Satellite Based Fraction of Absorbed Photosynthetically Active Radiation Is Congruent with Plant Diversity in India

 ,
,  ,
, 

Abstract
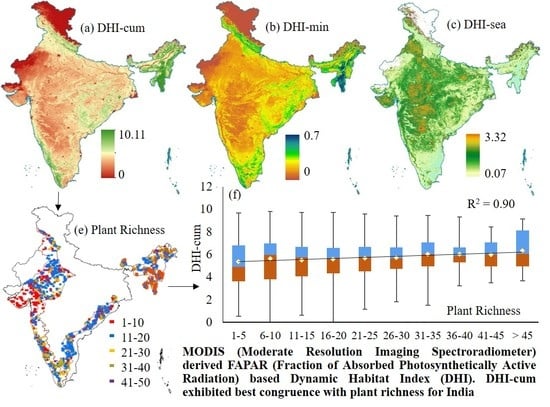
:
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Satellite Data
2.3. Generation of DHI Components and Their Composite
2.4. DHI Components and Plant Richness
3. Results
3.1. Visualizing Greenness Pattern Using DHI Components along Indian Biogeographic Regions
3.2. Significance of Greenness Variability from 2001 to 2015 Using DHI Components
3.3. Validation of DHI Components Using Plant Richness Data
4. Discussion
5. Conclusions
6. Highlights
- Characterized spatiotemporal variability of FAPAR-based DHI components (2001–2015) for India.
- Individual as well as composites of DHI components very well differentiated the biogeographic regions of India with high/low biodiversity levels.
- The inter-year correlation and regression of DHIs exhibited gradual decrease in vegetation greenness for Northeastern region, while the semi-arid and Deccan peninsular regions showed abrupt increase in vegetation greenness and seasonality.
- DHI-cum representing the annual greenness was strongly correlated with the plant richness thereby emerging as a suitable indicator to monitor the plant diversity.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Foley, J.A.; Levis, S.; Costa, M.H.; Cramer, W.; Pollard, D. Incorporating dynamic vegetation cover within global climate models. Ecol. Appl. 2000, 10, 1620–1632. [Google Scholar] [CrossRef]
- Cui, L.; Shi, J. Temporal and spatial response of vegetation NDVI to temperature and precipitation in eastern China. J. Geogr. Sci. 2010. [Google Scholar] [CrossRef]
- Wang, R.; Gamon, J.A. Remote sensing of terrestrial plant biodiversity. Remote Sens. Environ. 2019, 231, 111218. [Google Scholar] [CrossRef]
- Zhang, C.; Cai, D.; Guo, S.; Guan, Y.; Fraedrich, K.; Nie, Y.; Liu, X.; Bian, X. Spatial-temporal dynamics of China’s terrestrial biodiversity: A dynamic habitat index diagnostic. Remote Sens. 2016, 8, 227. [Google Scholar] [CrossRef] [Green Version]
- Erdelen, W.R. Shaping the Fate of Life on Earth: The Post-2020 Global Biodiversity Framework. Glob. Policy 2020. [Google Scholar] [CrossRef]
- Šímová, I.; Li, Y.M.; Storch, D. Relationship between species richness and productivity in plants: The role of sampling effect, heterogeneity and species pool. J. Ecol. 2013. [Google Scholar] [CrossRef]
- Jetz, W.; Cavender-Bares, J.; Pavlick, R.; Schimel, D.; Davis, F.W.; Asner, G.P.; Guralnick, R.; Kattge, J.; Latimer, A.M.; Moorcroft, P.; et al. Monitoring plant functional diversity from space. Nat. Plants 2016, 2, 16024. [Google Scholar] [CrossRef] [Green Version]
- Tucker, C.J.; Sellers, P.J. Satellite remote sensing of primary production. Int. J. Remote Sens. 1986, 7, 1395–1416. [Google Scholar] [CrossRef]
- Waring, R.H.; Running, S.W. Forest ecosystems: Analysis at multiple scales. Choice Rev. Online 1998. [Google Scholar] [CrossRef]
- Fensholt, R.; Sandholt, I.; Rasmussen, M.S.; Stisen, S.; Diouf, A. Evaluation of satellite based primary production modelling in the semi-arid Sahel. Remote Sens. Environ. 2006, 105, 173–188. [Google Scholar] [CrossRef]
- Anderson, R.G.; Canadell, J.G.; Randerson, J.T.; Jackson, R.B.; Hungate, B.A.; Baldocchi, D.D.; Ban-Weiss, G.A.; Bonan, G.B.; Caldeira, K.; Cao, L.; et al. Biophysical considerations in forestry for climate protection. Front. Ecol. Environ. 2011, 9, 174–182. [Google Scholar] [CrossRef] [Green Version]
