Response of Spectral Reflectances and Vegetation Indices on Varying Juniper Cone Densities


 ,
,
Abstract
:1. Introduction
2. Methods
2.1. Acquisition of Juniper Canopy Spectra
2.2. Analysis of Reflectances and Vegetation Indices
2.3. Variation Analysis of Juniper Canopy Spectra and Vegetation Indices
3. Results and Discussion
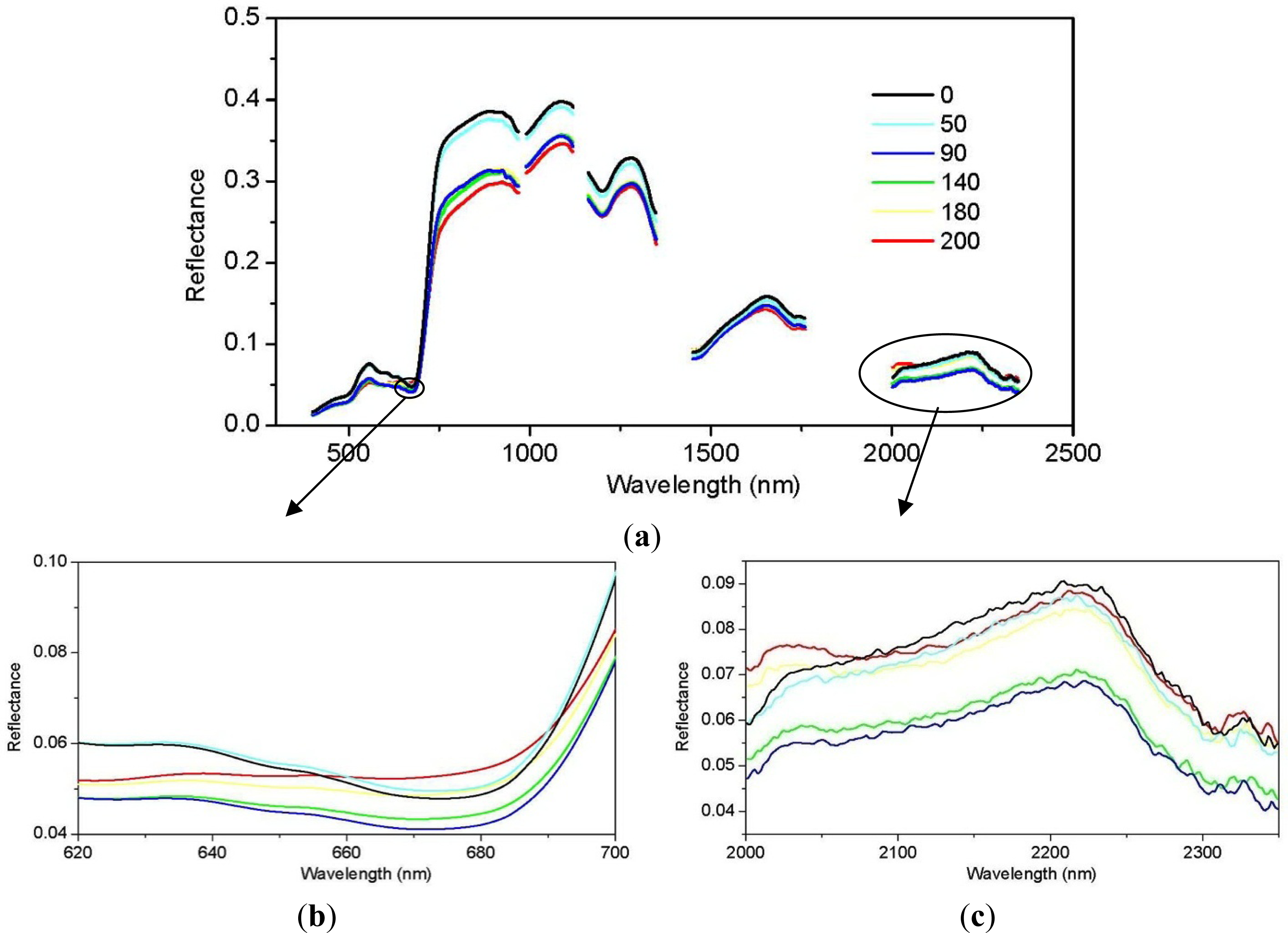
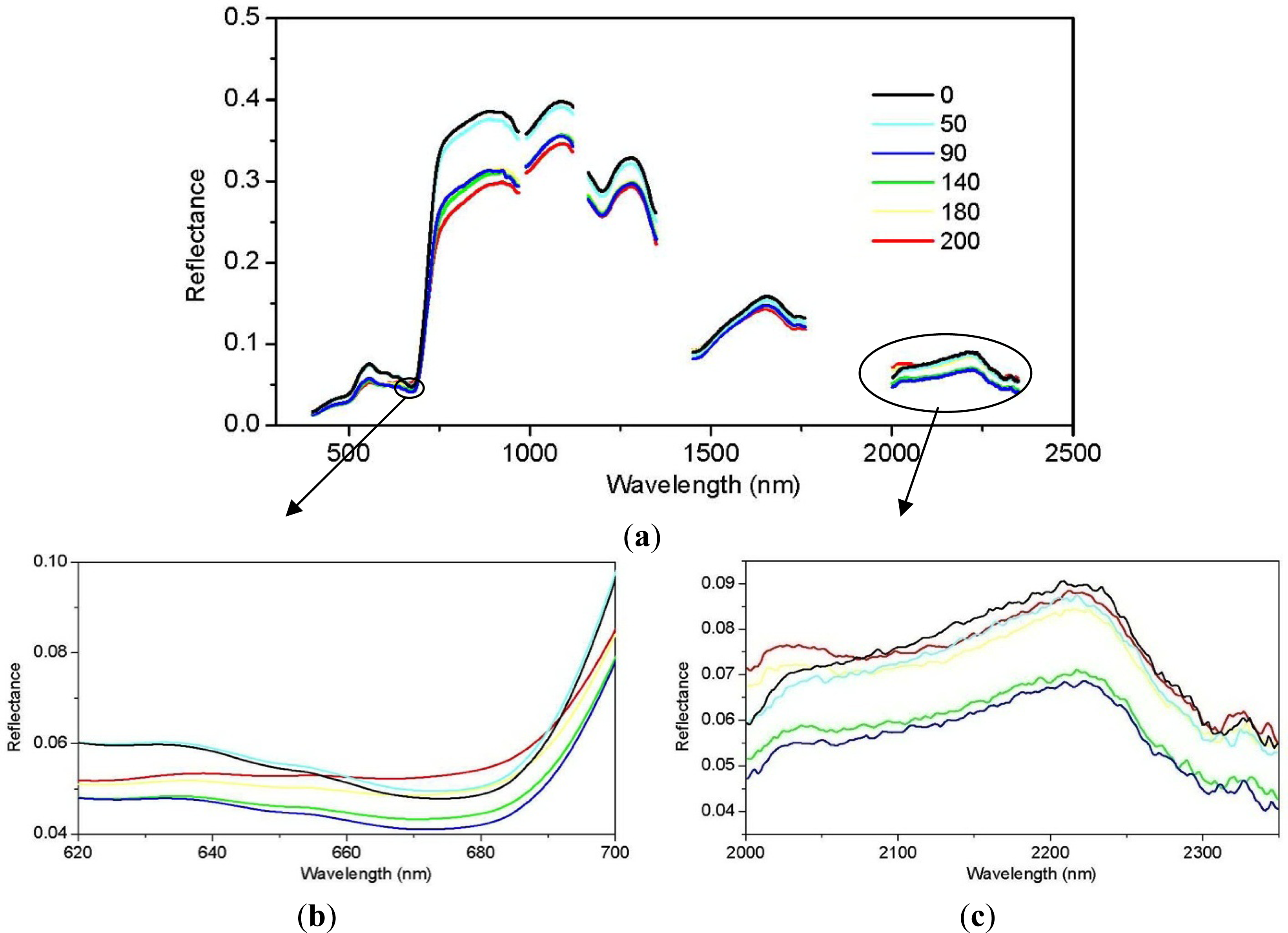
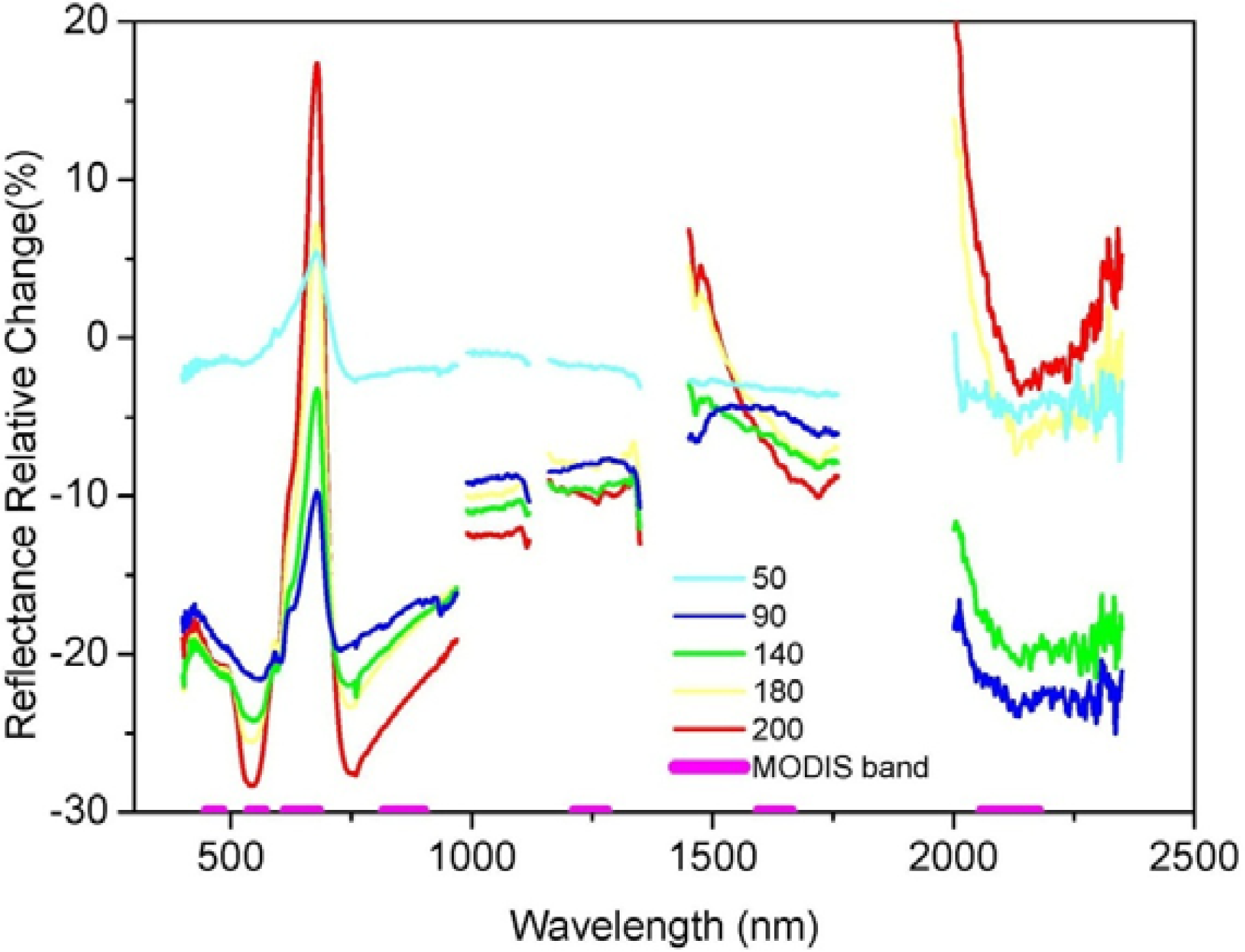
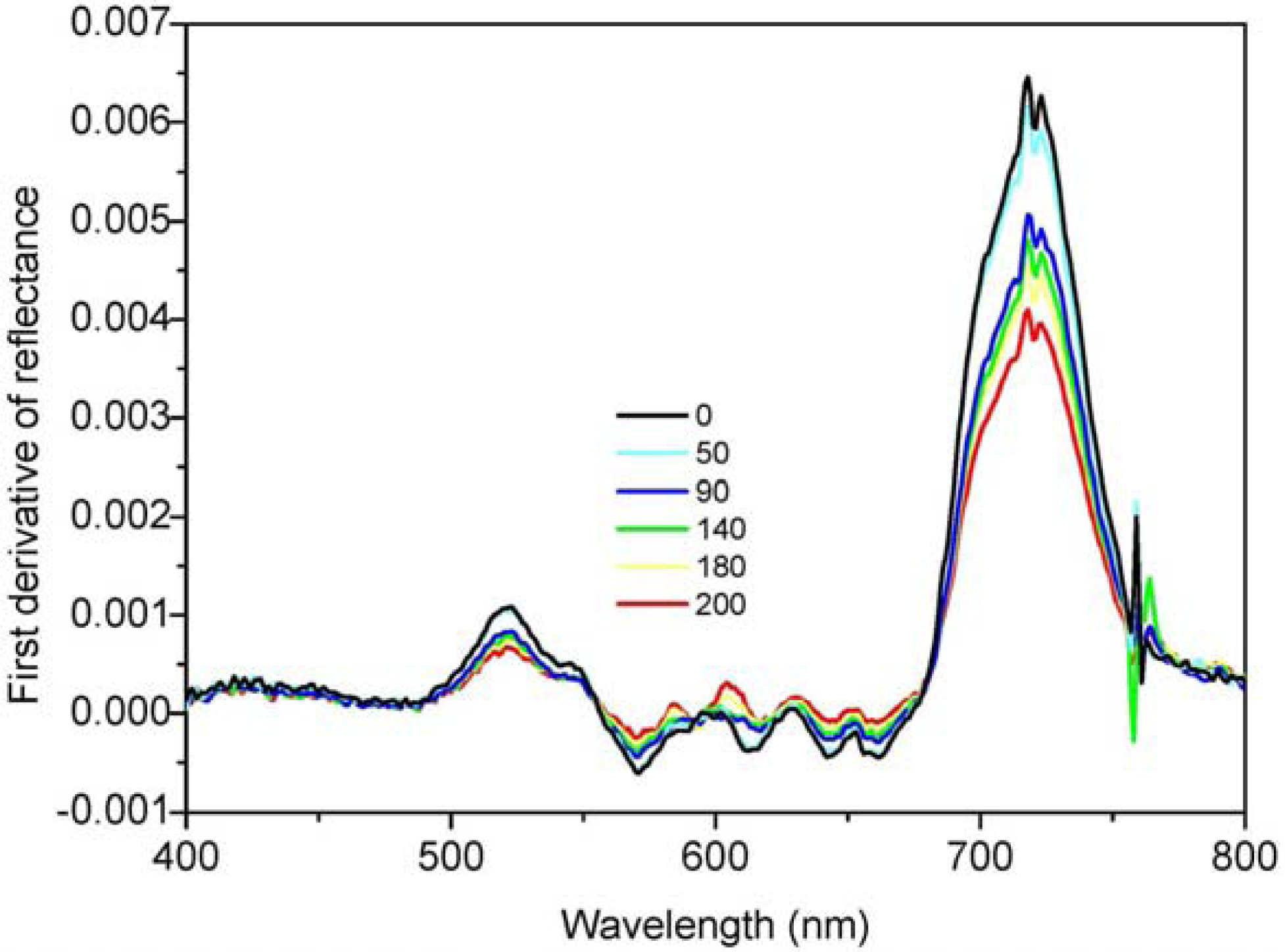
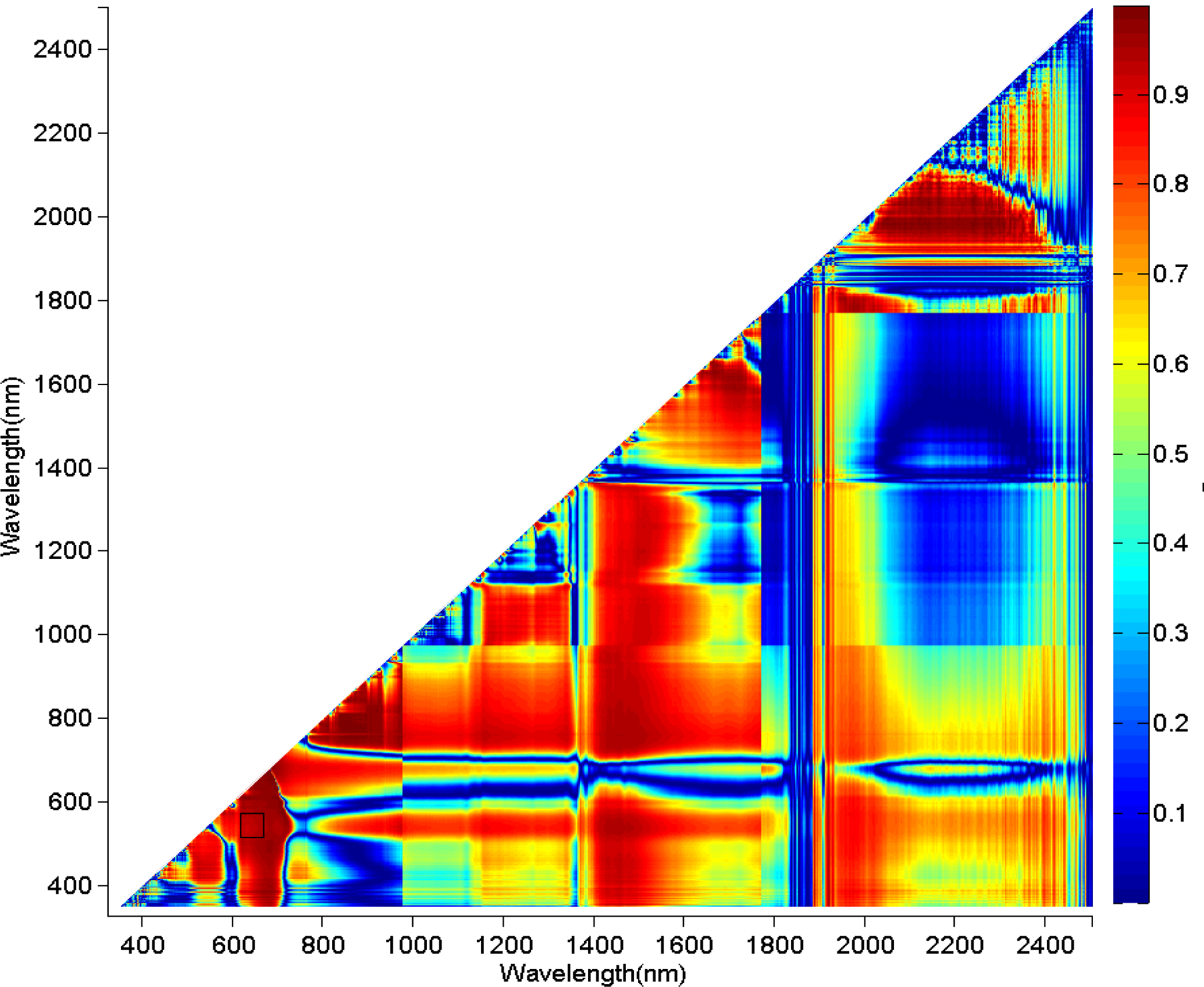
3.1. Juniper Canopy Spectral Signatures with Varying Cone Densities
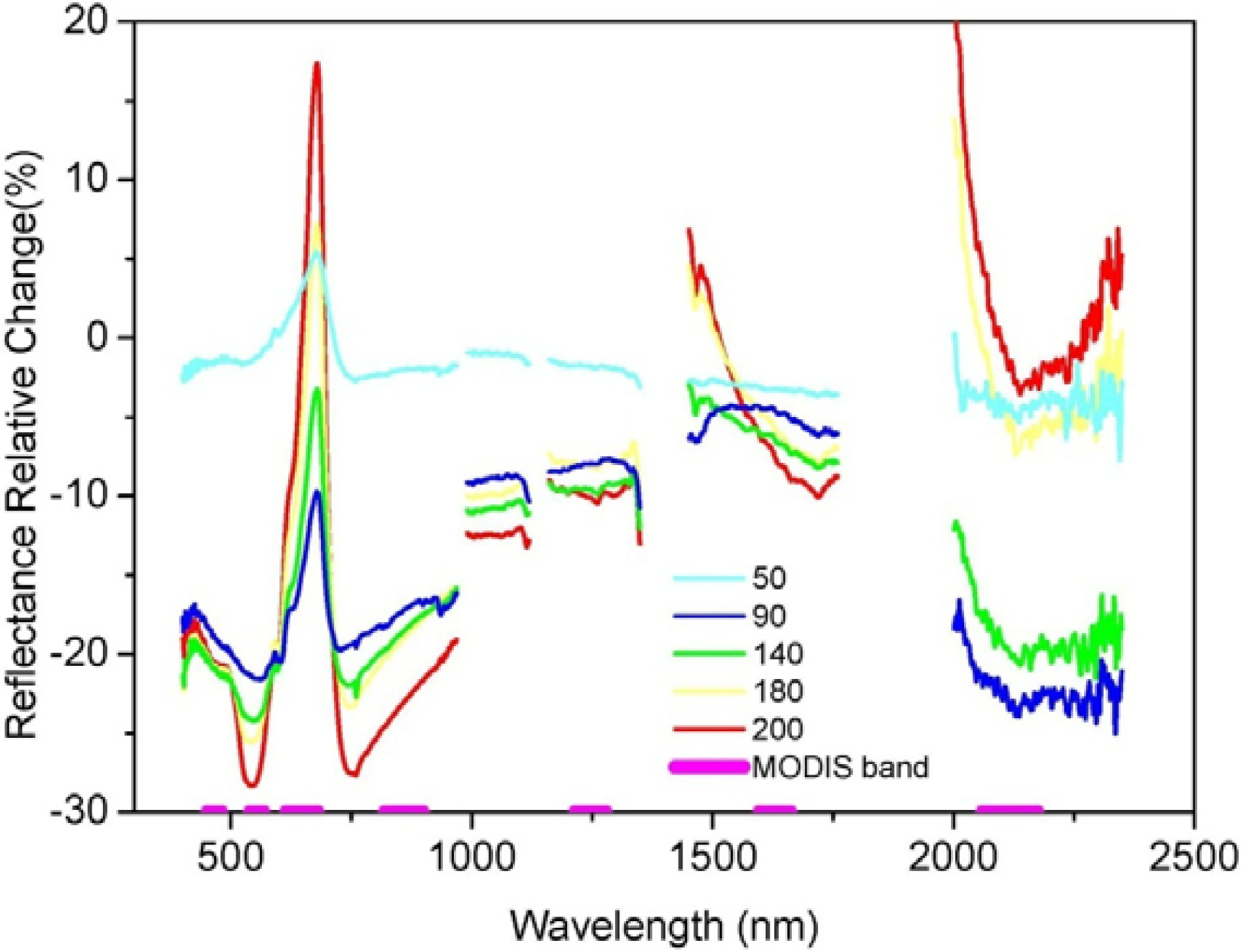
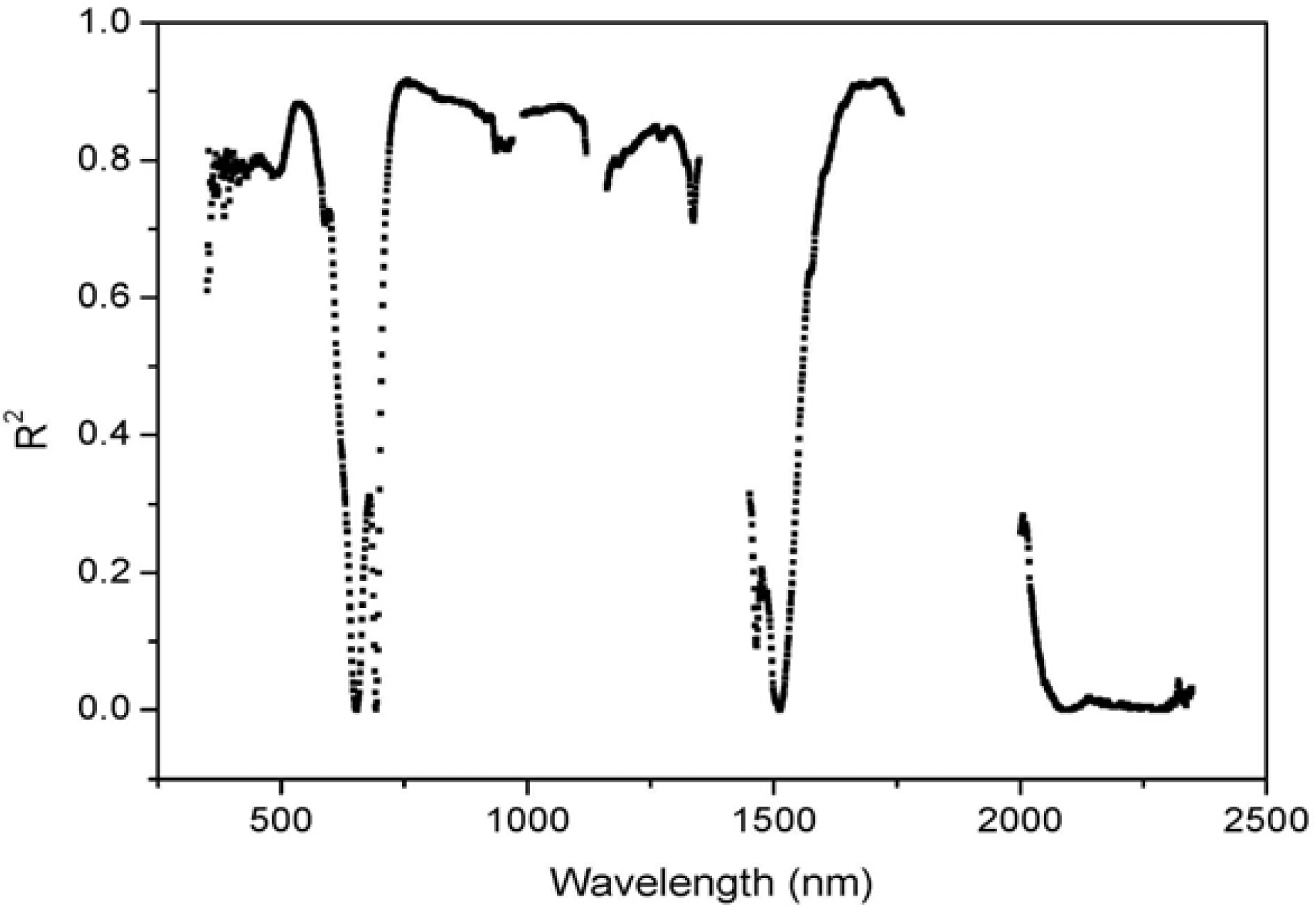
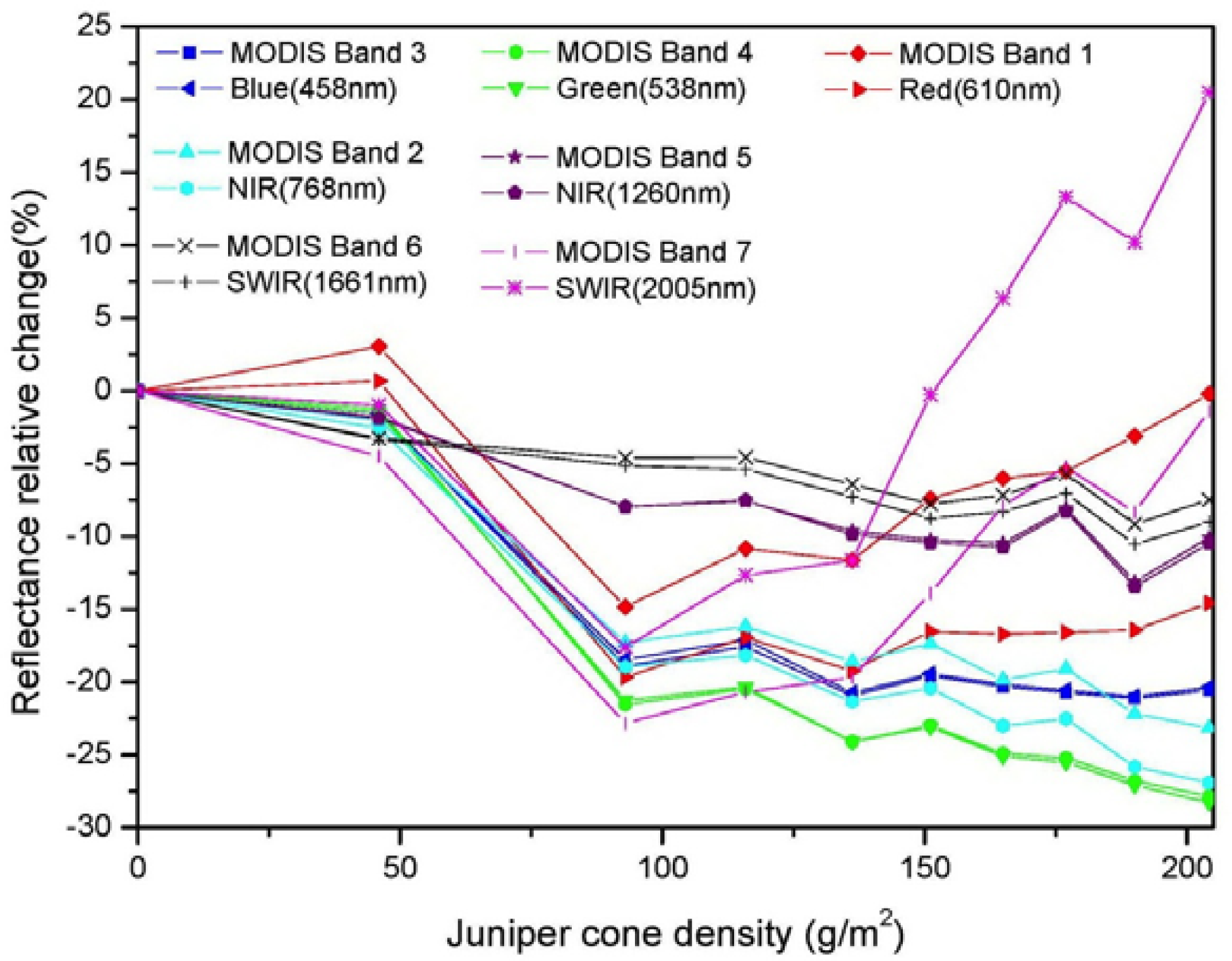
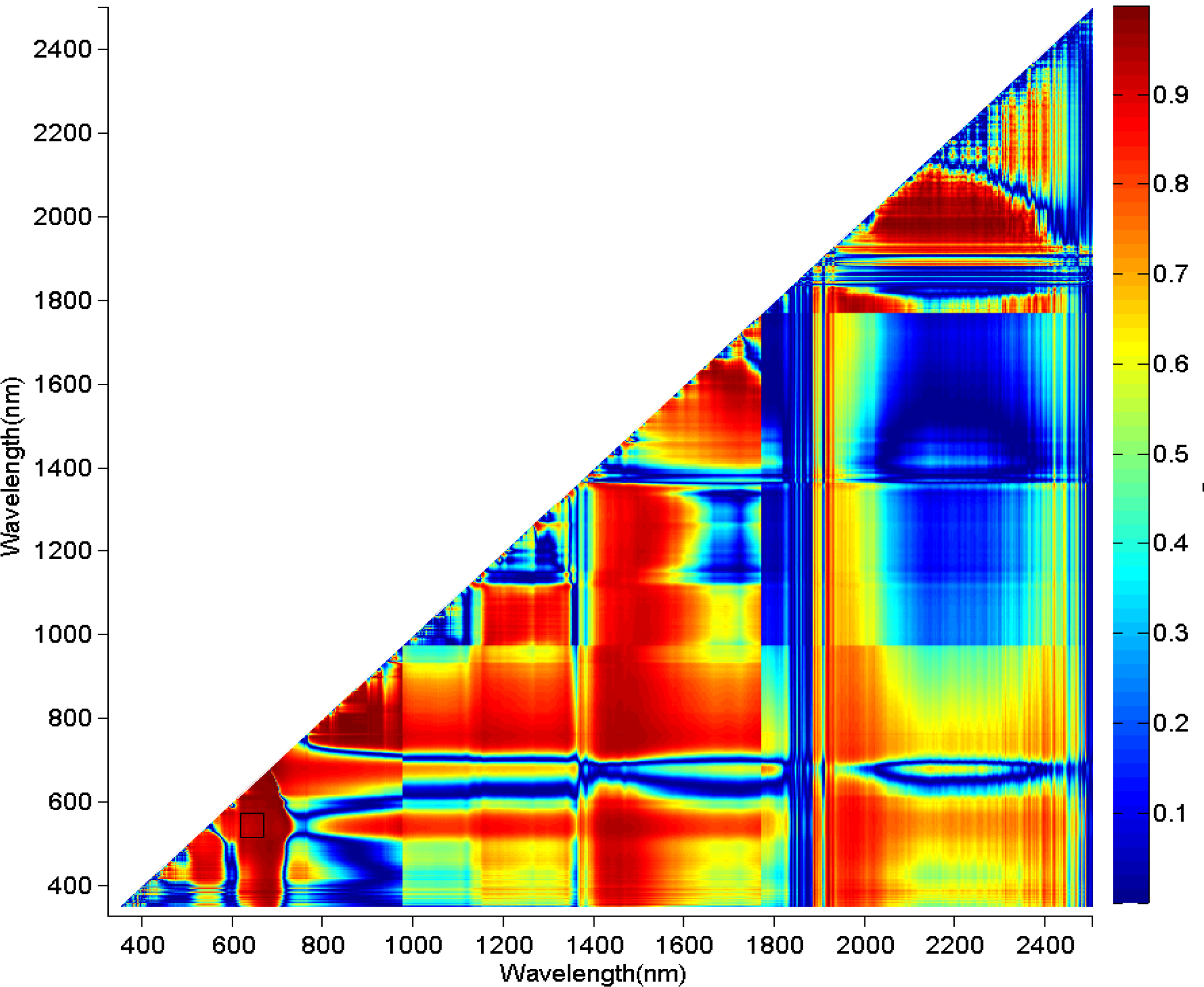
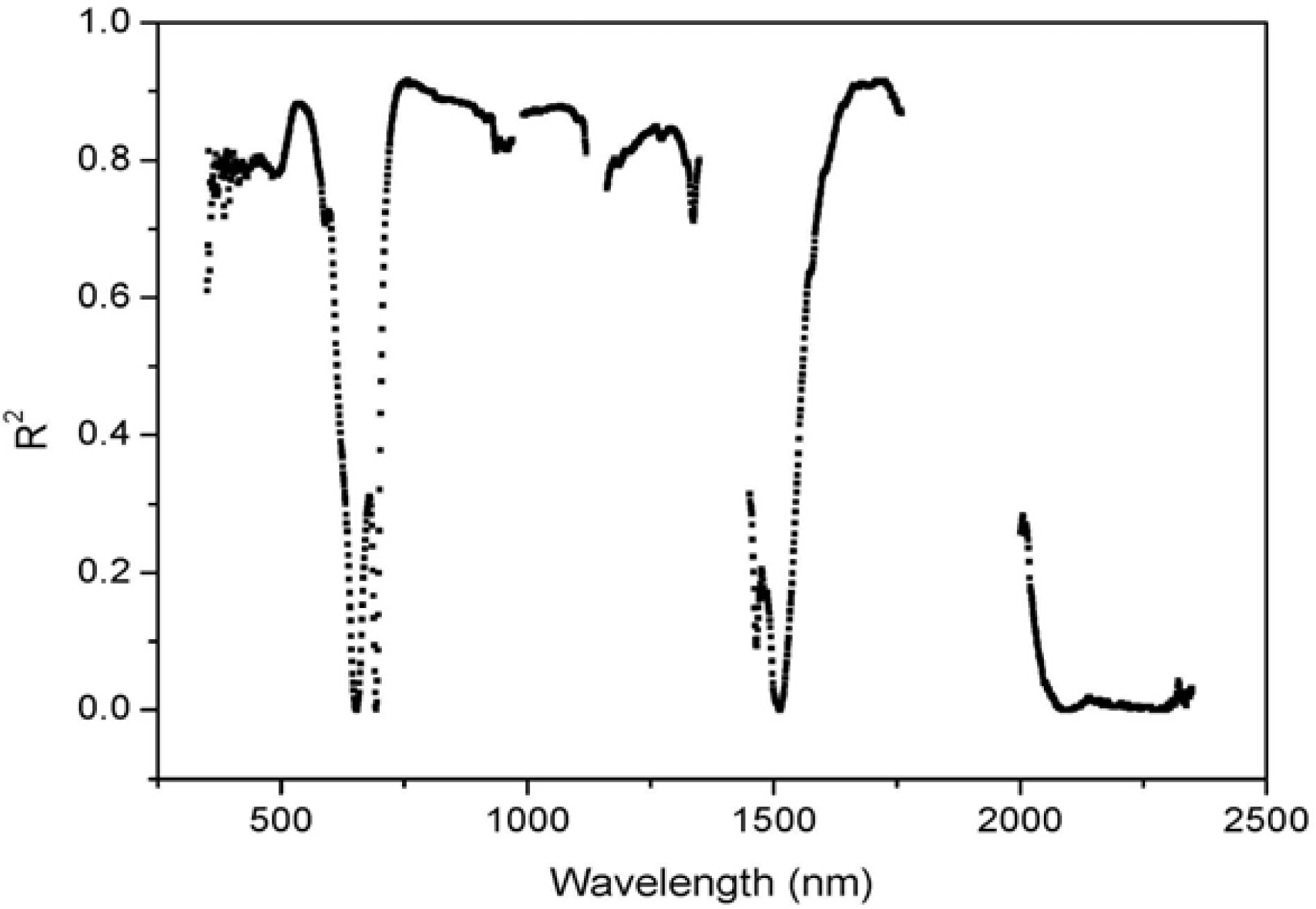
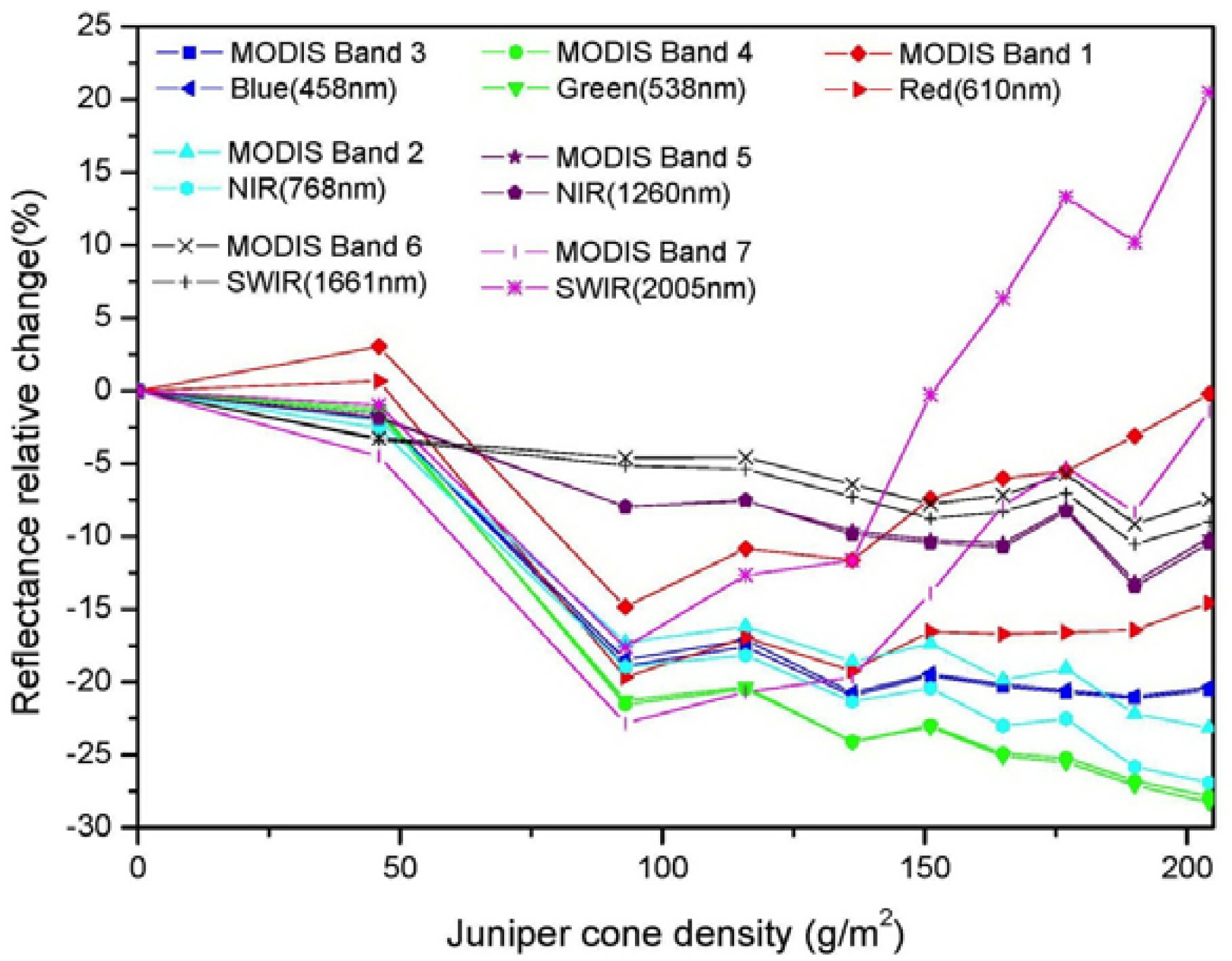
3.2. Comparisons of Narrow- and Broadband Sensitivity to Cone Density Variations
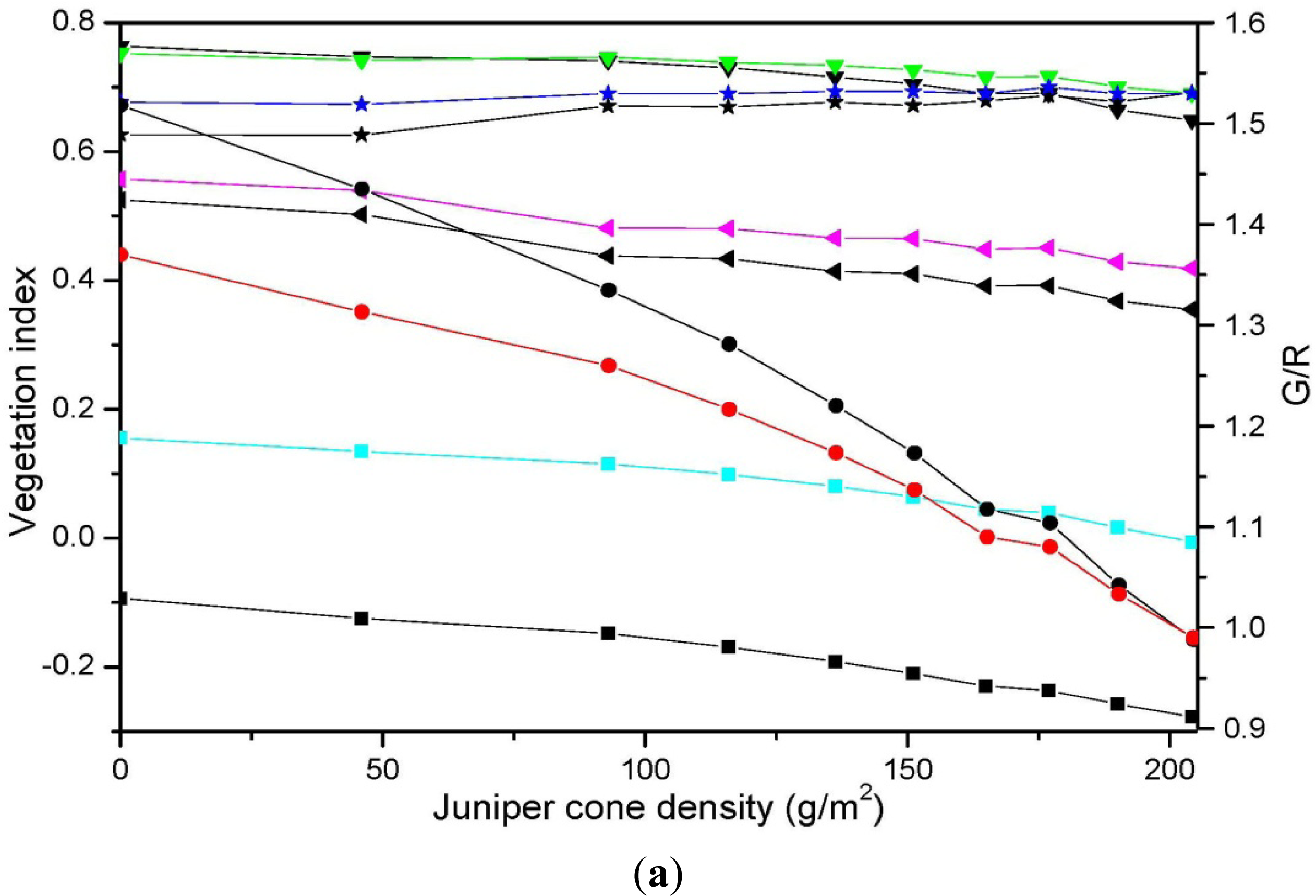
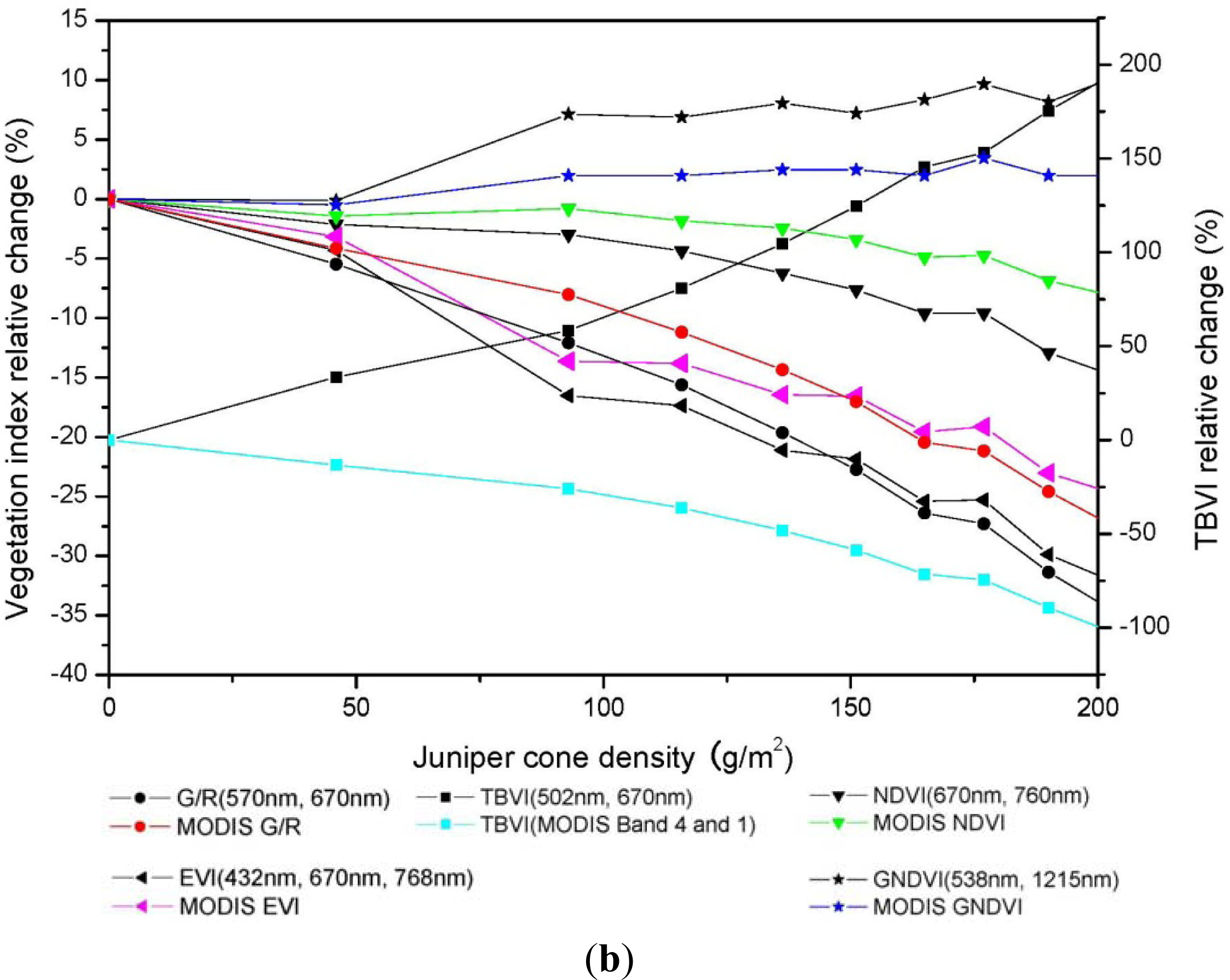
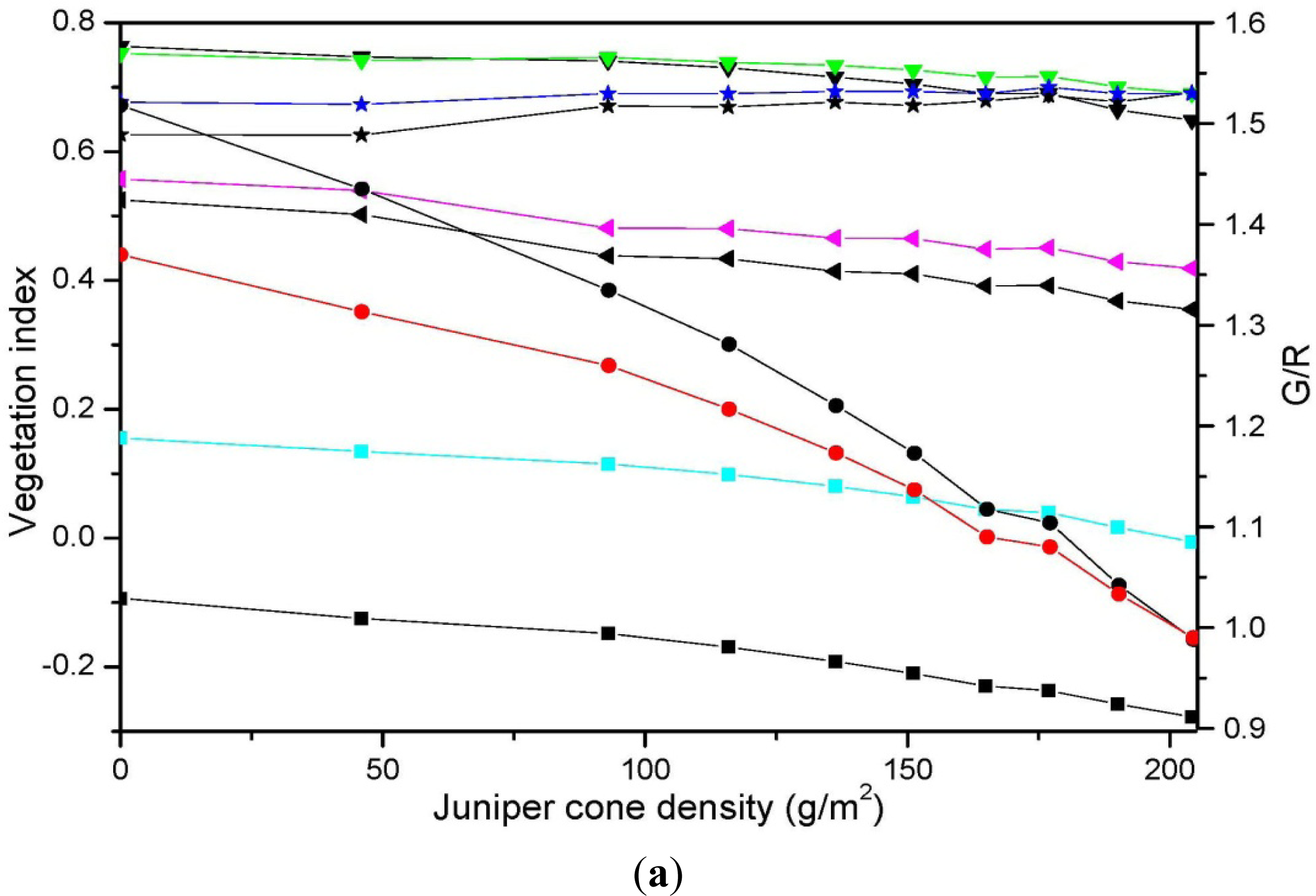
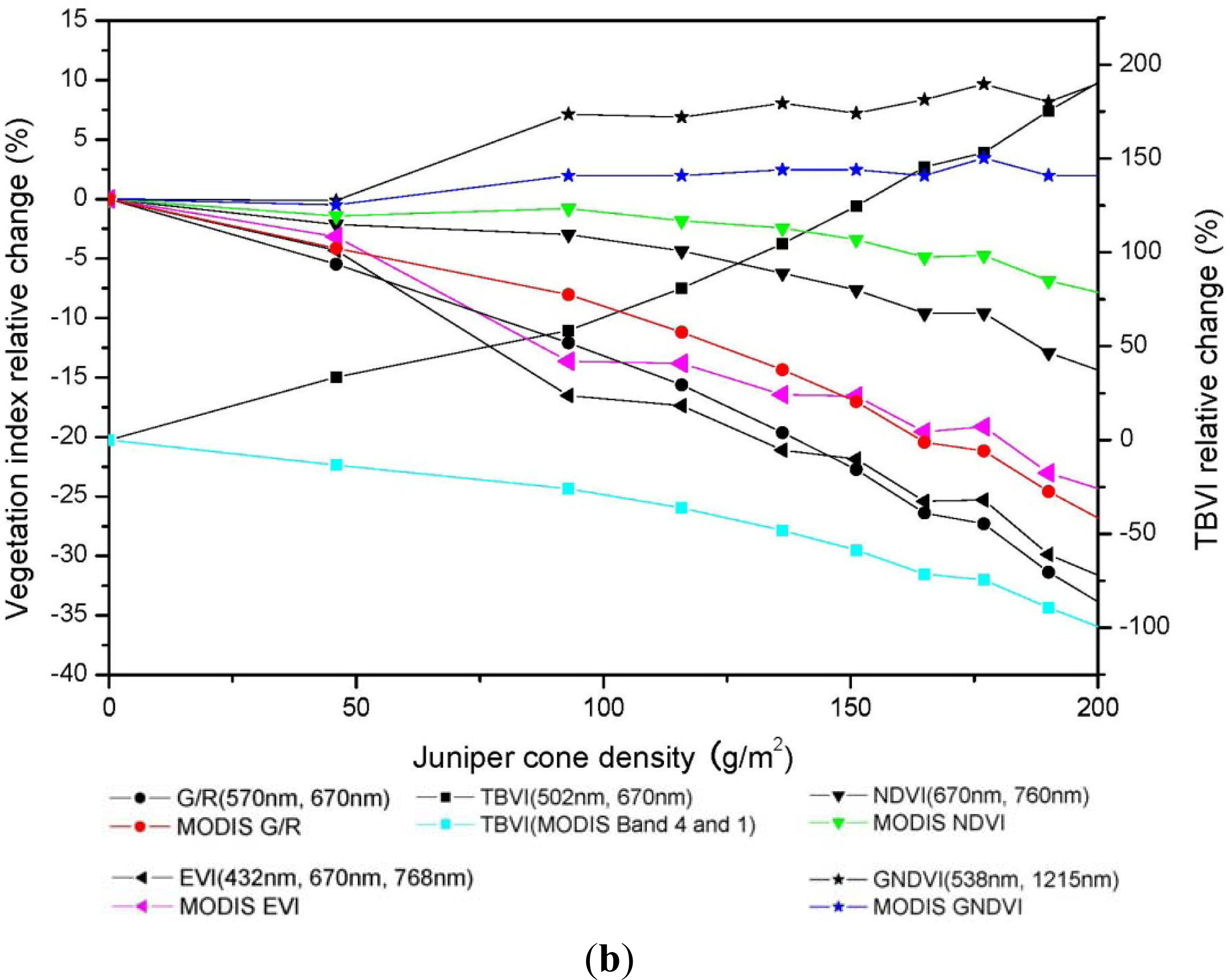
3.3. Relationship between Vegetation Indices and Juniper Cone Densities
