Early Detection of Bark Beetle Green Attack Using TerraSAR-X and RapidEye Data

Abstract
:1. Introduction
2. Material and Methods
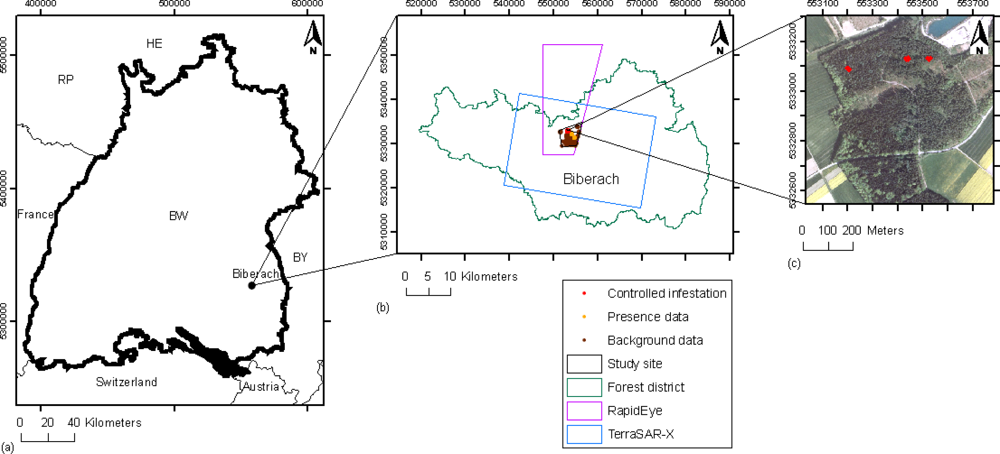
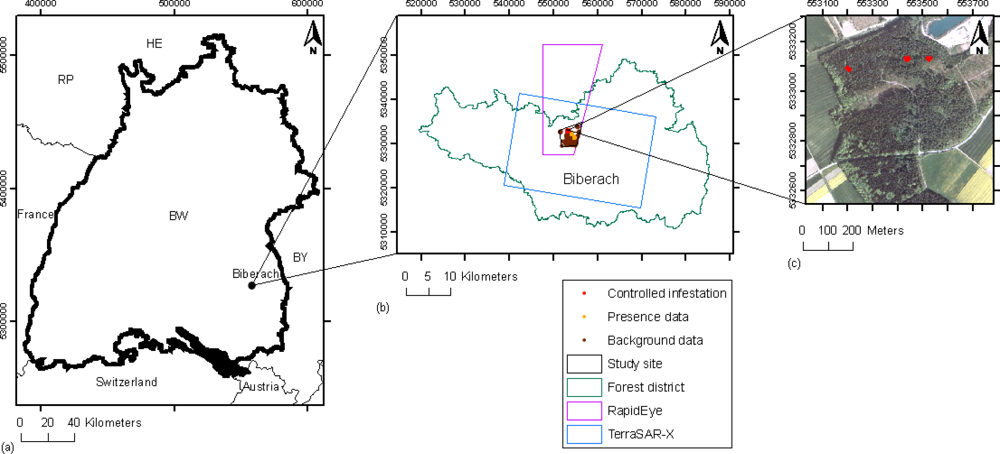
2.1. Study Site and Field Data
2.2. Satellite Data
2.2.1. Pre-Processing of RapidEye Data
2.2.2. Pre-Processing of TerraSAR-X Data
2.3. Statistical Modelling
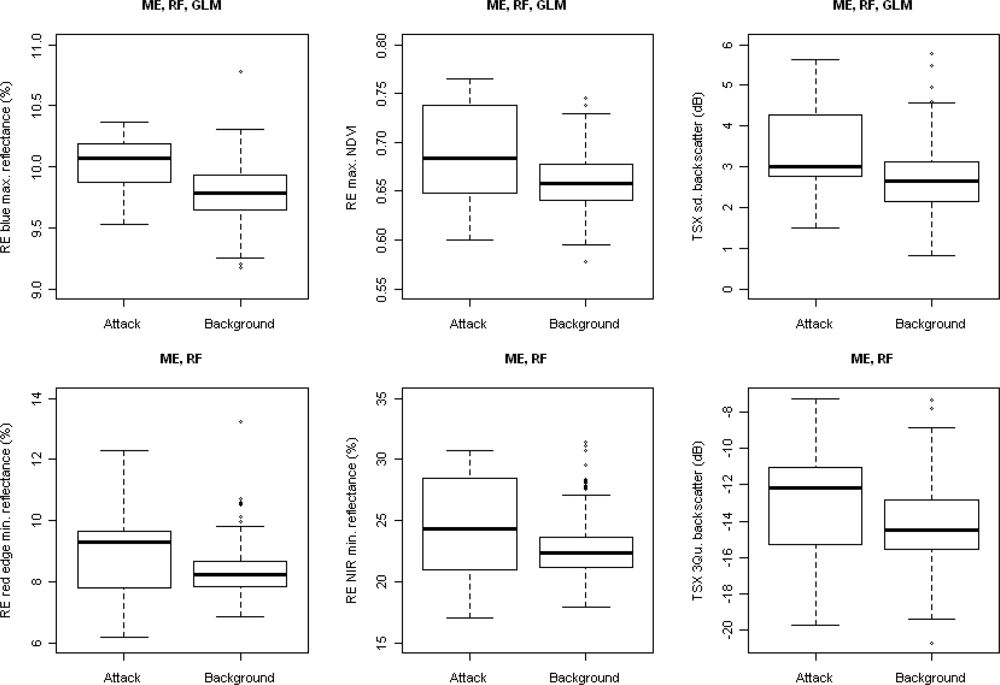
2.3.1. Explanatory Variables
2.3.2. Model Types
(a) Generalized Linear Model (GLM)
(b) Maximum Entropy (ME)
(c) Random Forest (RF)
2.3.3. Final Model Selection
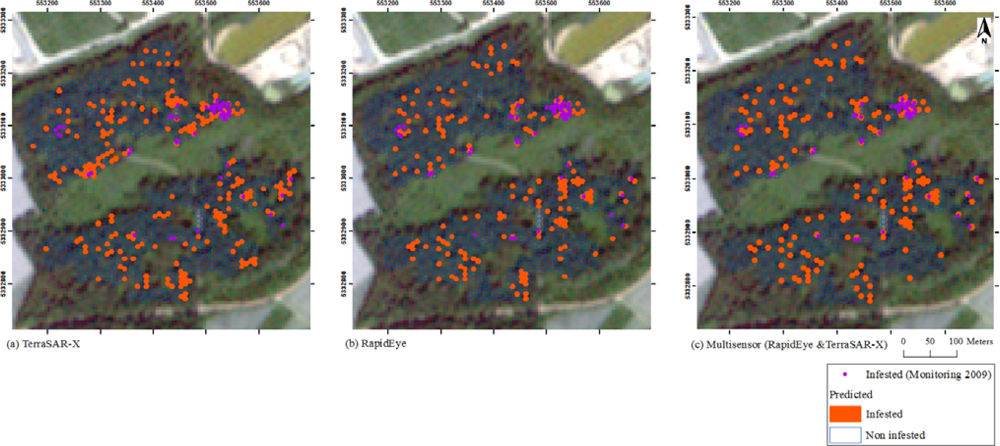
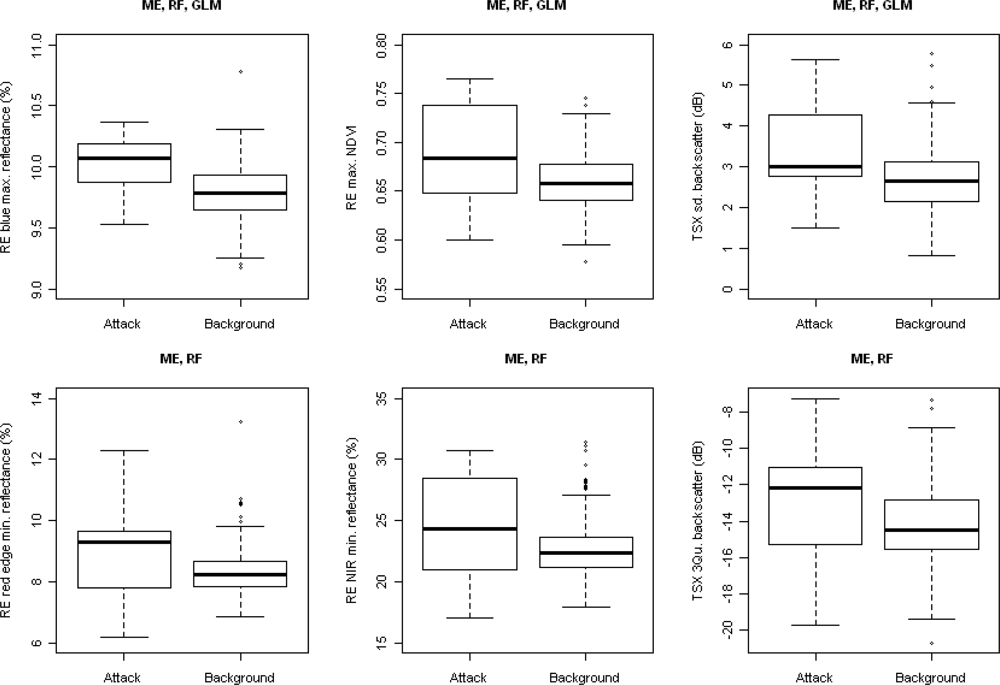
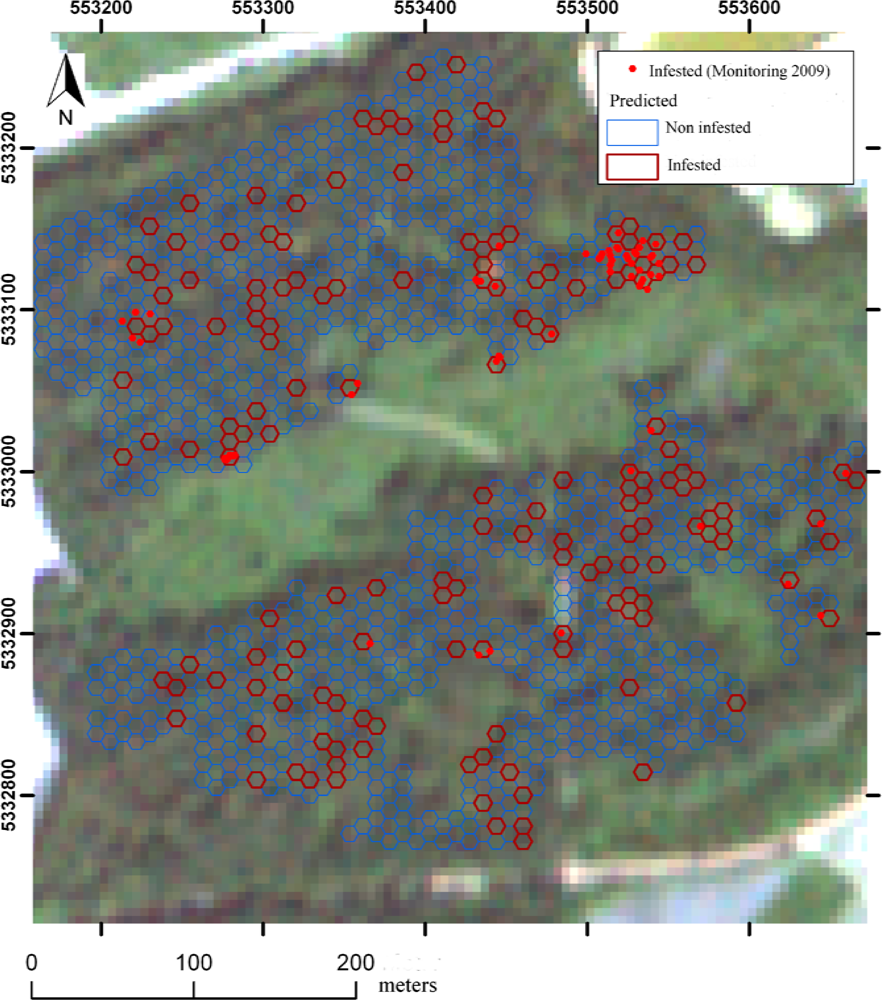
3. Results
4. Discussion
5. Conclusions
Acknowledgments
References and Notes
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Garcia, J.; Seidl, R.; Delzon, S.; Corona, P.; Kolström, M.; et al. Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. Forest Ecol. Manag 2010, 259, 698–709. [Google Scholar]
- Meining, S.; Wilpert, K.V.; Schröter, H.; Schäffer, J.; Morgenstern, Y.; Zirlewagen, D. Waldzustandsbericht 2009 der Forstlichen Versuchsund Forschungsanstalt Baden-Württemberg (FVA); ForstBW: Freiburg, Germany, 2010; pp. 1–72. [Google Scholar]
