
Influences of Satellite Sensor and Scale on Derivation of Ecosystem Functional Types and Diversity
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
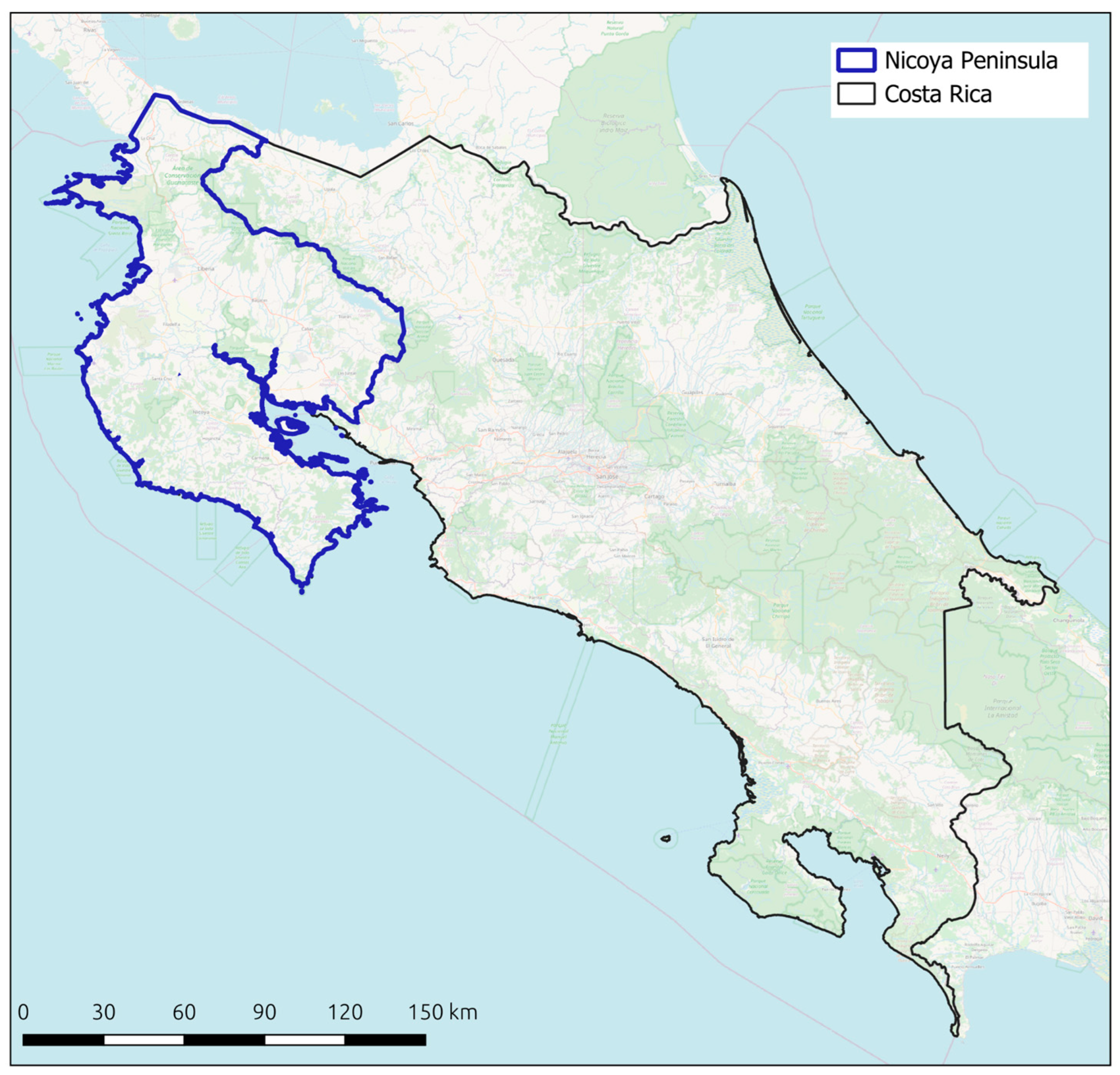
2.1. Costa Rica and the Nicoya Peninsula as a Case Study
2.2. Satellite Data
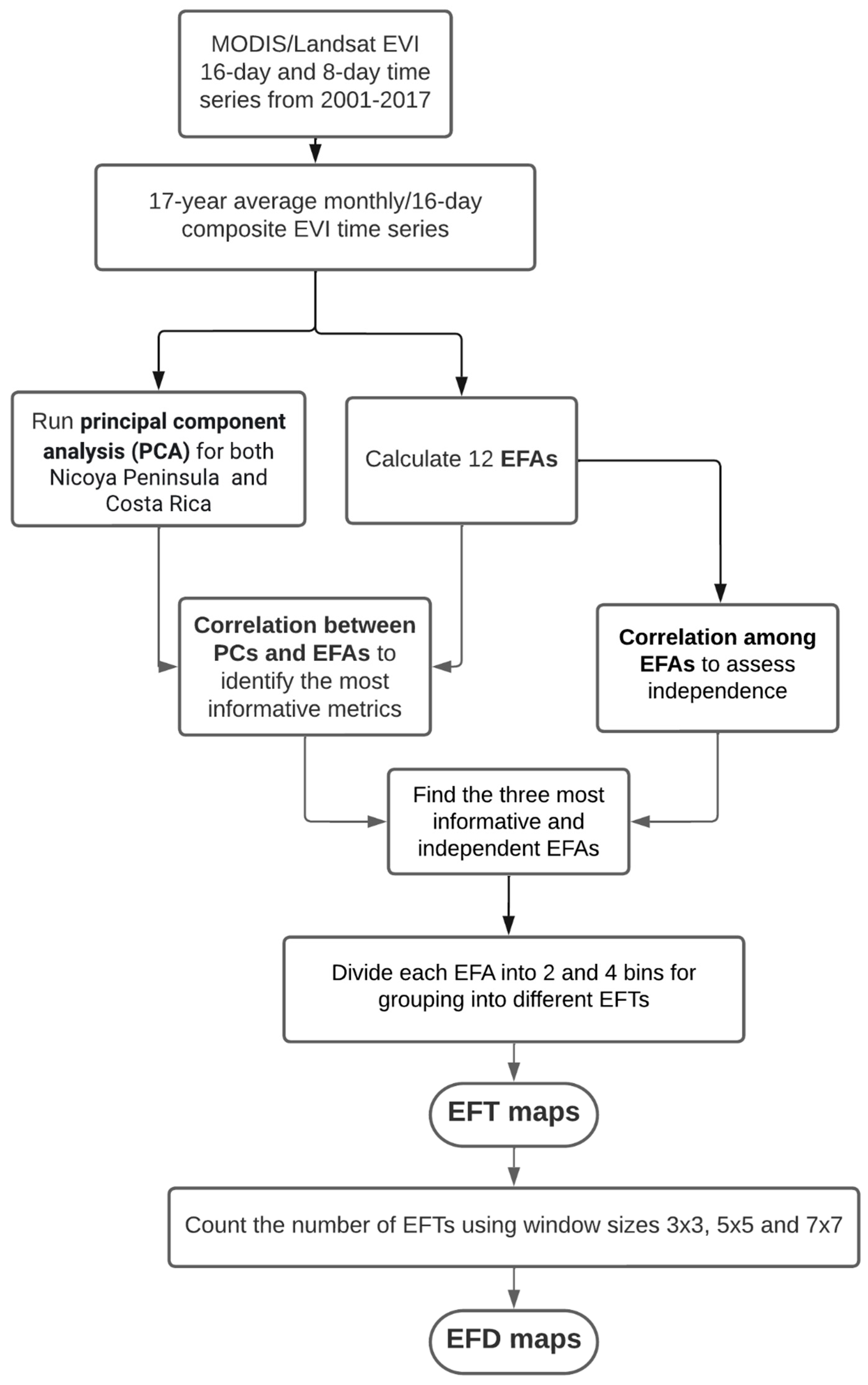
2.3. Obtaining Ecosystem Functional Types and Diversity
2.4. Comparison between MODIS-Based and Landsat-Based EFTs and EFDs in Costa Rica and EFTs and EFDs at National and Local Scales
3. Results
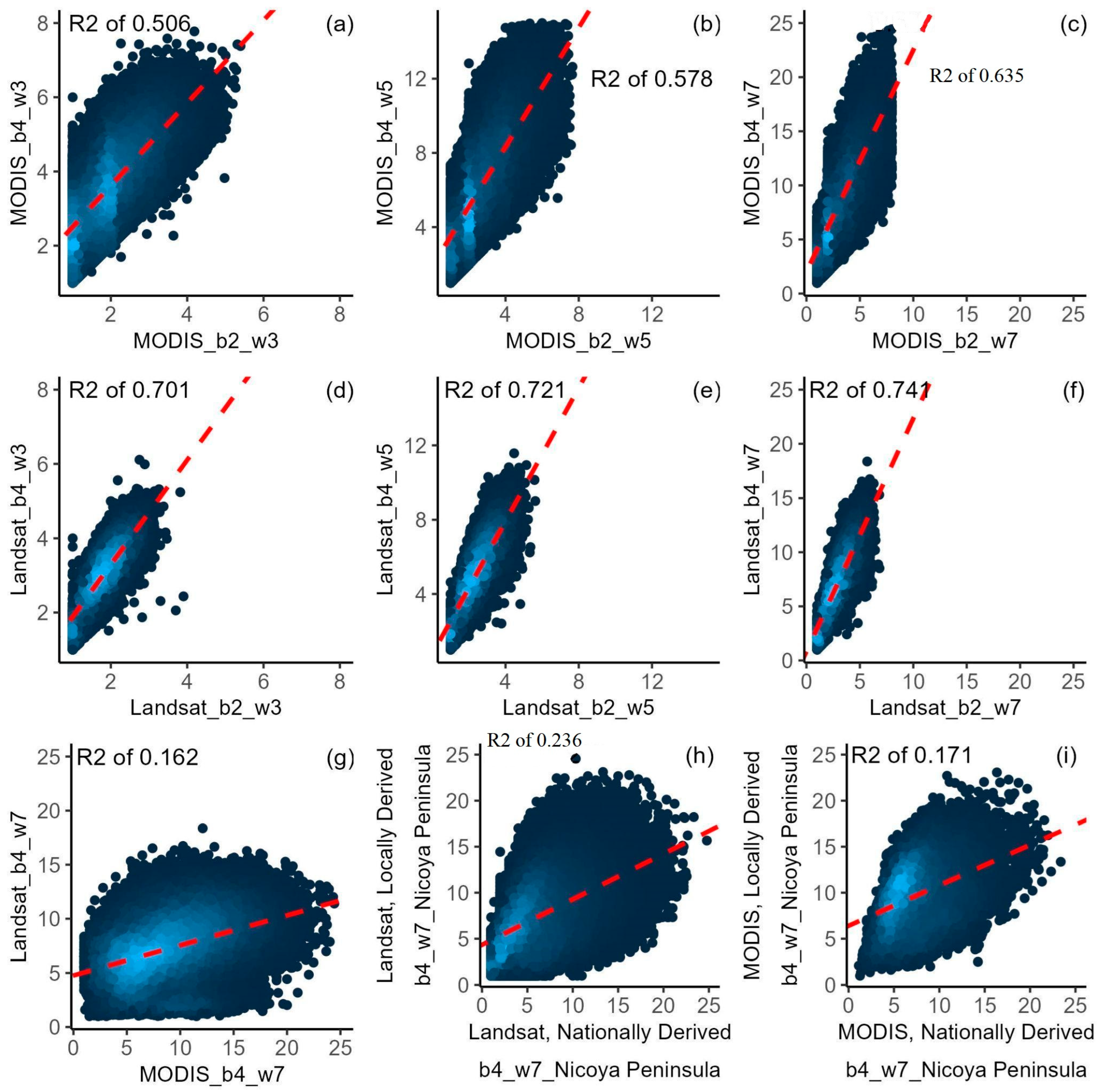
3.1. Ecosystem Functional Attributes (EFAs)
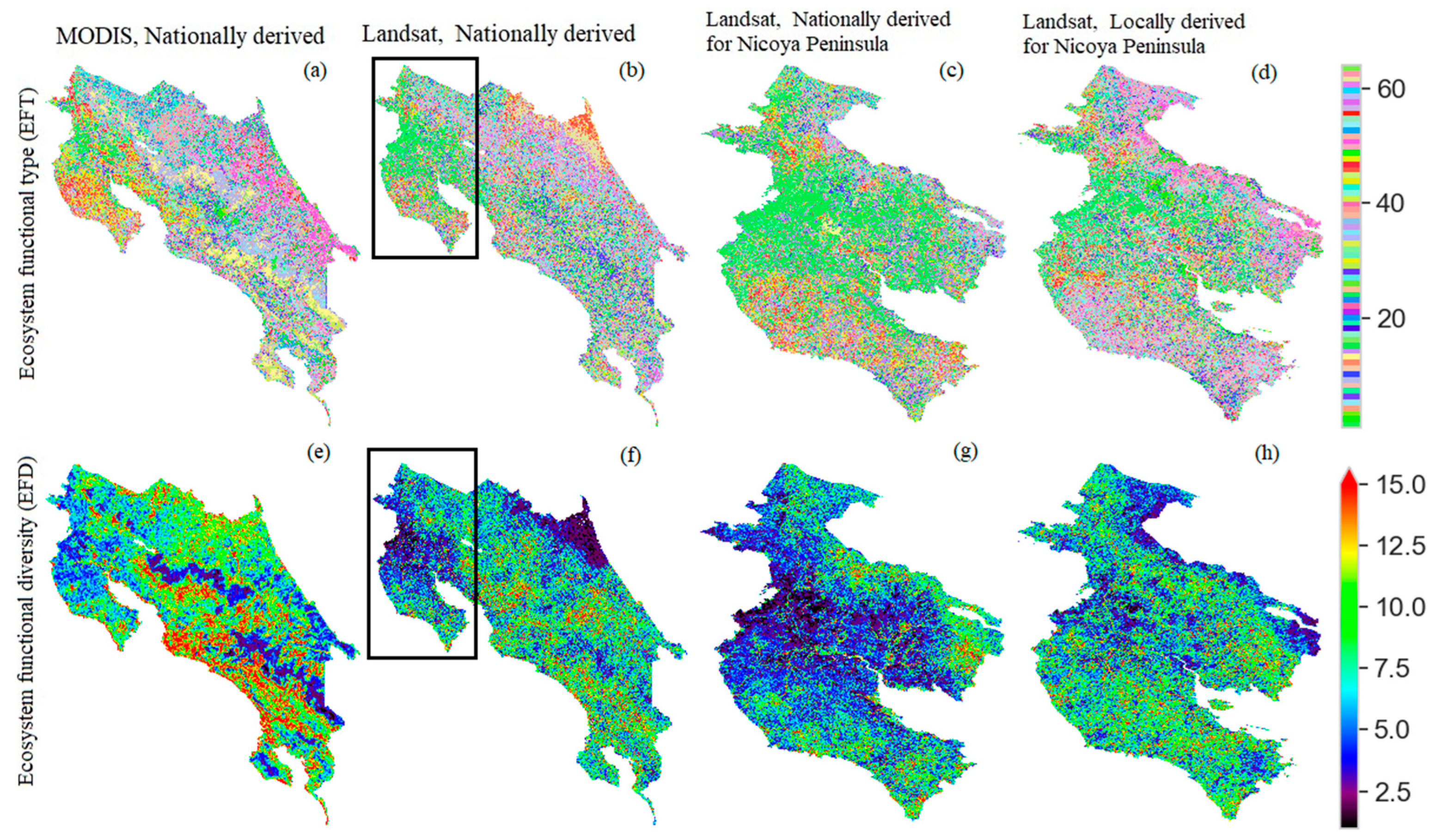
3.2. Ecosystem Functional Types (EFTs)
3.3. Ecosystem Functional Diversity (EFDs)
3.4. Comparison of EFDs Calculated Using Different Methods
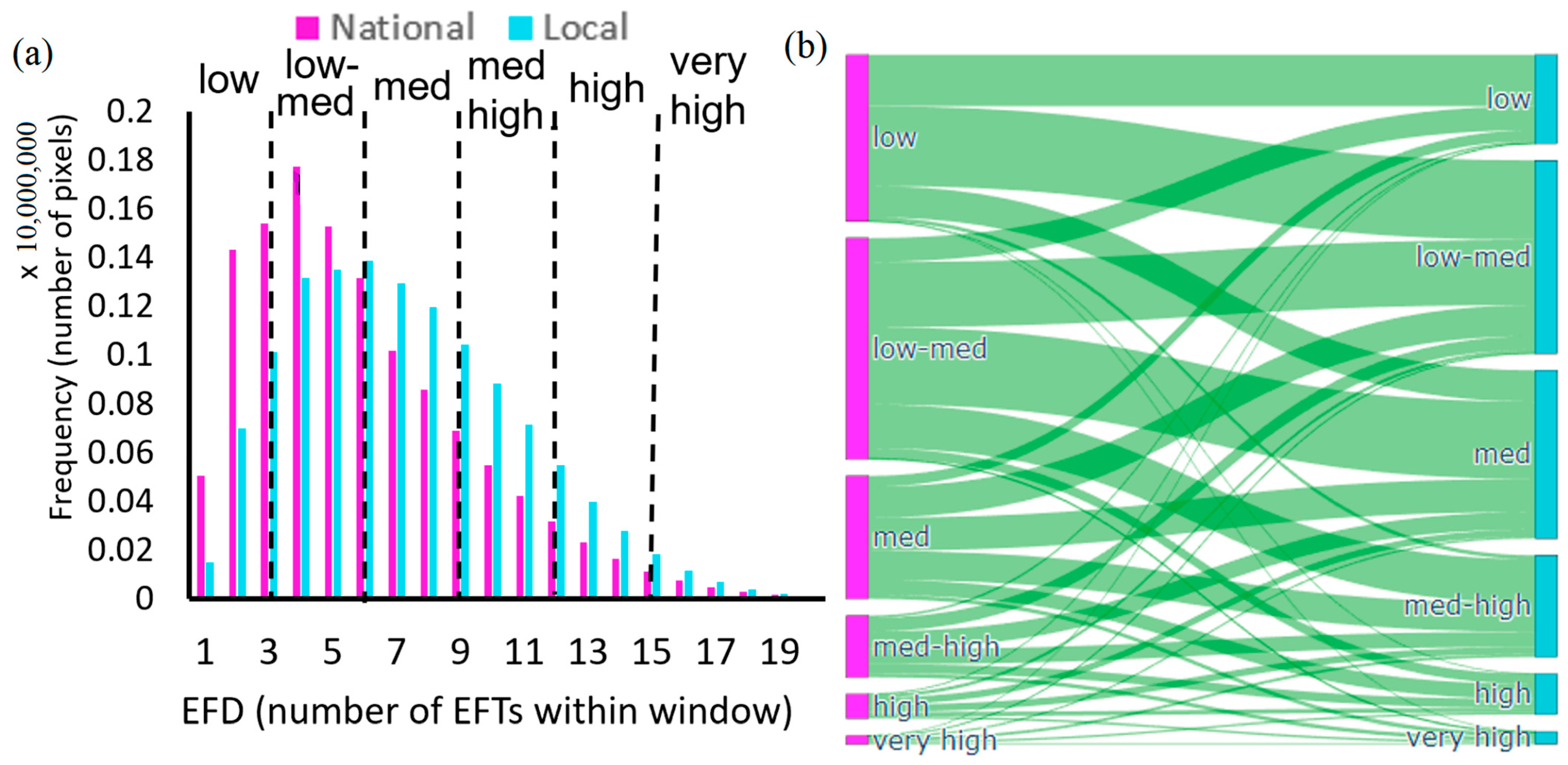
3.5. Comparison of EFDs Calculated across Scales
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mucina, L. Biome: Evolution of a Crucial Ecological and Biogeographical Concept. New Phytol. 2019, 222, 97–114. [Google Scholar] [CrossRef] [PubMed]
- Alcaraz, D.; Paruelo, J.; Cabello, J. Identification of Current Ecosystem Functional Types in the Iberian Peninsula. Glob. Ecol. Biogeogr. 2006, 15, 200–212. [Google Scholar] [CrossRef]
