
Vegetation Responses to Climate Variability in the Northern Arid to Sub-Humid Zones of Sub-Saharan Africa


Abstract
:
1. Introduction
2. Materials and Methods
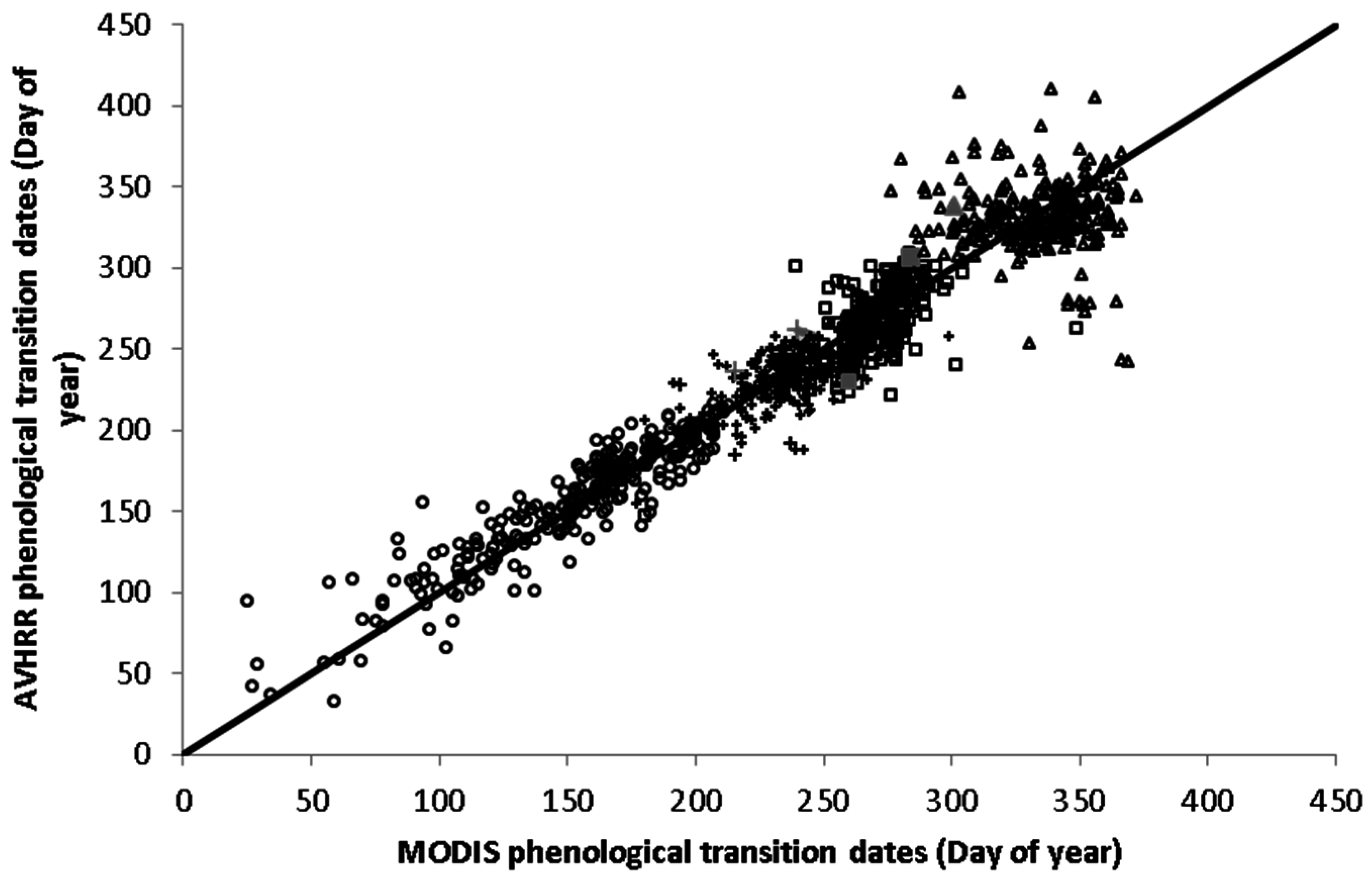
2.1. Remote Sensing Data
2.2. Meteorological and Land Cover Data
2.3. Estimating Phenological Transition Dates and the Length of the Growing Season
2.4. Relationship of Annual ΣNDVI with Annual Total Precipitation
2.5. Relationship of Growing Season ΣNDVI with Intraseasonal Precipitation Distribution
2.6. Relationship of Growing Season ΣNDVI with Temperature and Humidity
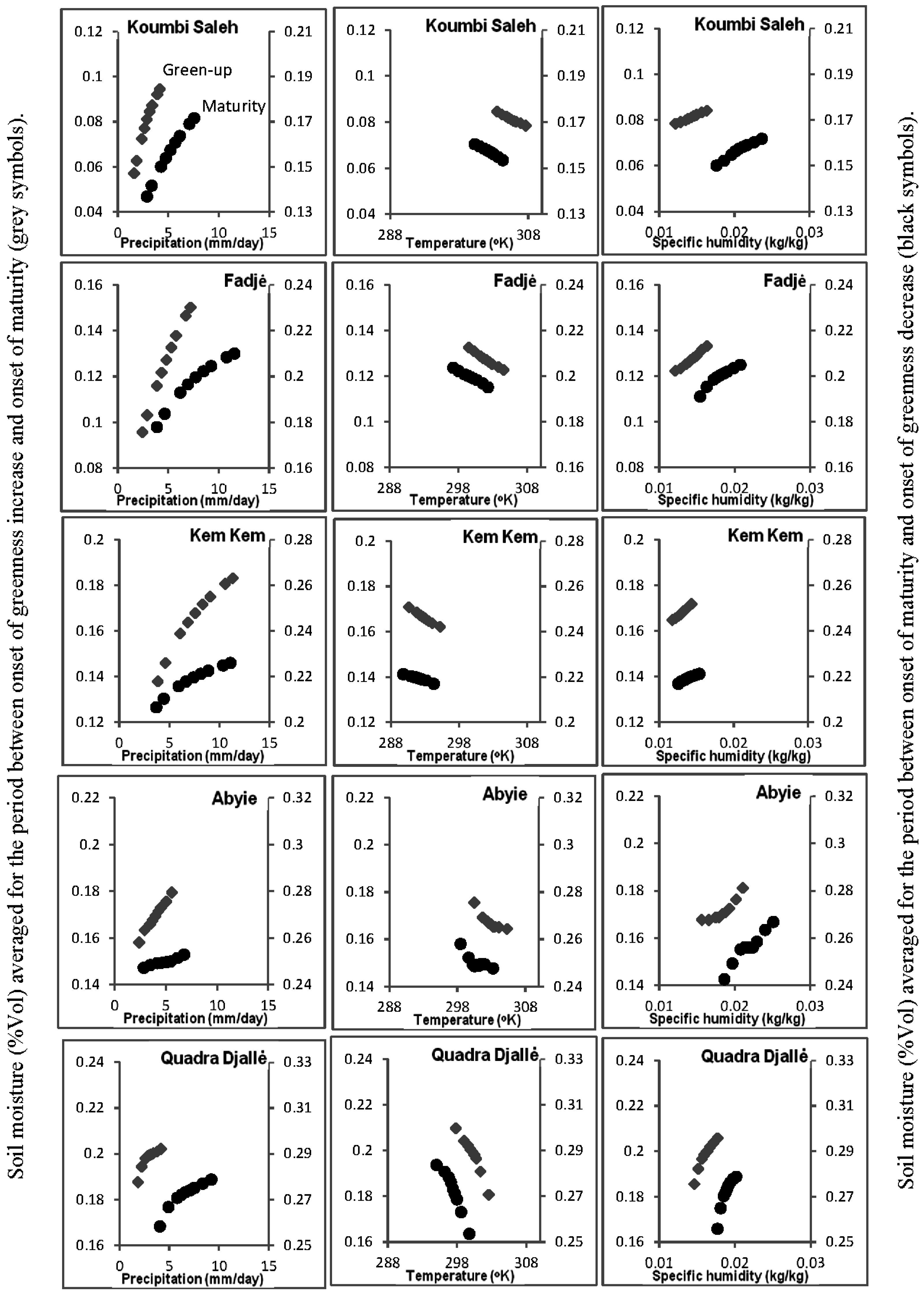
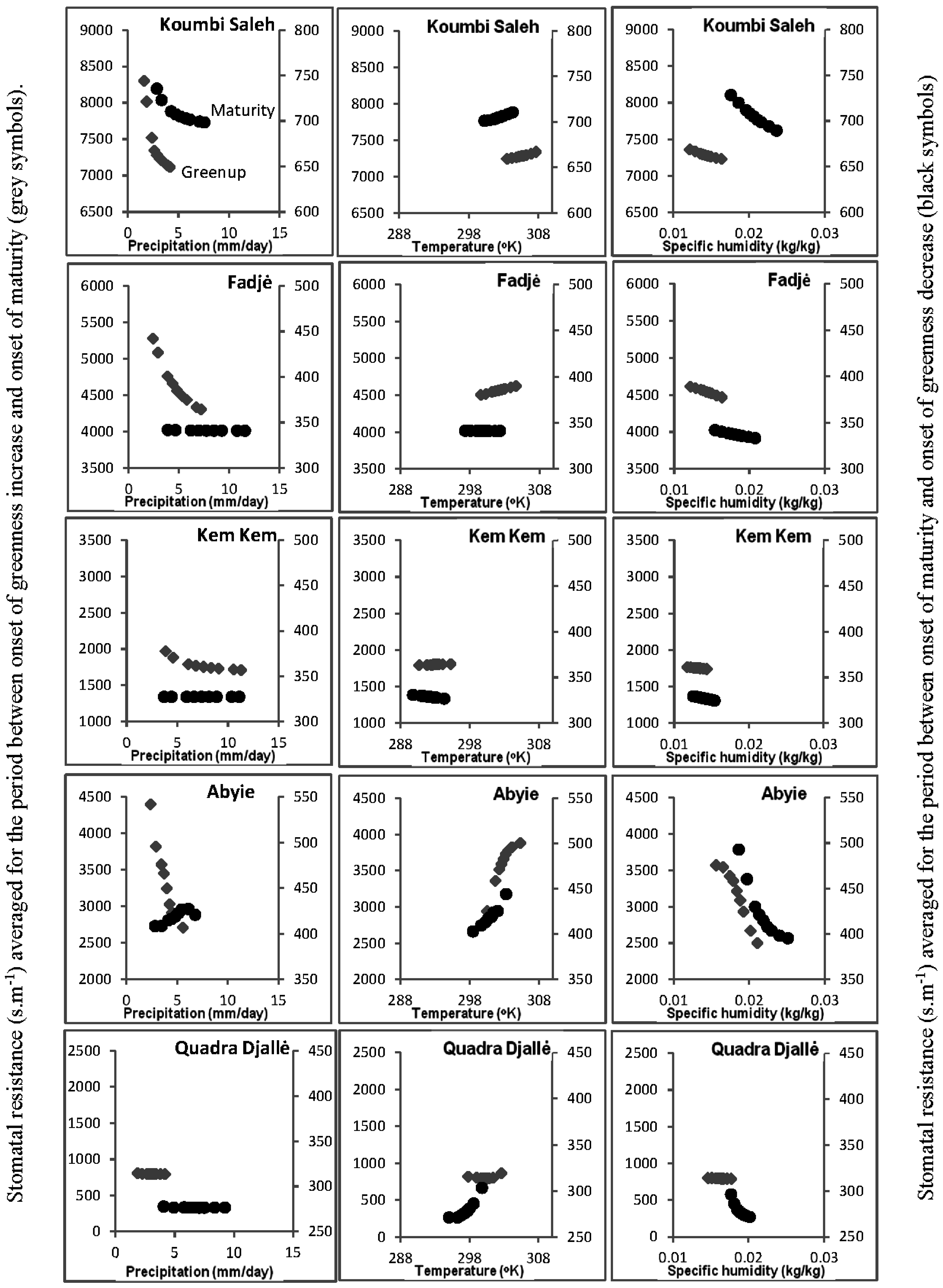
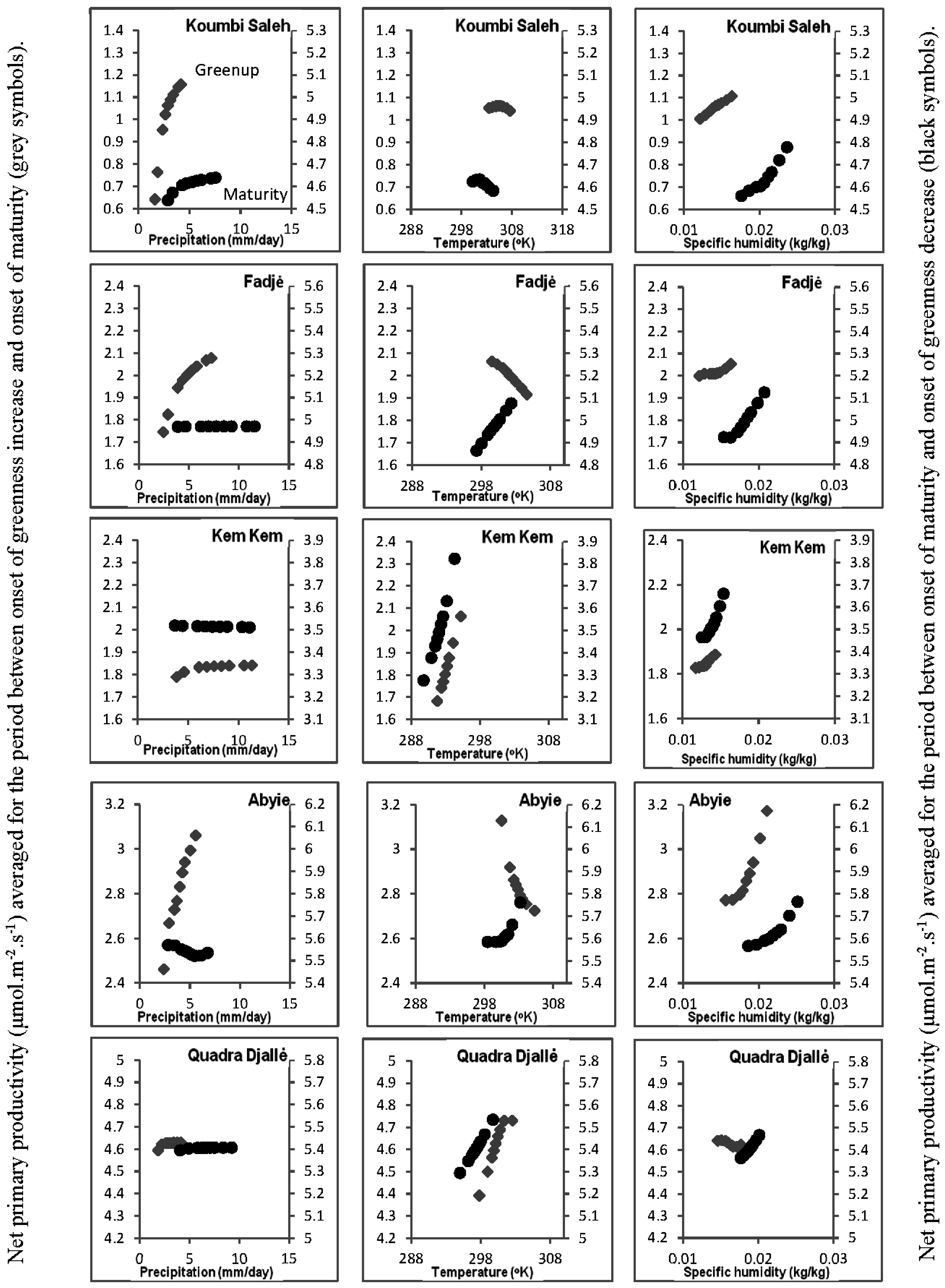
2.7. Soil-Vegetation-Atmosphere Transfer Modeling
3. Results
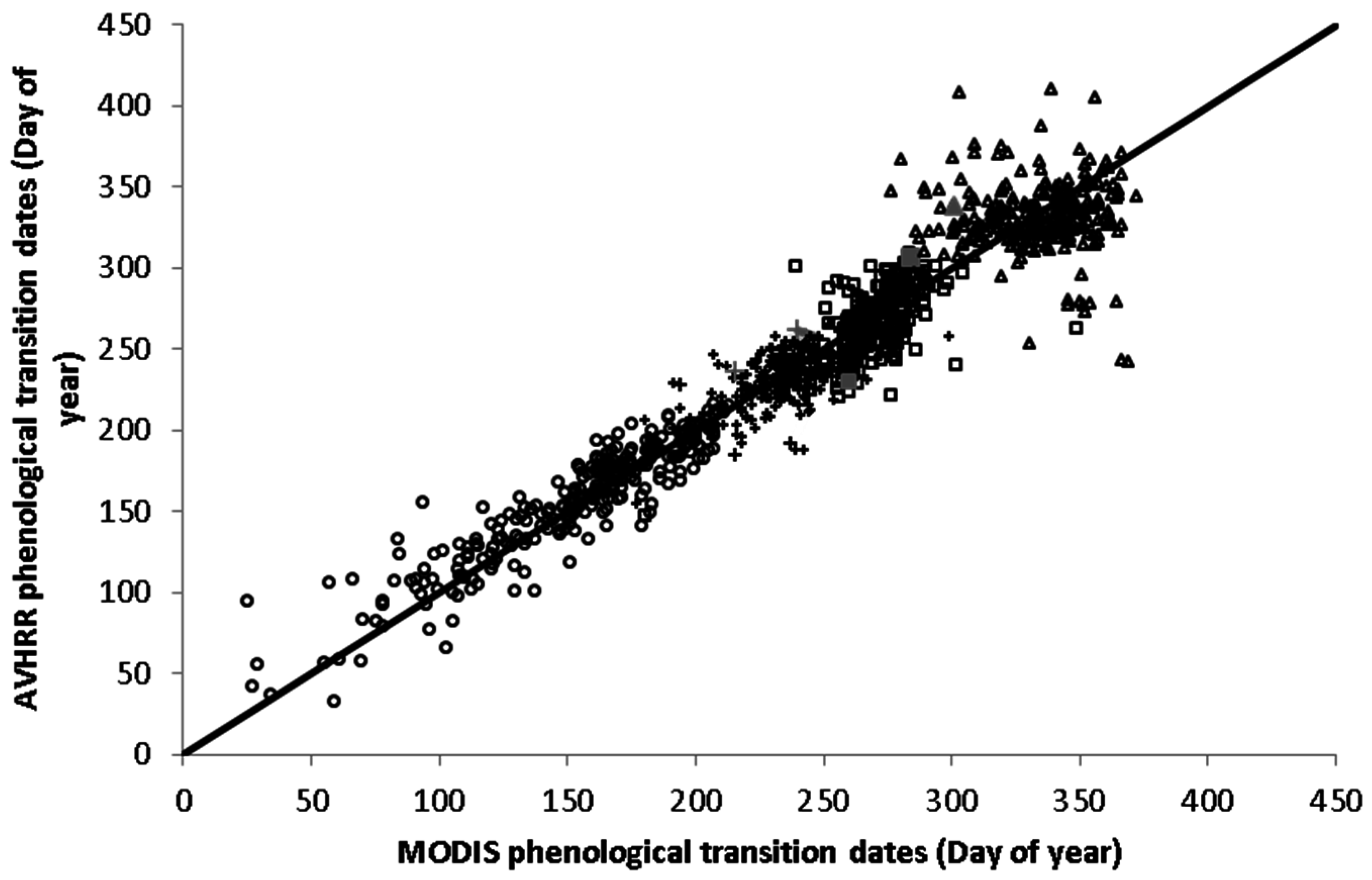
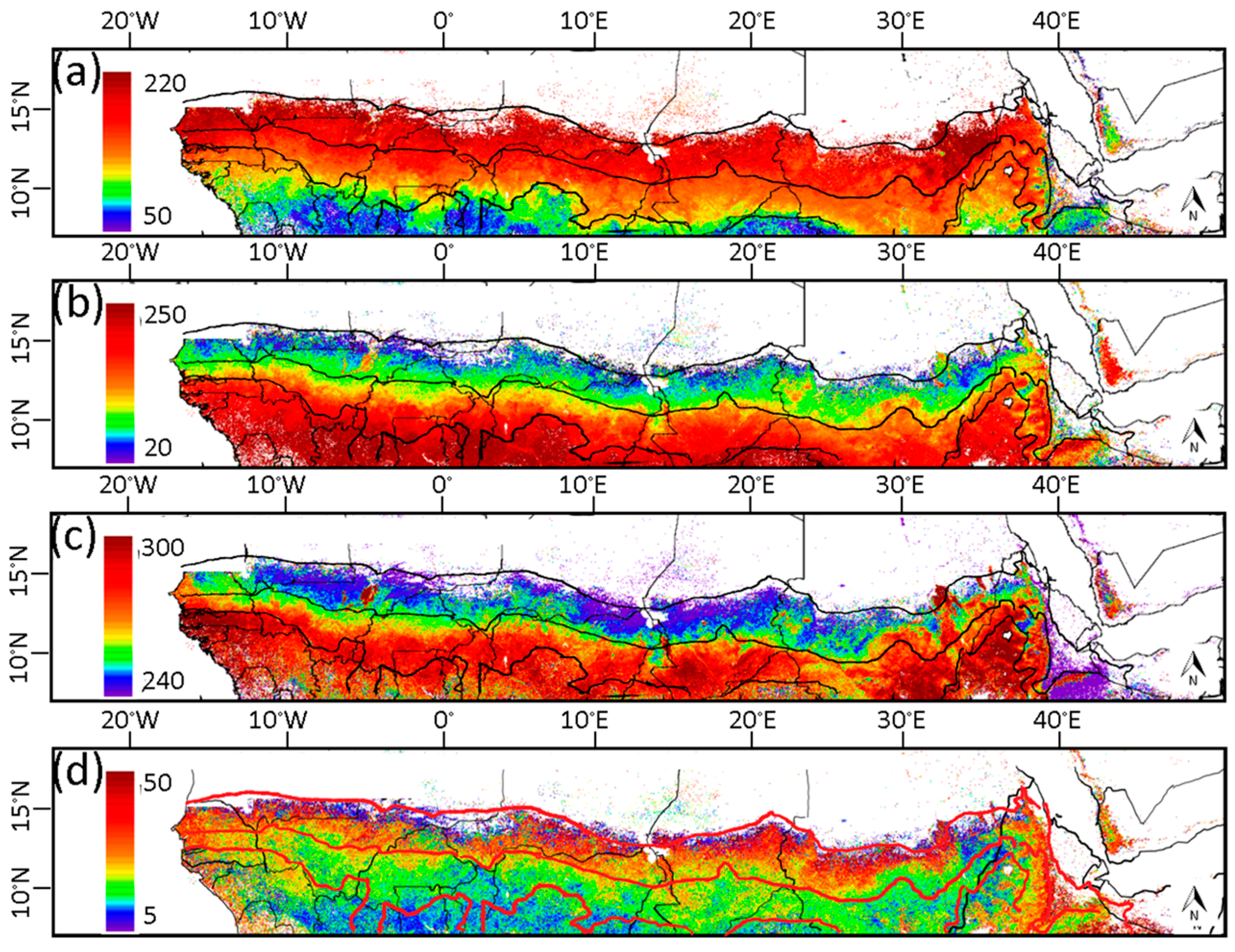
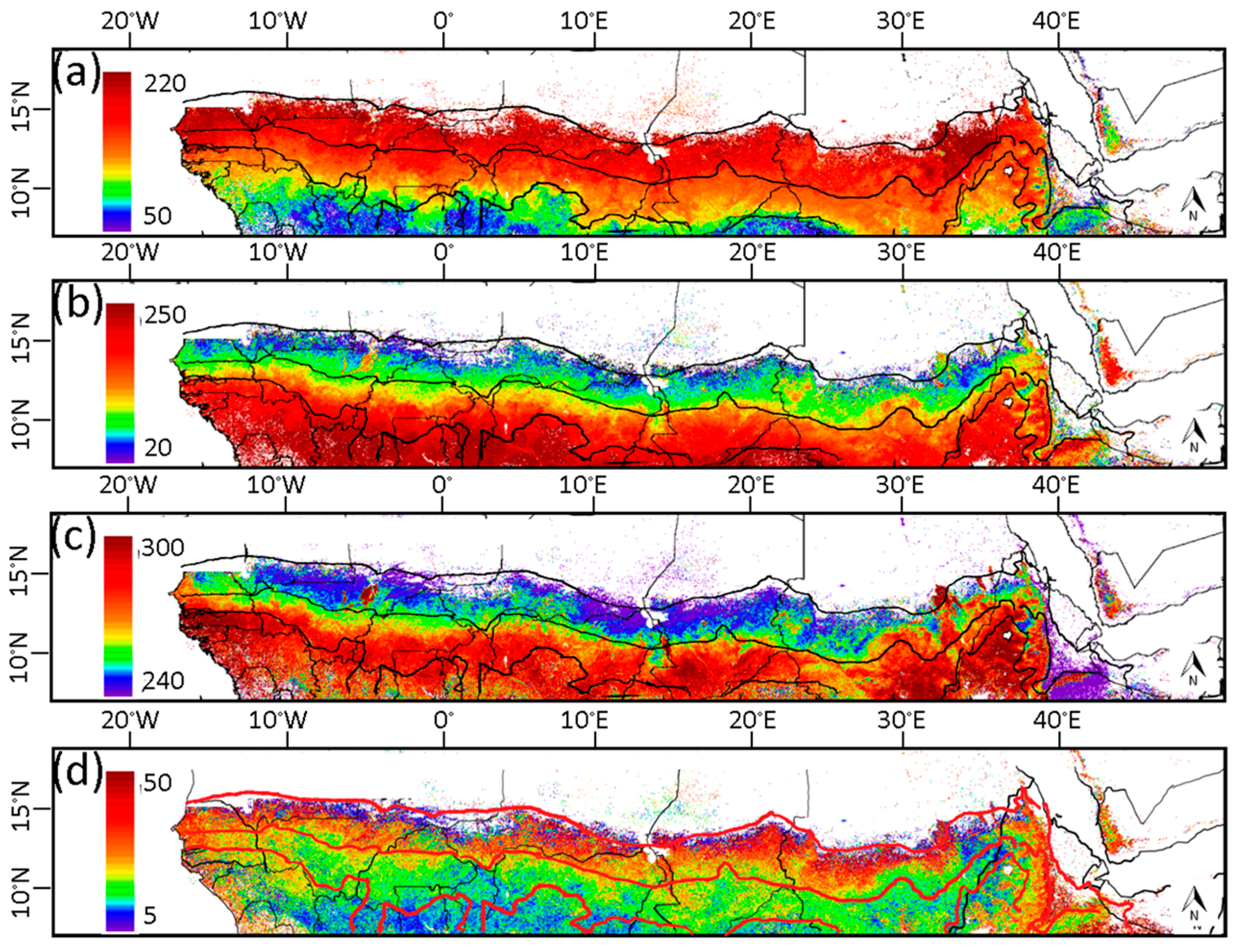
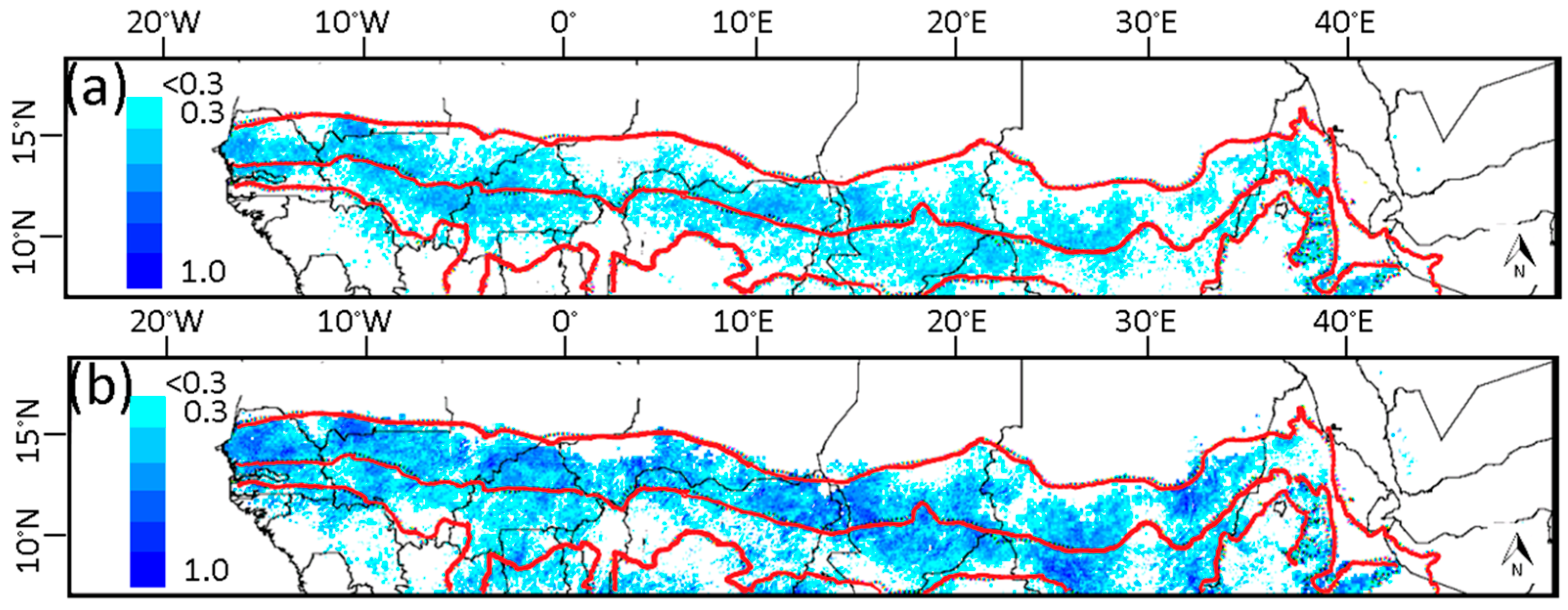
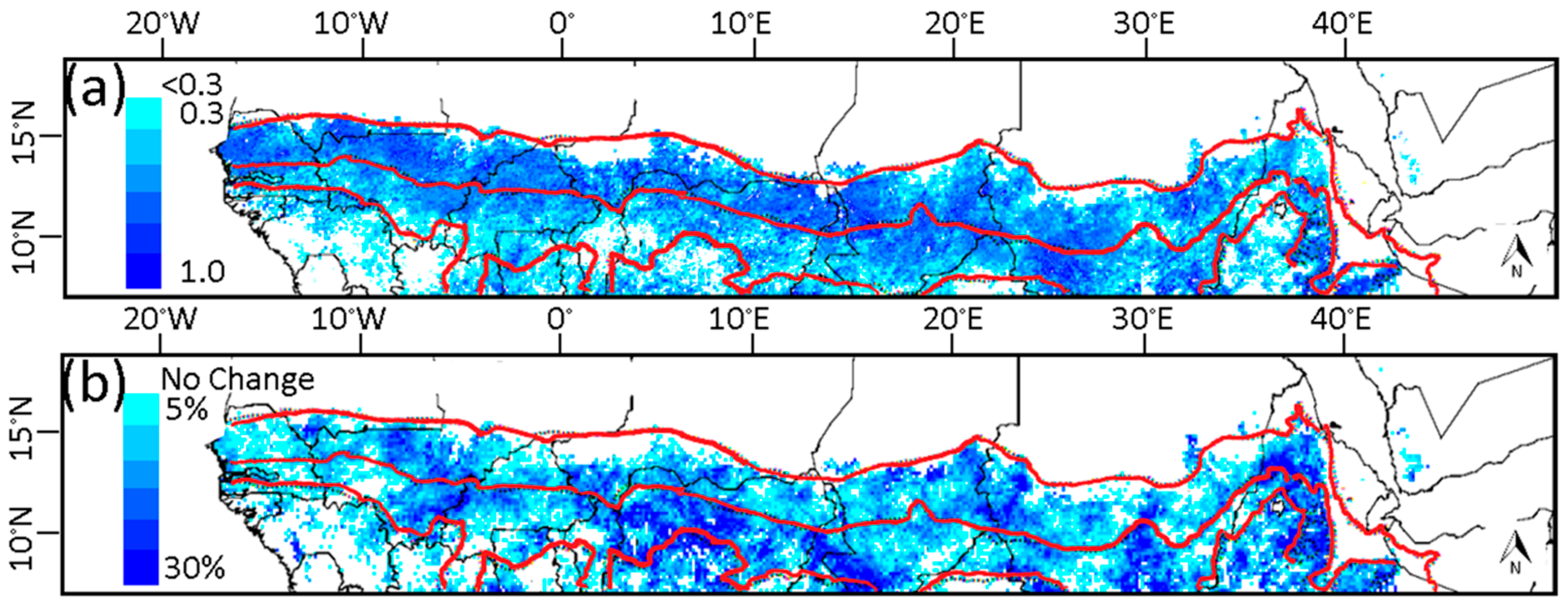
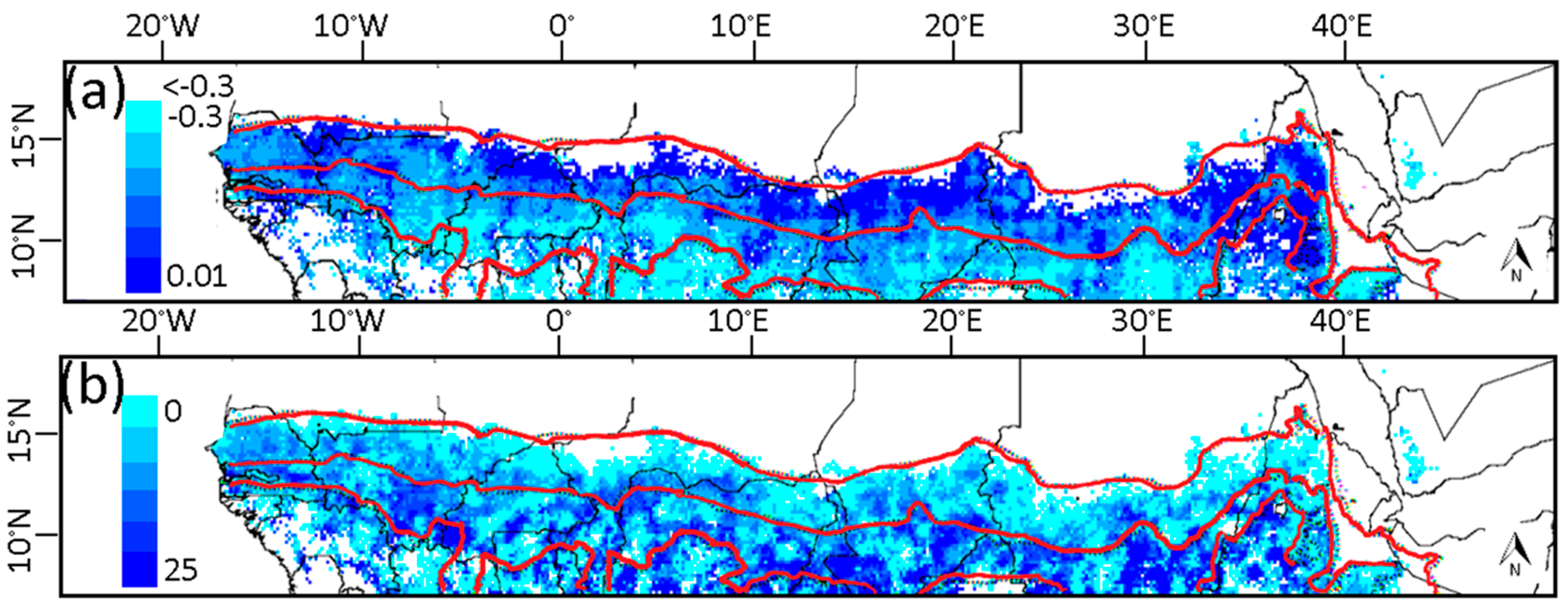
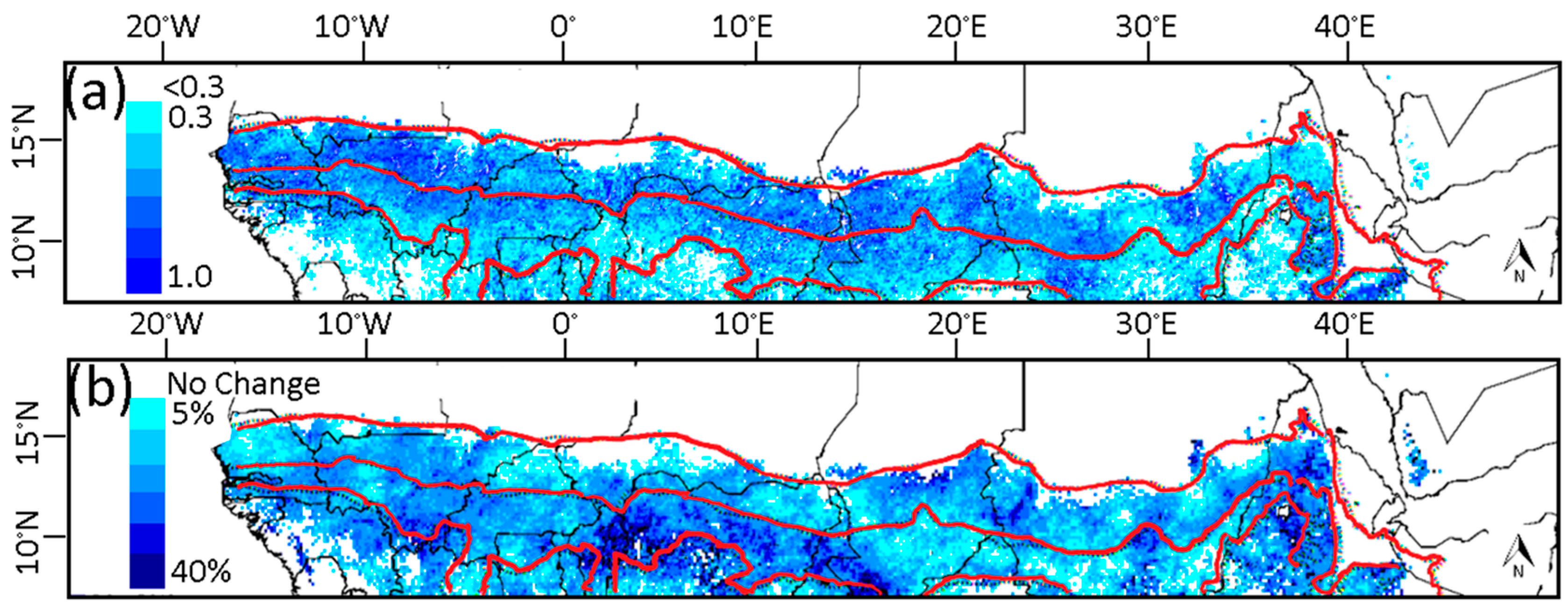
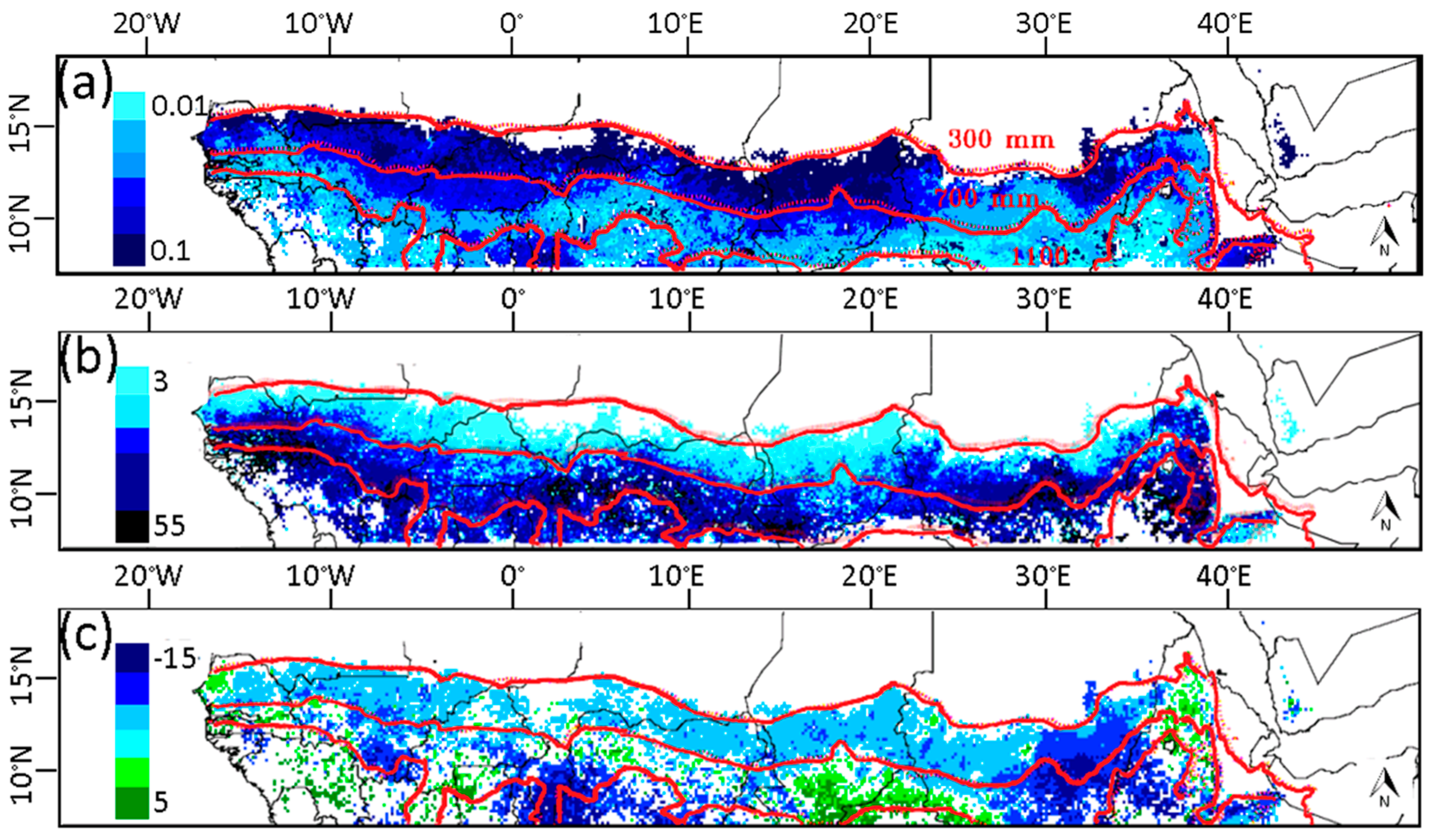
3.1. Phenological Transition Dates
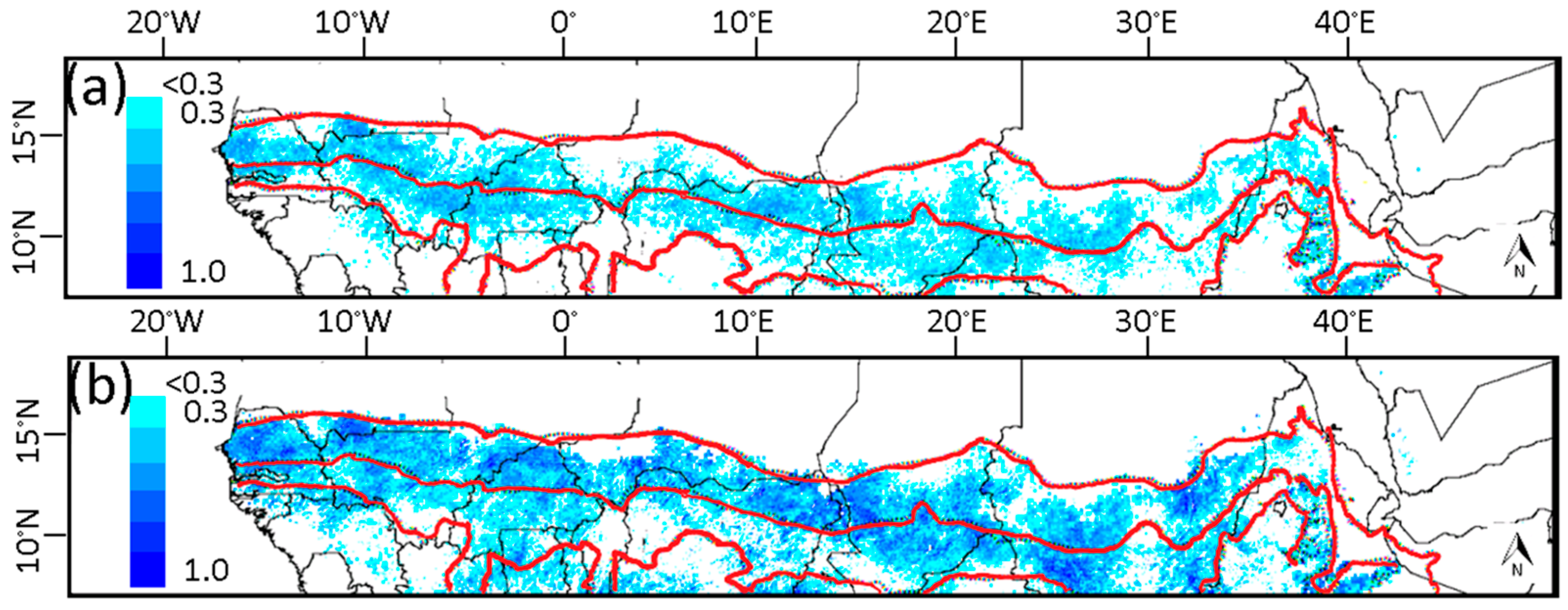
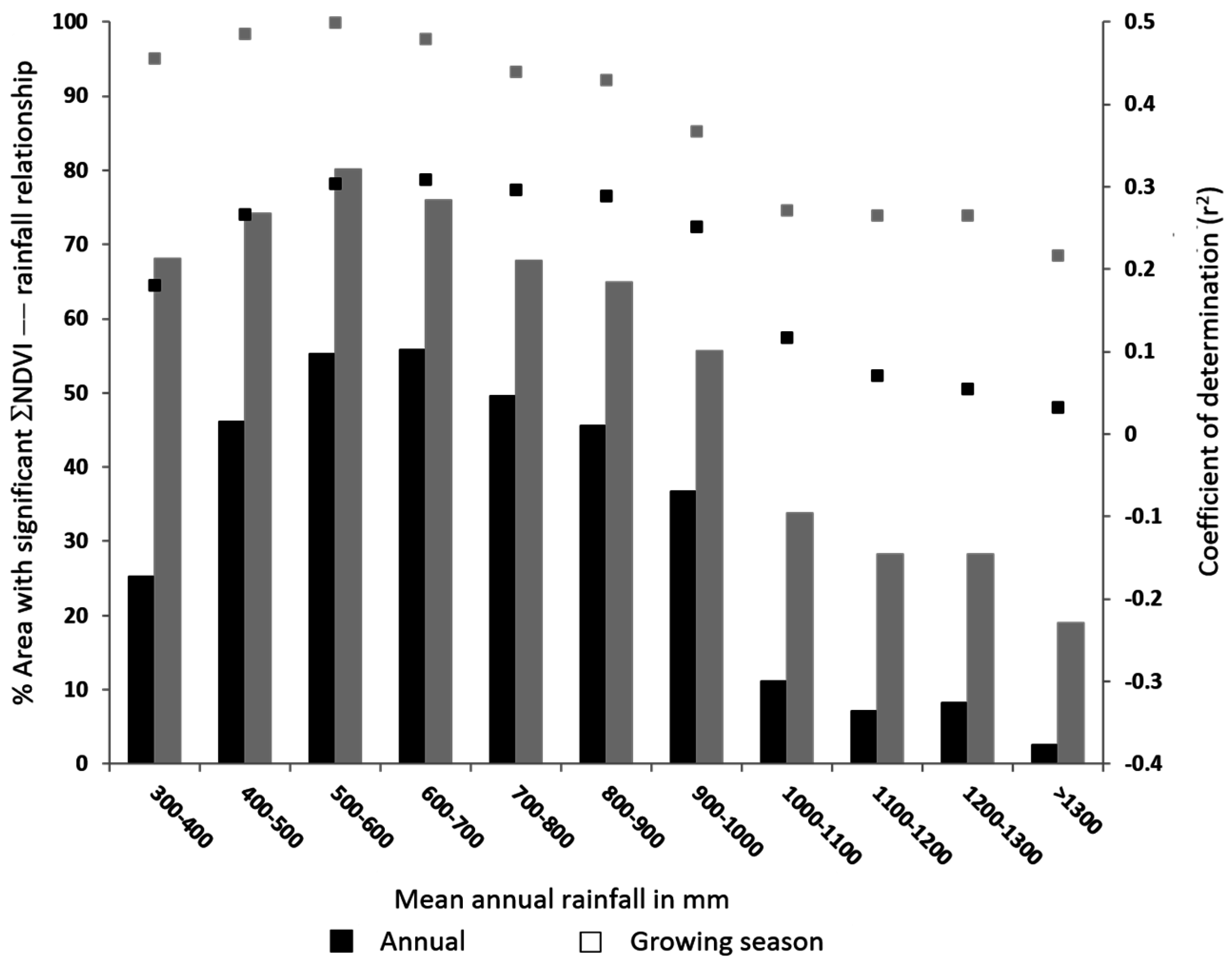
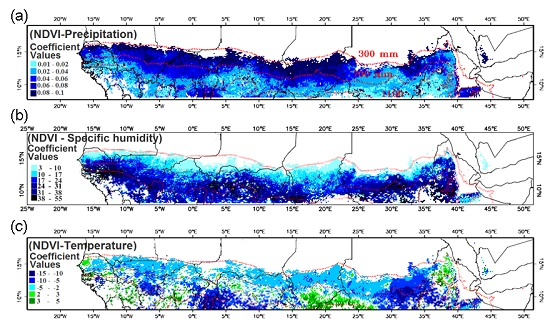
3.2. Relationship of NDVI with Rainfall
3.3. Relationship of Growing Season ΣNDVI with Seasonal Rainfall Distribution
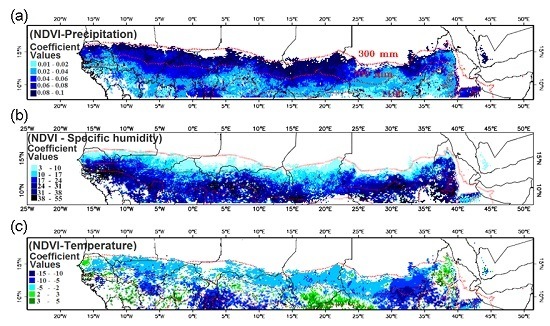
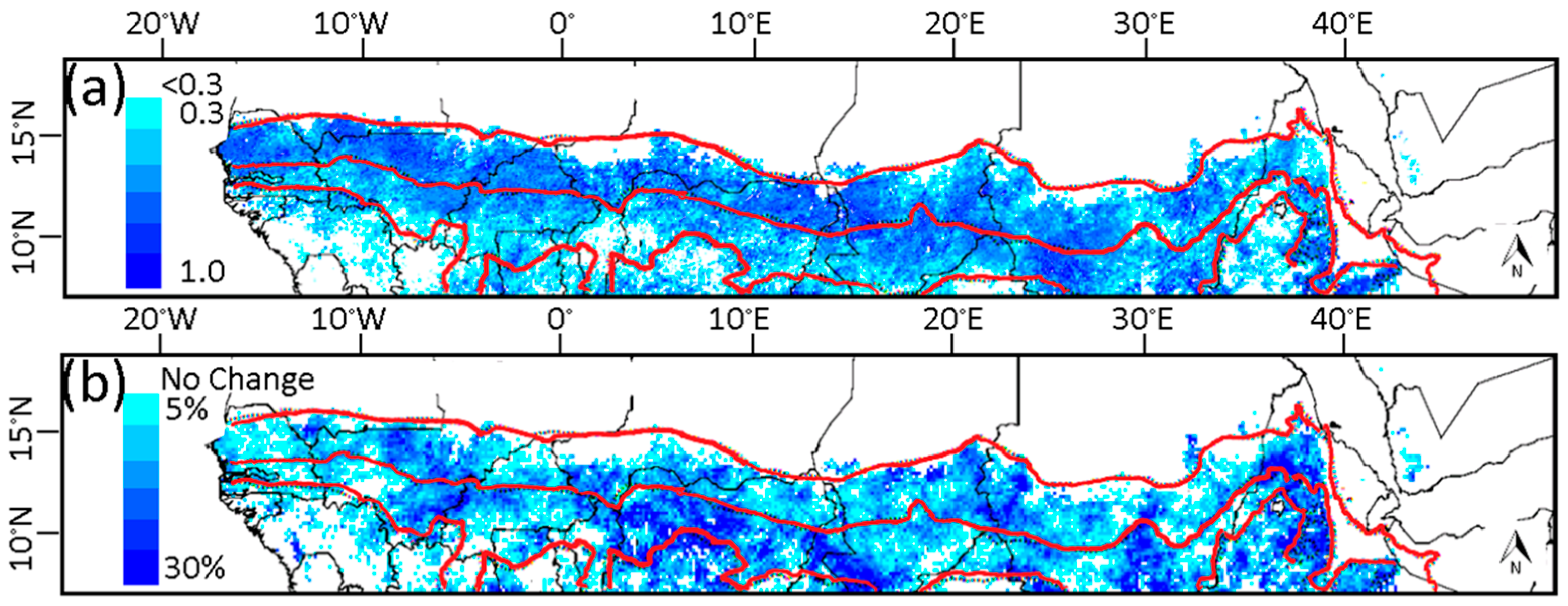
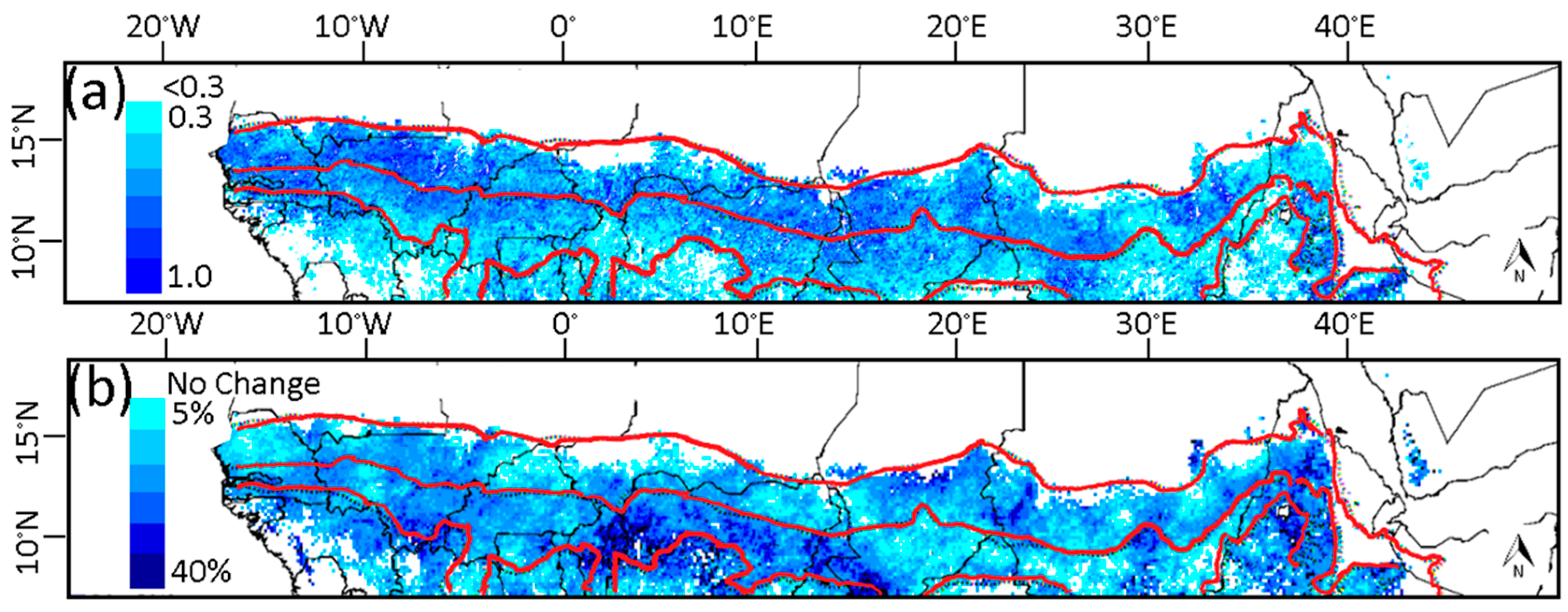
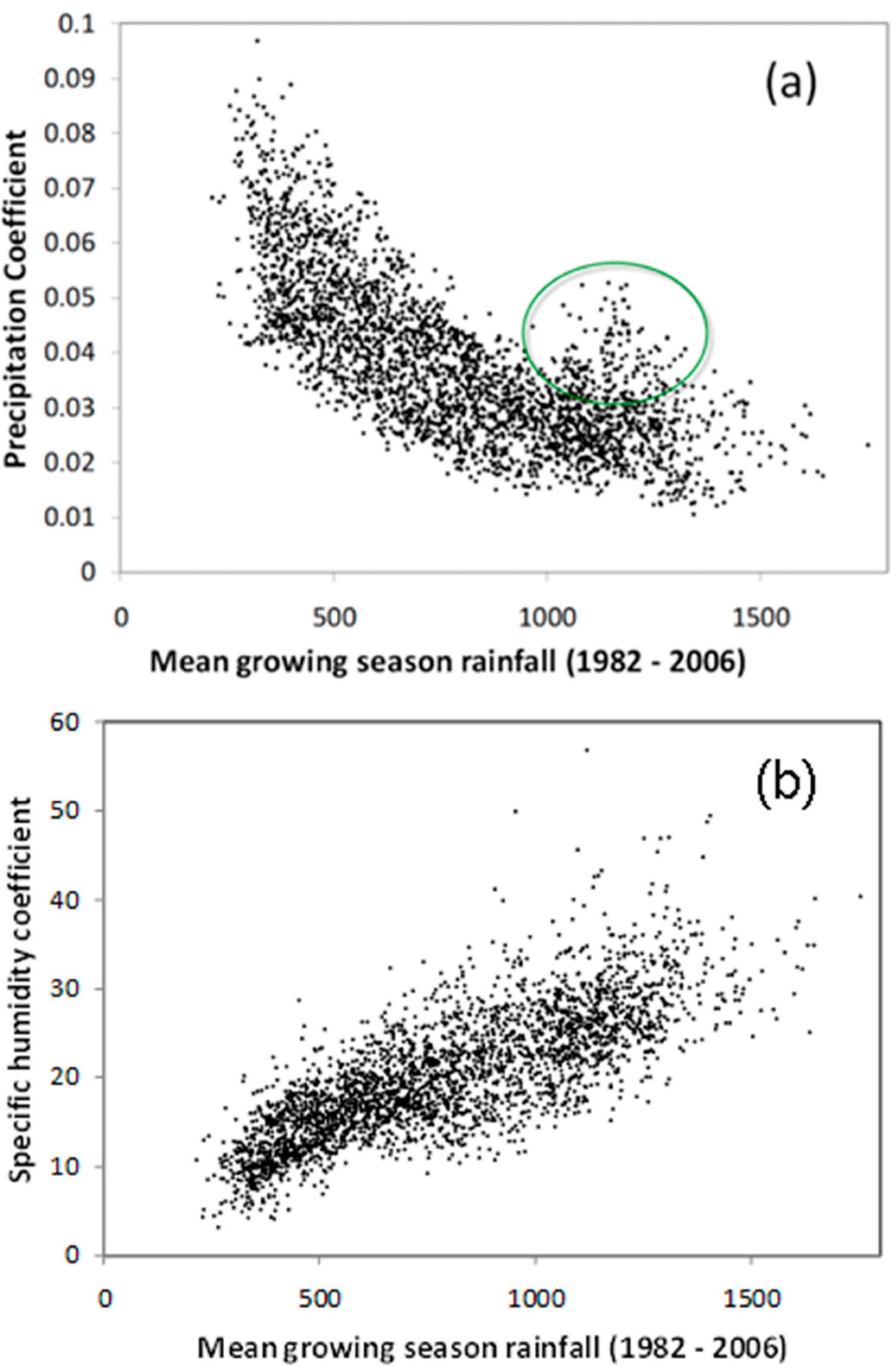
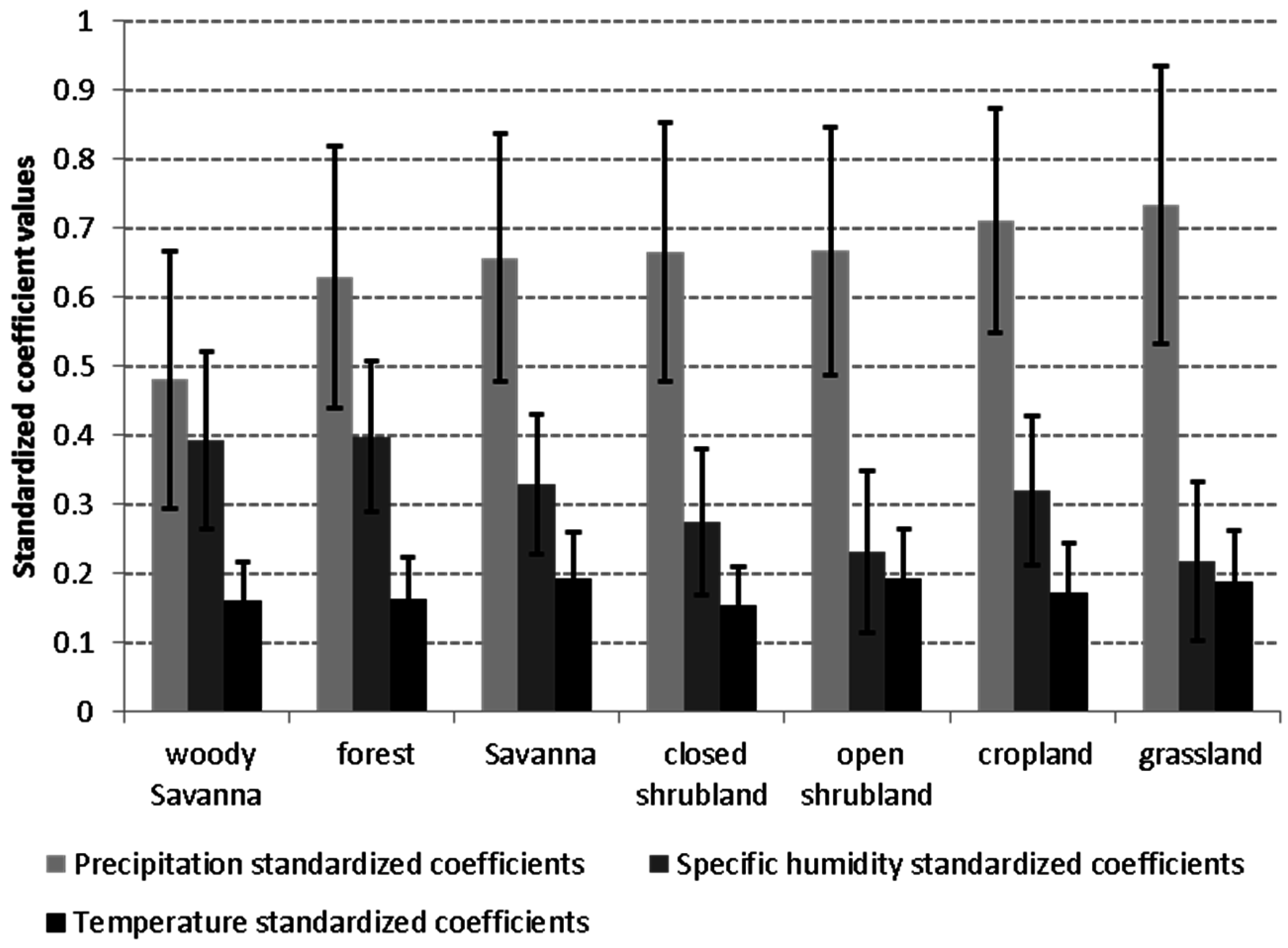
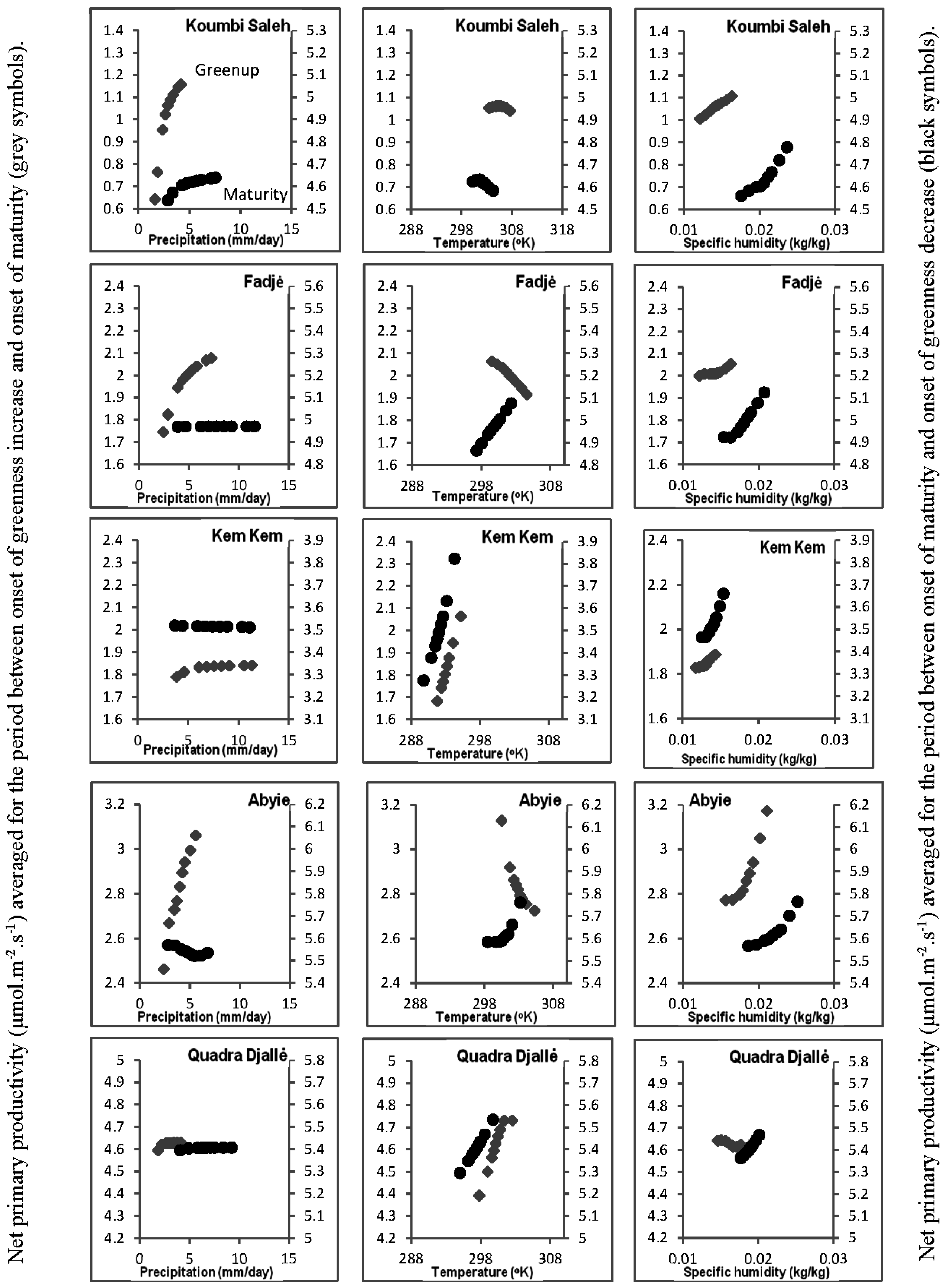
3.4. Relationship of Growing Season ΣNDVI with Air Humidity and Temperature
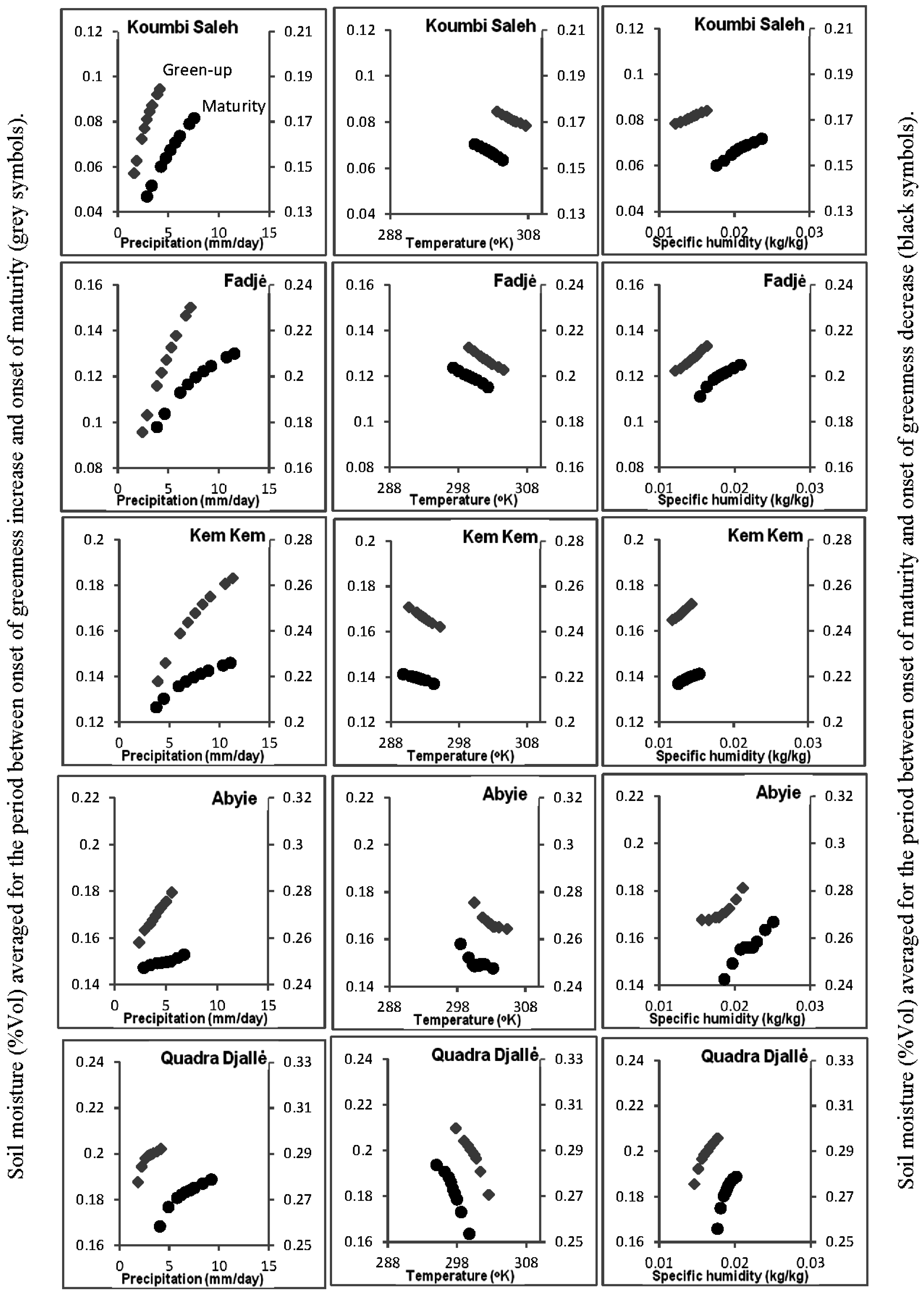
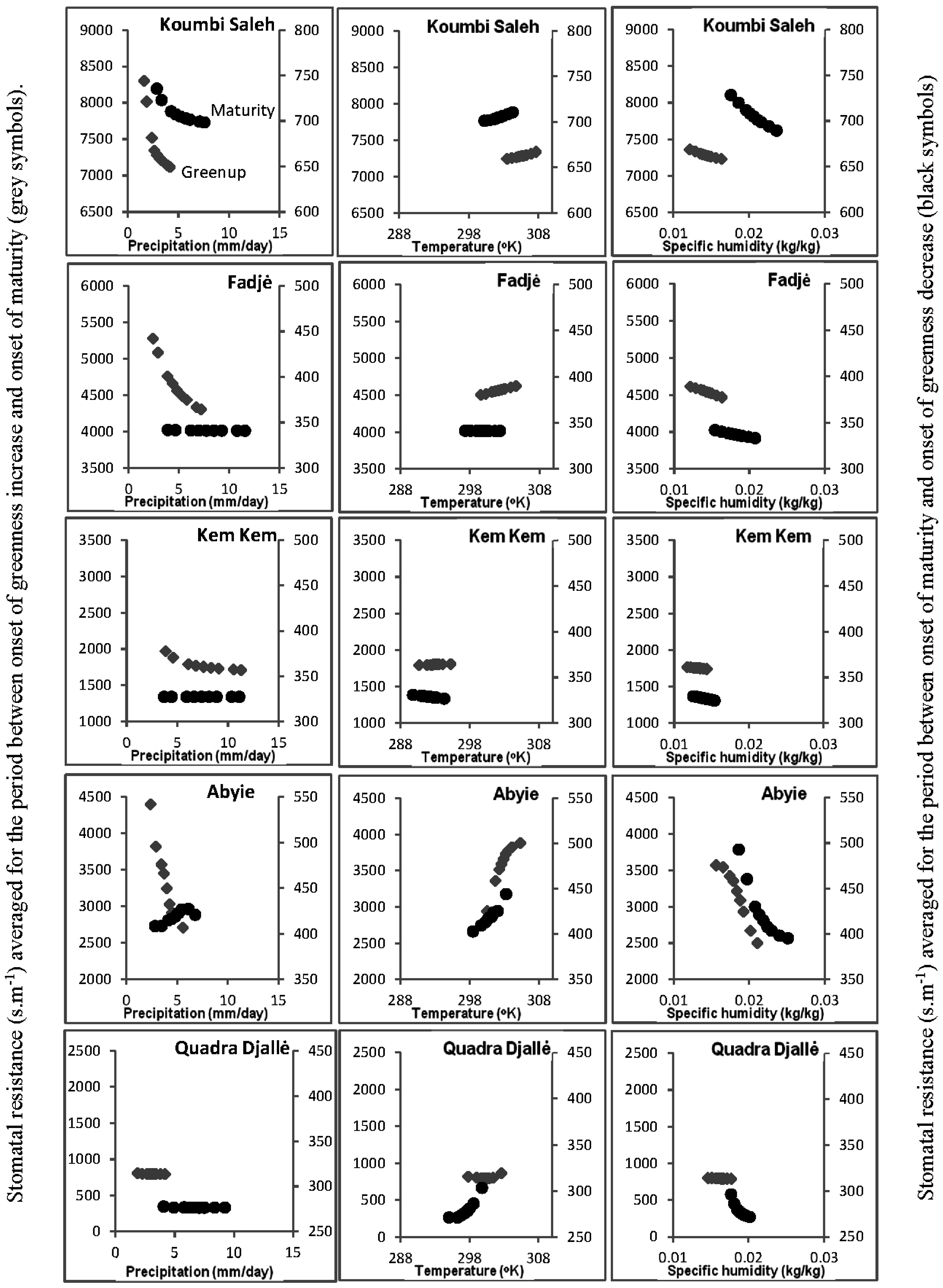
3.5. Soil-Vegetation-Atmosphere Transfer Modeling
4. Discussion
4.1. Relationship of ΣNDVI and Climate Variability
4.2. Phenological Transition Dates
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Fuller, D.O.; Prince, S.D. Rainfall and foliar dynamics in tropical southern Africa: Potential impacts of global climatic change on savanna vegetation. Clim. Chang. 1996, 33, 69–96. [Google Scholar] [CrossRef]
- Olsson, L.; Eklundh, L.; Ardo, J. A recent greening of the Sahel—Trends, patterns and potential causes. J. Arid Environ. 2005, 63, 556–566. [Google Scholar] [CrossRef]
