Remote Sensing-Based Biomass Estimation and Its Spatio-Temporal Variations in Temperate Grassland, Northern China

Abstract
:1. Introduction
2. Materials and Methods
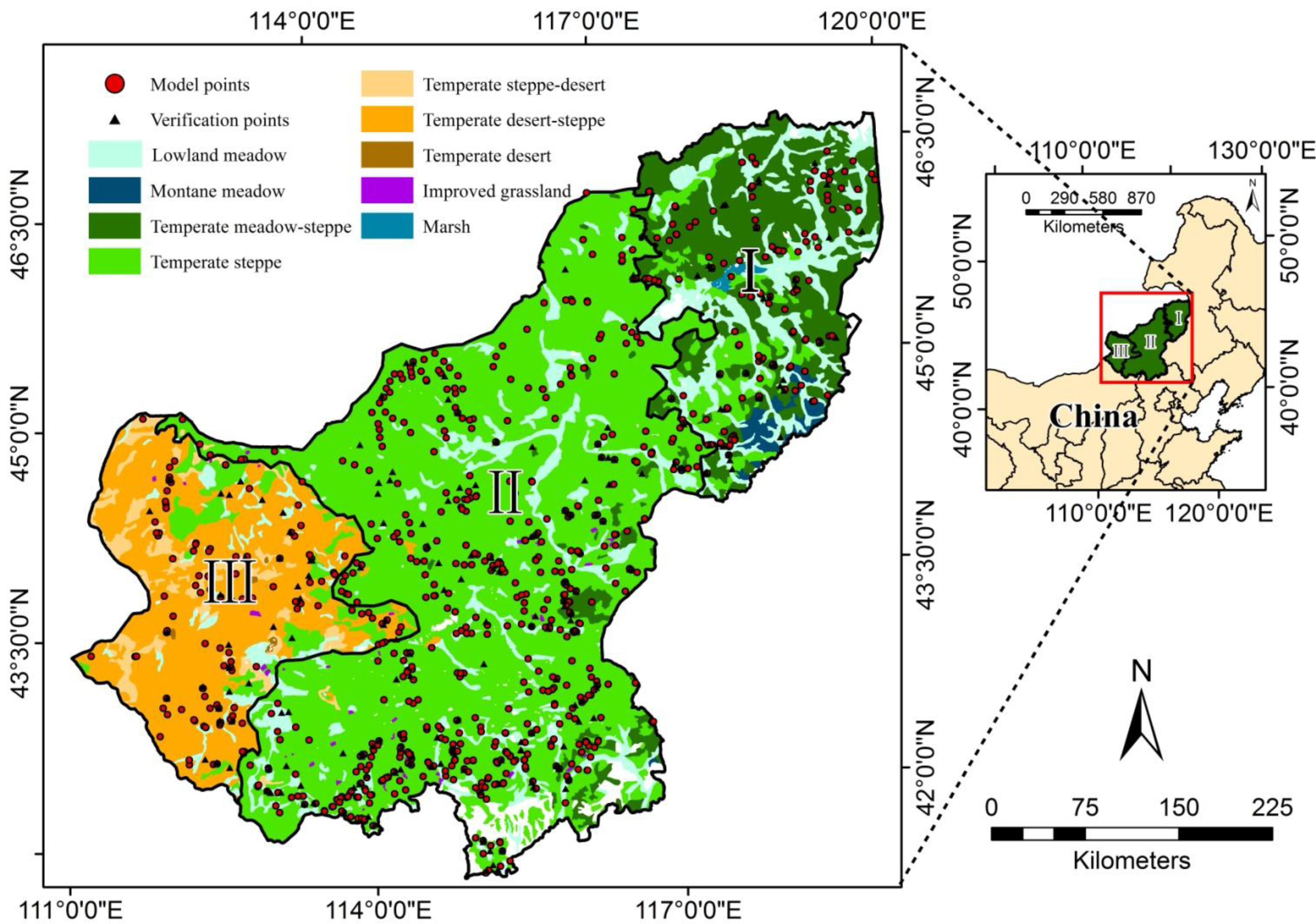
2.1. Study Area
2.2. Field data and Preprocessing
2.3. Remote Sensing Dataset and Preprocessing
2.4. Development and Verification of Estimation Models
3. Results
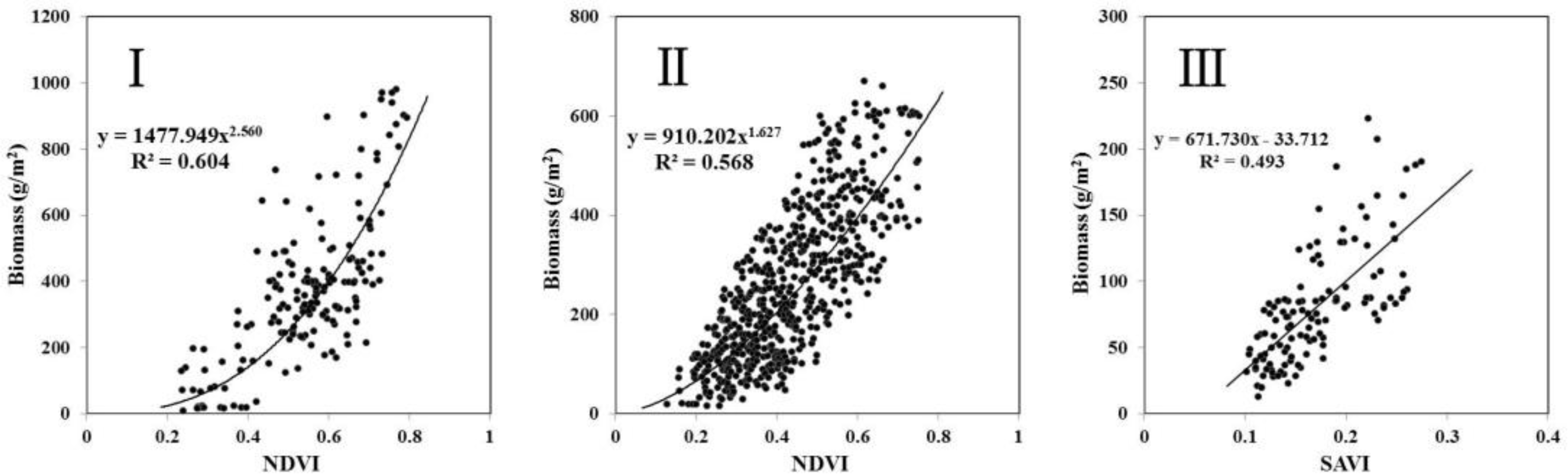
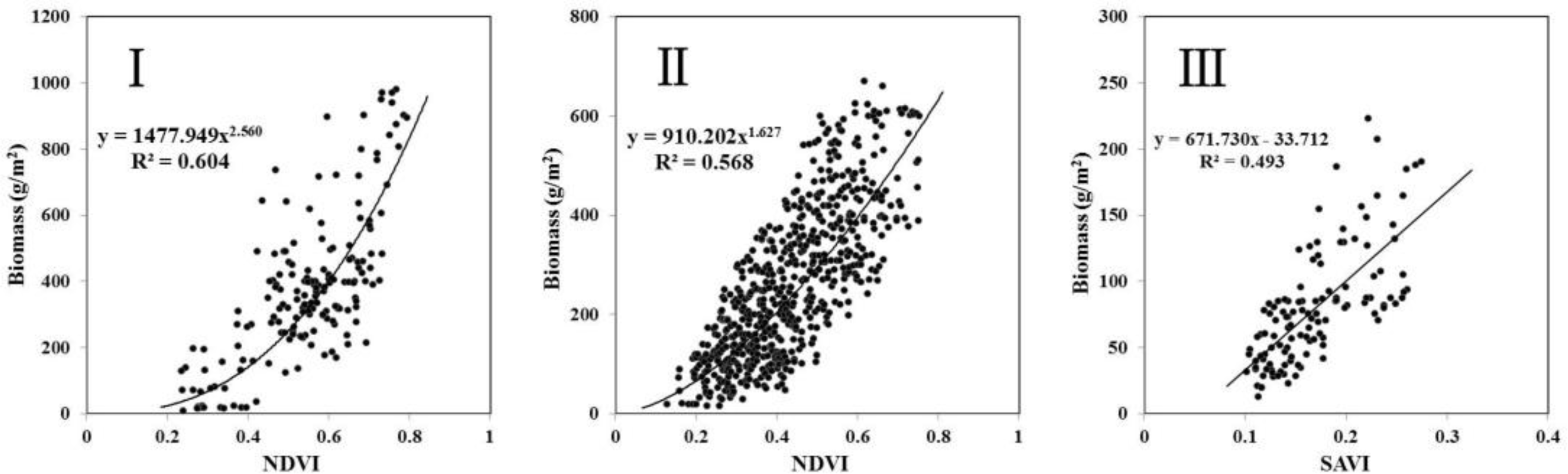
3.1. Relationship between Biomass and VIs
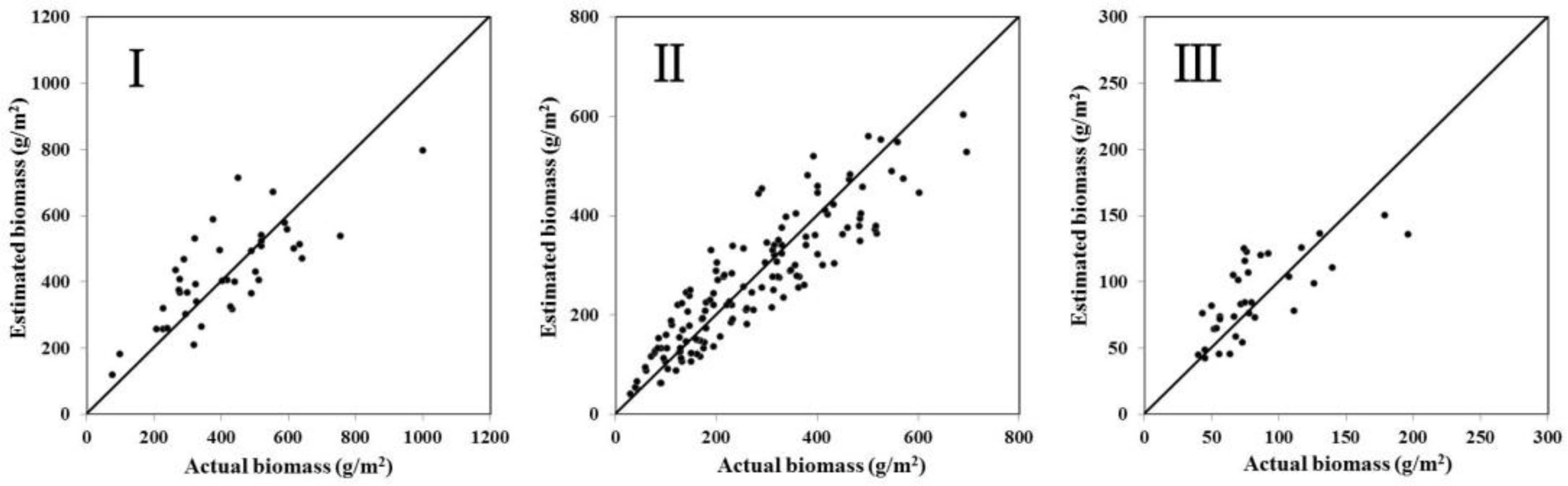
3.2. Development and Validation of Estimation Models
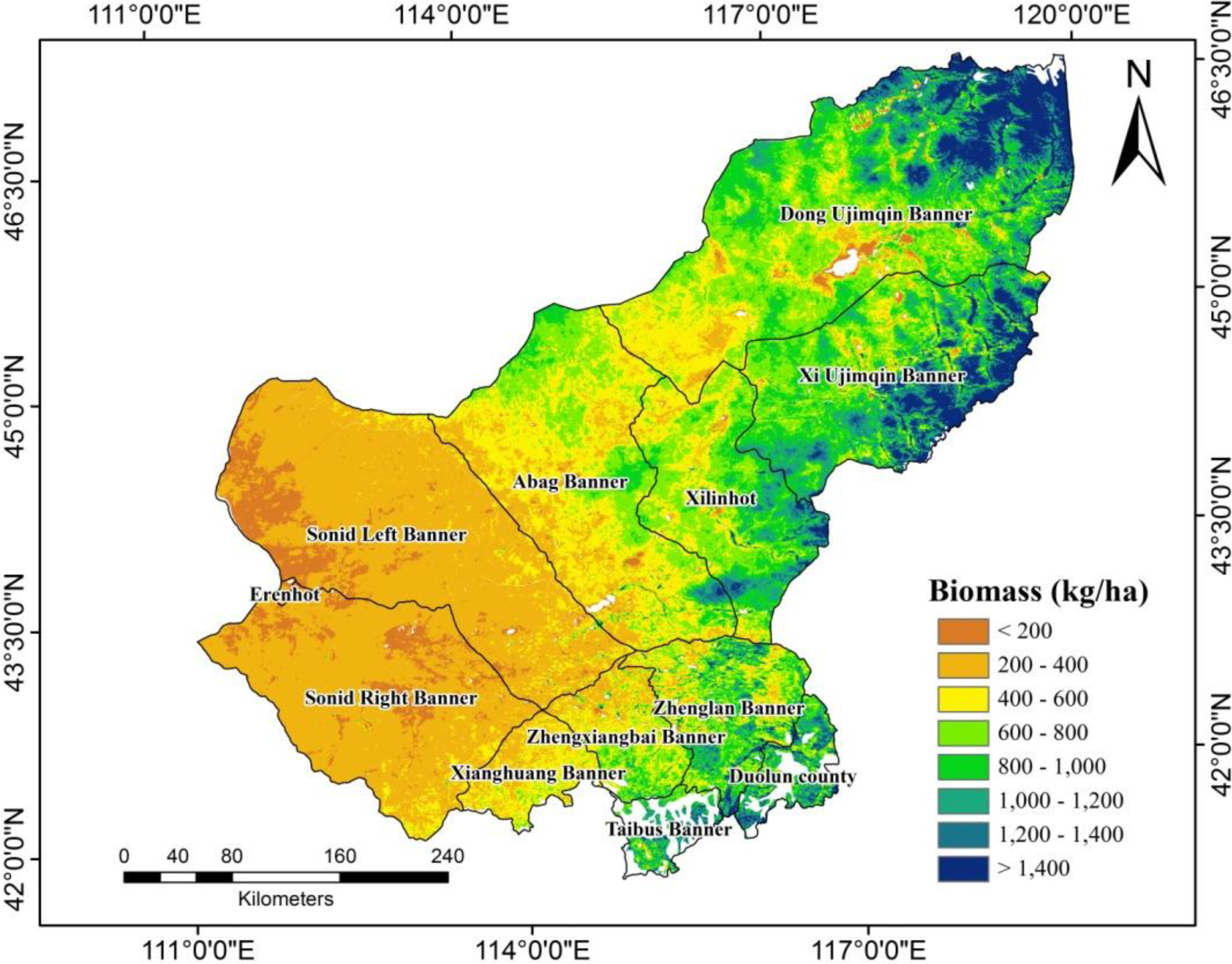
3.3. Spatial Distribution of Biomass in Xilingol
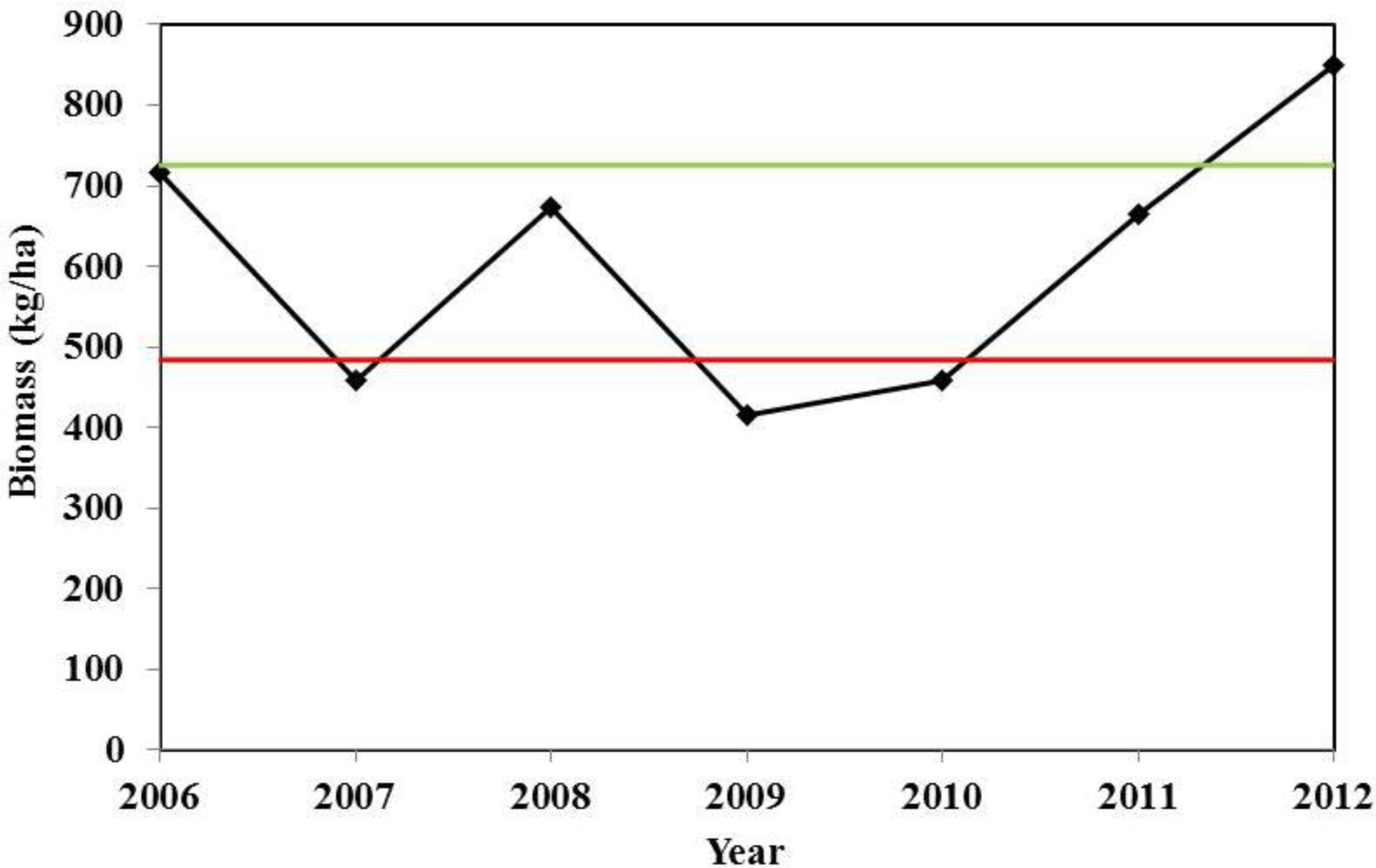
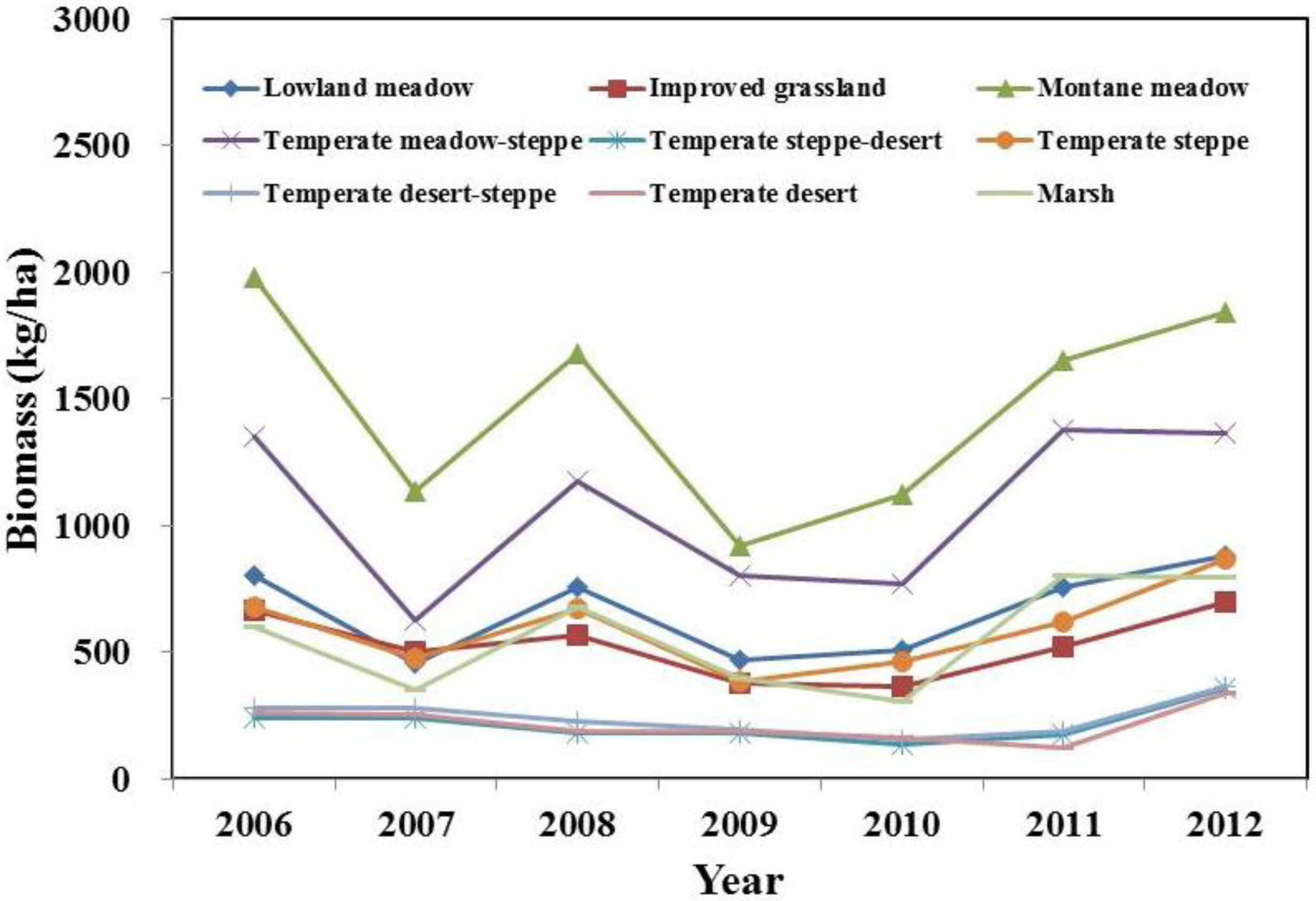
3.4. Interannual Variation of Biomass
4. Discussion
4.1. Model Development and Precision Validation
4.2. Temporal and Spatial Variation of Biomass
4.3. Differences between Biomass Estimates
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Adams, J.M.; Faure, H.; Faure-Denard, L.; McGlade, J.M.; Woodward, F.L. Increases in terrestrial carbon storage from the last glacial maximum to the present. Nature 1990, 348, 711–714. [Google Scholar]
- Scurlock, J.M.O.; Hall, D.O. The global carbon sink: A grassland perspective. Glob. Chang. Biol 1998, 4, 229–233. [Google Scholar]
- Peng, J.; Liu, Z.; Liu, Y.; Wu, J.; Han, Y. Trend analysis of vegetation dynamics in Qinghai—Tibet Plateau using Hurst Exponent. Ecol. Indic 2012, 14, 28–39. [Google Scholar]
- Department of Animal Husbandry and Veterinary, Rangeland Resources of China, 1st ed.; China Science and Technology Press: Beijing, China, 1996; pp. 188–207. (In Chinese)