3.4. Future Studies
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Brenzel, K.N. Sunset Western Garden Book; Oxmoor House: Birmingham, AL, USA, 1995; pp. 606–607. [Google Scholar]
- Steven, L.P.; Tamzen, K.S. Development of GIS-based models to predict plant community structure in relation to western juniper establishment. For. Ecol. Manage 2008, 256, 981–989. [Google Scholar]
- Gedney, D.R.; Azuma, D.L.; Bolsinger, C.L.; McKay, N. Western Juniper in Eastern Oregon; General Technical Report PNW-GTR-464; United States Department of Agriculture: Twin Cities, MN, USA, 1999. [Google Scholar]
- Miller, R.F.; Bates, J.D.; Svejcar, T.J.; Pierson, F.B.; Eddleman, L.E. Biology, Ecology, And Management of Western Juniper; Technical Bulletins 152; Agricultural Experiment Station, Oregon State University: Corvallis, OR, USA, 2005. [Google Scholar]
- José Carlos, M.R. Diversity of maritime juniper woodlands. For. Ecol. Manage 2004, 192, 267–276. [Google Scholar]
- Alice, C.M. Use of pinyon-juniper woodlands by bats in New Mexico. For. Ecol. Manage 2005, 204, 209–220. [Google Scholar]
- Johnsona, R.M.; Barnesa, C.; Dhar, M.; Portnoy, J. Increase in midwestern airborne juniper pollen over 13 years. J. Allergy Clin. Immun 2010, 125, 25. [Google Scholar]