- Hiernaux, P.; Mougin, E.; Diarra, L.; Soumaguel, N.; Lavenu, F.; Tracol, Y.; Diawara, M. Sahelian rangeland response to changes in rainfall over two decades in the Gourma region, Mali. J. Hydrol. 2009, 375, 114–127. [Google Scholar] [CrossRef]
- Goward, S.N.; Prince, S.D. Transient effects of climate on vegetation dynamics: Satellite observations. J. Biogeogr. 1995, 22, 549–564. [Google Scholar] [CrossRef]
- Fang, J.Y.; Piao, S.L.; Tang, Z.Y.; Peng, C.H.; Wei, J. Interannual variability in net primary production and precipitation. Science 2001, 293, U1–U2. [Google Scholar] [CrossRef] [PubMed]
- Nemani, R.R.; Keeling, C.D.; Hashimoto, H.; Jolly, W.M.; Piper, S.C.; Tucker, C.J.; Myneni, R.B.; Running, S.W. Climate-driven increases in global terrestrial net primary production from 1982 to 1999. Science 2003, 300, 1560–1563. [Google Scholar] [CrossRef] [PubMed]
- Easterling, D.R.; Meehl, G.A.; Parmesan, C.; Changnon, S.A.; Karl, T.R.; Mearns, L.O. Climate extremes: Observations, modeling, and impacts. Science 2000, 289, 2068–2074. [Google Scholar] [CrossRef] [PubMed]
- Solomon, S.; Qin, D.; Manning, M.; Chen, Z.; Marquis, M.; Averyt, K.B.; Tignor, M.; Miller, H.L. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change, 2007; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Nicholson, S.E. Climatic and environmental change in Africa during the last two centuries. Clim. Res. 2001, 17, 123–144. [Google Scholar] [CrossRef]
- Le Barbe, L.; Lebel, T.; Tapsoba, D. Rainfall variability in west Africa during the years 1950–90. J. Clim. 2002, 15, 187–202. [Google Scholar] [CrossRef]
- Hiernaux, P.; Diarra, L.; Trichon, V.; Mougin, E.; Soumaguel, N.; Baup, F. Woody plant population dynamics in response to climate changes from 1984 to 2006 in Sahel (Gourma, Mali). J. Hydrol. 2009, 375, 103–113. [Google Scholar] [CrossRef] [Green Version]
- Hulme, M. Climatic perspectives on sahelian desiccation: 1973–1998. Glob. Environ. Chang. Hum. Policy Dimens. 2001, 11, 19–29. [Google Scholar] [CrossRef]
- White, F. Vegetation of Africa—A Description Memoir to Accompany the UNESCO/AETFAT/UNSO Vegetation Map of Africa; Unesco: Paris, France, 1983; p. 356. [Google Scholar]