- Paruelo, J.M.; Jobbágy, E.G.; Sala, O.E. Current Distribution of Ecosystem Functional Types in Temperate South America. Ecosystems 2001, 4, 683–698. [Google Scholar] [CrossRef]
- Cazorla, B.P.; Cabello, J.; Peñas, J.; Garcillán, P.P.; Reyes, A.; Alcaraz-Segura, D. Incorporating Ecosystem Functional Diversity into Geographic Conservation Priorities Using Remotely Sensed Ecosystem Functional Types. Ecosystems 2021, 24, 548–564. [Google Scholar] [CrossRef]
- Fernández, N.; Paruelo, J.M.; Delibes, M. Ecosystem Functioning of Protected and Altered Mediterranean Environments: A Remote Sensing Classification in Doñana, Spain. Remote Sens. Environ. 2010, 114, 211–220. [Google Scholar] [CrossRef]
- Ivits, E.; Cherlet, M.; Mehl, W.; Sommer, S. Ecosystem Functional Units Characterized by Satellite Observed Phenology and Productivity Gradients: A Case Study for Europe. Ecol. Indic. 2013, 27, 17–28. [Google Scholar] [CrossRef]
- Alcaraz-Segura, D.; Paruelo, J.M.; Epstein, H.E.; Cabello, J. Environmental and Human Controls of Ecosystem Functional Diversity in Temperate South America. Remote Sens. 2013, 5, 127–154. [Google Scholar] [CrossRef]
