
Seed Burial Depth and Soil Water Content Affect Seedling Emergence and Growth of Ulmus pumila var. sabulosa in the Horqin Sandy Land


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Experimental Procedures
2.3. Determination of Seed Germination and Seedling Emergence
2.4. Statistical Analysis
3. Results
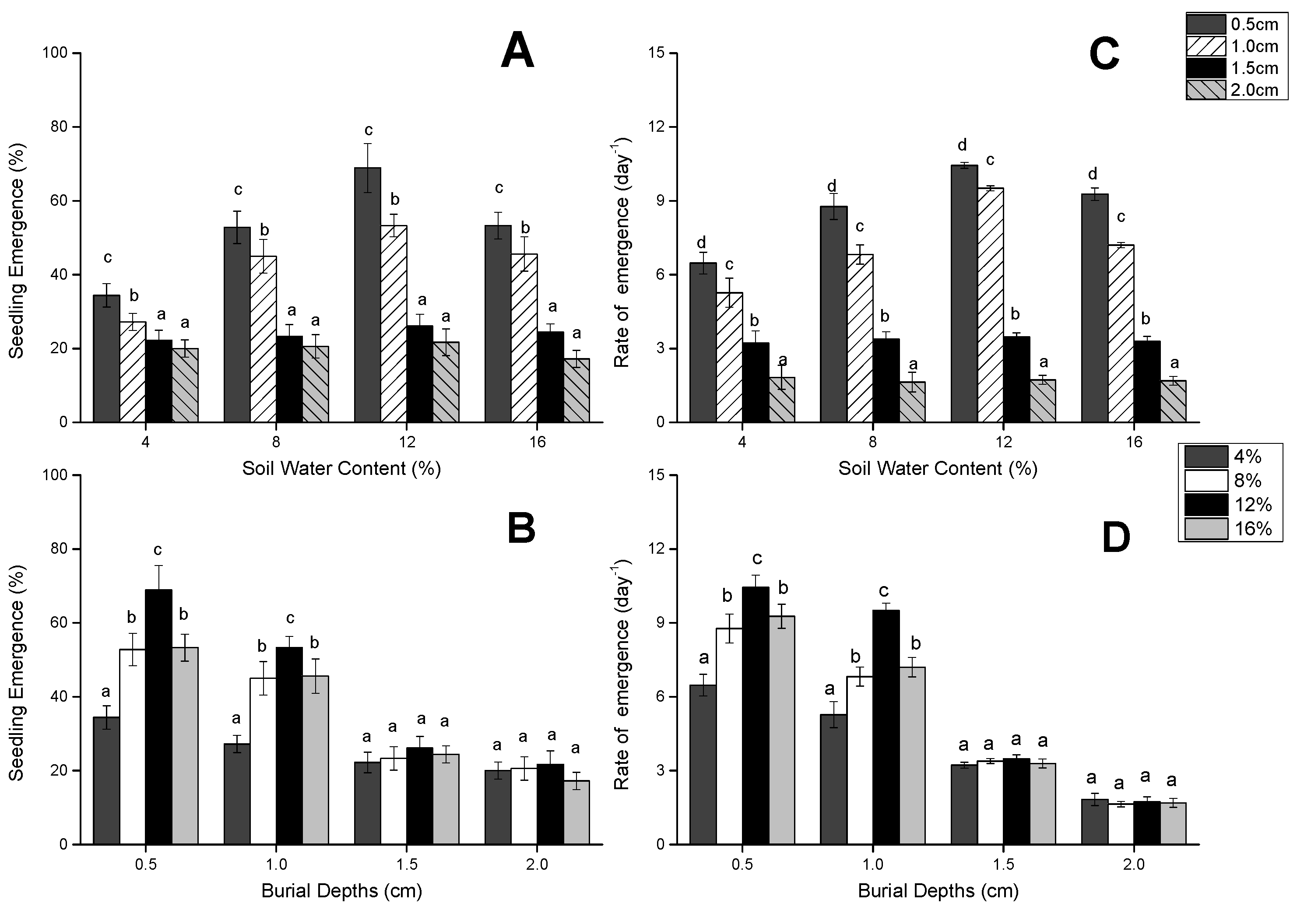
3.1. Seedling Emergence and Speed of Seedling Emergence

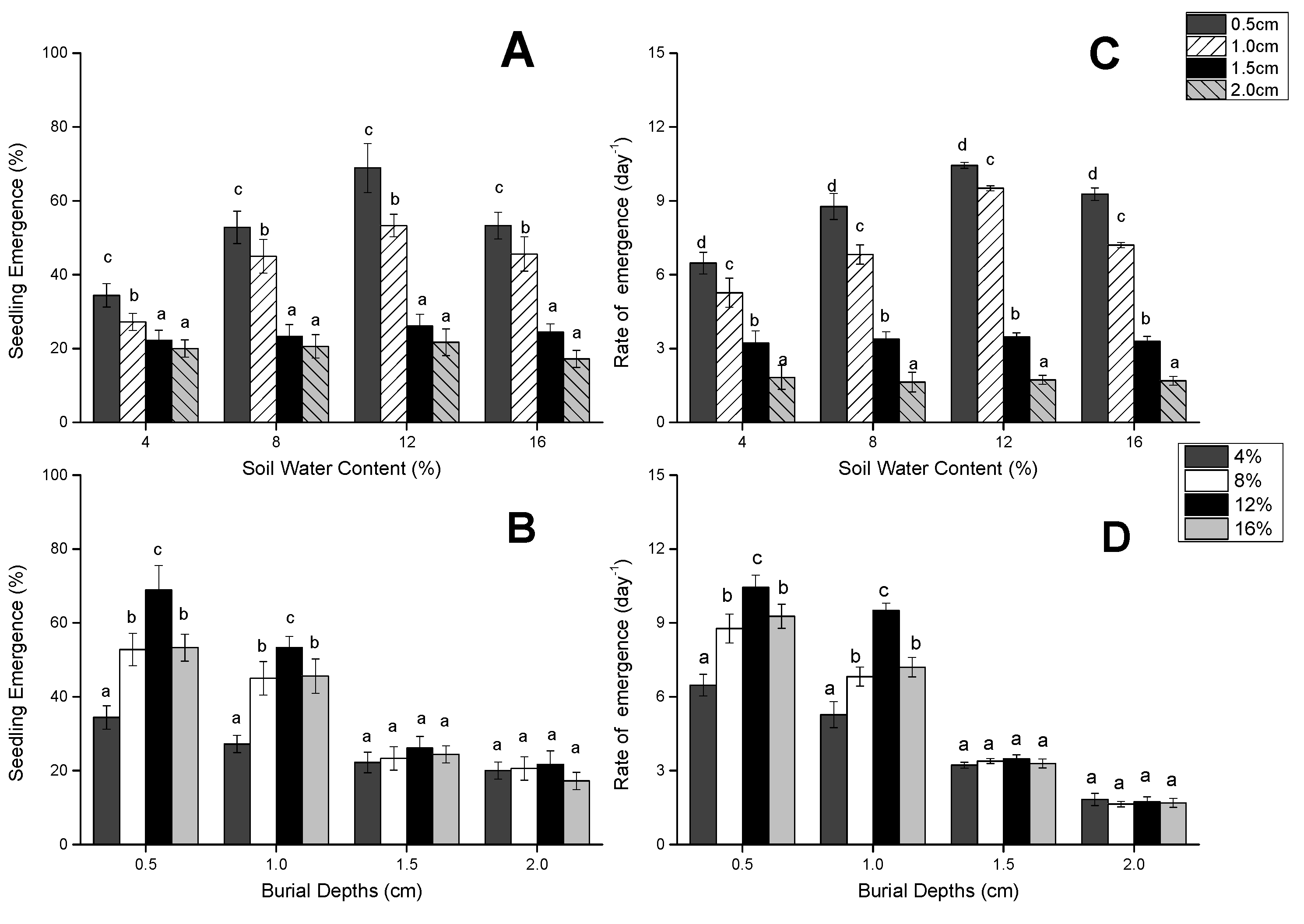{kind=link}
{kind=link}
{kind=link}
| Source of Variation | DF | Seedling Emergence Percentage (%) | Rate of Seedling Emergence (Day−1) | ||
|---|---|---|---|---|---|
| F-Value | p-Value | F-Value | p-Value | ||
| Depth of seed burial | 3 | 137.470 | 0 | 201.103 | 0 |
| Soil water content | 3 | 26.371 | <0.001 | 13.645 | <0.001 |
| Interaction | 9 | 6.276 | <0.001 | 4.459 | 0 |
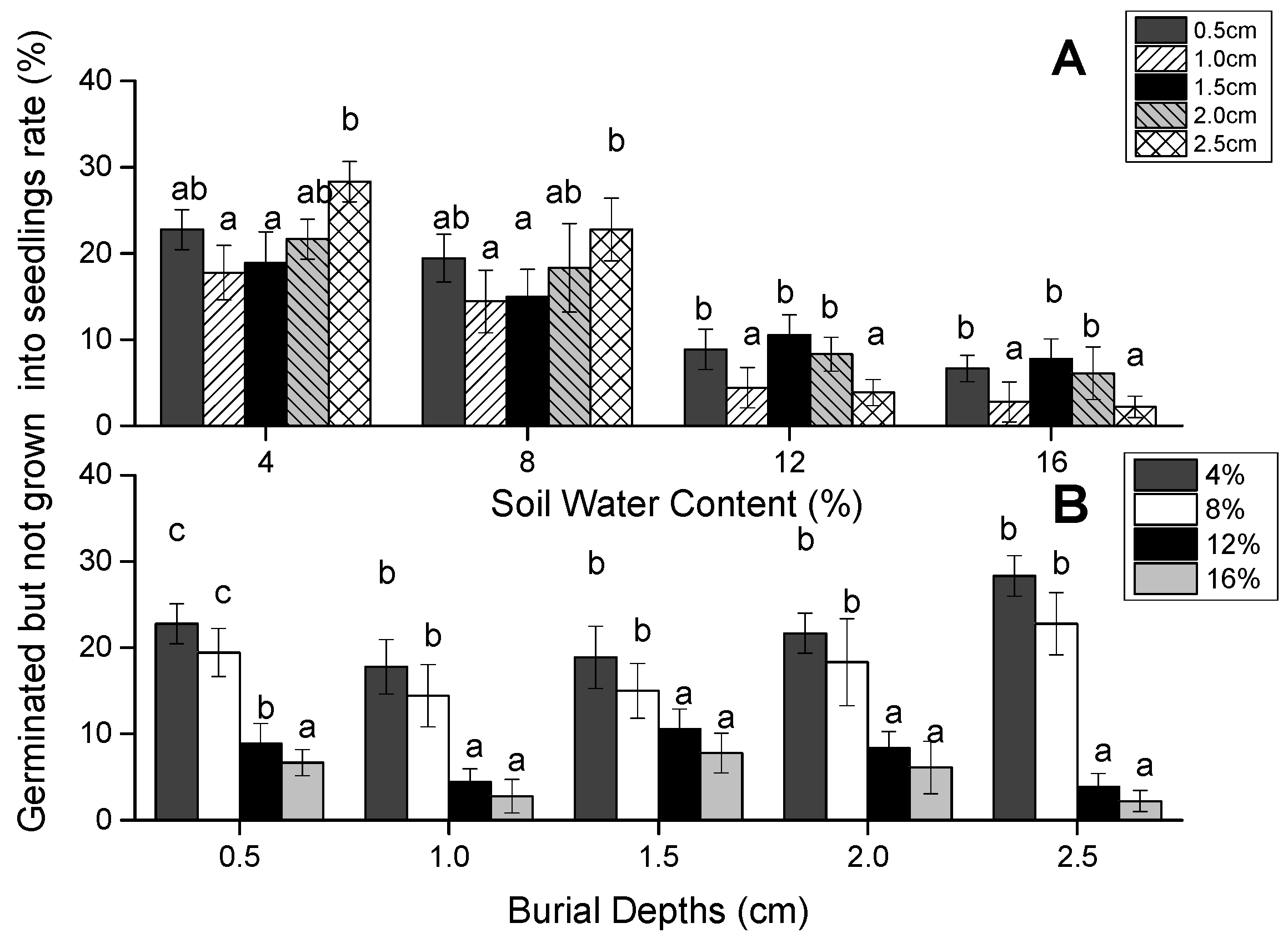
3.2. Germinated Seeds Which Did Not Grown into Seedlings
| Source of Variation | Germinated Seeds Which Did Not Grow into Seedlings (%) | |||
|---|---|---|---|---|
| DF | Mean Square | F-Value | p-Value | |
| Depth of seed burial | 4 | 65.922 | 6.631 | <0.001 |
| Soil water content | 3 | 1403.936 | 141.219 | 0 |
| Interaction | 12 | 45.198 | 4.546 | <0.001 |
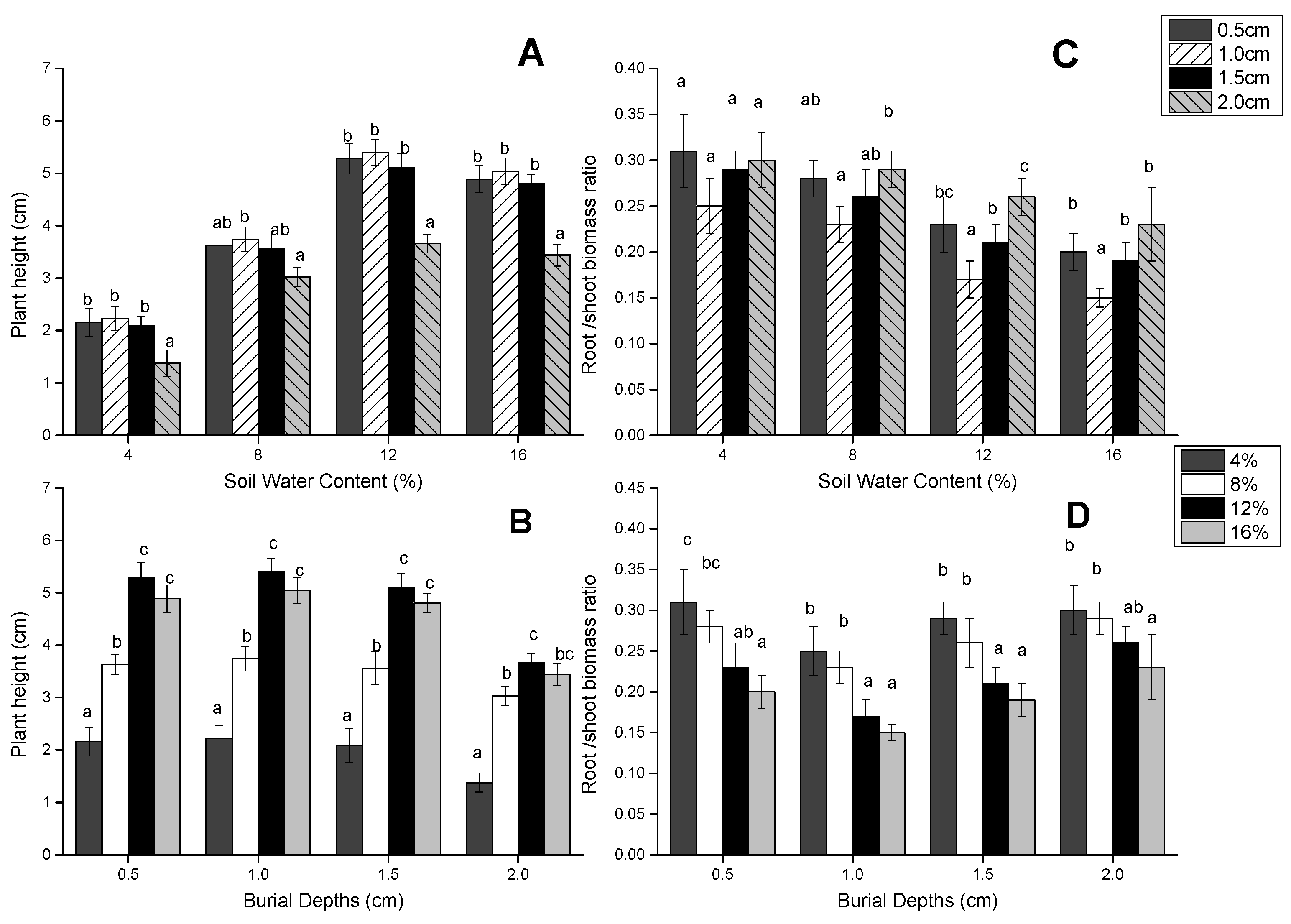
3.3. Seedling Height and Root/Shoot Ratio
| Source of Variation | DF | Seedling Height | Root/Shoot Ratio | ||
|---|---|---|---|---|---|
| F-Value | p-Value | F-Value | p-Value | ||
| Depth of seed burial | 3 | 50.344 | 0 | 20.271 | <0.001 |
| Soil water content | 3 | 279.845 | 0 | 42.422 | 0 |
| Interaction | 9 | 2.243 | 0.024 | 4.124 | 0.04 |
4. Discussion