- Lehouerou, H.N. The rangelands of the Sahel. J. Range Manag. 1980, 33, 41–46. [Google Scholar]
- Lebel, T.; Diedhiou, A.; Laurent, H. Seasonal cycle and interannual variability of the sahelian rainfall at hydrological scales. J. Geophys. Res. Atmos. 2003. [Google Scholar] [CrossRef]
- Dieng, O.; Roucou, P.; Louvet, S. Intra-seasonal variability of precipitation in Senegal (1951–1996). Science et Changements Planétaires/Sécheresse 2008, 19, 87–93. [Google Scholar]
- Nicholson, S.E. The West African Sahel: A review of recent studies on the rainfall regime and its interannual variability. ISRN Meteorol. 2013. [Google Scholar] [CrossRef]
- Sultan, B.; Janicot, S. The west African monsoon dynamics. Part II: The “preonset” and “onset” of the summer monsoon. J. Clim. 2003, 16, 3407–3427. [Google Scholar] [CrossRef]
- Dalu, G.A.; Gaetani, M.; Baldi, M. A hydrological onset and withdrawal index for the west African monsoon. Theor. Appl. Clim. 2009, 96, 179–189. [Google Scholar] [CrossRef]
- Sultan, B.; Janicot, S. Abrupt shift of the ITCZ over west Africa and intra-seasonal variability. Geophys. Res. Lett. 2000, 27, 3353–3356. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Friedl, M.A.; Schaaf, C.B.; Strahler, A.H.; Liu, Z. Monitoring the response of vegetation phenology to precipitation in Africa by coupling MODIS and TRMM instruments. J. Geophys. Res. Atmos. 2005. [Google Scholar] [CrossRef]
- Herrmann, S.M.; Anyamba, A.; Tucker, C.J. Recent trends in vegetation dynamics in the African Sahel and their relationship to climate. Glob. Environ. Chang. 2005, 15, 394–404. [Google Scholar] [CrossRef]
- Lehouerou, H.N.; Bingham, R.L.; Skerbek, W. Relationship between the variability of primary production and the variability of annual precipitation in world arid lands. J. Arid Environ. 1988, 15, 1–18. [Google Scholar]
- Wessels, K.J.; Prince, S.D.; Malherbe, J.; Small, J.; Frost, P.E.; VanZyl, D. Can human-induced land degradation be distinguished from the effects of rainfall variability? A case study in South Africa. J. Arid Environ. 2007, 68, 271–297. [Google Scholar] [CrossRef]
- Nicholson, S.E.; Davenport, M.L.; Malo, A.R. A comparison of the vegetation response to rainfall in the Sahel and East-Africa, using normalized difference vegetation index from NOAA AVHRR. Clim. Chang. 1990, 17, 209–241. [Google Scholar] [CrossRef]