- Cabello, J.; Alcaraz-Segura, D.; Altesor, A.; Delibes, M.; Baeza, S.; Liras, E. Funcionamiento Ecosistémico y Evaluación de Prioridades Geográficas En Conservación. Ecosistemas 2008, 17, 53–63. [Google Scholar]
- Cabello, J.; Lourenço, P.; Reyes, A.; Alcaraz-Segura, D. Ecosystem Services Assessment of National Parks Networks for Functional Diversity and Carbon Conservation Strategies Using Remote Sensing. Earth Obs. Ecosyst. Serv. 2013, 179, 430. [Google Scholar]
- Paruelo, J.; Alcaraz-Segura, D.; Volante, J.N. El seguimiento del nivel de provisión de los servicios ecosistémicos. Expans. E Intensificación Agríc. En Argent. Valorac. Bienes Serv. Ecosistémicos Para El Ordenamiento Territ. Ed INTA B. Aires. 2011. Available online: https://www.researchgate.net/profile/Esteban-Jobbagy/publication/249643891_Valoracion_de_Servicios_Ecosistemicos_Conceptos_herramientas_y_aplicaciones_para_el_ordenamiento_territorial/links/0deec51e963480928c000000/Valoracion-de-Servicios-Ecosistemicos-Conceptos-herramientas-y-aplicaciones-para-el-ordenamiento-territorial.pdf#page=147 (accessed on 25 September 2023).