4.1. Seed Germination, Seedling Emergence and Speed of Seedling Emergence
4.2. Seedling Height and Root/Shoot Ratio
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Peng, S.; Huang, Z.Y.; Peng, S.L.; Ou Yang, X.; Xu, G. Factors influencing mortality of seed and seedling in plant nature regeneration process. Guihaia 2004, 24, 113–124. [Google Scholar]
- Zheng, M.Q.; Zheng, Y.R.; Jiang, L.H. Effects of one-time water supply and sand burial on seed germination and seedling emergence of four populary psammophyte in Mu Us sandy land. Acta Ecol. Sin. 2006, 26, 2474–2484. [Google Scholar]
- Du, X.F.; Guo, Q.F.; Gao, X.M.; Ma, K.P. Seed rain, soil seed bank, seed loss and regeneration of Castanopsis fargesii (Fagaceae) in a subtropical evergreen broad-leaved forest. For. Ecol. Manag. 2007, 238, 212–219. [Google Scholar] [CrossRef]
- Su, H.; Li, Y.G.; Liu, W.; Xu, H.; Sun, J.X. Changes in water use with growth in Ulmus pumila in semiarid sandy land of northern China. Trees 2013, 28, 41–52. [Google Scholar] [CrossRef]
- Li, Y.G.; Jiang, G.M.; Liu, M.Z.; Niu, S.L.; Gao, L.M.; Cao, X.C. Photosynthetic response to precipitation/rainfall in predominant tree (Ulmus pumila) seedlings in Hunshandak Sandland, China. Photosynthetica 2007, 45, 133–138. [Google Scholar] [CrossRef]
- Jiang, D.M.; Liu, Z.M.; Cao, C.Y; Kou, Z.W.; Wang, R.Y. Desertification and Ecological Restoration of Keerqin Sandy Land; China Environmental Science Press: Beijing, China, 2003. [Google Scholar]
- Liu, Z.M. Plant Regenerative Strategies; China Meteorological Press: Beijing, China, 2010. [Google Scholar]
