Investigation on the Patterns of Global Vegetation Change Using a Satellite-Sensed Vegetation Index


Abstract
:1. Introduction
2. Materials and Methods
2.1. Data and Processing
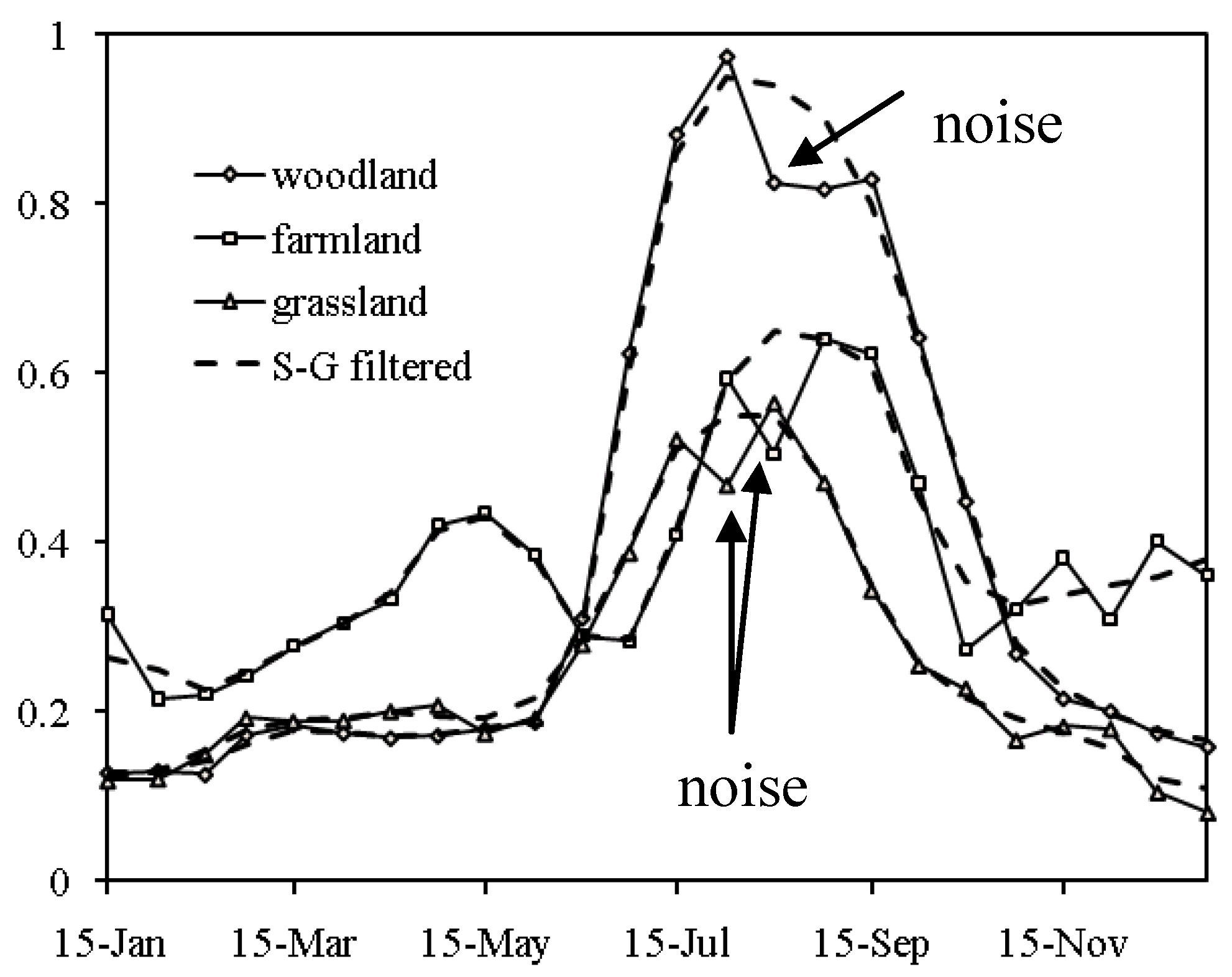
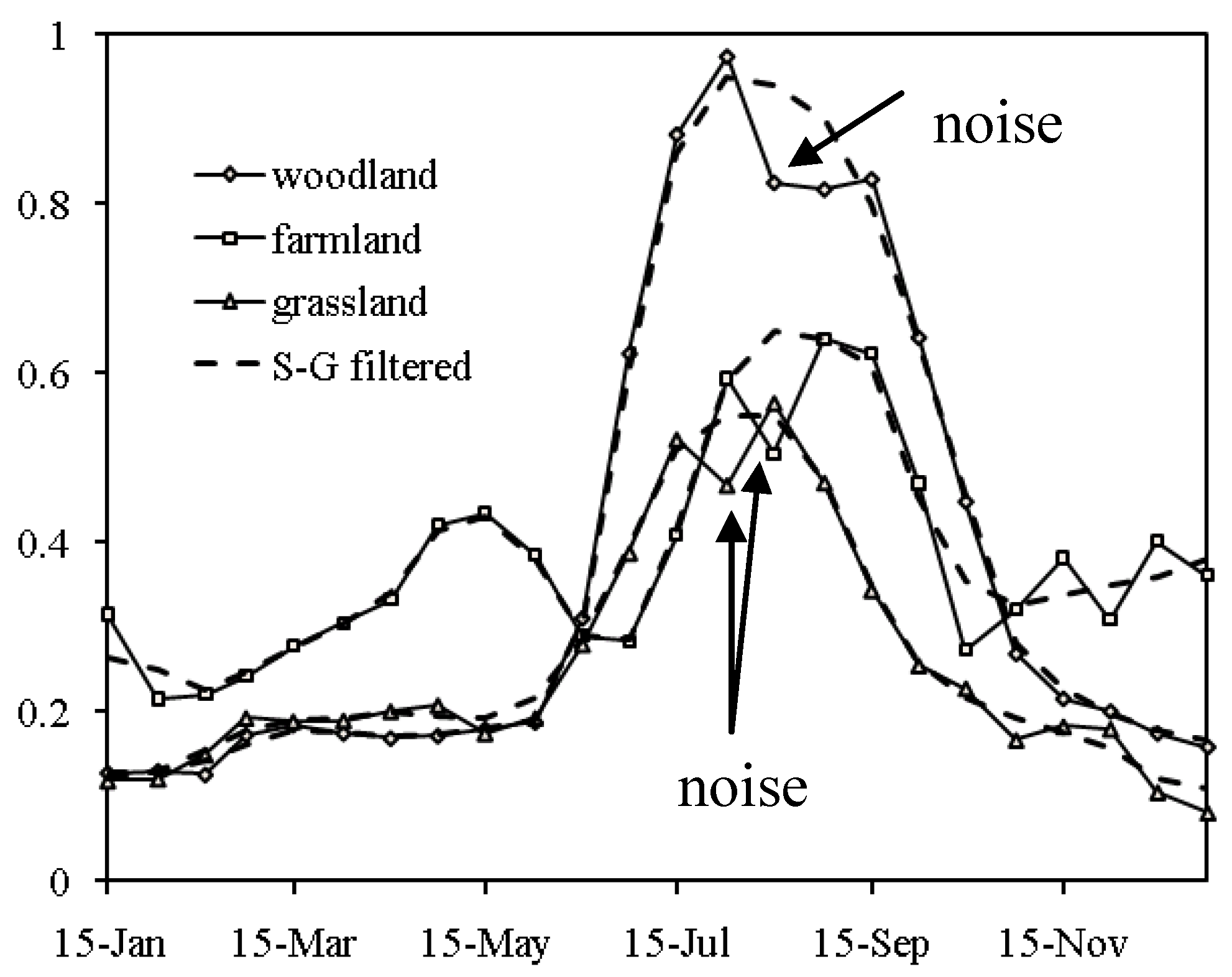
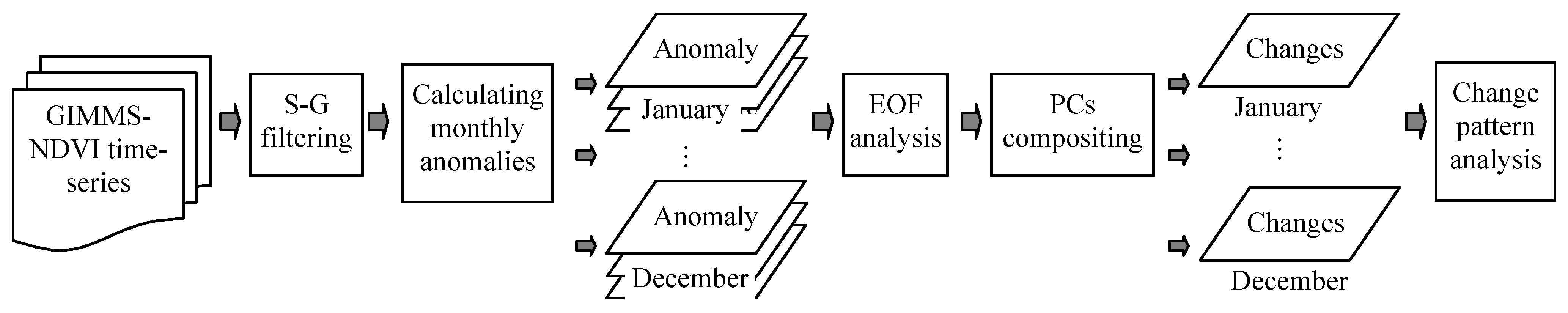
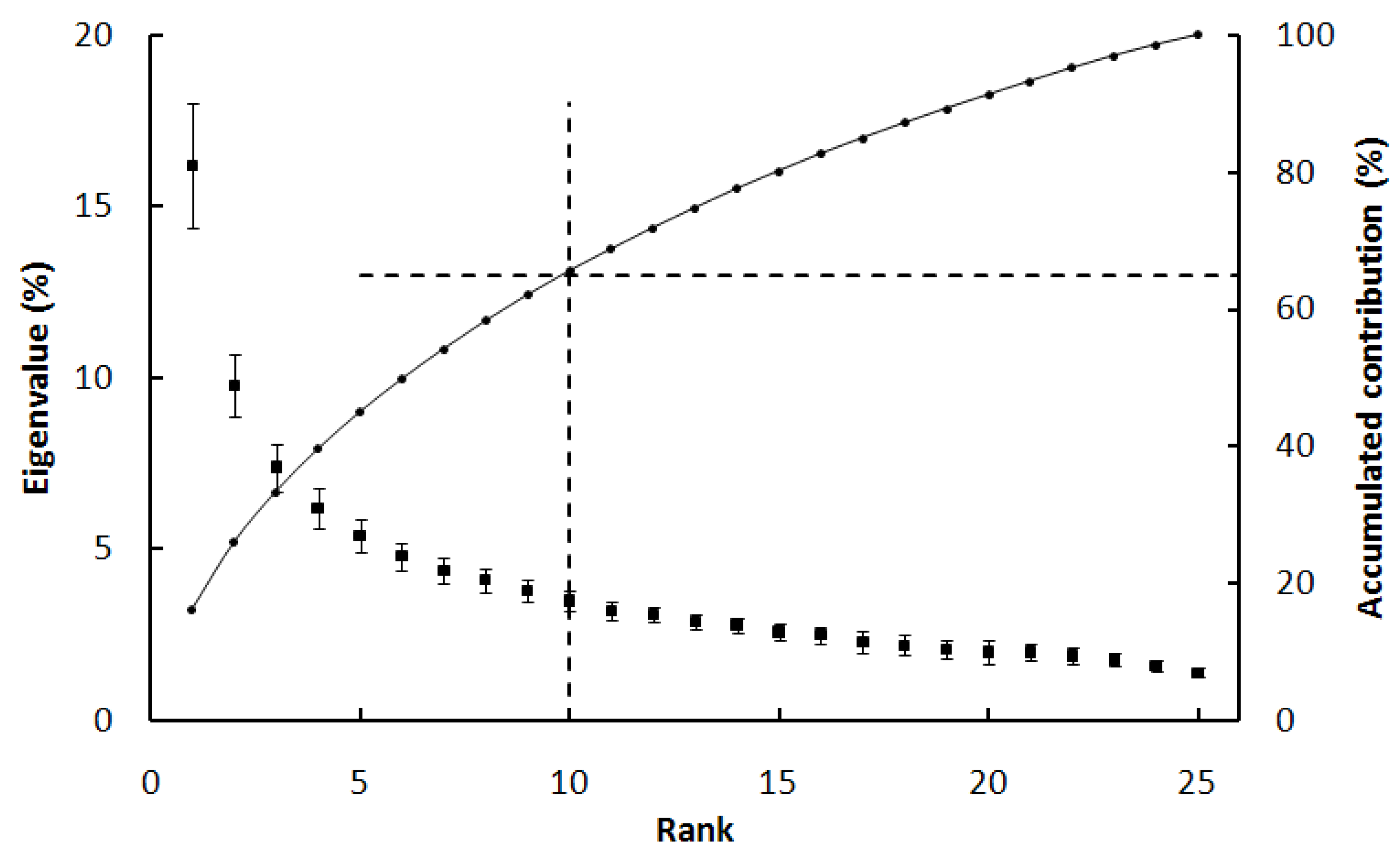
2.2. Extracting the Spatial-Temporal Change by Using an Empirical Orthogonal Function (EOF) Method
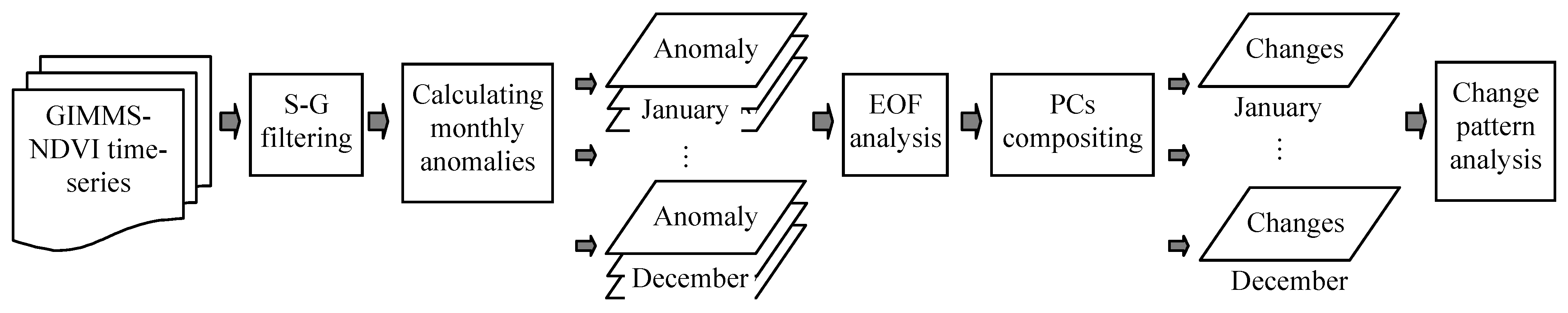
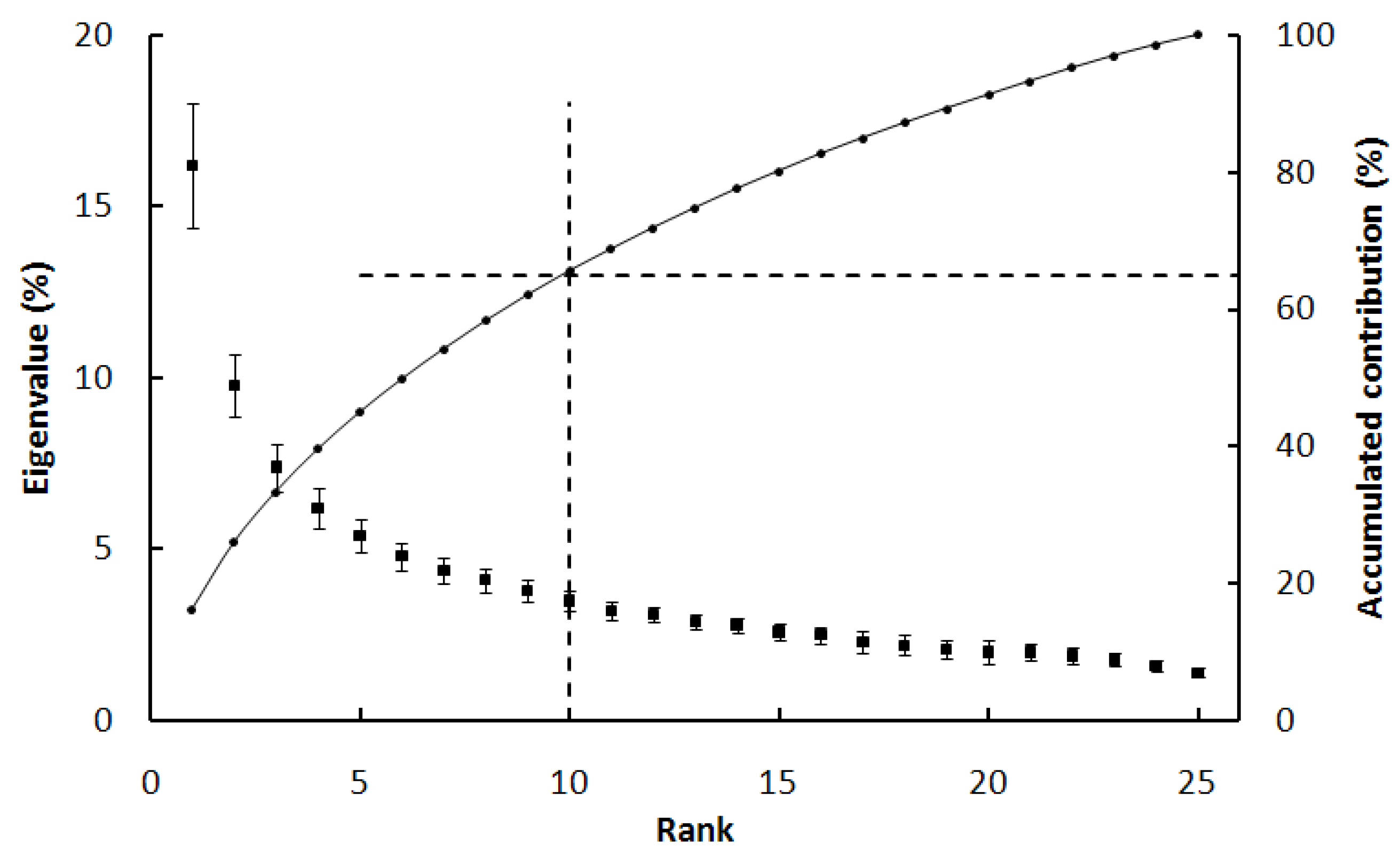
2.3. The Hypotheses of Vegetation Change Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Month | Jan | Feb | Mar | Apr | May | Jun | Jul | Aug | Sep | Oct | Nov | Dec |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| North hemisphere | Winter | Spring | Summer | Autumn | Winter | |||||||
| South hemisphere | Summer | Autumn | Winter | Spring | Summer | |||||||
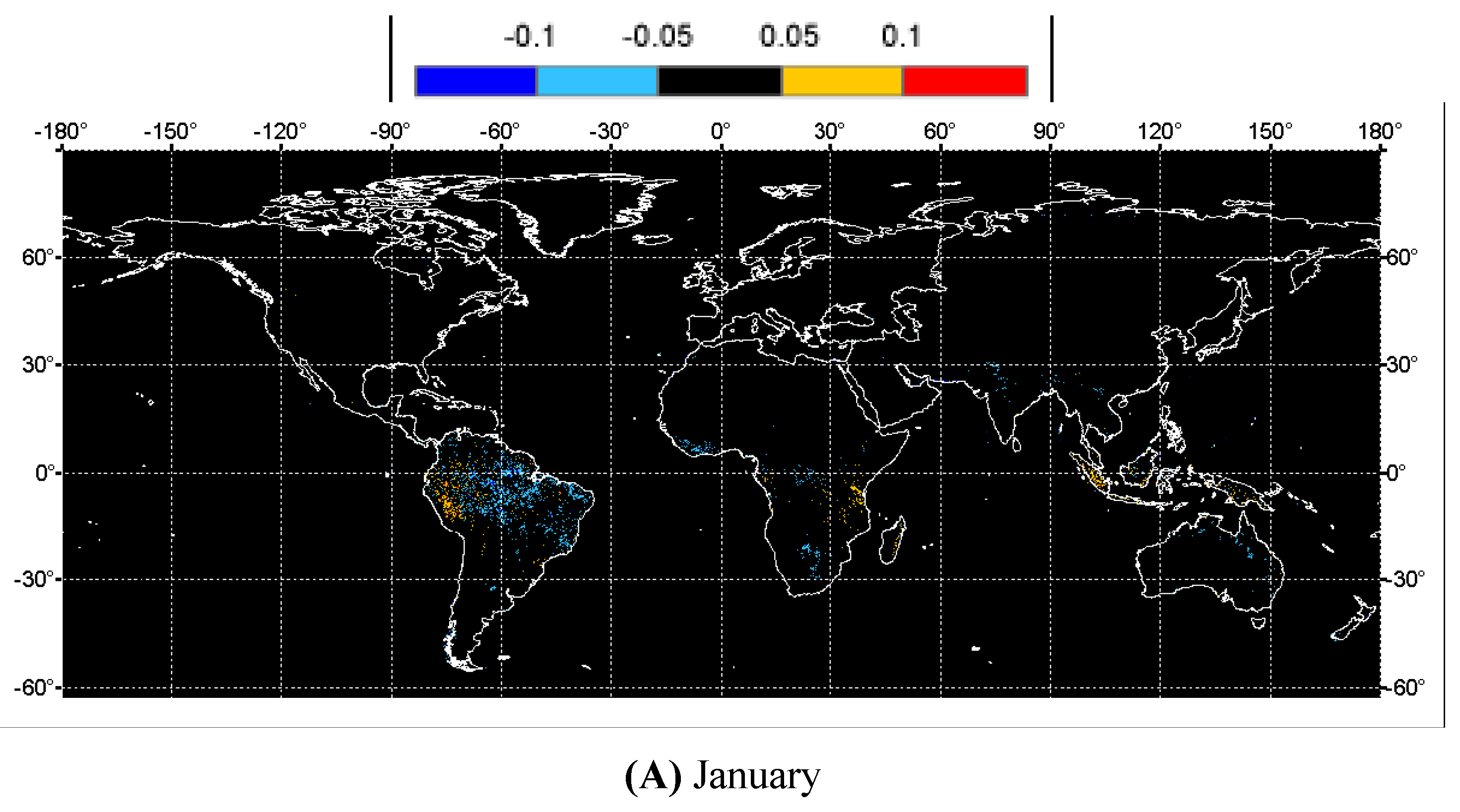
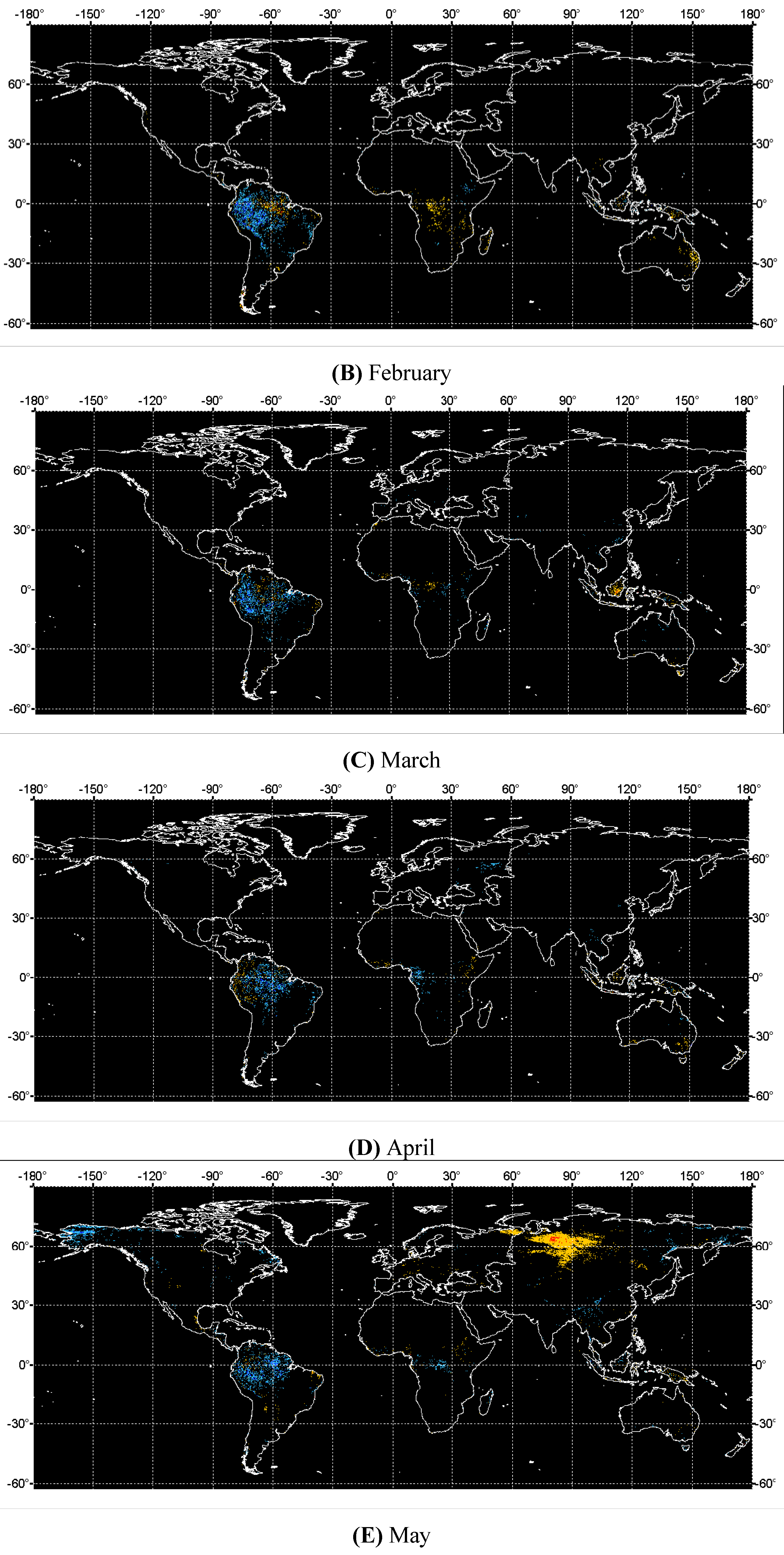
3. Results and Analysis

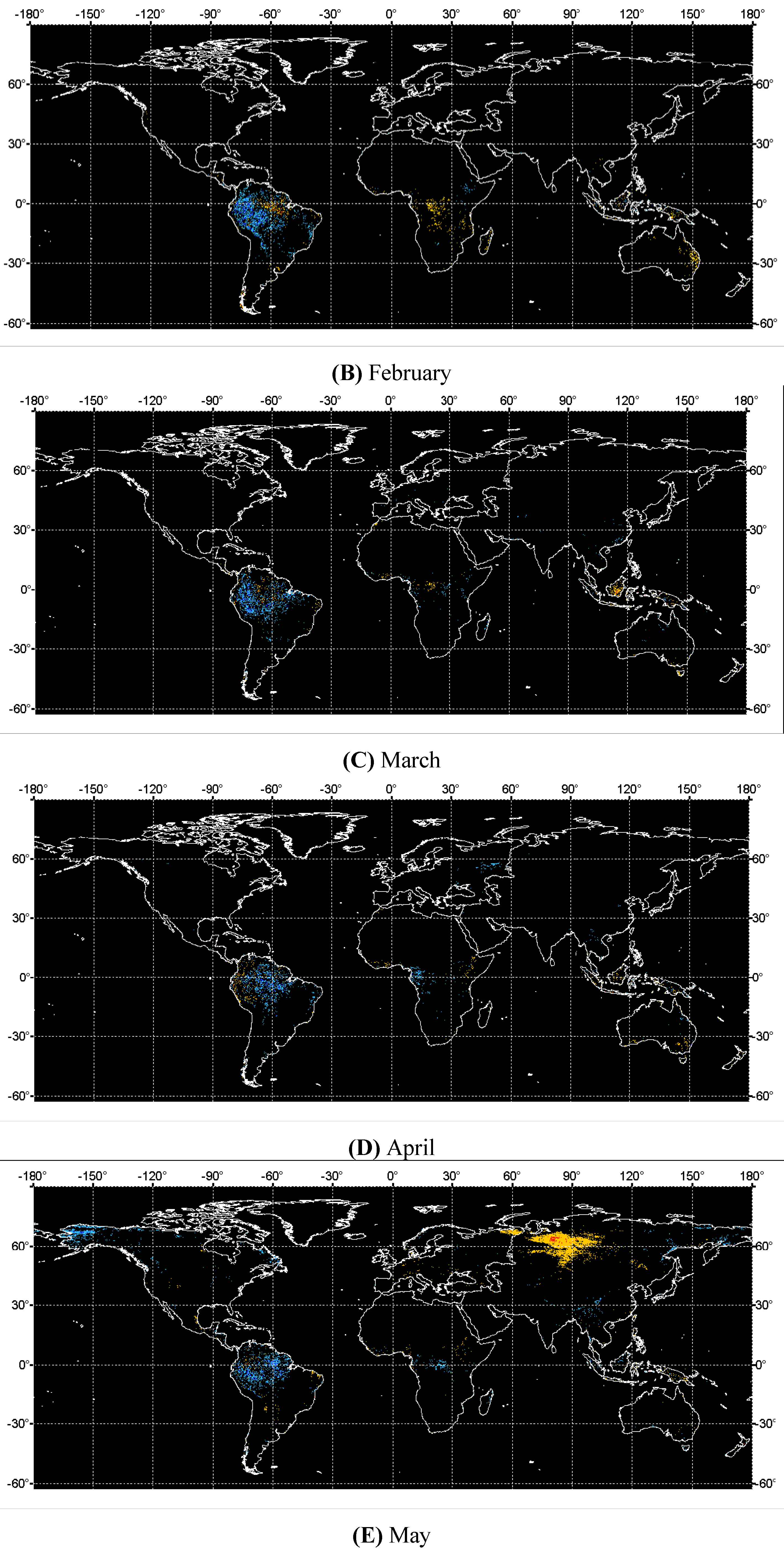
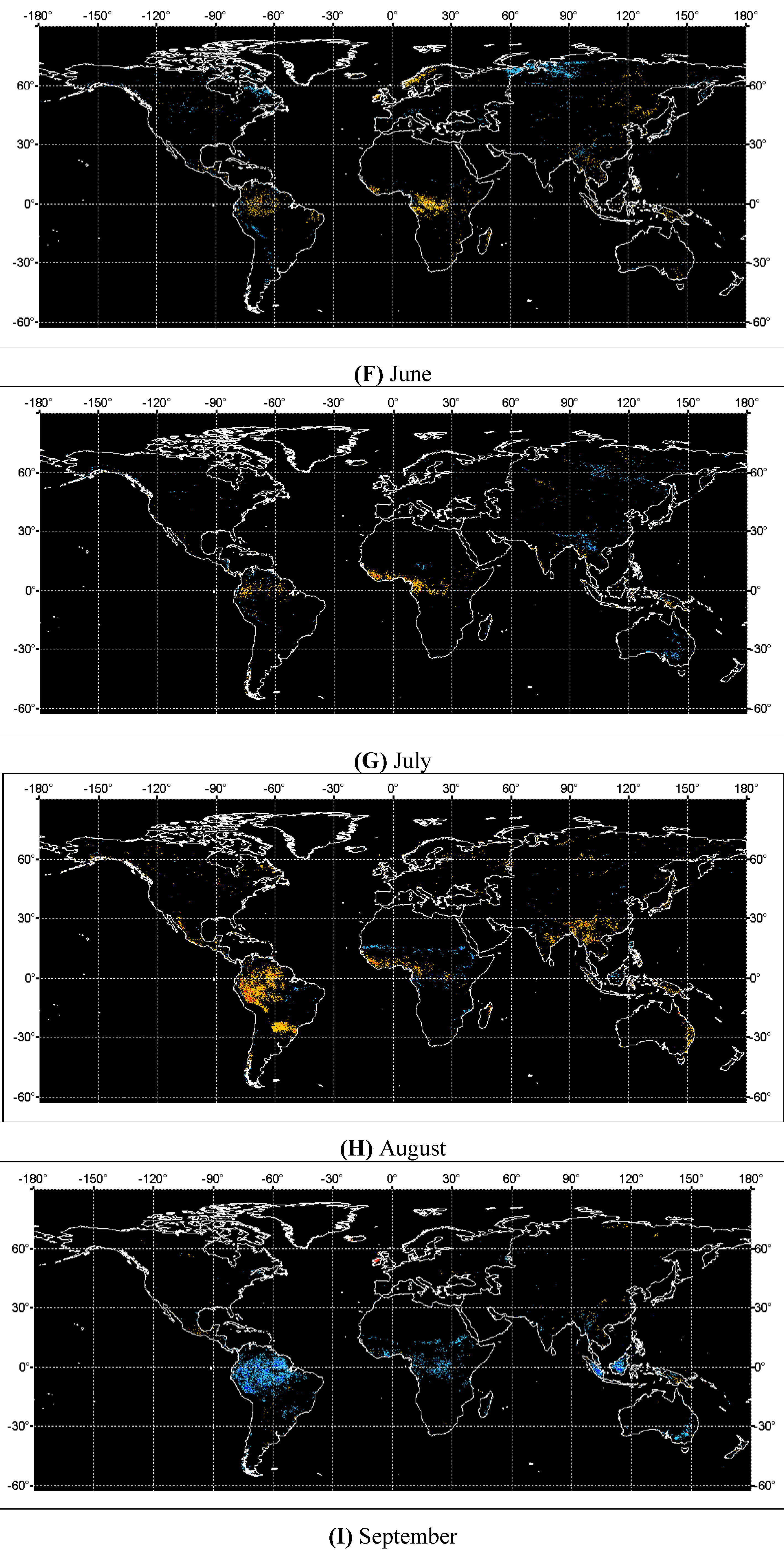
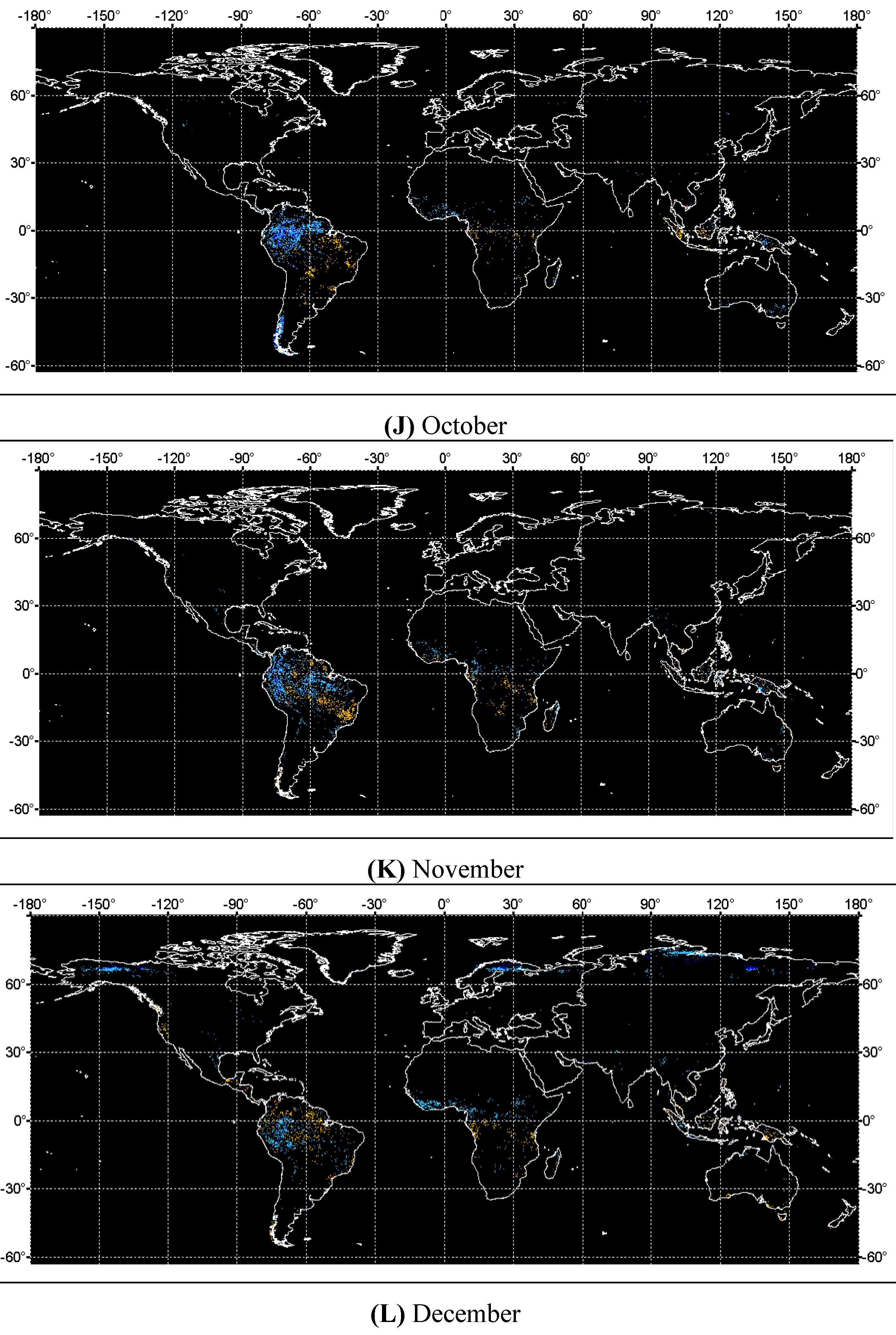
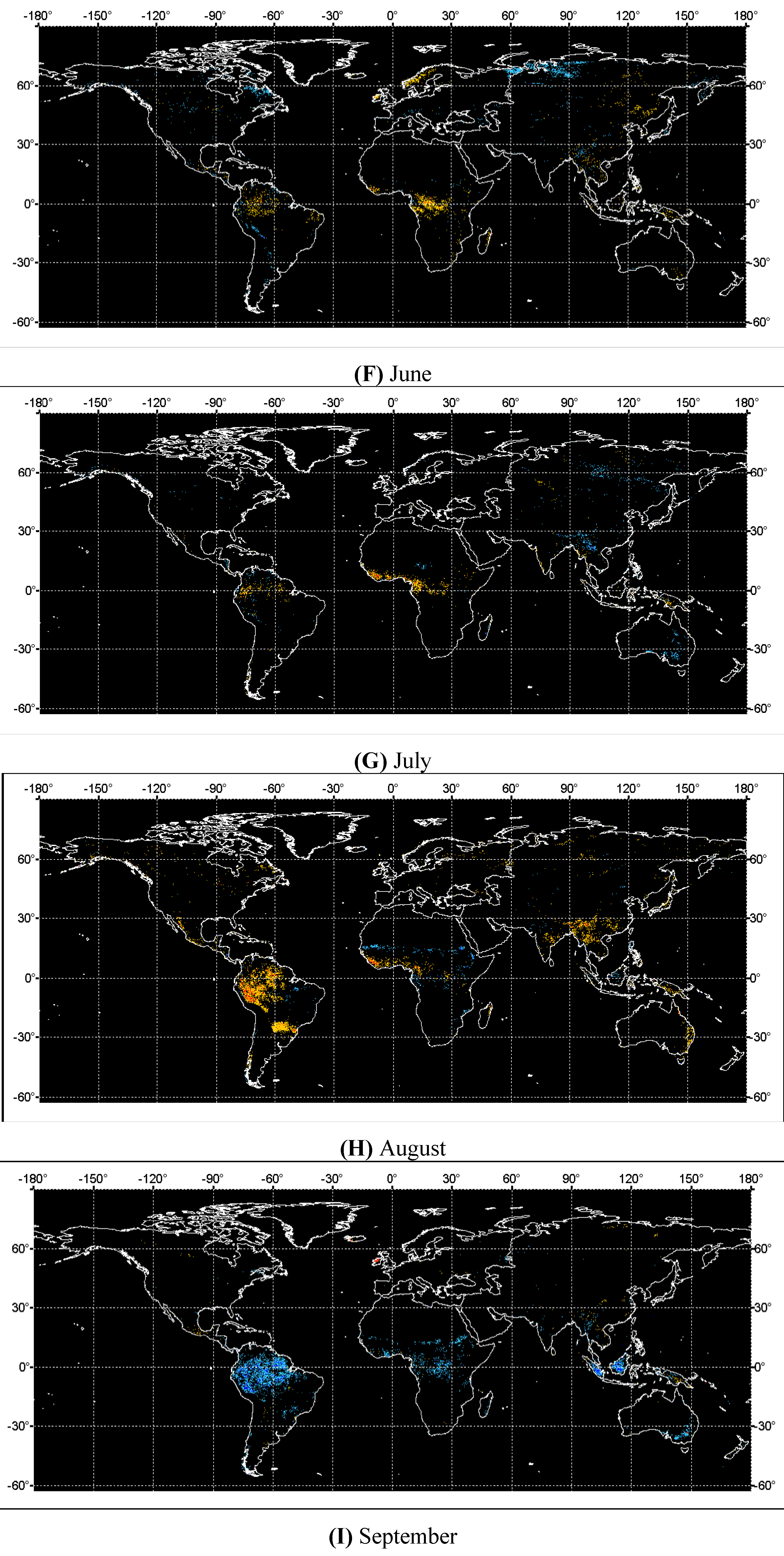
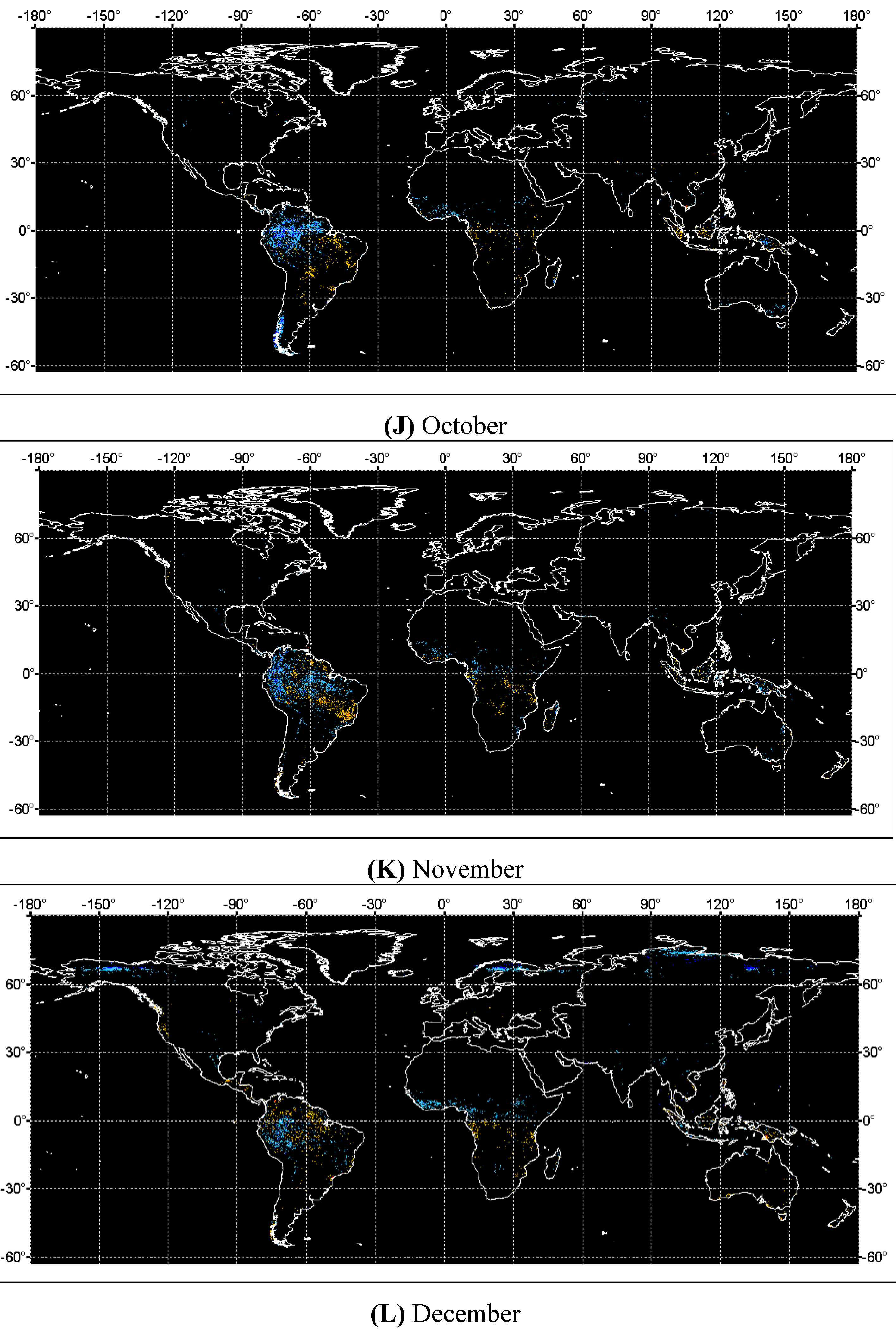
4. Discussions
5. Conclusions
Acknowledgements
References
- Walker, B.H. Landscape to regional-scale responses of terrestrial ecosystems to global change. Ambio 1994, 23, 67–73. [Google Scholar]
- Friedlingstin, P.; Dufrene, J.-L.; Cox, P.M.; Rayner, P. How positive is the feedback between climate change and the carbon cycle? Tellus 2003, 55B, 692–700. [Google Scholar] [CrossRef]
- Houghton, J.T.; Ding, Y.; Griggs, D.J.; Noguer, M.; van der Linden, P.J.; Xiaosu, D. Climate Change 2001: The Scientific Basis Contribution of Working Group I to the Third Assessment Report of the Intergovernmental Panel on Climate Change (IPCC); Cambridge University Press: Cambridge, UK, 2001. [Google Scholar]
- Fang, X.Q.; Yu, W.H. Progress in the studies on the phenological responding to globalwarming. Adv. Earth Sci. 2002, 17, 714–719. [Google Scholar]
- Houghton, J.T. Counting terrestrial sources and sinks of carbon. Climatic Change 2001, 48, 525–534. [Google Scholar] [CrossRef]
- Gurney, K.; Law, R.M.; Ddenning, A.S.; Rayner, P.J.; Baker, D.; Bousquet, P.; Bruhwiler, L.; Chen, Y.-H.; Ciais, P.; Fan, S.; Fung, I.Y.; Gloor, M.; Heimann, M.; Higuchi, K.; John, J.; Maki, T.; Maksyutov, S.; Masarie, K.; Peylin, P.; Prather, M.; Pak, B.C.; Randerson, J.; Sarmiento, J.; Taguchi, S.; Takahashi, T.; Yuen, C.-W. Towards robust regional estimates of CO2 sources and sinks using atmospheric transport models. Nature 2002, 415, 626–630. [Google Scholar] [CrossRef] [PubMed]