- Volante, J.N.; Alcaraz-Segura, D.; Mosciaro, M.J.; Viglizzo, E.F.; Paruelo, J.M. Ecosystem Functional Changes Associated with Land Clearing in NW Argentina. Agric. Ecosyst. Environ. 2012, 154, 12–22. [Google Scholar] [CrossRef]
- Oki, T.; Blyth, E.M.; Berbery, E.H.; Alcaraz-Segura, D. Land Use and Land Cover Changes and Their Impacts on Hydroclimate, Ecosystems and Society. In Climate Science for Serving Society; Springer: Berlin/Heidelberg, Germany, 2013; pp. 185–203. [Google Scholar]
- Müller, O.V.; Berbery, E.H.; Alcaraz-Segura, D.; Ek, M.B. Regional Model Simulations of the 2008 Drought in Southern South America Using a Consistent Set of Land Surface Properties. J. Clim. 2014, 27, 6754–6778. [Google Scholar] [CrossRef]
- Potter, C.S.; Brooks, V. Global Analysis of Empirical Relations between Annual Climate and Seasonality of NDVI. Int. J. Remote Sens. 1998, 19, 2921–2948. [Google Scholar] [CrossRef]
- Zhang, X.; Friedl, M.A.; Schaaf, C.B.; Strahler, A.H.; Hodges, J.C.F.; Gao, F.; Reed, B.C.; Huete, A. Monitoring Vegetation Phenology Using MODIS. Remote Sens. Environ. 2003, 84, 471–475. [Google Scholar] [CrossRef]