- Liu, Z.M.; Yan, Q.L.; Li, X.L.; Ma, J.L.; Ling, X. Seed mass and shape, germination and plant abundance in a desertified grassland in northeastern Inner Mongolia. J. Arid Environ. 2007, 69, 198–211. [Google Scholar] [CrossRef]
- Maun, M.A. Seed-Germination and Seedling Establishment of Calamovilfa-Longifolia on Lake Huron Sand Dunes. Can. J. Bot. 1981, 59, 460–469. [Google Scholar] [CrossRef]
- Maun, M.A. Adaptations of plants to burial in coastal sand dunes. Can.J. Bot. 1998, 76, 713–738. [Google Scholar]
- Tang, J.; Jiang, D.M.; Wang, Y.C. A review on the process of seed-seedling regeneration of Ulmus pumila in sparse forest grassland. Chin.J. Ecol. 2014, 33, 1114–1120. [Google Scholar]
- Dulamsuren, C.; Hauck, M.; Nyambayar, S.; Bader, M.; Osokhjargal, D.; Oyungerel, S.; Leuschner, C. Performance of Siberian elm (Ulmus pumila) on steppe slopes of the northern Mongolian mountain taiga: Drought Stress and Herbivory in Mature Trees. Environ. Exp. Bot. 2009, 66, 18–24. [Google Scholar] [CrossRef]
- Ma, C.G. A Provenance Test of White Elm (Ulmus-Pumila L) in China. Silvae Genet. 1989, 38, 37–44. [Google Scholar]
- Wesche, K.; Walther, D.; von Wehrden, H.; Hensen, I. Trees in the desert: Reproduction and Genetic Structure of Fragmented Ulmus pumila forests in Mongolian drylands. Flora-Morphol. Distrib. Funct. Ecol. 2011, 206, 91–99. [Google Scholar] [CrossRef]
- Dong, M.; Alaten, B. Clonal plasticity in response to rhizome severing and heterogeneous resource supply in the rhizomatous grass Psammochloa villosa in an Inner Mongolian dune. China. Plant. Ecol. Plant. 1999, 141, 53–58. [Google Scholar] [CrossRef]
