulating eicosanoids and cardiovascular risk factors. Clin. Nutr. 2013, 32, 686–696. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.-Y.; Chapkin, R.S. Importance of dietary γ-linolenic acid in human health and nutrition. J. Nutr. 1998, 128, 1411–1414. [Google Scholar] [PubMed]
- Lund, E.; Harvey, L.; Ladha, S.; Clark, D.; Johnson, I. Effects of dietary fish oil supplementation on the phospholipid composition and fluidity of cell membranes from human volunteers. Ann. Nutr. Metab. 1999, 43, 290–300. [Google Scholar] [CrossRef] [PubMed]
- Hodson, L.; Skeaff, C.; Fielding, B. Fatty acid composition of adipose tissue and blood in humans and its use as a biomarker of dietary intake. Prog. Lipid Res. 2008, 47, 348–380. [Google Scholar] [CrossRef] [PubMed]
- Popp-Snijders, C.; Schouten, J.A.; van Blitterswijk, W.J.; van der Veen, E.A. Changes in membrane lipid composition of human erythrocytes after dietary supplementation of (n-3) polyunsaturated fatty acids. Maintenance of membrane fluidity. Biochim. Biophys. Acta 1986, 854, 31–37. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walker, C.G.; West, A.L.; Browning, L.M.; Madden, J.; Gambell, J.M.; Jebb, S.A.; Calder, P.C. The Pattern of Fatty Acids Displaced by EPA and DHA Following 12 Months Supplementation Varies between Blood Cell and Plasma Fractions. Nutrients 2015, 7, 6281-6293. https://doi.org/10.3390/nu7085285
Walker CG, West AL, Browning LM, Madden J, Gambell JM, Jebb SA, Calder PC. The Pattern of Fatty Acids Displaced by EPA and DHA Following 12 Months Supplementation Varies between Blood Cell and Plasma Fractions. Nutrients. 2015; 7(8):6281-6293. https://doi.org/10.3390/nu7085285
Chicago/Turabian StyleWalker, Celia G., Annette L. West, Lucy M. Browning, Jackie Madden, Joanna M. Gambell, Susan A. Jebb, and Philip C. Calder. 2015. "The Pattern of Fatty Acids Displaced by EPA and DHA Following 12 Months Supplementation Varies between Blood Cell and Plasma Fractions" Nutrients 7, no. 8: 6281-6293. https://doi.org/10.3390/nu7085285