- Pérez-Hoyos, A.; Martínez, B.; García-Haro, F.J.; Moreno, Á.; Gilabert, M.A. Identification of Ecosystem Functional Types from Coarse Resolution Imagery Using a Self-Organizing Map Approach: A Case Study for Spain. Remote Sens. 2014, 6, 11391–11419. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, F. Identification and Analysis of Ecosystem Functional Types in the West of Songnen Plain, China, Based on Moderate Resolution Imaging Spectroradiometer Data. J. Appl. Remote Sens. 2015, 9, 096096. [Google Scholar] [CrossRef]
- Cadotte, M.W.; Carscadden, K.; Mirotchnick, N. Beyond Species: Functional Diversity and the Maintenance of Ecological Processes and Services. J. Appl. Ecol. 2011, 48, 1079–1087. [Google Scholar] [CrossRef]
- Díaz, S.; Lavorel, S.; Chapin, F.S.; Tecco, P.A.; Gurvich, D.E.; Grigulis, K. Functional Diversity—At the Crossroads between Ecosystem Functioning and Environmental Filters. In Terrestrial Ecosystems in a Changing World; Springer: Berlin/Heidelberg, Germany, 2007; pp. 81–91. [Google Scholar]
- Balvanera, P.; Pfisterer, A.B.; Buchmann, N.; He, J.-S.; Nakashizuka, T.; Raffaelli, D.; Schmid, B. Quantifying the Evidence for Biodiversity Effects on Ecosystem Functioning and Services. Ecol. Lett. 2006, 9, 1146–1156. [Google Scholar] [CrossRef] [PubMed]