- Behrens, T.; Muller, J.; Diepenbrock, W. Utilization of canopy reflectance to predict properties of oilseed rape (Brassica napus L.) and barley (Hordeumvalgare L.) during ontogenesis. Eur. J. Agoron 2006, 25, 345–355. [Google Scholar]
- Ge, S.; Everitt, J.; Carruthers, R.; Gong, P.; Anderson, G. Hyperspectral characteristics of canopy components and structure for phonological assessment of an invasive weed. Environ. Monit. Assess 2006, 120, 109–126. [Google Scholar]
- Peters, A.J.; Reed, B.C.; Eve, M.D.; Mcdaniel, K.C. Remote sensing of broom snakeweed (Gutierreziasarothrae) with NOAA-10 spectral image processing. Weed Technol 1992, 6, 1015–1020. [Google Scholar]
- Verma, K.S.; Saxena, R.K.; Hajare, T.N.; Kharche, V.K.; Kumari, P.A. Spectral response of gram varieties under variable soil conditions. Int. J. Remote Sens 2002, 23, 313–324. [Google Scholar]
- Shen, M.; Chen, J.; Zhu, X.; Tang, Y. Yellow flowers can decrease NDVI and EVI values: Evidence from a field experiment in an alping meadow. Can. J. Remote Sens 2009, 35, 99–106. [Google Scholar]