- Shi, L.; Zhang, Z.J.; Zhang, C.Y.; Zhang, J.Z. Effects of sand burial on survival, growth, gas exchange and biomass allocation of Ulmus pumila seedlings in the Hunshandak Sandland. Ann. Bot. 2004, 94, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Liu, Z.; Lü, X.T.; Maestre, F.T.; Wang, L.X. Sand burial compensates for the negative effects of erosion on the dune-building shrub Artemisia wudanica. Plant. Soil 2013, 374, 263–273. [Google Scholar] [CrossRef] [Green Version]
- Jiang, D.M.; Tang, Y.; Busso, C.A. Effects of vegetation cover on recruitment of Ulmus pumila L. in Horqin Sandy Land. J. Arid Land. 2013, 6, 343–351. [Google Scholar] [CrossRef]
- Dulamsuren, C.; Hauck, M.; Nyambayar, S.; Osokhjargal, D.; Leuschner, C. Establishment of Ulmus pumila seedlings on steppe slopes of the northern Mongolian mountain taiga. Acta Oecologica 2009, 35, 563–572. [Google Scholar] [CrossRef]
- Tang, Y.; Jiang, D.M.; Lü, X.T. Effects of Exclosure Management on Elm (Ulmus Pumila) Recruitment in Horqin Sandy Land. Arid Land Res. Manag. 2013, 28, 109–117. [Google Scholar] [CrossRef]
- Aguirre, L.; Johnson, D.A. Root morphological development in relation to shoot growth in seedlings of four range grasses. J. Range Manag. 1991, 44, 341–346. [Google Scholar] [CrossRef]
- Aguirre, L.; Johnson, D.A. Influence of temperature and cheatgrass competition on seedling development of two bunchgrasses. J. Range Manag. 1991, 44, 347–354. [Google Scholar] [CrossRef]
- Allen, M.F.; Hipps, L.E.; Woolridge, G.L. Wind dispersal and subsequent establishment of VA mycorrhizal fungi across a successional arid landscape. Landsc. Ecol. 1989, 2, 165–171. [Google Scholar] [CrossRef]