- Scurlock, J.M.O.; Johnson, K.; Olson, R.J. Estimating net primary productivity from grassland biomass dynamics measurements. Glob. Chang. Biol 2002, 8, 736–753. [Google Scholar]
- Lu, D.S. The potential and challenge of remote sensing-based biomass estimation. Int. J. Remote Sens 2006, 27, 1297–1328. [Google Scholar]
- Flombaum, P.; Sala, O.E. A non-destructive and rapid method to estimate biomass and aboveground net primary production in arid environments. J. Arid Environ 2007, 69, 352–358. [Google Scholar]
- Xu, B.; Yang, X.C.; Tao, W.G.; Qin, Z.H.; Liu, H.Q.; Miao, J.M.; Bi, Y.Y. MODIS-based remote sensing monitoring of grass production in China. Int. J. Remote Sens 2008, 29, 5313–5327. [Google Scholar]
- Gaitán, J.J.; Bran, D.; Oliva, G.; Ciari, G.; Nakamatsu, V.; Salomone, J.; Ferrante, D.; Buono, G.; Massara, V.; Humano, G.; et al. Evaluating the performance of multiple remote sensing indices to predict the spatial variability of ecosystem structure and functioning in Patagonian steppes. Ecol. Indic 2013, 34, 181–191. [Google Scholar]
- Gao, Y.; Liu, X.; Min, C.; He, H.; Yu, G.; Liu, M.; Zhu, X.; Wang, Q. Estimation of the North-South Transect of eastern China forest biomass using remote sensing and forest inventory data. Int. J. Remote Sens 2013, 34, 5598–5610. [Google Scholar]
- Piao, S.L.; Fang, J.Y.; Zhou, L.M.; Tan, K.; Tao, S. Changes in biomass carbon stocks in China’s grasslands between 1982 and 1999. Glob. Biogeochem. Cycles 2007, 21. [Google Scholar] [CrossRef]
- Butterfield, H.S.; Malmström, C.M. The effects of phenology on indirect measures of aboveground biomass in annual grasses. Int. J. Remote Sens 2009, 30, 3133–3146. [Google Scholar]