- Goulden, M.L.; Munger, J.W.; Fan, S.M.; Daube, B.C.; Wofsy, S.C. Exchange of carbon dioxide by a deciduous forest: Response to interannual climate variability. Science 1996, 271, 1576–1578. [Google Scholar] [CrossRef]
- Fang, J.Y.; Chen, A.; Peng, C.; Zhao, S.; Ci, L. Changes in forest biomass carbon storage in China between 1949 and 1998. Science 2001, 292, 2320–2322. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Gloor, M.; Mahlman, J.; Pacala, S.; Sarmiento, J.; Takahashi, T.; Tans, P. A large terrestrial carbon sink in North America implied by atmospheric and oceanic carbon dioxide data and models. Science 1998, 282, 442–446. [Google Scholar] [CrossRef] [PubMed]
- Pacala, S.; Hurtt, G.C.; Baker, D.; Peylin, P.; Houghton, R.A.; Birdsey, R.A.; Health, L.; Sundquist, E.T.; Stallard, R.F.; Ciais, P.; Moorcroft, P.; Caspersen, J.; Shevliakova, E.; Moore, B.; Kohlmaier, G.; Holland, E.; Gloor, M.; Harmon, M.E.; Fan, S.M.; Sarmiento, J.L.; Goodale, C.L.; Schimel, D.; Field, C.B. Consistent land-and atmosphere-based US carbon sink estimates. Science 2001, 292, 2316–2319. [Google Scholar] [CrossRef] [PubMed]
- Foley, J.A.; Levis, S.; Costa, M.H.; Cramer, W.; Pollard, D. Incorporating dynamic vegetation cover within global climate models. Ecol. Appl. 2000, 10, 1620–1632. [Google Scholar] [CrossRef]
- Bogaert, J.; Zhou, L.; Tucker, C.J. Evidence for a persistent and extensive greening trend in Eurasia inferred from satellite vegetation index data. J. Geophys. Res. 2002, 107, 4119–4135. [Google Scholar] [CrossRef]
- Zhou, L.; Tucker, C.J.; Kaufmann, R.K. Variations in northern vegetation activity inferred from satellite data of vegetation index during 1981 to 1999. J. Geophys. Res. 2001, 106, 20069–20083. [Google Scholar] [CrossRef]
- Burkett, V.R.; Wilcox, D.A.; Stottlemyer, R.; Barrow, W.; Fagre, D.; Baron, J.; Price, J.; Nielsen, J.L.; Allen, C.D.; Peterson, D.L.; Ruggerone, G.; Doyle, T. Nonlinear dynamics in ecosystem response to climatic change: Case studies and policy implications. Ecol. Complex. 2005, 2, 357–394. [Google Scholar] [CrossRef]
- Keeling, C.D.; Chin, J.F.S.; Whorf, T.P. Incressed activity of northern vegetation in inferred from atmospheric CO2 measurements. Nature 1996, 382, 146–149. [Google Scholar] [CrossRef]
- Menzel, A. Phenology: Its importance to the global change community. Climate Change 2002, 54, 379–395. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2007: Synthesis Report; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2007. [Google Scholar]