- Wulder, M.; White, J.; Coops, N.; Han, T.; Alvarez, M.; Butson, C.; Yuan, X. A Procedure for Mapping and Monitoring Mountain Pine Beetle Red Attack Forest Damage Using Landsat Imagery; Pacific Forestry Centre, Canadian Forest Service, Natural Resources Canada: Victoria, BC, Canada, 2006; pp. 1–40. [Google Scholar]
- Deshayes, M.; Guyon, D.; Jeanjean, H.; Stach, N.; Jolly, A.; Hagolle, O. The contribution of remote sensing to the assessment of drought effects in forest ecosystems. Ann. Forest Sci 2006, 63, 579–595. [Google Scholar]
- Dong, J.; Zhuang, D.; Huang, Y.; Fu, J. Advances in multi-sensor data fusion: Algorithms and Applications. Sensors 2009, 9, 7771–7784. [Google Scholar]
- Xia, J.; Du, P.; Cao, W. Classification of high resolution optical and SAR fusion image using fuzzy knowledge and object-oriented paradigm. Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 2010, XXXVIII-4/C7, 1–5. [Google Scholar]
- Eitel, J.; Vierling, L.; Litvak, M.; Long, D.; Schulthess, U.; Ager, A.; Krofcheck, D.; Stoscheck, L. Broadband, red-edge information from satellites improves early stress detection in a New Mexico conifer woodland. Remote Sens. Environ 2011, 115, 3640–3646. [Google Scholar]