- Jin, Y.X.; Xu, B.; Yang, X.C.; Li, J.Y.; Wang, D.L.; Ma, H.L. Remote sensing dynamic estimation of grass production in Xilingol, Inner Mongolia. Sci. Sin. Vitae 2011, 41, 1185–1195. [Google Scholar]
- Leisher, C.; Hess, S.; Boucher, T.M.; van Beukering, P.; Sanjayan, M. Measuring the impacts of community-based grasslands management in Mongolia’s Gobi. PLoS One 2012, 7. [Google Scholar] [CrossRef]
- Ouyang, W.; Hao, F.H.; Skidmore, A.K.; Groen, T.A.; Toxopeus, A.G.; Wang, T.J. Integration of multi-sensor data to assess grassland dynamics in a Yellow River sub-watershed. Ecol. Indic 2012, 18, 163–170. [Google Scholar]
- Gao, T.; Xu, B.; Yang, X.C.; Jin, Y.X.; Ma, H.L.; Li, J.Y.; Yu, H.D. Using MODIS time series data to estimate aboveground biomass and its spatio-temporal variation in Inner Mongolia’s grassland between 2001 and 2011. Int. J. Remote Sens 2013, 34, 7796–7810. [Google Scholar]
- Casady, G.; van Leeuwen, W.; Reed, B. Estimating winter annual biomass in the Sonoran and Mojave deserts with satellite- and ground-based observations. Remote Sens 2013, 5, 909–926. [Google Scholar]
- Lu, D.S. Aboveground biomass estimation using Landsat TM data in the Brazilian Amazon. Int. J. Remote Sens 2005, 26, 2509–2525. [Google Scholar]
- Mutanga, O.; Skidmore, A.K. Hyperspectral band depth analysis for a better estimation of grass biomass (Cenchrus ciliaris) measured under controlled laboratory conditions. Int. J. Appl. Earth Obs. Geoinf 2004, 5, 87–96. [Google Scholar]
- Anaya, J.A.; Chuvieco, E.; Palacios-Orueta, A. Aboveground biomass assessment in Colombia: A remote sensing approach. For. Ecol. Manag 2009, 257, 1237–1246. [Google Scholar]
- Ren, H.R.; Zhou, G.S.; Zhang, X.S. Estimation of green aboveground biomass of desert steppe in Inner Mongolia based on red-edge reflectance curve area method. Biosyst. Eng 2011, 109, 385–395. [Google Scholar]
