Increasing Woody Species Diversity for Sustainable Limestone Quarry Reclamation in Canada

Abstract
:1. Introduction
2. Results and Discussion
2.1. Survival and Growth of Out Planted Trees and Shrubs

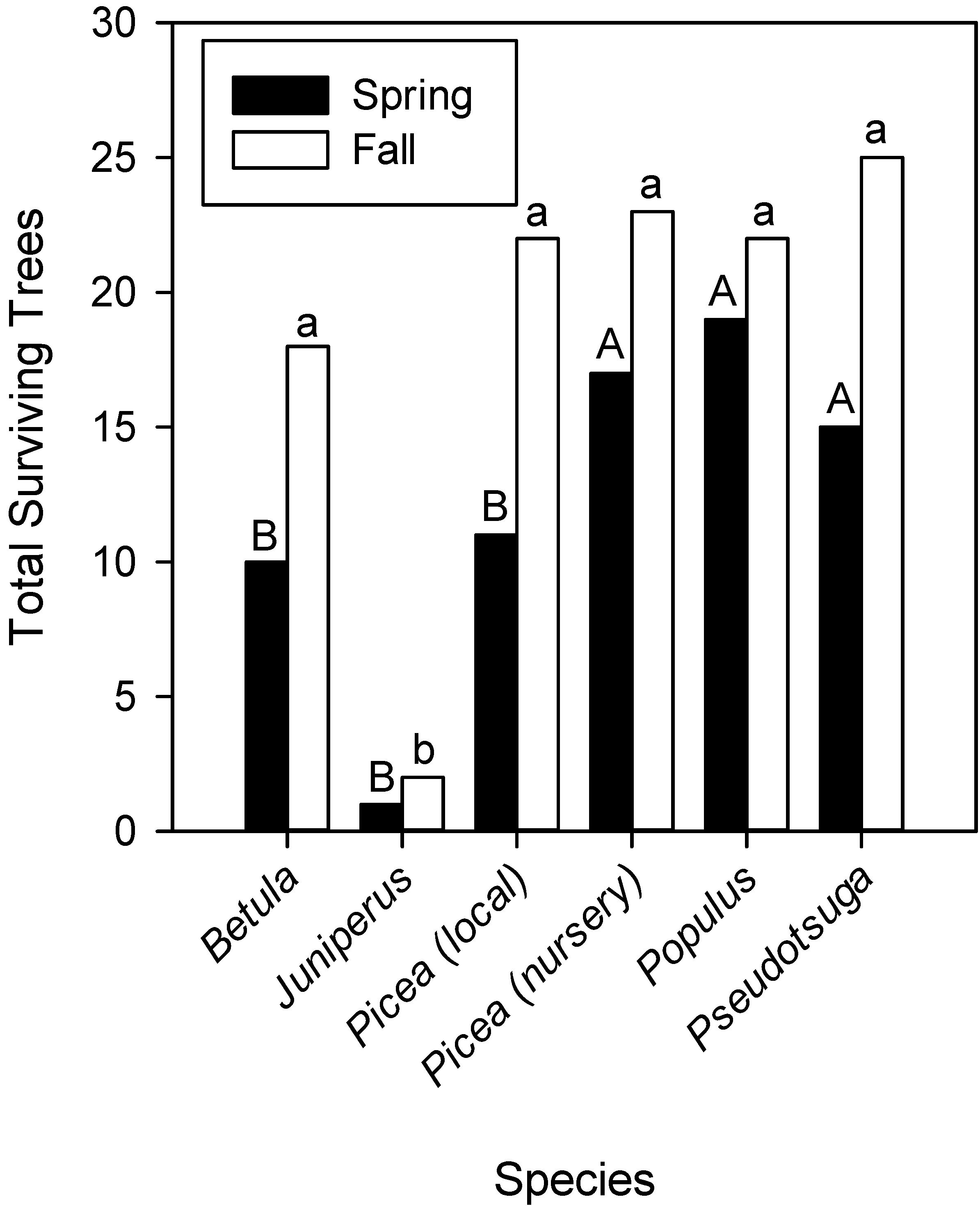{kind=link}
| 2007 | 2008 | 2009 | 2010 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| October | May | August | May | August | August | |||||||
| S | F | S | F | S | F | S | F | S | F | S | F | |
| Betula papyrifera | 21 | 26 | 18 | 25 | 18 | 24 | 12 | 24 | 10 | 23 | 10 b | 18 a |
| Juniperus horizontalis | 3 | 6 | 2 | 2 | 1 | 2 | 1 | 2 | 1 | 2 | 1 | 2 |
| Picea glauca (local) | 13 b | 27 a | 12 | 26 | 12 | 24 | 12 | 23 | 11 | 23 | 11 b | 22 a |
| Picea glauca (nursery) | 17 b | 30 a | 17 | 27 | 17 | 24 | 17 | 23 | 17 | 23 | 17 | 23 |
| Populus tremuloides | 21 b | 29 a | 20 | 29 | 19 | 26 | 19 | 23 | 19 | 22 | 19 | 22 |
| Pseudotsuga menziesii | 16 b | 29 a | 16 | 25 | 16 | 25 | 16 | 25 | 16 | 25 | 15 | 25 |
| Spring | Fall | ||||
|---|---|---|---|---|---|
| Monitoring Dates | Explanatory Variable | Residual Deviance | AIC | Residual Deviance | AIC |
| October 2007 | Species | 206.7 | 218.0 | 90.6 | 104.6 |
| May 2008 | 214.4 | 226.4 | 120.4 | 134.4 | |
| August 2008 | 209.8 | 221.8 | 152.0 | 166.0 | |
| May 2009 | 206.7 | 218.7 | 166.5 | 180.5 | |
| August 2009 | 206.6 | 218.6 | 171.3 | 185.3 | |
| August 2010 | 206.7 | 218.7 | 181.3 | 195.3 | |
| October 2007 | Soil treatment | 248.0 | 254.0 | 170.8 | 176.8 |
| May 2008 | 247.6 | 253.6 | 204.5 | 210.5 | |
| August 2008 | 247.1 | 253.1 | 220.2 | 226.2 | |
| May 2009 | 244.3 | 250.3 | 226.7 | 232.7 | |
| August 2009 | 242.3 | 248.3 | 229.9 | 235.9 | |
| August 2010 | 241.8 | 247.8 | 236.1 | 242.1 | |

| 2007 | 2008 | 2009 | 2010 | Final | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| October | May | August | May | August | August | |||||||||
| S | F | S | F | S | F | S | F | S | F | S | F | S | F | |
| Betula papyrifera | 30 | 13 | 10 | 3 | 0 | 3 | 20 | 0 | 7 | 3 | 0 | 17 | 67 | 40 |
| Juniperus horizontalis | 90 | 80 | 3 | 13 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 97 | 93 |
| Picea glauca (local) | 57 | 10 | 3 | 3 | 0 | 7 | 0 | 3 | 3 | 0 | 0 | 3 | 63 | 27 |
| Picea glauca (nursery) | 43 | 0 | 0 | 10 | 0 | 10 | 0 | 3 | 0 | 0 | 0 | 0 | 43 | 23 |
| Populus tremuloides | 30 | 3 | 3 | 0 | 3 | 10 | 0 | 10 | 0 | 3 | 0 | 0 | 37 | 27 |
| Pseudotsuga menziesii | 47 | 3 | 0 | 13 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 50 | 17 |