- Brown, R.W. The water relation of range plants: Adaptions to Water Deficits. In Wildland Plants:Physiological Ecology and Developmental Morphology; Bedunahy, D.J., Sosebee, R.E., Eds.; Society for Range Management: Littleton, CO, USA, 1995; pp. 219–413. [Google Scholar]
- Soriano, S.D.; Marti, V.P. Soil moisture determination at field capacity. In Edaphology and Climatology; Politenic University of Valencia: Alfaomega, Spain, 2004. [Google Scholar]
- Rozema, J. The influemce of salinity, inundation and temperature on germination of some halophytes and non-halophytes. Oecol. Plant. 1975, 10, 341–353. [Google Scholar]
- Sokal, R.R.; Rohlf, E.J. Biometry: The Principles and Practice of Statistics in Biological Research, 3rd ed.; W. H. Freeman: New York, NY, USA, 1995. [Google Scholar]
- Zhu, Y.J.; Yang, X.J.; Baskin, C.C.; Baskin, J.M.; Dong, M.; Huang, Z.Y. Effects of amount and frequency of precipitation and sand burial on seed germination, seedling emergence and survival of the dune grass Leymus secalinus in semiarid. Plant. Soil. 2014, 374, 399–409. [Google Scholar] [CrossRef]
- Nie, C.L.; Zheng, Y.R. Effects of water supply and sand burial on seed germination and seedling emergence of four dominant psammophytes in the ordos plateau. Acta Ecol. Sin. 2005, 29, 32–41. [Google Scholar]
- Chang, W.; Wu, J.G.; Liu, Y.H. Research advance in seed germination of desert woody plants. Chin. J. Appl. Ecol. 2007, 18, 436–444. [Google Scholar]
- Seiwa, K.; Watanabe, A.; Saitoh, T.; Kannu, H.; Akasaka, S. Effects of burying depth and seed size on seedling establishment of Japanese chestnuts, Castanea crenata. Forest Ecol. Manag. 2002, 164, 149–156. [Google Scholar] [CrossRef]