- Zarco-Tejada, P.; Sepulcre-Cantó, G. Remote Sensing of Vegetation Biophysical Parameters for Detecting Stress Condition and Land Cover Changes. In Estudios de la Zona no Saturada del Suelo Vol. VIII; Juan Vicente Giráldez Cervera y Francisco José Jiménez Hornero: Córdoba, Spain, 2007; pp. 37–44. [Google Scholar]
- Roberts, A.; Dragicevic, S.; Northrup, J.; Wolf, S.; Li, Y.; Coburn, C. Mountain Pine Beetle Detection and Monitoring: Remote Sensing Evaluations; FII Operational Report with Reference to Recipient Agreement R2003-0205; Department of Geography, Simon Fraser University: Vancouver, BC, Canada, 2003; pp. 1–45. Available online: http://www.for.gov.bc.ca/hfd/library/fia/html/FIA2003MR271.htm (accessed on 11 November 2012).
- Murtha, P.; Wiart, R. PC-based digital analysis of mountain pine beetle current-attacked and non-attacked lodgepole pine. Can. J. Remote Sens 1989, 15, 70–76. [Google Scholar]
- Heath, J. Detection of Mountain Pine Beetle Green Attacked Lodgepole Pine Using Compact Airborne Spectrographic Imagery (CASI) Data. 2001; pp. 1–72. Available online: https://circle.ubc.ca/handle/2429/11545 (accessed on 11 November 2012). [Google Scholar]