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Liang, S. Quantitative Remote Sensing of Land Surfaces; John Wiley & Sons: Hoboken, NJ, USA, 2004. [Google Scholar]
- Myneni, R.B.; Hall, F.G.; Sellers, P.J.; Marshak, A.L. The interpretation of spectral vegetation indexes. IEEE Trans. Geosci. Remot. Sens. 1995, 33, 481–486. [Google Scholar] [CrossRef]
- Tucker, C.J.; Fung, I.Y.; Hall, C.D. Relationship between atmospheric CO2 variations and a satellite-derived vegetation index. Nature 1986, 319, 195–199. [Google Scholar] [CrossRef]
- Holben, B.N. Characteristics of maximum-value composite images from Temporal AVHRR Data. Int. J. Remote Sens. 1986, 7, 1417–1434. [Google Scholar] [CrossRef]
- Li, A.; Liang, S.; Wang, A.; Qin, J. Estimating crop yield from multi-temporal satellite data using multivariate regression and neural network techniques. Photogramm. Eng. Remote Sensing 2007, 73, 1149–1159. [Google Scholar] [CrossRef]
- Doktor, D.; Bondeau, A.; Koslowski, D.; Badeck, F.-W. Influence of heterogeneous landscapes on computed green-up dates based on daily AVHRR NDVI observations. Remote Sens. Environ. 2009, 113, 2618–2632. [Google Scholar] [CrossRef]
- Bajgiran, P.R.; Darvishsefat, A.A.; Khalili, A.; Makhdoum, M.F. Using AVHRR-based vegetation indices for drought monitoring in the northwest of Iran. J. Arid Environ. 2008, 72, 1086–1096. [Google Scholar] [CrossRef]
- Paruelo, J.M.; Aguiar, M.R.; Golluscio, R.A.; Leon, R.J.C.; Pujol, G. Environmental controls of NDVI dynamics in patagonia based on noaa-avhrr satellite data. J. Veg. Sci. 1993, 4, 425–428. [Google Scholar] [CrossRef]
- de Beurs, K.M.; Henebry, G.M. A statistical framework for the analysis of long image time-series. Int. J. Remote Sens. 2005, 26, 1551–1573. [Google Scholar] [CrossRef]
- Bradley, B.A.; Jacob, R.W.; Hermance, J.F.; Mustard, J.F. A curve fitting procedure to derive inter-annual phenologies from time-series of noisy satellite NDVI data. Remote Sens. Environ. 2007, 106, 137–145. [Google Scholar] [CrossRef]
- Walker, B.; Steffen, W. An overview of the implications of global change for natural and managed terrestrial ecosystems. Conserv. Ecol. 1997, 1. Available online: http://www.ecologyandsociety.org/vol1/iss2/art2/ (accessed on 20 April 2010). [Google Scholar]
- Eastman, J.R.; Fulk, M. Long sequence time-series evaluation using standardized principal components. Photogramm. Eng. Remote Sensing 1993, 59, 991–996. [Google Scholar]