- Yan, F.; Wu, B.; Wang, Y.J. Estimating aboveground biomass in Mu Us Sandy Land using Landsat spectral derived vegetation indices over the past 30 years. J. Arid Land 2013, 5, 521–530. [Google Scholar]
- Boschetti, M.; Bocchi, S.; Brivio, P.A. Assessment of pasture production in the Italian Alps using spectrometric and remote sensing information. Agric. Ecosyst. Environ 2007, 118, 267–272. [Google Scholar]
- Shinoda, M.; Gillies, J.A.; Mikami, M.; Shao, Y. Temperate grasslands as a dust source: Knowledge, uncertainties, and challenges. Aeolian Res 2011, 3, 271–293. [Google Scholar]
- Shiyomi, M.; Akiyama, T.; Wang, S.P.; Yiruhan; Ailikun; Hori, Y.; Chen, Z.Z.; Yasuda, T.; Kawamura, K.; Yamamura, Y. A grassland ecosystem model of the Xilingol steppe, Inner Mongolia, China. Ecol. Model 2011, 222, 2073–2083. [Google Scholar]
- Yang, X.C.; Xu, B.; Zhu, X.H.; Tao, W.G.; Liu, T.K. Models of grass production based on remote sensing monitoring in northern agro-grazing Ecotone. Geogr. Res 2007, 26, 213–221. [Google Scholar]
- Piao, S.L.; Fang, J.Y.; He, J.S.; Xiao, Y. Spatial distribution of grassland biomass in China. Acta Phytoecol. Sin 2004, 28, 491–498. [Google Scholar]
- Fang, J.Y.; Liu, G.H.; Xu, S.L. Carbon Pools in Terrestrial Ecosystems in China. In Emissions and Their Relevant Processes of Greenhouse Gases in China, 1st ed; Wang, G.C., Wen, Y.P., Eds.; China Environment Science Press: Beijing, China, 1996; pp. 109–128. (In Chinese) [Google Scholar]
- Muukkonen, P.; Heiskanen, J. Biomass estimation over a large area based on standwise forest inventory data and ASTER and MODIS satellite data: A possibility to verify carbon inventories. Remote Sens. Environ 2007, 107, 617–624. [Google Scholar]