- Jackson, R.B.; Randerson, J.T.; Canadell, J.G.; Anderson, R.G.; Avissar, R.; Baldocchi, D.D.; Bonan, G.B.; Caldeira, K.; Diffenbaugh, N.S.; Field, C.B.; et al. Protecting climate with forests. Environ. Res. Lett. 2008. [Google Scholar] [CrossRef]
- Oindo, B.O.; Skidmore, A.K. Interannual variability of NDVI and species richness in Kenya. Int. J. Remote Sens. 2002. [Google Scholar] [CrossRef] [Green Version]
- Huston, M.A. Biological diversity: The coexistence of species on changing landscapes. Biol. Divers. Coexistence Species Chang. Landsc. 1994. [Google Scholar] [CrossRef]
- Muchoney, D.M. Earth observations for terrestrial biodiversity and ecosystems. Remote Sens. Environ. 2008. [Google Scholar] [CrossRef]
- Thakur, T.K.; Padwar, G.K.; Patel, D.K.; Bijalwan, A. Monitoring land use, species composition and diversity of moist tropical environ in Achanakmaar Amarkantak Biosphere reserve, India using satellite data. Biodivers. Int. J. 2019. [Google Scholar] [CrossRef]
- Mahanand, S.; Behera, M.D. Relationship between Field-Based Plant Species Richness and Satellite-Derived Biophysical Proxies in the Western Ghats, India. Proc. Natl. Acad. Sci. India Sect. A Phys. Sci. 2017, 87. [Google Scholar] [CrossRef]
- Chitale, V.S.; Behera, M.D.; Roy, P.S. Deciphering plant richness using satellite remote sensing: A study from three biodiversity hotspots. Biodivers. Conserv. 2019. [Google Scholar] [CrossRef]
- Zhou, L.; Tucker, C.J.; Kaufmann, R.K.; Slayback, D.; Shabanov, N.V.; Myneni, R.B. Variations in northern vegetation activity inferred from satellite data of vegetation index during 1981 to 1999. J. Geophys. Res. Atmos. 2001, 106, 20069–20083. [Google Scholar] [CrossRef]
- Nemani, R.R.; Keeling, C.D.; Hashimoto, H.; Jolly, W.M.; Piper, S.C.; Tucker, C.J.; Myneni, R.B.; Running, S.W. Climate-driven increases in global terrestrial net primary production from 1982 to 1999. Science 2003. [Google Scholar] [CrossRef] [Green Version]
- Fensholt, R.; Langanke, T.; Rasmussen, K.; Reenberg, A.; Prince, S.D.; Tucker, C.; Scholes, R.J.; Le, Q.B.; Bondeau, A.; Eastman, R.; et al. Greenness in semi-arid areas across the globe 1981-2007 - an Earth Observing Satellite based analysis of trends and drivers. Remote Sens. Environ. 2012, 121, 144–158. [Google Scholar] [CrossRef]
- Chen, C.; Park, T.; Wang, X.; Piao, S.; Xu, B.; Chaturvedi, R.K.; Fuchs, R.; Brovkin, V.; Ciais, P.; Fensholt, R.; et al. China and India lead in greening of the world through land-use management. Nat. Sustain. 2019, 2, 122–129. [Google Scholar] [CrossRef]
- Mondal, P.; Jain, M.; Robertson, A.W.; Galford, G.L.; Small, C.; DeFries, R.S. Winter crop sensitivity to inter-annual climate variability in central India. Clim. Chang. 2014. [Google Scholar] [CrossRef] [Green Version]
- Mishra, N.B.; Mainali, K.P. Greening and browning of the Himalaya: Spatial patterns and the role of climatic change and human drivers. Sci. Total Environ. 2017, 587–588, 326–339. [Google Scholar] [CrossRef] [PubMed]
- Mishra, N.B.; Chaudhuri, G. Spatio-temporal analysis of trends in seasonal vegetation productivity across Uttarakhand, Indian Himalayas, 2000-2014. Appl. Geogr. 2015, 56, 29–41. [Google Scholar] [CrossRef]