- Tracol, Y.; Mougin, E.; Hiernaux, P.; Jarlan, L. Testing a sahelian grassland functioning model against herbage mass measurements. Ecolo. Model. 2006, 193, 437–446. [Google Scholar] [CrossRef]
- Williams, C.A.; Hanan, N.P.; Baker, I.; Collatz, G.J.; Berry, J.; Denning, A.S. Interannual variability of photosynthesis across africa and its attribution. J. Geophys. Res. Biogeosci. 2008. [Google Scholar] [CrossRef]
- Fensholt, R.; Rasmussen, K. Analysis of trends in the sahelian ‘rain-use efficiency’ using GIMMS NDVI, RFE and GPCP rainfall data. Remote Sens. Environ. 2011, 115, 438–451. [Google Scholar] [CrossRef]
- Knapp, A.K.; Smith, M.D. Variation among biomes in temporal dynamics of aboveground primary production. Science 2001, 291, 481–484. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, J.F.; Kemp, P.R.; Ogle, K.; Fernandez, R.J. Modifying the ‘pulse-reserve’ paradigm for deserts of North America: Precipitation pulses, soil water, and plant responses. Oecologia 2004, 141, 194–210. [Google Scholar] [CrossRef] [PubMed]
- Potts, D.L.; Huxman, T.E.; Cable, J.M.; English, N.B.; Ignace, D.D.; Eilts, J.A.; Mason, M.J.; Weltzin, J.F.; Williams, D.G. Antecedent moisture and seasonal precipitation influence the response of canopy-scale carbon and water exchange to rainfall pulses in a semi-arid grassland. New Phytol. 2006, 170, 849–860. [Google Scholar] [CrossRef] [PubMed]
- Prince, S.D.; Goward, S.N.; Goetz, S.; Czajkowski, K. Inter-annual atmosphere-biosphere variation: Implications for observations and modeling. J. Geophys. Res. 2000, 105, 20–55. [Google Scholar] [CrossRef]
- Wiegand, T.; Snyman, H.A.; Kellner, K.; Paruelo, J.M. Do grasslands have a memory: Modeling phytomass production of a semiarid South African grassland. Ecosystems 2004, 7, 243–258. [Google Scholar] [CrossRef]
- Lauenroth, W.K.; Sala, O.E. Long-term forage production of north-american shortgrass steppe. Ecol. Appl. 1992, 2, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Nouvellon, Y.; Moran, M.S.; Lo Seen, D.; Bryant, R.; Rambal, S.; Ni, W.M.; Begue, A.; Chehbouni, A.; Emmerich, W.E.; Heilman, P.; et al. Coupling a grassland ecosystem model with Landsat imagery for a 10-year simulation of carbon and water budgets. Remote Sens. Environ. 2001, 78, 131–149. [Google Scholar] [CrossRef]