| Planting Season | Species | Number of Survivors | Number with Height Increase | % Trees with Increased Height | Mean Height Increase (cm) | Range of Increase from Individual Plants (cm) |
|---|---|---|---|---|---|---|
| Spring | Betula papyrifera | 10 | 4 | 40 | 2.9 | 1–5 |
| Juniperus horizontalis | 1 | 1 | 100 | 2.2 | 1.5–3 | |
| Picea glauca (local) | 11 | 10 | 91 | 4.2 | 1–9 | |
| Picea glauca (nursery) | 17 | 14 | 82 | 4.4 | 1–17 | |
| Populus tremuloides | 19 | 9 | 47 | 4.8 | 1–10.5 | |
| Pseudotsuga menziesii | 15 | 11 | 73 | 2.1 | 1–6 | |
| Fall | Betula papyrifera | 18 | 6 | 33 | 6.5 | 1–16 |
| Juniperus horizontalis | 2 | 2 | 100 | 5.3 | 4–6 | |
| Picea glauca (local) | 22 | 20 | 91 | 4.5 | 1–10 | |
| Picea glauca (nursery) | 23 | 14 | 61 | 3.1 | 1–13 | |
| Populus tremuloides | 22 | 9 | 44 | 2.8 | 1–8 | |
| Pseudotsuga menziesii | 25 | 15 | 60 | 2.5 | 1–9 |
2.2. Plant Establishment from Seeds
3. Experimental Section
3.1. Study Area
3.2. Treatments and Experimental Design
3.2.1. Experimental Design
| Plant Sources | |||
|---|---|---|---|
| Transplanting and Seeding Season | Soil Amendment | Transplants | Seed |
| Fall | Control | aspen | aspen |
| Cleanfill | douglas fir | bearberry | |
| Woodshavings | juniper | douglas fir | |
| paper birch | paper birch | ||
| spruce (local) | spruce | ||
| spruce (nursery) | green alder | ||
| LFH* | |||
| Spring | Control | aspen | aspen |
| Cleanfill | douglas fir | bearberry | |
| Woodshavings | juniper | douglas fir | |
| paper birch | paper birch | ||
| spruce (local) | spruce | ||
| spruce (nursery) | green alder | ||
| LFH * | |||
3.2.2. Soil Amendments and Fertilizer
3.2.3. Plant Species and Sources, Transplanting and Seeding
3.3. Vegetation Assessment
3.4. Statistical Analyses
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Martínez-Garza, C.; Howe, H.F. Restoring tropical diversity: Beating the time tax on species loss. J. Appl. Ecol. 2003, 40, 423–429. [Google Scholar] [CrossRef]
- Wickham, J.; Riitters, K.; Wade, T.; Coan, M.; Homer, C. The effect of Appalachian mountaintop mining on interior forest. Landsc. Ecol. 2007, 22, 179–187. [Google Scholar] [CrossRef]
- Mudd, G.M. The environmental sustainability of mining in Australia: Key mega-trends and looming constraints. Resour. Policy 2010, 35, 98–115. [Google Scholar] [CrossRef]
- Costa, S.; Scoble, M. An interdisciplinary approach to integrating sustainability into mining engineering education and research. J. Clean. Prod. 2004, 14, 366–373. [Google Scholar] [CrossRef]