- Díaz, S.; Fargione, J.; Chapin III, F.S.; Tilman, D. Biodiversity Loss Threatens Human Well-Being. PLoS Biol. 2006, 4, e277. [Google Scholar] [CrossRef]
- Duncan, C.; Thompson, J.R.; Pettorelli, N. The Quest for a Mechanistic Understanding of Biodiversity–Ecosystem Services Relationships. Proc. R. Soc. B Biol. Sci. 2015, 282, 20151348. [Google Scholar] [CrossRef]
- Mouchet, M.A.; Villéger, S.; Mason, N.W.H.; Mouillot, D. Functional Diversity Measures: An Overview of Their Redundancy and Their Ability to Discriminate Community Assembly Rules. Funct. Ecol. 2010, 24, 867–876. [Google Scholar] [CrossRef]
- Proença, V.; Martin, L.J.; Pereira, H.M.; Fernandez, M.; McRae, L.; Belnap, J.; Böhm, M.; Brummitt, N.; García-Moreno, J.; Gregory, R.D. Global Biodiversity Monitoring: From Data Sources to Essential Biodiversity Variables. Biol. Conserv. 2017, 213, 256–263. [Google Scholar] [CrossRef]
- Skidmore, A.K.; Coops, N.C.; Neinavaz, E.; Ali, A.; Schaepman, M.E.; Paganini, M.; Kissling, W.D.; Vihervaara, P.; Darvishzadeh, R.; Feilhauer, H. Priority List of Biodiversity Metrics to Observe from Space. Nat. Ecol. Evol. 2021, 5, 896–906. [Google Scholar] [CrossRef] [PubMed]