- Oesterheld, M.; Loreti, J.; Semmartin, M.; Sala, O.E. Inter-annual variation in primary production of a semi-arid grassland related to previous-year production. J. Veg. Sci. 2001, 12, 137–142. [Google Scholar] [CrossRef]
- Grist, J.; Nicholson, S.E.; Mpolokang, A. On the use of ndvi for estimating rainfall fields in the Kalahari of Botswana. J. Arid Environ. 1997, 35, 195–214. [Google Scholar] [CrossRef]
- Knapp, A.K.; Beier, C.; Briske, D.D.; Classen, A.T.; Luo, Y.; Reichstein, M.; Smith, M.D.; Smith, S.D.; Bell, J.E.; Fay, P.A.; et al. Consequences of more extreme precipitation regimes for terrestrial ecosystems. Bioscience 2008, 58, 811–821. [Google Scholar] [CrossRef]
- Huxman, T.E.; Smith, M.D.; Fay, P.A.; Knapp, A.K.; Shaw, M.R.; Loik, M.E.; Smith, S.D.; Tissue, D.T.; Zak, J.C.; Weltzin, J.F.; et al. Convergence across biomes to a common rain-use efficiency. Nature 2004, 429, 651–654. [Google Scholar] [CrossRef] [PubMed]
- Elberse, W.T.; Breman, H. Germination and establishment of sahelian rangeland species. I. Seed properties. Oecologia 1989, 80, 477–484. [Google Scholar] [CrossRef]
- Elberse, W.T.; Breman, H. Germination and establishment of sahelian rangeland species. II. Effects of water availability. Oecologia 1990, 85, 32–40. [Google Scholar] [CrossRef]
- Huxman, T.E.; Snyder, K.A.; Tissue, D.; Leffler, A.J.; Ogle, K.; Pockman, W.T.; Sandquist, D.R.; Potts, D.L.; Schwinning, S. Precipitation pulses and carbon fluxes in semiarid and arid ecosystems. Oecologia 2004, 141, 254–268. [Google Scholar] [CrossRef] [PubMed]
- Chabot, B.F.; Hicks, D.J. The ecology of leaf life spans. Annu. Rev. Ecol. Syst. 1982, 13, 229–259. [Google Scholar] [CrossRef]
- Noy-Meir, I. Desert ecosystems: Environment and producers. Annu. Rev. Ecol. Syst. 1973, 4, 25–51. [Google Scholar] [CrossRef]
- Sala, O.; Lauenroth, W. Small rainfall events: An ecological role in semiarid regions. Oecologia 1982, 53, 301–304. [Google Scholar] [CrossRef]
- Prince, S.D.; De Colstoun, E.B.; Kravitz, L.L. Evidence from rain-use efficiencies does not indicate extensive Sahelian desertification. Glob. Chang. Biol. 1998, 4, 359–374. [Google Scholar] [CrossRef]