- Roberts, A.; Northrup, J.; Reich, R. Mountain Pine Beetle Detection and Monitoring: Replication Trials for Early Detection. Proceedings of Third International Workshop on the Analysis of Multi-temporal Remote Sensing Images, Biloxi, MS, USA, 16–18 May 2005; pp. 20–24.
- Schweigler, P. Früherkennung von Buchdruckerbefall. 2007; pp. 1–73. [Google Scholar]
- Marx, A. Erkennung von borkenkäferbefall in fichtenreinbeständen mit multi-temporalen rapideye- satellitenbildern und datamining-techniken. Photogramm. Fernerk. Geoinf 2010, 4, 243–252. [Google Scholar]
- Tanase, M.; Santoro, M.; Wegmuller, U.; de la Riva, J.; Perez-Cabello, F. Properties of X-, C- and L-band repeat-pass interferometric SAR coherence in Mediterranean pine forests affected by fires. Remote Sens. Environ 2010, 114, 2182–2194. [Google Scholar]
- Tanase, M.; de la Riva, J.; Santoro, M.; Pérez, F.; Kasischke, E. Sensitivity of SAR data to post-fire forest regrowth in Mediterranean and boreal forests. Remote Sens. Environ 2011, 115, 2075–2085. [Google Scholar]
- Solberg, S.; Astrup, R.; Lyytikainen-Saarenmaa, P.; Kantola, T.; Holopainen, M.; Kaartinen, H. Testing TerraSAR-X for Forest Disturbance Mapping. Proceedings of 4th TerraSAR-X Science Team Meeting, Oberpfaffenhofen, Germany, 14–16 February 2011; pp. 1–7.