- Peng, D.; Huete, A.R.; Huang, J.; Wang, F.; Sun, H. Detection and estimation of mixed paddy rice cropping patterns with MODIS data. Int. J. Appl. Earth Obs. Geoinf 2011, 13, 13–23. [Google Scholar]
- Huete, A.R.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ 2002, 83, 195–213. [Google Scholar]
- Thenkabail, P.S.; Smith, R.B.; De-Pauw, E. Evaluation of narrowband and broadband vegetation indices for determining optimal hyperspectral wavebands for agricultural crop characterization. Photogramm. Eng. Rem. Sens 2002, 68, 607–621. [Google Scholar]
- Thenkabail, P.S.; Smith, R.B.; De-Pauw, E. Hyperspectral vegetation indices for determining agricultural crop characteristics. Remote Sens. Environ 2000, 71, 158–182. [Google Scholar]
- Thenkabail, P.S.; Mariotto, I.; Gumma, M.K.; Middleton, E.M.; Landis, D.R.; Huemmrich, K.F. Selection of hyperspectralnarrowbands (HNBs) and composition of hyperspectraltwoband vegetation indices (HVIs) for biophysical characterization and discrimination of crop types using field reflectance and Hyperion/EO-1 data. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens 2013, 6, 427–439. [Google Scholar]