- Rouse, J.W.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring Vegetation Systems in the Great Plains with ERTS. Proceedings of the Third Earth Resources Technology Satellite-1 Symposium, Washington, DC, USA, 10–14 December 1973; pp. 309–317.
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ 1979, 8, 127–150. [Google Scholar]
- Jiang, Z.; Huete, A.R.; Didan, K.; Miura, T. Development of a two-band enhanced vegetation index without a blue band. Remote Sens. Environ 2008, 112, 3833–3845. [Google Scholar]
- Huete, A.R. A soil adjusted vegetation index (SAVI). Remote Sens. Environ 1988, 25, 295–309. [Google Scholar]
- Qi, J.; Chehbouni, A.; Huete, A.R.; Kerr, Y.H.; Sorooshian, S. A modified soil adjusted vegetation index. Remote Sens. Environ 1994, 48, 119–126. [Google Scholar]
- Zhang, L.Y.; Zhang, J.X.; Sai, Y.J.Y.; Bao, H.X.; Shan, J.G.; Guo, Z.Z. Remote sensing monitoring model for grassland vegetation biomass monitoring in typical steppe: A case study from Xilingol. Pratacult. Sci 2008, 25, 31–36. [Google Scholar]
- Bai, Y.F.; Chen, Z.Z. Effects of long-term variability of plant species and functional groups on stability of a Leymus Chinensis community in the Xilin river basin, Inner Mongolia. Acta Phytoecol. Sin 2000, 24, 641–647. [Google Scholar]
- Hou, Y.Y.; Mao, L.X.; Li, C.S.; Qian, S. Spatiotemporal variation pattern of vegetation’s net primary productivity in China. Chin. J. Ecol 2008, 27, 1455–1460. [Google Scholar]
- Ni, J. Estimating net primary productivity of grasslands from field biomass measurements in temperate northern China. Plant Ecol 2004, 174, 217–234. [Google Scholar]