- Townshed, J.R.G.; Goff, T.E.; Tucker, C.J. Multitemporal dimensionality of images of Normalized Difference Vegetation Index at continental scales. IEEE Trans. Geosci. Remot. Sen. 1985, GE-23, 888–895. [Google Scholar] [CrossRef]
- Jönsson, P.; Eklundh, L. TIMESAT—A program for analyzing time-series of satellite sensor data. Comput. Geosci. 2004, 30, 833–845. [Google Scholar] [CrossRef]
- Azzali, A.; Menenti, M. Mapping vegetation—Soil complexes in southern Africa using temporal Fourier analysis of NOAA AVHRR NDVI data. Int. J. Remote Sens. 2000, 21, 973–996. [Google Scholar] [CrossRef]
- Jakubauskas, M.E.; Legates, D.R.; Kastens, J.H. Harmonic analysis of time-series AVHRR NDVI data. Photogramm. Eng. Remote Sensing 2001, 67, 461–470. [Google Scholar]
- Galford, G.L.; Mustard, J.F.; Melillo, J.; Gendrin, A.; Cerri, C.C.; Cerri, C.E.P. Wavelet analysis of MODIS time-series to detect expansion and intensification of row-crop agriculture in Brazil. Remote Sensi. Environ. 2008, 112, 576–587. [Google Scholar] [CrossRef]
- Martínez, B.; Gilabert, M.A. Vegetation dynamics from NDVI time-series analysis using the wavelet transform. Remote Sensi. Environ. 2009, 113, 1823–1842. [Google Scholar] [CrossRef]
- Suzuki, R.; Masuda, K.; Dye, D.G. Interannual covariability between actual evapotranspiration and PAL and GIMMS NDVIs of northern Asia. Remote Sensi. Environ. 2007, 106, 387–398. [Google Scholar] [CrossRef]
- Heumann, B.W.; Seaquist, J.W.; Eklundh, L.; Jönsson, P. AVHRR derived phenological change in the Sahel and Soudan, Africa, 1982–2005. Remote Sensi. Environ. 2007, 108, 385–392. [Google Scholar] [CrossRef]
- Slayback, D.A.; Pinzon, J.E.; Los, S.O. Northern hemisphere photosynthetic trends 1982–99. Glob. Change Biol. 2003, 9, 1–15. [Google Scholar] [CrossRef]
- Davenport, M.L.; Nicholson, S.E. On the Relation between rainfall and the Normalized Difference Vegetation Index for diverse vegetation types in East Africa. Int. J. Remote Sens. 1993, 14, 2369–2389. [Google Scholar] [CrossRef]
- Fensholt, R.; Rasmussen, K.; Nielsen, T.T.; Mbow, C. Evaluation of earth observation based long term vegetation trends—Intercomparing NDVI time-series trend analysis consistency of Sahel from AVHRR GIMMS, Terra MODIS and SPOT VGT data. Remote Sens. Environ. 2009, 113, 1886–1898. [Google Scholar] [CrossRef]
- Tucker, C.J.; Pinzon, J.E.; Brown, M.E.; Slayback, D.A.; Pak, E.W.; Mahoney, R.; Vermote, E.F.; El Saleous, N. An extended AVHRR 8-km NDVI dataset compatible with MODIS and SPOT vegetation NDVI data. Int. J. Remote Sens. 2005, 26, 4485–4498. [Google Scholar] [CrossRef]