- Ackermann, J.; Klaus, M. Erfassung großflächiger Insektenfraßschäden an Kiefer durch Radardaten. AFZ-Der Wald 2011, 6, 20–22. [Google Scholar]
- Reid, R. Moisture changes in lodgepole pine before and after attack by the mountain pine beetle. Forestry Chron 1961, 37, 368–375. [Google Scholar]
- Wermelinger, B. Ecology and management of the spruce bark beetle Ips typographus—A review of recent research. Forest Ecol. Manag 2004, 202, 67–82. [Google Scholar]
- de Jong, J.; de Groot, H.; Klaassen, W.; Kuiper, P. Radar Backscatter Change from an Ash in Relation to Its Hydrological Properties. Proceedings of the IEEE International Geoscience and Remote Sensing Symposium IGARSS 2000, Honolulu, HI, USA, 24–28 July 2000; 7, pp. 2927–2929.
- Albertz, J.; Wiggenhage, M. Guide for Photogrammetry and Remote Sensing; Herbert Wichmann Verlag: Berlin und Offenbach, Germany, 2009; p. 334. [Google Scholar]
- Hernandez, PA.; Graham, C.; Master, L.; Albert, D. The effect of sample size and species characteristics on performance of different species distribution modeling methods. Ecography 2006, 29, 773–785. [Google Scholar]
- Coggins, S.; Coops, N.; Wulder, M. Improvement of low level bark beetle damage estimates with adaptive cluster sampling. Silva Fenn 2010, 44, 289–301. [Google Scholar]
- Valavanis, V.; Pierce, G.; Zuur, A.; Palialexis, A.; Saveliev, A.; Katara, I.; Wang, J. Modelling of essential fish habitat based on remote sensing, spatial analysis and GIS. Hydrobiologia 2008, 612, 5–20. [Google Scholar]
- Elith, J.; Phillips, S.; Hastie, T.; Dudk, M.; Chee, Y.; Yates, C. A statistical explanation of MaxEnt for ecologists. Divers. Distrib 2011, 17, 43–57. [Google Scholar]
- Forst, BW. Forest inventory in the forest district of Biberach (internal digital database). 2003. [Google Scholar]
- RapidEye AG. RapidEye Standard Image Product Specifications; RapidEye AG: Brandenburg an der Havel, Germany, 2011; pp. 1–54. [Google Scholar]
- Düring, R.; Koudogbo, F.; Weber, M. TerraSAR-X and TanDEM-X: Revolution in Spaceborne Radar. Int .Arch. Photogramm. Remote Sens. Spat. Inf. Sci 2008, 37, 227–234. [Google Scholar]
- DLR. TerraSAR-X, Ground Segment, Basic Products Specification Document; Deutsches Zentrum für Luft und Raumfahrt: Oberpfaffenhofen, Germany, 2006; pp. 1–45. [Google Scholar]
- Nowak, J.; Tokarczyk, P. Analysis of the Geometric Quality of the LPIS-Based RapidEye Level 3A; JRC Scientific and Technical Reports; JRC European Commission: Ispra, Italy, 2010; pp. 1–47. [Google Scholar]
- Jackson, R.; Huete, R. Interpreting vegetation indices. Preventive Veterinary Medicine 1991, 11, 185–200. [Google Scholar]
- Infoterra. Radiometric Calibration of TerraSAR-X Data; Infoterra GmbH: Friedrichshafen, Germany, 2008; pp. 1–16. [Google Scholar]
- Schleyer, A. Das Laserscan-DGM von Baden-Wuerttemberg. Photogramm. Week 2001, 1, 217–225. [Google Scholar]
- Weinacker, H.; Koch, B.; Weinacker, R. TREESVIS: A Software System for Simultaneous ED-Real-Time Visualisation of DTM, DSM, Laser Raw Data, Multispectral Data, Simple Tree and Building Models. Proceedings of the ISPRS Working Group VIII (2), Freiburg, Germany, 3–6 October 2004; pp. 90–95.
- Wegmüller, U. Automated Terrain Corrected SAR Geocoding. Proceedings of the IEEE International Geoscience and Remote Sensing Symposium IGARSS’99, Hamburg, Germany, 28 June–2 July 1999; pp. 1712–1714.
- Werner, C.; Wegmüller, U.; Strozzi, T.; Wiesmann, A. Gamma SAR and Interferometry Software. Proceedings of ERS-ENVISAT Symposium, Gothenburg, Sweden, 16–20 October 2000; pp. 1–9.
- Rouse, J.; Hass, R.; Schell, J.; Harlan, C. Monitoring the Vernal Advancement of Retrogradation (Greenwave Effect) of Natural Vegetation. NASA Final Report E75-10354, NASA-CR-144661, RSC-1978-4. 1974, pp. 1–390. Available online: http://ntrs.nasa.gov/search.jsp?R=19750020419 (accessed on 11 November 2012).