- Paruelo, J.M.; Lauenroth, W.K.; Burke, I.C.; Sala, O.E. Grassland precipitation-use efficiency varies across a resource gradient. Ecosystems 1999, 2, 64–68. [Google Scholar]
- Bai, Y.; Han, X.G.; Wu, J.G.; Chen, Z.Z.; Li, L.H. Ecosystem stability and compensatory effects in the Inner Mongolia grassland. Nature 2004, 431, 181–184. [Google Scholar]
- Hu, Z.; Fan, J.W.; Zhong, H.P.; Yu, G.R. Spatiotemporal dynamics of aboveground primary productivity along a precipitation gradient in Chinese temperate grassland. Sci. China Earth Sci 2007, 50, 754–764. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
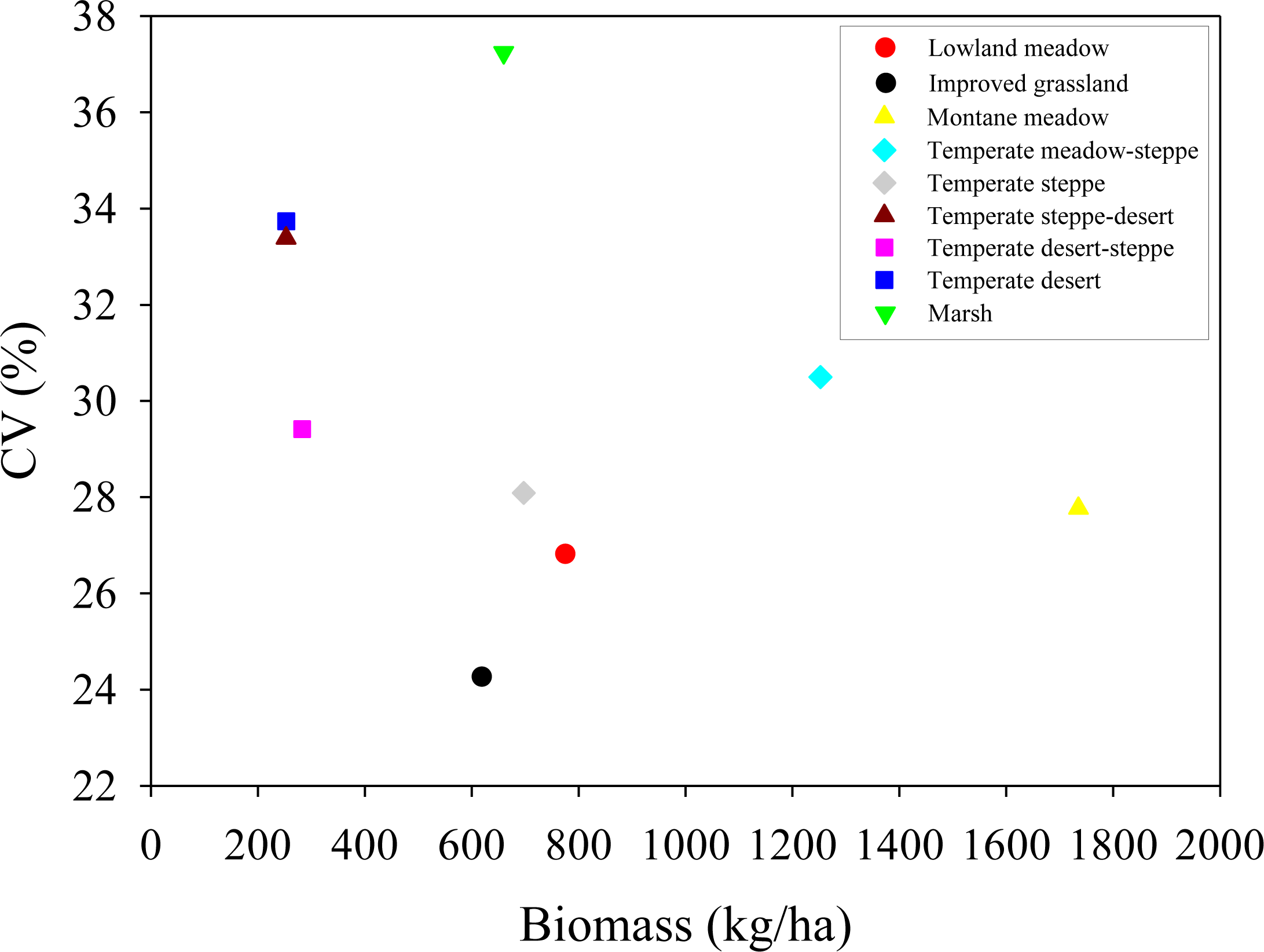
| Grassland Types | Conversion Coefficients | Grassland Types | Conversion Coefficients |
|---|---|---|---|
| Lowland meadow | 1/3.5 | Temperate steppe | 1/3.0 |
| Improved grassland | 1/3.2 | Temperate desert-steppe | 1/2.7 |
| Montane meadow | 1/3.5 | Temperate desert | 1/2.5 |
| Temperate meadow-steppe | 1/3.2 | Marsh | 1/4.0 |
| Temperate steppe-desert | 1/2.5 |
| Biomass | NDVI | DVI | EVI2 | SAVI | MSAVI |
|---|---|---|---|---|---|
| I Meadow steppe region (n = 214) | 0.731 | 0.664 | 0.709 | 0.711 | 0.708 |
| II Typical steppe region (n = 766) | 0.791 | 0.641 | 0.736 | 0.743 | 0.728 |
| III Desert steppe region (n = 154) | 0.686 | 0.693 | 0.702 | 0.702 | 0.702 |
| Vegetation Index | Model | R2 | F Value | RMSE (kg/ha) | REE | Precision (%) | |
|---|---|---|---|---|---|---|---|
| I Meadow steppe region (n = 173, test_n = 41) | NDVI | y = 1477.949x2.560 | 0.604 | 261.096 | 1141.45 | 0.264 | 73.6 |
| DVI | y = 507.373ln x + 1240.408 | 0.464 | 148.016 | 1544.24 | 0.346 | 65.4 | |
| EVI2 | y = 1711.819x − 182.167 | 0.515 | 181.910 | 1376.94 | 0.316 | 68.4 | |
| SAVI | y = 1829.285x − 235.148 | 0.518 | 183.889 | 1370.66 | 0.313 | 68.7 | |