- Cihlar, J.; Chen, J.; Li, Z.; Huang, F.; Pokrant, H. Can interannual land surface signal be discerned in composite AVHRR data? J. Geophys. Res. 1998, 103, 23163–23172. [Google Scholar] [CrossRef]
- Nagol, J.R.; Vermote, E.F.; Prince, S.D. Effects of atmospheric variation on AVHRR NDVI data. Remote Sens. Environ. 2009, 113, 392–397. [Google Scholar] [CrossRef]
- Diwakar, P.G.; Prabhakar, P.; Venkataramana, I.; Devarajan, R. Derivation of maximum value composite images using multidate NOAA-AVHRR data. J. Indian Soc. Remote Sens. 1989, 17, 1–6. [Google Scholar] [CrossRef]
- Savitzky, A.; Golay, M.J.E. Smoothing and differentiation of data by simplified least squares procedures. Anal. Chem. 1964, 36, 1627–1639. [Google Scholar] [CrossRef]
- Chen, J.; Jönsson, P.; Tamura, M.; Gu, Z.; Matsushita, B.; Eklundh, L. A simple method for reconstructing a high-quality NDVI time-series data set based on the Savitzky–Golay filter. Remote Sens. Environ. 2004, 91, 332–344. [Google Scholar] [CrossRef]
- Bian, J.; Li, A.; Song, M.; Ma, L.; Jiang, J. Reconstructing NDVI time-series data set based on the Savitzky-Golay filter. J. Remote Sens. 2010, 14, 001–009. [Google Scholar]
- Steinier, J.; Termonia, Y.; Deltour, J. Comments on smoothing and differentiation of data by simplified least squares procedure. Anal. Chem. 1972, 44, 1906–1909. [Google Scholar] [CrossRef] [PubMed]
- Richardson, A.J.; Wiegand, C.L. Distinguishing vegetation from soil background information. Photogramm. Eng. Remote Sensing 1977, 43, 1541–1552. [Google Scholar]
- Hannachi, A.; Unkel, S.; Trendafilov, N.T.; Jolliffe, I.T. Independent component analysis of climate data: A new look at EOF rotation. J. Climate 2009, 22, 2797–2812. [Google Scholar] [CrossRef]
- Obukhov, A.M. Statistically homogeneous fields on a sphere. Uspekhi Matematicheskikh Nauk 1947, 2, 196–198. [Google Scholar]
- Carleton, A.M. Methodology in climatology. Ann. Assn. Amer. Geogr. 1999, 89, 713–735. [Google Scholar] [CrossRef]
- Jolliffe, I.T. Principal Component Analysis; Springer: Berlin, Germany, 2002. [Google Scholar]
- Prasad, A.K.; Sarkar, S.; Singh, R.P.; Kafatos, M. Inter-annual variability of vegetation cover and rainfall over india. Adv. Space Res. 2007, 39, 79–87. [Google Scholar] [CrossRef]
- Nezlin, N.P.; Kostianoy, A.G.; Li, B.-L. Inter-annual variability and interaction of remote-sensed vegetation index and atmospheric precipitation in the Aral Sea region. J. Arid Environ. 2005, 62, 677–700. [Google Scholar] [CrossRef]
- Sarkar, S.; Kafatos, M. Interannual variability of vegetation over the Indian sub-continent and its relation to the different meteorological parameters. Remote Sens. Environ. 2004, 90, 268–280. [Google Scholar] [CrossRef]