- de Jong, R.; Verbesselt, J.; Schaepman, M.E.; de Bruin, S. Trend changes in global greening and browning: Contribution of short-term trends to longer-term change. Glob. Chang. Biol. 2012, 18, 642–655. [Google Scholar] [CrossRef]
- Murthy, K.; Bagchi, S. Spatial patterns of long-term vegetation greening and browning are consistent across multiple scales: Implications for monitoring land degradation. Land Degrad. Dev. 2018, 29, 2485–2495. [Google Scholar] [CrossRef]
- Chakraborty, A.; Seshasai, M.V.R.; Reddy, C.S.; Dadhwal, V.K. Persistent negative changes in seasonal greenness over different forest types of India using MODIS time series NDVI data (2001–2014). Ecol. Indic. 2018, 85, 887–903. [Google Scholar] [CrossRef]
- Rocchini, D. Effects of spatial and spectral resolution in estimating ecosystem α-diversity by satellite imagery. Remote Sens. Environ. 2007, 111, 423–434. [Google Scholar] [CrossRef]
- Mackey, B.; Bryan, J.; Randall, L. Australia’ s Dynamic Habitat Template for 2003; ANU Research Publications: Canberra, Australia, 2003. [Google Scholar]
- Coops, N.C.; Wulder, M.A.; Iwanicka, D. Demonstration of a satellite-based index to monitor habitat at continental-scales. Ecol. Indic. 2009, 9, 948–958. [Google Scholar] [CrossRef]
- Wright, D.H. Species-Energy Theory: An Extension of Species-Area Theory. Oikos 1983. [Google Scholar] [CrossRef] [Green Version]
- Currie, D.J.; Mittelbach, G.G.; Cornell, H.V.; Field, R.; Guégan, J.F.; Hawkins, B.A.; Kaufman, D.M.; Kerr, J.T.; Oberdorff, T.; O’Brien, E.; et al. Predictions and tests of climate-based hypotheses of broad-scale variation in taxonomic richness. Ecol. Lett. 2004, 7, 1121–1134. [Google Scholar] [CrossRef]
- Hurlbert, A.H. Linking species-area and species-energy relationships in Drosophila microcosms. Ecol. Lett. 2006. [Google Scholar] [CrossRef] [PubMed]
- Connell, J.H.; Orias, E. The Ecological Regulation of Species Diversity. Am. Nat. 1964. [Google Scholar] [CrossRef]
- Hu, G.; Jin, Y.; Liu, J.; Yu, M. Functional diversity versus species diversity: Relationships with habitat heterogeneity at multiple scales in a subtropical evergreen broad-leaved forest. Ecol. Res. 2014. [Google Scholar] [CrossRef]
- Mason, N.W.H.; Mouillot, D.; Lee, W.G.; Wilson, J.B. Functional richness, functional evenness and functional divergence: The primary components of functional diversity. Oikos 2005. [Google Scholar] [CrossRef]
- Williams, S.E.; Middleton, J. Climatic seasonality, resource bottlenecks, and abundance of rainforest birds: Implications for global climate change. Divers. Distrib. 2008. [Google Scholar] [CrossRef]
- Coops, N.C.; Fontana, F.M.A.; Harvey, G.K.A.; Nelson, T.A.; Wulder, M.A. Monitoring of a national-scale indirect indicator of biodiversity using a long time-series of remotely sensed imagery. Can. J. Remote Sens. 2014. [Google Scholar] [CrossRef]
- Nelson, T.A.; Coops, N.C.; Wulder, M.A.; Perez, L.; Fitterer, J.; Powers, R.; Fontana, F. Predicting climate change impacts to the canadian boreal forest. Diversity 2014, 6, 133–157. [Google Scholar] [CrossRef] [Green Version]
- Berry, S.; Mackey, B.; Brown, T. Potential applications of remotely sensed vegetation greenness to habitat analysis and the conservation of dispersive fauna. In Proceedings of the Pacific Conservation Biology. 2007. Available online: https://www.publish.csiro.au/PC/PC070120 (accessed on 4 January 2021).