- Rouse, W.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring Vegetation Systems in the Great Plains with ERTS; NASA SP-351; NASA: Washington, DC, USA, 1974. [Google Scholar]
- Jiang, Z.Y.; Huete, A.R.; Didan, K.; Miura, T. Development of a two-band enhanced vegetation index without a blue band. Remote Sens. Environ 2008, 112, 3833–3845. [Google Scholar]
- Gitelson, A.A.; Kaufman, Y.J.; Merzlyak, M.N. Use of a green channel in remote sensing of global vegetation from EOS-MODIS. Remote Sens. Environ 1996, 58, 289–298. [Google Scholar]
- Hunt, E.R., Jr.; Williams, A.E.P. Detection of flowering leafy spurge with satellite multispectral imagery. Rangel. Ecol. Manage 2006, 59, 494–499. [Google Scholar]
- Stone, C.; Chisholm, L.; Coops, N. Spectral reflectance characteristics of eucalypt foliagedamaged by insects. Aust. J. Bot 2001, 49, 687–698. [Google Scholar]
- Carter, G.A.; Knapp, A.K. Leaf optical properties in higher plants: Linking spectral characteristics to stress and chlorophyllconcentration. Am. J. Bot 2001, 88, 677–684. [Google Scholar]
- Skopec, M.M.; Dearing, M.D. Differential expression and activity of catechol-O-methyl transferase (COMT) in a generalist (Neotomaalbigula) and juniper specialist (Neotomastephensi) woodrat. Comp. Biochem. Physiol. C 2011, 154, 383–390. [Google Scholar]