- Lobo, A.; Maisongrande, P. Searching for trends of change through exploratory data analysis of time-series of remotely sensed images of SW Europe and NW Africa. Int. J. Remote Sens. 2008, 29, 5237–5245. [Google Scholar] [CrossRef]
- Golub, G.H.; van Loan, C.F. Matrix Computations; The John Hopkins University Press: Baltimore, MD, USA, 1996. [Google Scholar]
- Hansen, M.; DeFries, R.; Townshend, J.R.G.; Sohlberg, R. Global land cover classification at 1km resolution using a decision tree classifier. Int. J. Remote Sens. 2000, 21, 1331–1365. [Google Scholar] [CrossRef]
- Huang, Y.; Zhang, W.; Sun, W.; Zheng, X. Net primary production of Chinese croplands from 1950 to 1999. Ecol. Appl. 2007, 17, 692–701. [Google Scholar] [CrossRef] [PubMed]
- Jia, G.J.; Epstein, H.E.; Walker, D.A. Spatial characteristics of AVHRR-NDVI along latitudinal transects in northern Alaska. J. Veg. Sci. 2002, 13, 315–326. [Google Scholar] [CrossRef]
- Dewar, R.C.; Franklin, O.; Mäkelä, A.; Mcmurtrie, R.E.; Valentine, H.T. Optimal Function Explains Forest Responses to Global Change. BioScience 2009, 59, 127–139. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2001: The Science of Climate Change; Contribution of Working Group I to the Third Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2001; p. 658. [Google Scholar]
- Hansen, J.; Sato, M.; Ruedy, R.; Lo, K.; Lea, D.W.; Medina-Elizade, M. Global temperature change. PNAS 2006, 103, 14288–14293. [Google Scholar] [CrossRef] [PubMed]
- Kawabata, A.; Yamaguchi, Y. Global monitoring of interannnual changes in vegetation activities using NDVI and its relationships to temperature and precipitation. Int. J. Remote Sens. 2001, 22, 1377–1382. [Google Scholar] [CrossRef]
- Piao, S.L.; Fang, J.Y.; Chen, A.P. Seasonal dynamics of terrestrial primary production in response to climate change in China. Acta Bot. Sin. 2003, 45, 269–275. [Google Scholar]
- Notaro, M. Response of the mean global vegetation distribution to interannual climate variability. Clim. Dynam. 2008, 30, 845–854. [Google Scholar] [CrossRef]
- Lotsch, A.; Friedl, M.A.; Anderson, B.T.; Tucker, C.J. Coupled vegetation-precipitation variability observed from satellite and climate records. Geophys. Res. Lett. 2003, 30, 1774. [Google Scholar] [CrossRef]
- Nemani, R.; White, M.; Thornton, P.; Nishida, K.; Reddy, S.; Jenkins, J.; Running, S. Recent trends in hydrologic balance have enhanced the terrestrial carbon sink in the United States. Geophys. Res. Lett. 2002, 29, 1468. [Google Scholar] [CrossRef]