- Liu, H.J.; Guo, K. The impacts of sand burial on seedling development of Caragana intermedia. Acta Ecol. Sin. 2005, 25, 2550–2555. [Google Scholar]
- Baskin, C.C.; Baskin, J.M. Seeds Ecology, Biogeography and Evolution of Dormancy and Germination; Academic Press: San Diego, CA, USA, 1998. [Google Scholar]
- Huang, Z.Y.; Gutterman, Y. Comparison of germination strategies of Artemisia ordosica with its two congeners from deserts of China and Israel. Acta Ecol. Sin. 2000, 42, 71–80. [Google Scholar]
- Huang, Z.Y.; Gutterman, Y.; Osborne, D.J. Value of the mucilaginous pellicle to seeds of the sand-stabilizing desert woody shrub Artemisia sphaerocephala (Asteraceae). Trees-Struct Funct. 2004, 18, 669–676. [Google Scholar] [CrossRef]
- Liu, G.J.; Zhang, X.M.; Li, J.G.; Fan, D.D.; Deng, C.Z.; Hou, J.G.; Xin, R.M. Effects of Water Supply and Sand Burial on Seed Germination and Seedling Emergence of Haloxylon Ammodendronand Haloxylon Persicum. J. Desert Res. 2010, 30, 1085–1091. [Google Scholar]
- Huang, Z.Y.; Gutterman, Y. Artemisia monosperma achene germination in sand: Effects of Sand Depth, Sand/Water Content, Cyanobacterial Sand Crust and Temperature. J. Arid Environ. 1998, 38, 27–43. [Google Scholar] [CrossRef]
- Cheplick, G.P.; Grandstaff, K. Effects of sand burial on purple sandgrass (Triplasis purpurea): The Significance of Seed Heteromorphism. Plant Ecol. 1997, 133, 79–89. [Google Scholar] [CrossRef]
- Sun, Z.G.; Mou, X.J.; Lin, G.H.; Wang, L.L.; Song, H.Q.; Jiang, H.H. Effects of sediment burial disturbance on seedling survival and growth of Suaeda salsa in the tidal wetland of the Yellow River estuary. Plant. Soil. 2010, 337, 457–468. [Google Scholar] [CrossRef]