- Mining, Minerals And Sustainable Development Project (MMSD). Breaking new ground, mining, minerals and sustainable development. International Institute for Environment and Development. Earthscan Publications: London, UK, 2002. Available online: http://www.iied.org/mmsd/finalreport/index.html (accessed on 5 November 2012).
- Global Reporting Initiative (GRI). Sustainability reporting guidelines and mining and metals sector supplement. RG version 3.0/MMSS final version. Available online: https://www.globalreporting.org/resourcelibrary/MMSS-Complete.pdf (accessed on 5 November 2012).
- Petchey, O.; Gaston, K. Functional diversity (FD), species richness and community composition. Ecol. Lett. 2002, 5, 402–411. [Google Scholar] [CrossRef]
- Menninger, H.L.; Palmer, M.A. Chapter 5. Restoring Ecological Communities: From Theory to Practice. In Foundations of Restoration Ecology; Falk, D.A., Palmer, M.A., Zedler, J.B., Eds.; Island Press: Washington, DC, USA, 2006; pp. 88–112. [Google Scholar]
- Lamb, T.; Naeth, M.A.; Rothwell, R. Reclamation evaluation of the Rogers Pass Project. Prepared for Parks Canada and Canadian Pacific Railway. 1998; p. 44. [Google Scholar]
- Macyk, T.M. Thirty Years of Reclamation Research in the Alpine and Subalpine Regions near Grande Cache, Alberta. In Proceedings of the 26th Annual British Columbia Mine Reclamation Symposium, Dawson Creek, BC, Canada, 9–13 September 2002; pp. 22–33.
- Naeth, M.A.; Archibald, H.A.; Nemirsky, C.L.; Leskiw, L.A.; Brierley, J.A.; Bock, M.D.; VandenBygaart, A.J.; Chanasyk, D.S. Proposed classification for human modified soils in Canada: Anthroposolic order. Can. J. Soil. Sci. 2012, 92, 7–18. [Google Scholar]
- Čermák, P. Forest reclamation of dumpsites of coal combustion by-products (CCB). J. For. Sci. 2008, 54, 273–280. [Google Scholar]
- Cohen-Fernández, A.C.; Naeth, M.A. Anthroposol development from limestone quarry substrates. Can. J. Soil. Sci. 2013. submitted for publication. [Google Scholar]
- Cohen-Fernández, A.C.; Naeth, M.A. Erosion control blankets, organic amendments and site variability influenced the initial plant community at a limestone quarry in the Canadian rocky mountains. Biogeosciences Discuss 2013, 10, 3009–3037. [Google Scholar] [CrossRef]
- Solondz, D.M. Improving Rehabilitation Options through Research. In Proceedings of the Mining and Environment V Conference, Sudbury, Canada, 25–30 June 2011.