| MSAVI | y = 1721.413x − 160.114 | 0.515 | 181.423 | 1379.73 | 0.318 | 68.2 | |
| II Typical steppe region (n = 629, test_n = 137) | NDVI | y = 910.202x1.627 | 0.568 | 825.644 | 673.88 | 0.261 | 73.9 |
| DVI | y = 2073.410x − 98.397 | 0.387 | 396.641 | 1027.50 | 0.403 | 59.7 | |
| EVI2 | y = 1344.033x − 110.769 | 0.513 | 611.698 | 843.02 | 0.321 | 67.9 | |
| SAVI | y = 1403.755x − 140.048 | 0.523 | 687.838 | 826.64 | 0.325 | 67.5 | |
| MSAVI | y = 1379.324x − 104.273 | 0.502 | 631.358 | 862.30 | 0.331 | 66.9 | |
| III Desert steppe region (n = 119, test_n = 35) | NDVI | y = 486.989x − 27.719 | 0.484 | 109.625 | 281.91 | 0.267 | 73.3 |
| DVI | y = 978.067x − 33.273 | 0.475 | 105.913 | 255.29 | 0.266 | 73.4 | |
| EVI2 | y = 677.344x − 28.493 | 0.491 | 113.082 | 256.68 | 0.260 | 74.0 | |
| SAVI | y = 671.730x − 33.712 | 0.493 | 113.708 | 258.96 | 0.261 | 73.9 | |
| MSAVI | y = 706.893x − 30.412 | 0.490 | 112.399 | 255.45 | 0.261 | 73.9 | |
| Banner* | Grassland Area (km2) | Wet Weight of Biomass (t) | Yield of Biomass (t) | Biomass (kg/ha) |
|---|---|---|---|---|
| Abag Banner | 27,325 | 5,085,442 | 1,428,672 | 522.84 |
| Dong Ujimqin Banner | 43,861 | 13,929,648 | 3,701,216 | 843.85 |
| Duolun County | 2,996 | 1,124,055 | 300,395 | 1,002.65 |
| Erenhot | 174 | 9,618 | 3,028 | 174.02 |
| Sonid Left Banner | 34,618 | 2,985,264 | 895,988 | 258.82 |
| Sonid Right Banner | 25,148 | 2,089,824 | 614,094 | 244.19 |
| Xilinhot | 15,713 | 4,175,067 | 1,152,669 | 733.58 |
| Xi Ujimqin Banner | 23,587 | 8,736,553 | 2,323,326 | 985.00 |
| Xianghuang Banner | 4,885 | 744,731 | 208,519 | 426.86 |
| Zhenglan Banner | 10,142 | 2,689,253 | 743,844 | 733.43 |