- Buschmann, C.; Nagel, E. In vivo spectroscopy and internal optics of leaves as basis for remote sensing of vegetation. Int. J. Remote Sens 1993, 14, 711–722. [Google Scholar]
- Gitelson, A.; Merzlyak, M. Remote estimation of chlorophyll content in higher plant leaves. Int. J. Remote Sens 1996, 18, 2691–2697. [Google Scholar]
- Barnes, E.; Richard, S.; Colaizzi, P.; Haberland, J.; Kostrzewski, M.; Waller, P.; Choi, C. Coincindent Detection of Crop Water Stress, Nitrogen Status and Canopy Density Using Ground-Based Multispectral Data. Proceedings of the Fifth International Conference on Precision Agriculture, Bloomington, IN, USA, 16–19 July 2000; pp. 1–15.
- Gitelson, A.; Viña, A.; Ciganda, V.; Rundquist, D.; Arkebauer, T. Remote estimation of canopy chlorophyll content in crops. Geophys. Res. Lett 2005, 32, L08403. [Google Scholar] [CrossRef]
- Phillips, S.; Anderson, R.; Schapire, R. Maximum entropy modeling of species geographic distributions. Ecol. Model 2006, 190, 231–259. [Google Scholar]
- Venables, W.; Ripley, B. Generalized Linear Models. In Modern Applied Statistics with S. Statistics and Computing, 4th ed.; Springer: New York, NY, USA, 2002; pp. 183–208. [Google Scholar]
- McCullagh, P.; Nelder, J. Generalized Linear Models, 2nd ed.; Chapman and Hall: London, UK, 1989; pp. 1–506. [Google Scholar]
- R Development Core Team. R A Language and Environment for Statistical Computing. 2010. Available online: http://www.r-project.org/ (accessed on 15 September 2012).
- Pinheiro, J.; Bates, D. Mixed-Effects Models in S and S-PLUS; Springer: New York, NY, USA, 2000; pp. 1–523. [Google Scholar]
- Braunisch, V.; Patthey, P.; Arlettaz, R. Spatially explicit modeling of conflict zones between wildlife and snow sports: Prioritizing areas for winter refuges. Ecol. Appl 2011, 21, 955–967. [Google Scholar]
- Faraway, J. Practical Regression and Anova Using R. 2002. Available online: http://cran.r-project.org/doc/contrib/Faraway-PRA.pdf (accessed on 15 September 2012).
- Phillips, S. A Brief Tutorial on Maxent; 2005; Available online: www.cs.princeton.edu/~schapire/-maxent/tutorial/tutorial.doc (accessed on 15 September 2012).
- Breiman, L. Random forests. Mach. Learn 2001, 45, 5–32. [Google Scholar]
- Morgan, J.; Sonquist, J. Problem in the analysis of survey data, and a proposal. J. Am. Statist. Assoc 1963, 58, 415–434. [Google Scholar]
- Breidenbach, J.; Nothdurft, A.; Kändler, G. Comparison of nearest neighbour approaches for small area estimation of tree species-specific forest inventory attributes in central Europe using airborne laser scanner data. Eur. J. Forest Res 2010, 129, 833–846. [Google Scholar]
- Crookston, N.; Finley, A. yaImpute: An R Package for k-NN Imputation. J. Statist. Softw 2008, 23, 1–16. [Google Scholar]
- Strobl, C.; Boulesteix, A.; Kneib, T.; Augustin, T.; Zeileis, A. Conditional variable importance for random forests. BMC Bioinf 2008. [Google Scholar] [CrossRef]
- Landis, J.; Koch, G. The measurement of observer agreement for categorical data. Biometrics 1977, 33, 159–174. [Google Scholar]
- Boyce, M.; Vernier, P.; Nielsen, S.; Schmiegelow, F. Evaluating resource selection functions. Ecol. Model 2002, 157, 281–300. [Google Scholar]
- Phillips, S.; Dudik, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar]
- Richardson, A.; Berlyn, G. Changes in foliar spectral reflectance and chlorophyll fluorescence of four temperate species following branch cutting. Tree Physiol 2002, 22, 499–506. [Google Scholar]
- Carter, G. Responses of leaf spectral reflectance to plant stress. Am. J. Bot 1993, 80, 239–243. [Google Scholar]
- Carter, G. Primary and secondary effects of water content on the spectral reflectance of leaves. Am. J. Bot 1991, 78, 916–924. [Google Scholar]
- Lichtenthaler, H.; Rinderle, U. The role of chlorophyll fluorescence in the detection of stress conditions in plants. CRC Critical Rev. Anal. Chem 1988, 19, 29–85. [Google Scholar]
- Zwiggelaar, R. A review of spectral properties of plants and their potential use for crop/weed discrimination on row-crops. Crop Protect 1998, 17, 189–206. [Google Scholar]
- Stockburger, K.; Mitchell, A. Photosynthetic Pigments: A Bibliography; BC-X-383; Information Report; Canadian Forest Service, Pacific Forestry Centre: Victoria, BC, Canada, 1999; pp. 1–30. [Google Scholar]
- Mohammed, G.; Noland, T.; Irving, D.; Sampson, P.; Zarco-Tejada, P.; Miller, J. Natural and Stress-Induced Effects on Leaf Spectral Reflectance in Ontario Species; Ontario Forest Research Institute: Marie, ON, Canada, 2000; pp. 1–34. [Google Scholar]
- Schelling, K.; Schulthess, U. Abschaetzung des Chlorophyllgehaltes von Pflanzenbestaenden mit RapidEye Satellitenbilddaten. Proceedings of Precision Agriculture Reloaded; Informationsgestuetzte Landwirtschaft. 30 GIL-Jahrestagung, Stuttgart, Germany, 24–25 February 2010; pp. 167–170.