- Landon, D.B.; Peter, V.W.; Harrington, W.; Estelle, L. Predicting and quantifying pollen production in Juniperus Ashei forests. Phytologia 2012, 94, 417–438. [Google Scholar]
- Huang, C.Y.; Asner, G.P.; Martin, R.E.; Barger, N.N.; Neff, J.C. Multiscale analysis of tree cover and aboveground carbon stocks in pinyon-juniper woodlands. Ecol. Appl 2009, 19, 668–681. [Google Scholar]
- William, H.R.; Craig, D.A.; John, D.B.; William, L.B.; Brandon, T.B.; Peter, M.B.; Karen, S.E.; Lisa, M.F.; David, W.H.; Brian, F.J.; et al. Historical and modern disturbance regimes, stand structures, and landscape dynamics in piñon-juniper vegetation of the Western United States. Rangel. Ecol. Manage 2009, 62, 203–222. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Band Ranges | R2 | S | CV |
|---|---|---|---|
| Blue (430–480 nm) | 0.81 *** (458 nm) | 7.1 × 10−4 (480 nm) | 0.10 (480 nm) |
| MODIS Band 3 | 0.80 *** | 6.6 × 10−4 | 0.09 |
| Green (500–570 nm) | 0.88*** (538 nm) | 2.1 × 10−3 (555 nm) | 0.13 (546 nm) |
| MODIS Band 4 | 0.87 *** | 2.0 × 10−3 | 0.12 |