- Wainwright, J.; Mulligan, M.; Thornes, J. Plants and water in drylands. In Eco-Hydrology: Plants and Water in Terrestrial and Aquatic Environments; Baird, A.J., Wilby, R.L., Eds.; Routledge: London, UK, 1999; pp. 78–126. [Google Scholar]
- Jobbagy, E.G.; Sala, O.E. Controls of grass and shrub aboveground production in the Patagonian Steppe. Ecol. Appl. 2000, 10, 541–549. [Google Scholar] [CrossRef]
- Paruelo, J.M.; Sala, O.E.; Beltran, A.B. Long-term dynamics of water and carbon in semi-arid ecosystems: A gradient analysis in the Patagonian Steppe. Plant Ecol. 2000, 150, 133–143. [Google Scholar] [CrossRef]
- Heisler-White, J.L.; Blair, J.M.; Kelly, E.F.; Harmoney, K.; Knapp, A.K. Contingent productivity responses to more extreme rainfall regimes across a grassland biome. Glob. Chang. Biol. 2009, 15, 2894–2904. [Google Scholar] [CrossRef]
- Knapp, A.K.; Fay, P.A.; Blair, J.M.; Collins, S.L.; Smith, M.D.; Carlisle, J.D.; Harper, C.W.; Danner, B.T.; Lett, M.S.; McCarron, J.K. Rainfall variability, carbon cycling, and plant species diversity in a mesic grassland. Science 2002, 298, 2202–2205. [Google Scholar] [CrossRef] [PubMed]
- Schwinning, S.; Sala, O.E. Hierarchy of responses to resource pulses in and and semi-arid ecosystems. Oecologia 2004, 141, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Robertson, T.R.; Bell, C.W.; Zak, J.C.; Tissue, D.T. Precipitation timing and magnitude differentially affect aboveground annual net primary productivity in three perennial species in a Chihuahuan desert grassland. New Phytol. 2009, 181, 230–242. [Google Scholar] [CrossRef] [PubMed]
- Good, S.P.; Caylor, K.K. Climatological determinants of woody cover in Africa. Proc. Natl. Acad. Sci. USA 2011, 108, 4902–4907. [Google Scholar] [CrossRef] [PubMed]
- Belnap, J.; Welter, J.R.; Grimm, N.B.; Barger, N.; Ludwig, J.A. Linkages between microbial and hydrologic processes in arid and semiarid watersheds. Ecology 2005, 86, 298–307. [Google Scholar] [CrossRef]
- Collatz, G.J.; Ball, J.T.; Grivet, C.; Berry, J.A. Physiological and environmental-regulation of stomatal conductance, photosynthesis and transpiration—A model that includes a laminar boundary-layer. Agric. For. Meteorol. 1991, 54, 107–136. [Google Scholar] [CrossRef]