- Bonneau, L.; Shields, K.; Civco, D. Using satellite images to classify and analyze the health of hemlock forests infested by the hemlock woolly adelgid. Biol. Invasion 1999, 1, 255–267. [Google Scholar]
- Niemann, K.; Visintini, F. Assessment of Potential for Remote Sensing Detection of Bark Beetle-Infested Areas during Green Attack: A Literature Review; Natural Resources Canada, Canadian Forest Service: Victoria, BC, Canada, 2005; pp. 1–14. Available online: http://cfs.nrcan.gc.ca/pubwarehouse/pdfs/25269.pdf (accessed on 15 September 2012).
- Barsch, H.; Steinhardt, U.; Soellner, R. The development of forest damage—Control and prognosis on the basis of remote sensing data. GeoJournal 1994, 32, 39–46. [Google Scholar]
- McDonald, K.; Zimmermann, R.; Way, J. An Investigation of the Relationship between Tree Water Potential and Dielectric Constant. Proceedings of the IEEE International Geoscience and Remote Sensing Symposium IGARSS’92, Houston, TX, USA, 26–29 May 1992; pp. 523–525.
- Hoekman, D. Radar backscattering of forest stands. Int. J. Remote Sens 1985, 6, 325–343. [Google Scholar]
- Hickey, J.; Birnie, R.; Zhao, M. Oleoresin, Chemistry and Spectral Reflectance in Stressed Lodgepole and White Bark Pine, Mammoth Mountain, California. Summary of the Tenth Annual JPL Airborne Earth Science Workshop, Pasadena, CA, USA, 27 February–2 March 2001; pp. 1–10.
- Hildebrandt, G. Fernerkundung und Luftbildmessung: Für Forstwirtschaft, Vegetationskartierung und Landschaftsoekologie; Wichmann: Heidelberg, Germany, 1996. [Google Scholar]
- Ranson, K.; Kovacs, K.; Sun, G.; Kharuk, V. Disturbance recognition in the boreal forest using radar and Landsat-7. Can. J. Remote Sens 2003, 29, 271–285. [Google Scholar]
- Wulder, M.; White, J.; Carroll, A.; Coops, N. Challenges for the operational detection of mountain pine beetle green attack with remote sensing. For. Chron 2009, 85, 32–38. [Google Scholar]
- Carter, G.; Knapp, A. Leaf optical properties in higher plants: linking spectral characteristics to stress and chlorophyll concentration. Am. J. Bot 2001, 88, 677–684. [Google Scholar]
- Xie, Y.; Sha, Z.; Yu, M. Remote sensing imagery in vegetation mapping: a review. J. Plant Ecol 2008, 1, 9–23. [Google Scholar]
- Guisan, A.; Zimmermann, N.; Elith, J.; Graham, C.; Phillips, S.; Peterson, A. What matters for predicting the occurrences of trees: techniques, data, or species characteristics? Ecol. Monogr 2007, 77, 615–630. [Google Scholar]
- Meynard, C.; Quinn, J. Predicting species distributions: a critical comparison of the most common statistical models using artificial species. J. Biogeogr 2007, 34, 1455–1469. [Google Scholar]
- Renner, I.; Warton, D. Equivalence of MAXENT and poisson point process models for species distribution modeling in ecology. Biometrics 2013, 69, 274–281. [Google Scholar]
Appendix


{kind=link}
{kind=link}
{kind=link}
| RapidEye | TerraSAR-X | |
|---|---|---|
| Acquisition date | 25 May 09 | 24 May 09 |