| Red (610–670 nm) | 0.55** (610 nm) | 8.7 × 10−4 (610 nm) | 0.09 (610 nm) |
| MODIS Band 1 | 0.03 | 0.7 × 10−4 | 0.06 |
| NIR (760–898 nm) | 0.91*** (768 nm) | 9.4 × 10−3 (760 nm) | 0.12 (760 nm) |
| MODIS Band 2 | 0.88 *** | 8.5 × 10−3 | 0.09 |
| NIR (1215–1278 nm) | 0.85*** (1,260 nm) | 3.6 × 10−3 (1,262 nm) | 0.05 (1,262 nm) |
| MODIS Band 5 | 0.84 | 3.4 × 10−3 | 0.04 |
| SWIR (1597–1661 nm) | 0.91*** (1,661 nm) | 1.8 × 10−3 (2,001 nm) | 0.13 (2,001 nm) |
| MODIS Band 6 | 0.86 *** | 1.1 × 10−3 | 0.03 |
| SWIR (2000–2174 nm) | 0.28(2,005 nm) | 1.7 × 10−3 (2,000 nm) | 0.13 (2,001 nm) |
| MODIS Band 7 | 0.01 | 0.4 × 10−3 | 0.09 |
| Vegetation Index | R2 | S | CV |
|---|---|---|---|
| Narrowband TBVI | 0.97 *** (502 nm, 670 nm) | 2.5 × 10−2 (538 nm, 670 nm) | 0.99 (547 nm, 638 nm) |
| MODIS TBVI | 0.94 *** (MODIS Band 4 and 1) | 1.9 × 10−2 (MODIS Band 7 and 4) | 0.79 (MODIS Band 7 and 4) |
| Narrowband EVI | 0.98 *** (432 nm, 670 nm, 768 nm) | 1.8 × 10−2 (480 nm, 670 nm, 760 nm) | 0.14 (480 nm, 670 nm, 760 nm) |
| MODIS EVI | 0.97 *** | 1.2 × 10−2 | 0.09 |
| Narrowband NDVI | 0.89 *** (670 nm, 760 nm) | 1.2 × 10−2 (670 nm, 760 nm) | 0.05 (670 nm, 760 nm) |