- Pereira, H.M.; Ferrier, S.; Walters, M.; Geller, G.N.; Jongman, R.H.G.; Scholes, R.J.; Bruford, M.W.; Brummitt, N.; Butchart, S.H.M.; Cardoso, A.C. Essential Biodiversity Variables. Science 2013, 339, 277–278. [Google Scholar] [CrossRef] [PubMed]
- Epstein, H.; Armstrong, A.; Alcaraz-Segura, D.; Tassone, M.; Raynolds, M. Assessing the Controls on Ecosystem Functional Diversity in the Arctic Tundra at Circumpolar and Regional Scales. In Proceedings of the AGU Fall Meeting Abstracts, New Orleans, LA, USA, 13–17 December 2021; Volume 2021, p. B13A-01. [Google Scholar]
- Obando, V. Biodiversidad En Costa Rica: Estado Del Conocimiento y Gestión; INBio: Heredia, CR, USA, 2002. [Google Scholar]
- Herrera, W. Climate of Costa Rica. In Costa Rican Ecosystems; The University of Chicago Press: Chicago, IL, USA, 2016; pp. 19–29. [Google Scholar]
- Kappelle, M. Costa Rica’s Ecosystems: Setting the Stage. In Costa Rican Ecosystems; The University of Chicago Press: Chicago, IL, USA, 2016; p. 3. [Google Scholar]
- Steyn, D.G.; Moisseeva, N.; Harari, O.; Welch, W.J. Temporal and Spatial Variability of Annual Rainfall Patterns in Guanacaste, Costa Rica. Available online: https://open.library.ubc.ca/collections/52383/items/1.0340318 (accessed on 25 September 2023).