| Zhengxiangbai Banner | 6,084 | 1,170,427 | 325,048 | 534.27 |
| Taibus Banner | 1,652 | 590,210 | 162,600 | 984.26 |
| Total | 196,185 | 43,330,094 | 1,1859,399 | 604.50 |
| Grassland Types | Grassland Area (km2) | Wet Weight of Biomass (t) | Yield of Biomass (t) | Biomass (kg/ha) |
|---|---|---|---|---|
| Lowland meadow | 25,955 | 7,041,867 | 1,710,168 | 658.90 |
| Montane meadow | 1,593 | 967,275 | 234,910 | 1,474.64 |
| Temperate meadow-steppe | 24,658 | 9,883,646 | 2,625,343 | 1,064.70 |
| Temperate steppe | 108,370 | 2,266,8551 | 6,422,757 | 592.67 |
| Temperate steppe-desert | 5,082 | 321,088 | 109,170 | 214.82 |
| Temperate desert-steppe | 29,576 | 2,255,945 | 710,205 | 240.13 |
| Temperate desert | 140 | 8,855 | 3,011 | 215.04 |
| Improved grassland | 473 | 93,678 | 24,883 | 526.07 |
| Marsh | 338 | 89,189 | 18,953 | 560.73 |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Jin, Y.; Yang, X.; Qiu, J.; Li, J.; Gao, T.; Wu, Q.; Zhao, F.; Ma, H.; Yu, H.; Xu, B. Remote Sensing-Based Biomass Estimation and Its Spatio-Temporal Variations in Temperate Grassland, Northern China. Remote Sens. 2014, 6, 1496-1513. https://doi.org/10.3390/rs6021496
Jin Y, Yang X, Qiu J, Li J, Gao T, Wu Q, Zhao F, Ma H, Yu H, Xu B. Remote Sensing-Based Biomass Estimation and Its Spatio-Temporal Variations in Temperate Grassland, Northern China. Remote Sensing. 2014; 6(2):1496-1513. https://doi.org/10.3390/rs6021496
Chicago/Turabian StyleJin, Yunxiang, Xiuchun Yang, Jianjun Qiu, Jinya Li, Tian Gao, Qiong Wu, Fen Zhao, Hailong Ma, Haida Yu, and Bin Xu. 2014. "Remote Sensing-Based Biomass Estimation and Its Spatio-Temporal Variations in Temperate Grassland, Northern China" Remote Sensing 6, no. 2: 1496-1513. https://doi.org/10.3390/rs6021496