- Berry, S.L.; Roderick, M.L. Estimating mixtures of leaf functional types using continental-scale satellite and climatic data. Glob. Ecol. Biogeogr. 2002. [Google Scholar] [CrossRef]
- Cramer, M.J.; Willig, M.R. Habitat heterogeneity, species diversity and null models. Oikos 2005. [Google Scholar] [CrossRef]
- Turner, M.G.; Donato, D.C.; Romme, W.H. Consequences of spatial heterogeneity for ecosystem services in changing forest landscapes: Priorities for future research. Landsc. Ecol. 2013. [Google Scholar] [CrossRef]
- Barik, S.K.; Behera, M.D. Studies on ecosystem function and dynamics in Indian sub-continent and emerging applications of satellite remote sensing technique. Trop. Ecol. 2020, 61, 1–4. [Google Scholar] [CrossRef]
- Zhao, M.; Heinsch, F.A.; Nemani, R.R.; Running, S.W. Improvements of the MODIS terrestrial gross and net primary production global data set. Remote Sens. Environ. 2005. [Google Scholar] [CrossRef]
- Heinsch, F.A.; Zhao, M.; Running, S.W.; Kimball, J.S.; Nemani, R.R.; Davis, K.J.; Bolstad, P.V.; Cook, B.D.; Desai, A.R.; Ricciuto, D.M.; et al. Evaluation of remote sensing based terrestrial productivity from MODIS using regional tower eddy flux network observations. IEEE Trans. Geosci. Remote Sens. 2006. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Zhang, Y.; Knyazikhin, Y.; Myneni, R.B.; Glassy, J.M.; Dedieu, G.; Running, S.W. Prototyping of MODIS LAI and FPAR algorithm with LASUR and LANDSAT data. IEEE Trans. Geosci. Remote Sens. 2000. [Google Scholar] [CrossRef] [Green Version]
- Coops, N.C.; Wulder, M.A.; Duro, D.C.; Han, T.; Berry, S. The development of a Canadian dynamic habitat index using multi-temporal satellite estimates of canopy light absorbance. Ecol. Indic. 2008. [Google Scholar] [CrossRef]
- Roy, P.S.; Kushwaha, S.P.S.; Murthy, M.S.R.; Roy, A.; Kushwaha, D.; Reddy, C.S.; Behera, M.D.; Mathur, V.B.; Padalia, H.; Saran, S.; et al. Biodiversity Characterisation at Landscape Level: National Assessment; Indian Institute of Remote Sensing: Dehradun, India, 2012; ISBN 8190141880. [Google Scholar]
- Tripathi, P.; Dev Behera, M.; Roy, P.S. Optimized grid representation of plant species richness in India-Utility of an existing national database in integrated ecological analysis. PLoS ONE 2017, 12, e0173774. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, M.D.; Ahas, R.; Aasa, A. Onset of spring starting earlier across the Northern Hemisphere. Glob. Chang. Biol. 2006. [Google Scholar] [CrossRef]
- Gaston, K.J.; Blackburn, T.M. Pattern and Process in Macroecology. 2007. Available online: https://www.wiley.com/en-ag/Pattern+and+Process+in+Macroecology-p-9780470999592 (accessed on 4 January 2021).