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km Spatial Resolution Climate Surfaces for Global Land Areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Rauscher, S.A.; Giorgi, F.; Diffenbaugh, N.S.; Seth, A. Extension and Intensification of the Meso-American Mid-Summer Drought in the Twenty-First Century. Clim. Dyn. 2008, 31, 551–571. [Google Scholar] [CrossRef]
- Hund, S.V.; Grossmann, I.; Steyn, D.G.; Allen, D.M.; Johnson, M.S. Changing Water Resources under El Niño, Climate Change, and Growing Water Demands in Seasonally Dry Tropical Watersheds. Water Resour. Res. 2021, 57, e2020WR028535. [Google Scholar] [CrossRef]
- Karp, D.S.; Frishkoff, L.O.; Echeverri, A.; Zook, J.; Juárez, P.; Chan, K.M. Agriculture Erases Climate-driven Β-diversity in Neotropical Bird Communities. Glob. Chang. Biol. 2018, 24, 338–349. [Google Scholar] [CrossRef]
- Amani, M.; Ghorbanian, A.; Ahmadi, S.A.; Kakooei, M.; Moghimi, A.; Mirmazloumi, S.M.; Moghaddam, S.H.A.; Mahdavi, S.; Ghahremanloo, M.; Parsian, S.; et al. Google Earth Engine Cloud Computing Platform for Remote Sensing Big Data Applications: A Comprehensive Review. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2020, 13, 5326–5350. [Google Scholar] [CrossRef]
- Huete, A.R.; Didan, K.; Shimabukuro, Y.E.; Ratana, P.; Saleska, S.R.; Hutyra, L.R.; Yang, W.; Nemani, R.R.; Myneni, R. Amazon Rainforests Green-up with Sunlight in Dry Season. Geophys. Res. Lett. 2006, 33. [Google Scholar] [CrossRef]
- Xiao, X.; Hollinger, D.; Aber, J.; Goltz, M.; Davidson, E.A.; Zhang, Q.; Moore, B. Satellite-Based Modeling of Gross Primary Production in an Evergreen Needleleaf Forest. Remote Sens. Environ. 2004, 89, 519–534. [Google Scholar] [CrossRef]
- Xiao, X.; Braswell, B.; Zhang, Q.; Boles, S.; Frolking, S.; Moore, B. Sensitivity of Vegetation Indices to Atmospheric Aerosols: Continental-Scale Observations in Northern Asia. Remote Sens. Environ. 2003, 84, 385–392. [Google Scholar] [CrossRef]
- Kappelle, M. Costa Rican Ecosystems; University of Chicago Press: Chicago, IL, USA, 2016. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, L.; Smith, J.R.; Armstrong, A.H.; Alcaraz-Segura, D.; Epstein, H.E.; Echeverri, A.; Langhans, K.E.; Schmitt, R.J.P.; Chaplin-Kramer, R. Influences of Satellite Sensor and Scale on Derivation of Ecosystem Functional Types and Diversity. Remote Sens. 2023, 15, 5593. https://doi.org/10.3390/rs15235593
Liu L, Smith JR, Armstrong AH, Alcaraz-Segura D, Epstein HE, Echeverri A, Langhans KE, Schmitt RJP, Chaplin-Kramer R. Influences of Satellite Sensor and Scale on Derivation of Ecosystem Functional Types and Diversity. Remote Sensing. 2023; 15(23):5593. https://doi.org/10.3390/rs15235593
Chicago/Turabian StyleLiu, Lingling, Jeffrey R. Smith, Amanda H. Armstrong, Domingo Alcaraz-Segura, Howard E. Epstein, Alejandra Echeverri, Kelley E. Langhans, Rafael J. P. Schmitt, and Rebecca Chaplin-Kramer. 2023. "Influences of Satellite Sensor and Scale on Derivation of Ecosystem Functional Types and Diversity" Remote Sensing 15, no. 23: 5593. https://doi.org/10.3390/rs15235593