| MODIS NDVI | 0.76*** | 0.6 × 10−2 | 0.03 |
| Narrowband GNDVI | 0.86 *** (538 nm, 1,215 nm) | 6.7 × 10−3 (552 nm, 1215 nm) | 0.04 (554 nm, 1,215 nm) |
| MODIS GNDVI | 0.76 *** | 2.0 × 10−3 | 0.01 |
| Narrowband G/R | 0.98 *** (570 nm, 670 nm) | 6.3 × 10−2 (555 nm, 670 nm) | 0.15 (546 nm, 670 nm) |
| MODIS G/R | 0.96 *** | 4.0 × 10−2 | 0.11 |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Peng, D.; Jiang, Z.; Huete, A.R.; Ponce-Campos, G.E.; Nguyen, U.; Luvall, J.C. Response of Spectral Reflectances and Vegetation Indices on Varying Juniper Cone Densities. Remote Sens. 2013, 5, 5330-5345. https://doi.org/10.3390/rs5105330
Peng D, Jiang Z, Huete AR, Ponce-Campos GE, Nguyen U, Luvall JC. Response of Spectral Reflectances and Vegetation Indices on Varying Juniper Cone Densities. Remote Sensing. 2013; 5(10):5330-5345. https://doi.org/10.3390/rs5105330
Chicago/Turabian StylePeng, Dailiang, Zhangyan Jiang, Alfredo R. Huete, Guillermo E. Ponce-Campos, Uyen Nguyen, and Jeffrey C. Luvall. 2013. "Response of Spectral Reflectances and Vegetation Indices on Varying Juniper Cone Densities" Remote Sensing 5, no. 10: 5330-5345. https://doi.org/10.3390/rs5105330