- Shigehara, K. Phonological observation data in Japan to be utilized as an indicator of climatic variation. In Proceedings of the International Conference on Climatic Impacts on the Environment and Society; University of Tsukuba: Ibaraki, Japan, 1991; pp. C1–C6. [Google Scholar]
- Poveda, G.; Salazar, L.F. Annual and interannual (ENSO) variability of spatial scaling properties of a vegetation index (NDVI) in Amazonia. Remote Sens. Environ. 2004, 93, 391–401. [Google Scholar] [CrossRef]
- Ottersen, G.; Planque, B.; Belgravio, A. Ecological effects of the Northern Atlantic Oscillation. Oecologia 2001, 128, 1–14. [Google Scholar] [CrossRef]
- Li, A.; Liang, S.; Huang, C.; Wang, A. Investigating the impacts of the north Atlantic Oscillation on global vegetation changes. J. Geophys. Res. submitted.
- Souza, C.M.; Roberts, D.A.; Monteiro, A.L. Multitemporal analysis of degraded forests in the southern Brazilian Amazon. Earth Interactions 2005, 9, 1–25. [Google Scholar] [CrossRef]
- Potter, C.; Tan, P. N.; Kumar, V.; Kucharik, C.; Klooster, S.; Genovese, V.; Cohen, W.; Healey, S. Recent history of large-scale ecosystem disturbances in North America derived from the AVHRR satellite record. Ecosystems 2005, 8, 808–824. [Google Scholar] [CrossRef]
- Hansen, M.C.; DeFries, R.S. Detecting long-term global forest change using continuous fields of tree-cover maps from 8-km advanced very high resolution radiometer (AVHRR) data for the years 1982–99. Ecosystems 2004, 7, 695–716. [Google Scholar] [CrossRef]
- Zhou, H.J.; Van Rompaey, A.; Wang, J.A. Detecting the impact of the “Grain for Green” program on the mean annual vegetation cover in the Shaanxi province, China using SPOT-VGT NDVI data. Land Use Policy 2009, 26, 954–960. [Google Scholar] [CrossRef]
- Chen, X.G.; Zhang, X.Q.; Zhang, Y.P.; Wan, C.B. Carbon sequestration potential of the stands under the grain for green program in Yunnan province, China. Forest Ecol. Manage. 2009, 258, 199–206. [Google Scholar] [CrossRef]
- Erda, L. Agricultural vulnerability and adaptation to global, warming in China. Water Air Soil Pollut. 1996, 92, 63–73. [Google Scholar]
- Ma, M.G.; Dong, L.X.; Wang, X.M. Study on the dynamically monitoring and simulating the vegetation cover in northwest China in the past 21 years. J. Glaciol. Geocryol. 2003, 25, 232–236. [Google Scholar]
- Piao, S.L.; Fang, J.Y. Dynamic vegetation cover change over the last 18 years in China. Quaternary Sci. 2001, 21, 294–302. [Google Scholar]
- Cao, M.K.; Prince, S.D.; Li, K.P. Response of terrestrial carbon uptake to climate interannual variability in China. Glob. Change Biol. 2003, 9, 536–546. [Google Scholar] [CrossRef]