- Shulz, F.; Wiegleb, G. Development options of natural habitats in a post-mining landscape. Land Degrad. Dev. 2000, 11, 99–110. [Google Scholar] [CrossRef]
- Wunderle, J.M., Jr. The role of animal seed dispersal in accelerating native forest regeneration on degraded tropical lands. For. Ecol. Manage 1997, 99, 223–235. [Google Scholar] [CrossRef]
- Rodrigues, R.R.; Martins, S.V.; de Barros, L.C. Tropical rain forest regeneration in an area degraded by mining in Mato Grosso State, Brazil. For. Ecol. Manage 2004, 190, 323–333. [Google Scholar] [CrossRef]
- Gerling, H.S.; Willoughby, M.G.; Schoepf, A.; Tannas, K.E.; Tannas, C.A. A Guide to Using Native Plants on Disturbed Lands; Alberta Agriculture, Food and Rural Development: Edmonton, AB, Canada, 1996. [Google Scholar]
- Callaway, R.M.; Davis, F.W. Recruitment of Quercus. agrifolia in central California: The importance of shrub-dominated patches. J. Veg. Sci. 1998, 9, 647–656. [Google Scholar]
- Rousset, O.; Lepart, J. Shrub facilitation of Quercus. humilis regeneration in succession on calcareous grasslands. J. Veg. Sci. 1999, 10, 493–502. [Google Scholar] [CrossRef]
- Del Moral, R.; Wood, D.M. Early primary succession on the volcano Mount St. Helens. J. Veg. Sci. 1993, 4, 223–234. [Google Scholar] [CrossRef]
- Gomez-Aparicio, L. The role of plant interactions in the restoration of degraded ecosystems: A meta-analysis across life-forms and ecosystems. J. Ecol. 2009, 97, 1202–1214. [Google Scholar] [CrossRef]
- Campbell, R.K.; Sorensen, F.C. Effect of test environment on expression of clines and on delimitation of seed zones in douglas-fir. Theoret. Appl. Genet. 1978, 51, 233–246. [Google Scholar] [CrossRef]
- Smith, B.M.; Diaz, A.; Winder, L.; Daniels, R. The effect of provenance on the establishment and performance of Lotus corniculatus L. in a re-creation environment. Biol. Conserv. 2005, 125, 37–46. [Google Scholar]
- Holmes, P. Shrubland restoration following woody alien invasion and mining: Effects of topsoil depth, seed source, and fertilizer addition. Restor. Ecol. 2001, 9, 71–84. [Google Scholar] [CrossRef]
- Mackenzie, D.D.; Naeth, M.A. The role of the forest soil propagule bank in assisted natural recovery after oil sands mining. Restor. Ecol. 2009, 18, 418–427. [Google Scholar] [CrossRef]
- Kramer, P.J.; Kozlowski, T.T. Physiology of Woody Plants; Academic Press: New York, NY, USA, 1979. [Google Scholar]
- Harrington, G.N. Effects of soil moisture on shrub seedling survival in semi-arid grassland. Ecology 1991, 72, 1138–1149. [Google Scholar] [CrossRef]
- Environment Canada. Banff station. Canadian climate normals 1971–2000. Available online: http://www.climate.weatheroffice.gc.ca (accessed on 18 June 2011).