- Zhu, Y.; Dong, M.; Huang, Z.Y. Effects of sand burial and seed size on seed germination and seedling emergence of Psammochola villosa. Acta Ecol. Sin. 2005, 29, 730–739. [Google Scholar]
- Wang, J.H.; Chai, Y.F.; Zhang, Y. Effects of sand burial depth of Achnatherum inebrians seed on its germination and seedling growth. Chin. J. Ecol. 2010, 29, 324–328. [Google Scholar]
- Yang, H.L.; Cao, Z.P.; Dong, M.; Ye, Y.Z.; Huang, Z.Y. Effects of sand burying on caryopsis germination and seedling growth of Bromus inermis Leyss. Chin. J. Appl Ecol. 2007, 18, 2438–2443. [Google Scholar]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, J.; Busso, C.A.; Jiang, D.; Wang, Y.; Wu, D.; Musa, A.; Miao, R.; Miao, C. Seed Burial Depth and Soil Water Content Affect Seedling Emergence and Growth of Ulmus pumila var. sabulosa in the Horqin Sandy Land. Sustainability 2016, 8, 68. https://doi.org/10.3390/su8010068
Tang J, Busso CA, Jiang D, Wang Y, Wu D, Musa A, Miao R, Miao C. Seed Burial Depth and Soil Water Content Affect Seedling Emergence and Growth of Ulmus pumila var. sabulosa in the Horqin Sandy Land. Sustainability. 2016; 8(1):68. https://doi.org/10.3390/su8010068
Chicago/Turabian StyleTang, Jiao, Carlos A. Busso, Deming Jiang, Yongcui Wang, Dafu Wu, Ala Musa, Renhui Miao, and Chunping Miao. 2016. "Seed Burial Depth and Soil Water Content Affect Seedling Emergence and Growth of Ulmus pumila var. sabulosa in the Horqin Sandy Land" Sustainability 8, no. 1: 68. https://doi.org/10.3390/su8010068