- Yang, J.P.; Ding, Y.J.; Chen, R.S. NDVI reflection of alpine vegetation changes in the source regions of the Yangtze and Yellow rivers. Acta Geographica Sinica 2005, 60, 467–478. [Google Scholar]
- Huete, A.R. A soil-adjusted vegetation index (SAVI). Remote Sens. Environ. 1988, 25, 53–70. [Google Scholar] [CrossRef]
- McCloy, K.R.; Los, S.; Lucht, W.; Højsgaard, S. A comparative analysis of three long-term NDVI datasets derived from AVHRR satellite data. EARSeL eProceedings 2005, 4, 52–69. [Google Scholar]
- Fontana, F.; Rixen, C.; Jonas, T.; Aberegg, G.; Wunderle, S. Alpine grassland phenology as seen in AVHRR, VEGETATION, and MODIS NDVI time-series—A comparison with in situ measurements. Sensors 2008, 8, 2833–2853. [Google Scholar] [CrossRef] [Green Version]
- Justice, C.O.; Townshend, J.R.G.; Holben, B.N.; Tucker, C.J. Analysis of the phenology of global vegetation using meteorological satellite data. Int. J. Remote Sens. 1985, 6, 1271–1318. [Google Scholar] [CrossRef]
- Knight, J.F.; Lunetta, R.S.; Ediriwickrema, J.; Khorrarn, S. Regional scale land cover characterization using MODIS-NDVI 250 m multi-temporal imagery: A phenology-based approach. GISci. Remote Sens. 2006, 43, 1–23. [Google Scholar] [CrossRef]
- van Leeuwen, W.J.D.; Davison, J.E.; Casady, G.M.; Marsh, S.E. Phenological characterization of desert sky island vegetation communities with remotely sensed and climate time-series data. Remote Sensing 2010, 2, 388–415. [Google Scholar] [CrossRef]
- Gu, Y.; Brown, J.F.; Miura, T.; van Leeuwen, W.J.D.; Reed, B.C. Phenological classification of the United States: A geographic framework for extending multi-sensor time-series data. Remote Sensing 2010, 2, 526–544. [Google Scholar] [CrossRef]
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an Open Access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, A.; Deng, W.; Liang, S.; Huang, C. Investigation on the Patterns of Global Vegetation Change Using a Satellite-Sensed Vegetation Index. Remote Sens. 2010, 2, 1530-1548. https://doi.org/10.3390/rs2061530
Li A, Deng W, Liang S, Huang C. Investigation on the Patterns of Global Vegetation Change Using a Satellite-Sensed Vegetation Index. Remote Sensing. 2010; 2(6):1530-1548. https://doi.org/10.3390/rs2061530
Chicago/Turabian StyleLi, Ainong, Wei Deng, Shunlin Liang, and Chengquan Huang. 2010. "Investigation on the Patterns of Global Vegetation Change Using a Satellite-Sensed Vegetation Index" Remote Sensing 2, no. 6: 1530-1548. https://doi.org/10.3390/rs2061530