| Extent | 25 by 25 km (41% coverage) | 10 by 10 km |
| Orbital direction | Descending | Descending |
| Incidence angle | 6.7° | 37.5° |
| Illumination azimuth angle | 175.5° | --- |
| Illumination elevation angle | 62.9° | --- |
| Pixel size | 5 m | 2 m |
| Spatial resolution | 6.5 m | Slant range resolution: 1.18 m |
| Azimuth resolution: 2.05 m | ||
| Spectral coverage | Blue: 410–510 nm | X-band |
| Green: 520–590 nm | --- | |
| Red: 630–690 nm | --- | |
| Red-edge: 690–730 nm | --- | |
| Near infrared: 760–850 nm | --- | |
| Radiometric resolution | 12 bit | 16 bit |
| Index | Formula |
|---|---|
| NDVI [38] | |
| Red-edge Green NDVI [39] | |
| Green NDVI (GNDVI) [40] | |
| Red-edge index (NDRE) [41] | |
| Chlorophyll Green Model (CGM) [42] | |
| Chlorophyll Red-edge Model (CRM) [42] | |
| Red-edge NDVI |
| TerraSAR-X | RapidEye | Multisensor (TerraSAR-X & RapidEye) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ME | RF | GLM | ||||||||||||
| Predicted | Observed | |||||||||||||
| 0 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | |||||
| 0 | 206 | 7 | 220 | 5 | 227 | 4 | 228 | 10 | 228 | 13 | ||||
| 1 | 24 | 8 | 10 | 10 | 3 | 11 | 2 | 5 | 2 | 2 | ||||
| PA (%) | 89.6 | 53.3 | 95.7 | 66.7 | 98.7 | 73.3 | 99.1 | 33.3 | 99.1 | 13.3 | ||||
| UA (%) | 96.7 | 25.0 | 97.8 | 50.0 | 98.3 | 78.6 | 95.8 | 71.4 | 94.6 | 50.0 | ||||
| OA (%) | 87.3 | 93.9 | 97.1 | 95.1 | 93.9 | |||||||||
| AUC | 0.70 | 0.80 | 0.80 | 0.66 | 0.56 | |||||||||
| Kappa | 0.28 | 0.5 | 0.74 | 0.43 | 0.19 | |||||||||
| Kappa 95% CI | 0.04–0.5 | 0.3–0.76 | 0.55–0.93 | 0.11–0.74 | (−0.2) −0.58 | |||||||||
| TerraSAR-X | RapidEye | Multisensor | |
|---|---|---|---|
| Variable (Metric) | Contribution (%) | ||
| Backscatter (Q3) | 39.5 | 16.9 | |
| Backscatter (Q2) | 22.9 | ||
| Backscatter (SD) | 18.8 | 14.5 | |
| Backscatter (Q1) | 10.7 | ||
| Backscatter (max) | 8.1 | ||
| Blue band (max and min) | 31.2 | 14.4 | |
| GNDVI (max) | 28 | ||
| NDVI (max) | 17.13 | 39.1 | |
| Red-edge band (min) | 11.9 | 12 | |
| Red-edge NDVI (mean) | 8.3 | ||
| NIR band (min) | 3.3 | 3.1 | |
| Sum | 100 | 100 | 100 |
Share and Cite
Ortiz, S.M.; Breidenbach, J.; Kändler, G. Early Detection of Bark Beetle Green Attack Using TerraSAR-X and RapidEye Data. Remote Sens. 2013, 5, 1912-1931. https://doi.org/10.3390/rs5041912
Ortiz SM, Breidenbach J, Kändler G. Early Detection of Bark Beetle Green Attack Using TerraSAR-X and RapidEye Data. Remote Sensing. 2013; 5(4):1912-1931. https://doi.org/10.3390/rs5041912
Chicago/Turabian StyleOrtiz, Sonia M., Johannes Breidenbach, and Gerald Kändler. 2013. "Early Detection of Bark Beetle Green Attack Using TerraSAR-X and RapidEye Data" Remote Sensing 5, no. 4: 1912-1931. https://doi.org/10.3390/rs5041912
APA StyleOrtiz, S. M., Breidenbach, J., & Kändler, G. (2013). Early Detection of Bark Beetle Green Attack Using TerraSAR-X and RapidEye Data. Remote Sensing, 5(4), 1912-1931. https://doi.org/10.3390/rs5041912