- Collatz, G.J.; Ribas-Carbo, M.; Berry, J.A. Coupled photosynthesis-stomatal conductance model for leaves of C4 plants. Aust. J. Plant Physiol. 1992, 19, 519–538. [Google Scholar] [CrossRef]
- Reichstein, M.; Ciais, P.; Papale, D.; Valentini, R.; Running, S.; Viovy, N.; Cramer, W.; Granier, A.; OgÉE, J.; Allard, V.; et al. Reduction of ecosystem productivity and respiration during the european summer 2003 climate anomaly: A joint flux tower, remote sensing and modelling analysis. Glob. Chang. Biol. 2007, 13, 634–651. [Google Scholar] [CrossRef]
- Xue, Y.; Sellers, P.J.; Kinter, J.L.; Shukla, J. A simplified biosphere model for global climate studies. J. Clim. 1991, 4, 345–364. [Google Scholar] [CrossRef]
- Merbold, L.; Ardo, J.; Arneth, A.; Scholes, R.J.; Nouvellon, Y.; de Grandcourt, A.; Archibald, S.; Bonnefond, J.M.; Boulain, N.; Brueggemann, N.; et al. Precipitation as driver of carbon fluxes in 11 African ecosystems. Biogeosciences 2009, 6, 1027–1041. [Google Scholar] [CrossRef]
- Beer, C.; Reichstein, M.; Tomelleri, E.; Ciais, P.; Jung, M.; Carvalhais, N.; Roedenbeck, C.; Arain, M.A.; Baldocchi, D.; Bonan, G.B.; et al. Terrestrial gross carbon dioxide uptake: Global distribution and covariation with climate. Science 2010, 329, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, S.E.; Tucker, C.J.; Ba, M.B. Desertification, drought, and surface vegetation: An example from the West African Sahel. Bull. Am. Meteorol. Soc. 1998, 79, 815–829. [Google Scholar] [CrossRef]
- Camberlin, P.; Martiny, N.; Philippon, N.; Richard, Y. Determinants of the interannual relationships between remote sensed photosynthetic activity and rainfall in tropical Africa. Remote Sens. Environ. 2007, 106, 199–216. [Google Scholar] [CrossRef]
- Hellden, U.; Tottrup, C. Regional desertification: A global synthesis. Glob. Planet. Change 2008, 64, 169–176. [Google Scholar] [CrossRef]
- Sheffield, J.; Goteti, G.; Wood, E.F. Development of a 50-year high-resolution global dataset of meteorological forcings for land surface modeling. J. Clim. 2006, 19, 3088–3111. [Google Scholar] [CrossRef]
- Pedelty, J.; Devadiga, S.; Masuoka, E.; Brown, M.; Roy, D.; Pinheiro, A. Generating a long-term land data record from the AVHRR and MODIS instruments. In Proceedings of the 2007 IEEE International Geoscience and Remote Sensing Symposium, Barcelona, Spain, 23–28 July 2007.