- Kleijn, D.; Rundlöf, M.; Scheper, J.; Smith, H.G.; Tscharntke, T. Does conservation on farmland contribute to halting the biodiversity decline? Trends Ecol. Evol. 2011, 26, 474–481. [Google Scholar] [CrossRef]
- Negi, V.S.; Joshi, B.C.; Pathak, R.; Rawal, R.S.; Sekar, K.C. Assessment of fuelwood diversity and consumption patterns in cold desert part of Indian Himalaya: Implication for conservation and quality of life. J. Clean. Prod. 2018, 196, 23–31. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Cabello, D.; Tomás-Burguera, M.; Martín-Hernández, N.; Beguería, S.; Azorin-Molina, C.; Kenawy, A. El Drought variability and land degradation in semiarid regions: Assessment using remote sensing data and drought indices (1982-2011). Remote Sens. 2015, 7, 4391–4423. [Google Scholar] [CrossRef] [Green Version]
- MEA. Ecosystems and Human Well-Being. Synthesis. 2005. Available online: http://www.bioquest.org/wp-content/blogs.dir/files/2009/06/ecosystems-and-health.pdf (accessed on 4 January 2021).
- Saikia, A. NDVI variability in North East India. Scottish Geogr. J. 2009. [Google Scholar] [CrossRef]
- Roy, P.S.; Roy, A.; Joshi, P.K.; Kale, M.P.; Srivastava, V.K.; Srivastava, S.K.; Dwevidi, R.S.; Joshi, C.; Behera, M.D.; Meiyappan, P.; et al. Development of decadal (1985-1995-2005) land use and land cover database for India. Remote Sens. 2015, 7, 2401–2430. [Google Scholar] [CrossRef] [Green Version]
- Pasha, S.V.; Behera, M.D.; Mahawar, S.K.; Barik, S.K.; Joshi, S.R. Assessment of shifting cultivation fallows in Northeastern India using Landsat imageries. Trop. Ecol. 2020. [Google Scholar] [CrossRef]
- Roy, P.S.; Behera, M.D. Assessment of biological richness in different altitudinal zones in the Eastern Himalayas, Arunachal Pradesh, India. Curr. Sci. 2005, 88, 250–257. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BG Regions | DHI-Cum | DHI-Min | DHI-Sea |
|---|---|---|---|
| Semi-Arid | 2.15–3.54 | 0.15–0.27 | 0.27–0.44 |
| Eastern Ghats | 3.43–4.9 | 0.28–0.48 | 0.2–0.31 |
| Western Ghats | 3.83–5.1 | 0.31–0.52 | 0.21–0.27 |
| Northeast | 4.5–6.0 | 0.33–0.61 | 0.21–0.32 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahanand, S.; Behera, M.D.; Roy, P.S.; Kumar, P.; Barik, S.K.; Srivastava, P.K. Satellite Based Fraction of Absorbed Photosynthetically Active Radiation Is Congruent with Plant Diversity in India. Remote Sens. 2021, 13, 159. https://doi.org/10.3390/rs13020159
Mahanand S, Behera MD, Roy PS, Kumar P, Barik SK, Srivastava PK. Satellite Based Fraction of Absorbed Photosynthetically Active Radiation Is Congruent with Plant Diversity in India. Remote Sensing. 2021; 13(2):159. https://doi.org/10.3390/rs13020159
Chicago/Turabian StyleMahanand, Swapna, Mukunda Dev Behera, Partha Sarathi Roy, Priyankar Kumar, Saroj Kanta Barik, and Prashant Kumar Srivastava. 2021. "Satellite Based Fraction of Absorbed Photosynthetically Active Radiation Is Congruent with Plant Diversity in India" Remote Sensing 13, no. 2: 159. https://doi.org/10.3390/rs13020159