- Young, B.J.; Harrington, J.T.; Loveall, M.W.; Wagner, A.; Sanders, J.; Buchanan, B.A. Short and Long Term Transplant Performance on Mine Rock Material, Questa mine, New Mexico. In National Meeting of the American Society of Mining and Reclamation, Billings, MT, USA, 30 May–5 June, 2009; Barnhisel, R.I., Ed.; American Society of Mining and Reclamation: Lexington, KY, USA, 2009; pp. 1698–1710. [Google Scholar]
- Johnson, P.S.; Rogers, R. A method for estimating the contribution of planted hardwoods to future stocking. For. Sci. 1985, 31, 883–891. [Google Scholar]
- Haase, D.L.; Rose, R. Soil moisture stress induces transplant shock in stored and unstored 2 + 0 douglas-fir seedlings of varying root volumes. For. Sci. 1993, 39, 275–294. [Google Scholar]
- Vyse, A. Growth of young spruce plantations in interior British Columbia. For. Chron. 1981, 57, 174–180. [Google Scholar]
- Harrington, J.T.; Loveall, M.W. Evaluating Forest Productivity on Reclaimed Mine Land in Western United States. In Proceedings of the 7th International Conference on acid rock drainage, 26–30 March 2006; American Society of Mining and Reclamation: Lexington, Kentucky, USA, 2006; pp. 721–737. [Google Scholar]
- McCann, L.J. Ecology of the mountain sheep. Am. Midl. Nat. 1956, 56, 297–324. [Google Scholar] [CrossRef]
- Shannon, N.H.; Hudson, R.J.; Brink, V.C.; Kitts, W.D. Determinants of spatial distribution of rocky mountain bighorn sheep. J. Wildlife Manag. 1975, 39, 387–401. [Google Scholar] [CrossRef]
- Galvez, D.A.; Landhäusser, S.M.; Tyree, M.T. Low root reserve accumulation during drought may lead to winter mortality in Poplar seedlings. New Phytol. 2013, 198, 139–148. [Google Scholar] [CrossRef]
- Cole, E.C.; Newton, M.; Youngblood, A. Regenerating white spruce, paper birch, and willow in south-central Alaska. Can. J. For. Res. 1999, 29, 993–1001. [Google Scholar] [CrossRef]
- Cattelino, P.J.; Noble, I.R.; Slatyer, R.O.; Kessell, S.R. Predicting the multiple pathways of plant succession. Environ. Manag. 1979, 3, 41–50. [Google Scholar] [CrossRef]
- Shannon, N.H.; Hudson, R.H.; Brink, V.C.; Kitts, W.D. Determinants of spatial distribution of Rocky Mountain bighorn sheep. J. Wildl. Manage 1975, 39, 387–401. [Google Scholar] [CrossRef]
- Steven, D.D. Experiments on mechanisms of tree establishment in old-field succession: Seedling emergence. Ecology 1991, 72, 1066–1075. [Google Scholar] [CrossRef]
- DeLong, H.B.; Lieffers, V.J.; Blenis, P.V. Microsite effects on first-year establishment and overwinter survival of white spruce in aspen-dominated boreal mixedwoods. Can. J. For. Res. 1997, 27, 1452–1457. [Google Scholar] [CrossRef]
- Eis, S. Development of white spruce and alpine fir seedlings on cut-over areas in central interior of British Columbia. For. Chron. 1965, 41, 419–431. [Google Scholar]
- Moss, E.H. Longevity of seed and establishment of seedlings in species of populus. Bot. Gaz. 1938, 99, 529–542. [Google Scholar]
- Densmore, R.V.; Page, J.C. Paper birch regeneration on scarified logged areas in southcentral Alaska. N. J. Appl. For. 1992, 9, 63–66. [Google Scholar]
- Alberta Parks and Protected Areas Division. In Bow valley protected areas management plan, 0–7785–2221–0; Alberta Community Development, Parks and Protected Areas: Edmonton, Canada, 2002.
- Agresti, A. Categorical Data Analysis, 2nd ed; Wiley-Interscience: New York, NY, USA, 2002. [Google Scholar]
- Jones, O.R.; Crawley, M.J.; Pilkington, J.G.; Pemberton, J.M. Predictors of early survival in Soay sheep: cohort-, maternal- and individual-level variation. Proc. Royal. Soc. Biol. Sci. 2005, 272, 2619–2625. [Google Scholar] [CrossRef]
- Agresti, A. An Introduction to Categorical Data Analysis, 2nd ed; Wiley-Interscience: Hoboken, NJ, USA, 2007. [Google Scholar]
- R Development Core Team, R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2011.
- Systat Software. In SigmaPlot for Windows version 12; Systat Software Inc.: Chicago, IL, USA, 2011.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cohen-Fernandez, A.C.; Naeth, M.A. Increasing Woody Species Diversity for Sustainable Limestone Quarry Reclamation in Canada. Sustainability 2013, 5, 1340-1355. https://doi.org/10.3390/su5031340
Cohen-Fernandez AC, Naeth MA. Increasing Woody Species Diversity for Sustainable Limestone Quarry Reclamation in Canada. Sustainability. 2013; 5(3):1340-1355. https://doi.org/10.3390/su5031340
Chicago/Turabian StyleCohen-Fernandez, Anayansi C., and M. Anne Naeth. 2013. "Increasing Woody Species Diversity for Sustainable Limestone Quarry Reclamation in Canada" Sustainability 5, no. 3: 1340-1355. https://doi.org/10.3390/su5031340