- Vermote, E.; Kaufman, Y.J. Absolute calibration of avhrr visible and near-infrared channels using ocean and cloud views. Int. J. Remote Sens. 1995, 16, 2317–2340. [Google Scholar] [CrossRef]
- Vermote, E.F.; Saleous, N.Z. Calibration of noaa16 AVHRR over a desert site using MODIS data. Remote Sens. Environ. 2006, 105, 214–220. [Google Scholar] [CrossRef]
- Nagol, J.N. Quantification of Error in AVHRR NDVI Data. Ph.D. Thesis, University of Maryland, College Park, MD, USA, 2011. [Google Scholar]
- Nagol, J.N. Quantification of uncertainty in LTDR AVHRR NDVI data by comparison with MODIS Aqua. Remote Sens. Environ. 2016. submitted for publication. [Google Scholar]
- Rishmawi, K.N. Spatial Patterns and Potential Mechanisms of Land Degradation in the Sahel. Ph.D. Thesis, University of Maryland, College Park, MD, USA, 2013. [Google Scholar]
- Vermote, E.; Justice, C.O.; Breon, F.M. Towards a generalized approach for correction of the BRDF effect in MODIS directional reflectances. IEEE Trans. Geosci. Remote Sens. 2009, 47, 898–908. [Google Scholar] [CrossRef]
- Fensholt, R.; Sandholt, I.; Proud, S.R.; Stisen, S.; Rasmussen, M.O. Assessment of MODIS sun-sensor geometry variations effect on observed NDVI using MSG SEVIRI geostationary data. Int. J. Remote Sens. 2010, 31, 6163–6187. [Google Scholar] [CrossRef]
- Gutman, G.G. Vegetation indexes from AVHRR—An update and future-prospects. Remote Sens. Environ. 1991, 35, 121–136. [Google Scholar] [CrossRef]
- Cihlar, J.; Howarth, J. Detection and removal of cloud contamination from AVHRR images. IEEE Trans. Geosci. Remote Sens. 1994, 32, 583–589. [Google Scholar] [CrossRef]
- Viovy, N.; Arino, O.; Belward, A.S. The best index slope extraction (BISE)—A method for reducing noise in NDVI time-series. Int. J. Remote Sens. 1992, 13, 1585–1590. [Google Scholar] [CrossRef]
- Hansen, M.C.; DeFries, R.S.; Townshend, J.R.G.; Carroll, M.; Dimiceli, C.; Sohlberg, R.A. Global percent tree cover at a spatial resolution of 500 meters: First results of the modis vegetation continuous fields algorithm. Earth Interact. 2003. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Friedl, M.A.; Schaaf, C.B.; Strahler, A.H.; Hodges, J.C.F.; Gao, F.; Reed, B.C.; Huete, A. Monitoring vegetation phenology using MODIS. Remote Sens. Environ. 2003, 84, 471–475. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Friedl, M.A.; Schaaf, C.B. Global vegetation phenology from Moderate Resolution Imaging spectroradiometer (MODIS): Evaluation of global patterns and comparison with in situ measurements. J. Geophys. Res. Biogeosci. 2006. [Google Scholar] [CrossRef]
- Dielman, T.E. Applied Regression Analysis: A Second Course in Business and Economic Statistics, 4th ed.; Brooks/Cole: Belmont, CA, USA, 2005. [Google Scholar]
- Furnival, G.M.; Wilson, R.W. Regressions by leaps and bounds. Technometrics 1974, 16, 499–511. [Google Scholar] [CrossRef]
- Freund, R.; Wilson, W.J. Regression Analysis: Statistical Modeling of a Response Variable; Academic Press: San Diego, CA, USA, 1998. [Google Scholar]
- Friedl, M.A.; McIver, D.K.; Hodges, J.C.F.; Zhang, X.Y.; Muchoney, D.; Strahler, A.H.; Woodcock, C.E.; Gopal, S.; Schneider, A.; Cooper, A.; et al. Global land cover mapping from MODIS: Algorithms and early results. Remote Sens. Environ. 2002, 83, 287–302. [Google Scholar] [CrossRef]
- Zhan, X.W.; Xue, Y.K.; Collatz, G.J. An analytical approach for estimating CO2 and heat fluxes over the amazonian region. Ecolo. Model. 2003, 162, 97–117. [Google Scholar] [CrossRef]
- Wood, E.F.; Lettenmaier, D.P.; Liang, X.; Lohmann, D.; Boone, A.; Chang, S.; Chen, F.; Dai, Y.J.; Dickinson, R.E.; Duan, Q.Y.; et al. The project for intercomparison of land-surface parameterization schemes (PILPS) phase 2(c) red-arkansas river basin experiment: I. Experiment description and summary intercomparisons. Glob. Planet. Chang. 1998, 19, 115–135. [Google Scholar] [CrossRef]
- Baret, F.; Hagolle, O.; Geiger, B.; Bicheron, P.; Miras, B.; Huc, M.; Berthelot, B.; Nino, F.; Weiss, M.; Samain, O.; et al. LAI, fAPAR and fCover cyclopes global products derived from vegetation—Part I: Principles of the algorithm. Remote Sens. Environ. 2007, 110, 275–286. [Google Scholar] [CrossRef] [Green Version]
- Sellers, P.J. Canopy reflectance, photosynthesis, and transpiration. II. The role of biophysics in the linearity of their interdependence. Remote Sens. Environ. 1987, 21, 143–183. [Google Scholar] [CrossRef]
- Malo, A.R.; Nicholson, S.E. A Study of Rainfall and Vegetation Dynamics in the African Sahel Using Normalized Vegetation Index; Elsevier: Kidlington, UK, 1990. [Google Scholar]
- Davenport, M.L.; Nicholson, S.E. On the relation between rainfall and the normalized difference vegetation index for diverse vegetation types in East-Africa. Int. J. Remote Sens. 1993, 14, 2369–2389. [Google Scholar] [CrossRef]
- Ramankutty, N.; Evan, A.T.; Monfreda, C.; Foley, J.A. Farming the planet: I. Geographic distribution of global agricultural lands in the year 2000. Glob. Biogeochem. Cycles 2008. [Google Scholar] [CrossRef]
- Gruber, I.; Kloos, J.; Schopp, M. Seasonal water demand in Benin’s agriculture. J. Environ. Manag. 2009, 90, 196–205. [Google Scholar] [CrossRef] [PubMed]
- African Development Bank Group. Oueme Valley Irrigated Agricultural Development Project; Project Performance Evaluation Report; African Development Bank Group: Abidjan, Côte d’Ivoire, 1998. [Google Scholar]
- Hanjra, M.A.; Gichuki, F. Investments in agricultural water management for poverty reduction in Africa: Case studies of Limpopo, Nile, and Volta River Basins. Nat. Resour. Forum 2008, 32, 185–202. [Google Scholar] [CrossRef]
- Bai, Z.G.; Dent, D.L.; Olsson, L.; Schaepman, M.E. Proxy global assessment of land degradation. Soil Use Manag. 2008, 24, 223–234. [Google Scholar] [CrossRef]
- Bui, E.N.; Hancock, G.J.; Wilkinson, S.N. ‘Tolerable’ hillslope soil erosion rates in Australia: Linking science and policy. Agric. Ecosyst. Environ. 2011, 144, 136–149. [Google Scholar] [CrossRef]
- LeHouérou, H.N. The Grazing Land Ecosystems of the African Sahelthe Grazing Land Ecosystems of the African Sahel; Springer: Heidelberg, Germany, 1989. [Google Scholar]
- Lieth, H. Modeling the primary productivity of the world. In Primary Productivity of the Biosphere; Lieth, H., Whittaker, R.H., Eds.; Springer: Heidelberg, Germany, 1975; pp. 237–263. [Google Scholar]
- Breman, H.; Dewit, C.T. Rangeland productivity and exploitation in the Sahel. Science 1983, 221, 1341–1347. [Google Scholar] [CrossRef] [PubMed]
- Hiernaux, P.; Ayantunde, A.; Kalilou, A.; Mougin, E.; Gerard, B.; Baup, F.; Grippa, M.; Djaby, B. Trends in productivity of crops, fallow and rangelands in southwest Niger: Impact of land use, management and variable rainfall. J. Hydrol. 2009, 375, 65–77. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, Y.; Balzter, H.; Kaduk, J.; Tucker, C. Land degradation assessment using residual trend analysis of GIMMS NDVI3g, soil moisture and rainfall in Sub-Saharan West Africa from 1982 to 2012. Remote Sens. 2015, 7, 5471–5494. [Google Scholar] [CrossRef]
- Granger, C.W.J. Spurious regressions in econometrics. J. Econ. 1974, 2, 111–120. [Google Scholar] [CrossRef]
- Yang, L.; Wylie, B.K.; Tieszen, L.L.; Reed, B.C. An analysis of relationships among climate forcing and time-integrated NDVI of grasslands over the U.S. Northern and central great plains. Remote Sens. Environ. 1998, 65, 25–37. [Google Scholar] [CrossRef]
- Wang, G.L.; Eltahir, E.A.B. Modeling the biosphere-atmosphere system: The impact of the subgrid variability in rainfall interception. J. Clim. 2000, 13, 2887–2899. [Google Scholar] [CrossRef]
- Wessels, K.J. Monitoring land Degradation in Southern Africa by Assessing Changes in Primary Productivity. Ph.D. Thesis, University of Maryland, College Park, MD, USA, 2005. [Google Scholar]
- Barron, J.; Rockstrom, J.; Gichuki, F.; Hatibu, N. Dry spell analysis and maize yields for two semi-arid locations in east Africa. Agric. For. Meteorol. 2003, 117, 23–37. [Google Scholar] [CrossRef]
- Frappart, F.; Hiernaux, P.; Guichard, F.; Mougin, E.; Kergoat, L.; Arjounin, M.; Lavenu, F.; Koite, M.; Paturel, J.E.; Lebel, T. Rainfall regime across the Sahel band in the Gourma Region, Mali. J. Hydrol. 2009, 375, 128–142. [Google Scholar] [CrossRef] [Green Version]
- Yengoh, G.T.; Armah, F.A.; Onumah, E.E.; Odoi, J.O. Trends in agriculturally-relevant rainfall characteristics for small-scale agriculture in northern Ghana. J. Agric. Sci. 2010, 2, 3–16. [Google Scholar] [CrossRef]
- Penning de Vries, F.; Djitèye, M.A. La Productivité des Pasturages Sahélien: Une étude des Sols des Végétations et de L’exploitation de Cette Ressource Naturelle; Pudoc, Center for Agricultural Publication and Documentation: Wageningen, The Netherlands, 1983; p. 525. [Google Scholar]
- Huber, S.; Fensholt, R.; Rasmussen, K. Water availability as the driver of vegetation dynamics in the African Sahel from 1982 to 2007. Glob. Planet. Chang. 2011, 76, 186–195. [Google Scholar] [CrossRef]
- Churkina, G.; Running, S.W. Contrasting climatic controls on the estimated productivity of global terrestrial biomes. Ecosystems 1998, 1, 206–215. [Google Scholar] [CrossRef]
- D’Amato, N.; Lebel, T. On the characteristics of the rainfall events in the Sahel with a view to the analysis of climatic variability. Int. J. Clim. 1998, 18, 955–974. [Google Scholar] [CrossRef]
- LeBarbe, L.; Lebel, T. Rainfall climatology of the hapex-Sahel region during the years 1950–1990. J. Hydrol. 1997, 189, 43–73. [Google Scholar]
- Prince, S.D. Where does desertification occur? Mapping dryland degradation at regional to global scales. In The End of Desertification? Disrupting Environmental Change in Drylands; Behnke, R., Matimore, M., Eds.; Springer: Heidelberg, Germany, 2016. [Google Scholar]
- Jackson, H.; Prince, S.D. Degradation of net primary production in a semi-arid rangeland. Biogeosciences 2016, 13, 4721–4734. [Google Scholar] [CrossRef]
- Jackson, H.; Prince, S.D. Degradation of non-photosynthetic vegetation in a semi-arid rangeland. Remote Sens. 2016. [Google Scholar] [CrossRef]
- Dardel, C.; Kergoat, L.; Hiernaux, P.; Grippa, M.; Mougin, E.; Ciais, P.; Nguyen, C.C. Rain-use-efficiency: What it tells us about the conflicting Sahel greening and sahelian paradox. Remote Sens. 2014, 6, 3446–3474. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Land Cover | Latitude | Longitude | Elevation (m) | Growing Season Means and Standard Deviations | Maximum LAI and Interannual Variation | Mean Duration | |||
|---|---|---|---|---|---|---|---|---|---|---|
| Cumulative Precipitation (mm) | Temperature (°C) | Specific Humidity (gH2O/kgdry air) | GUMA (Days) | MASE (Days) | ||||||
| Koumbi Saleh, Mauritania | Grasslands | 15.775°N | 7.991°W | 187 | 300 ± 60 | 30.6 ± 0.71 | 18 ± 1.0 | 1.39 ± 0.37 | 75 ± 23 | 21 ± 3 |
| Fadjė, Chad | Shrublands with grass ground cover | 11.625°N | 15.925°W | 325 | 410 ± 99 | 28.2 ± 0.64 | 16 ± 0.8 | 2.79 ± 0.35 | 66 ± 21 | 38 ± 10 |
| Kem Kem, Ethiopia | Cropland | 12.075°N | 37.825°W | 1870 | 678 ± 103 | 19.7 ± 0.52 | 13 ± 0.5 | 2.51 ± 0.13 | 65 ± 18 | 72 ± 11 |
| Abyie, Sudan | Woody savannas | 9.525°N | 28.425°W | 400 | 792 ± 110 | 28.3 ± 0.70 | 21 ± 1.0 | 2.40 ± 0.20 | 95 ± 15 | 70 ± 12 |
| Quarda Djallė, Central African Republic | Savannas with grass ground cover | 8.775°N | 22.375°W | 682 | 861 ± 108 | 26.5 ± 0.62 | 18 ± 0.6 | 3.25 ± 0.25 | 128 ± 11 | 105 ± 11 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rishmawi, K.; Prince, S.D.; Xue, Y. Vegetation Responses to Climate Variability in the Northern Arid to Sub-Humid Zones of Sub-Saharan Africa. Remote Sens. 2016, 8, 910. https://doi.org/10.3390/rs8110910
Rishmawi K, Prince SD, Xue Y. Vegetation Responses to Climate Variability in the Northern Arid to Sub-Humid Zones of Sub-Saharan Africa. Remote Sensing. 2016; 8(11):910. https://doi.org/10.3390/rs8110910
Chicago/Turabian StyleRishmawi, Khaldoun, Stephen D. Prince, and Yongkang Xue. 2016. "Vegetation Responses to Climate Variability in the Northern Arid to Sub-Humid Zones of Sub-Saharan Africa" Remote Sensing 8, no. 11: 910. https://doi.org/10.3390/rs8110910