Assessment of the Role of Local Strawberry Rhizosphere—Associated Streptomycetes on the Bacterially—Induced Growth and Botrytis cinerea Infection Resistance of the Fruit



Abstract
:1. Introduction
2. Experimental Section
2.1. Isolation of the Pathogen
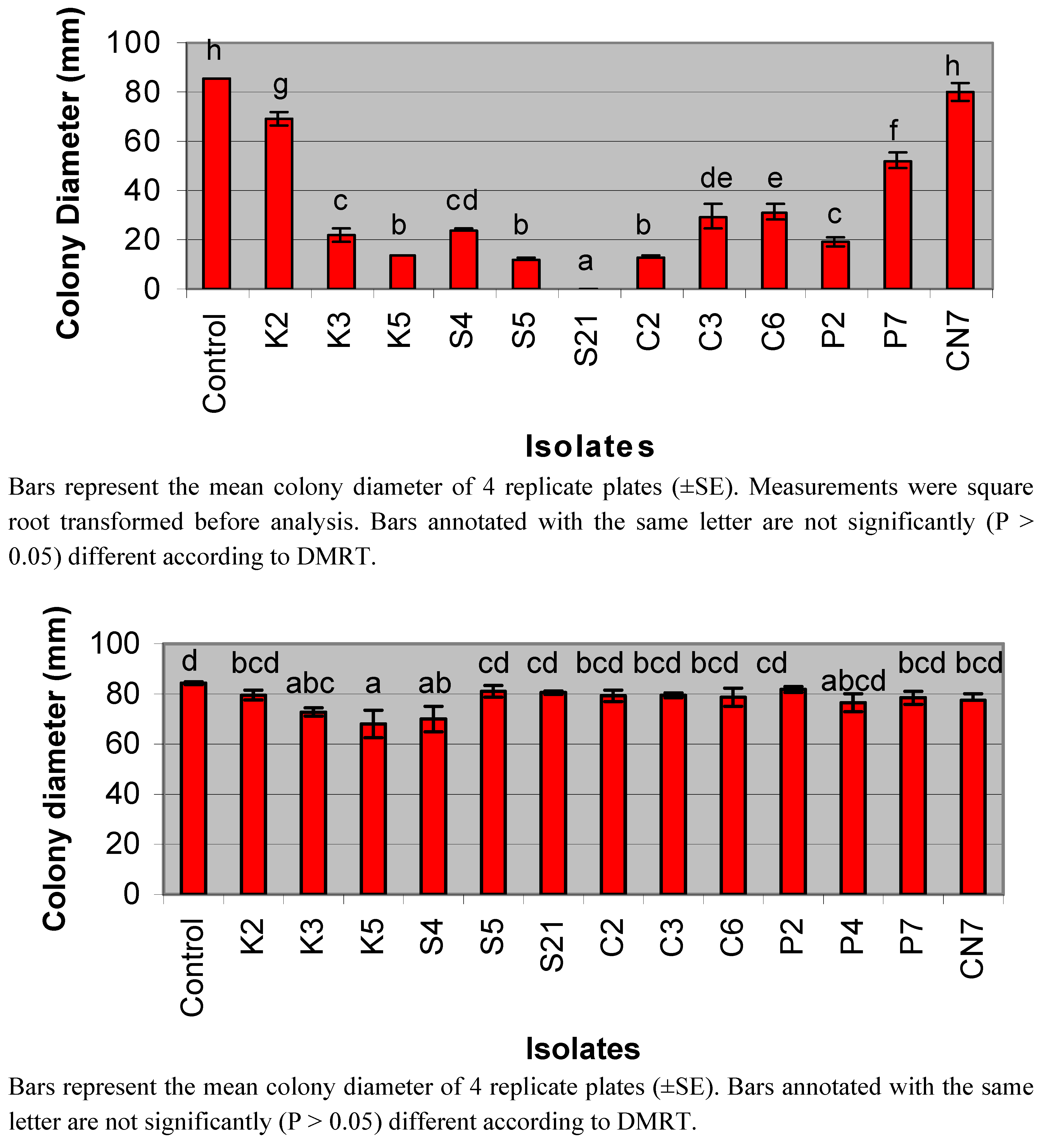
2.2. Isolation and Antifungal Screening of Streptomycetes against the Pathogen

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Total number of isolates | Isolate codes |
|---|---|---|
| Selva | 8 | S1≠, S2, S5≠, S6, S7, S8, S9, S10 |
| Kabala | 9 | K1≠, K2≠, K3≠, K4≠, K5≠, K6≠, K7, K8≠ |
| Camarosa | 16 | C1, C2≠, C3≠, C4≠, C6≠, C7≠, C8, C9, C10≠ CN1≠, CN2≠, CN3≠, CN4≠, CN5≠, CN6≠, CN7≠ |
| Parker | 7 | P1, P2≠, P3, P4≠, P5, P6, P7≠ |
| USC isolates* | S4≠, S21≠ |
2.3. Antifungal Compound Extractions and Purifications
2.4. Pot Trial
| Pot treatment | Description |
|---|---|
| Treatment-1 | Control |
| Treatment-2 | Disease control |
| Treatment-3 | C3-LP-EtOAc extract spray solution on aerial plant surfaces |
| Treatment-4 | Soil inoculated with non-inhibitory streptomycete isolates C1, C8, P6, S2 |
| Treatment-5 | Soil inoculated with inhibitory streptomycete isolates K3, K5, S4, S21 |
| Treatment-6 | Rye grass seed inoculum control (Soil inoculated only with sterilised rye grass seeds) |
2.5. Pathogen Inoculations
2.6. Application of Antifungal Extract
2.7. Harvesting of Strawberry Fruit
3. Results and Discussion
3.1. Detection of Antifungal Activity from Streptomycete Isolates
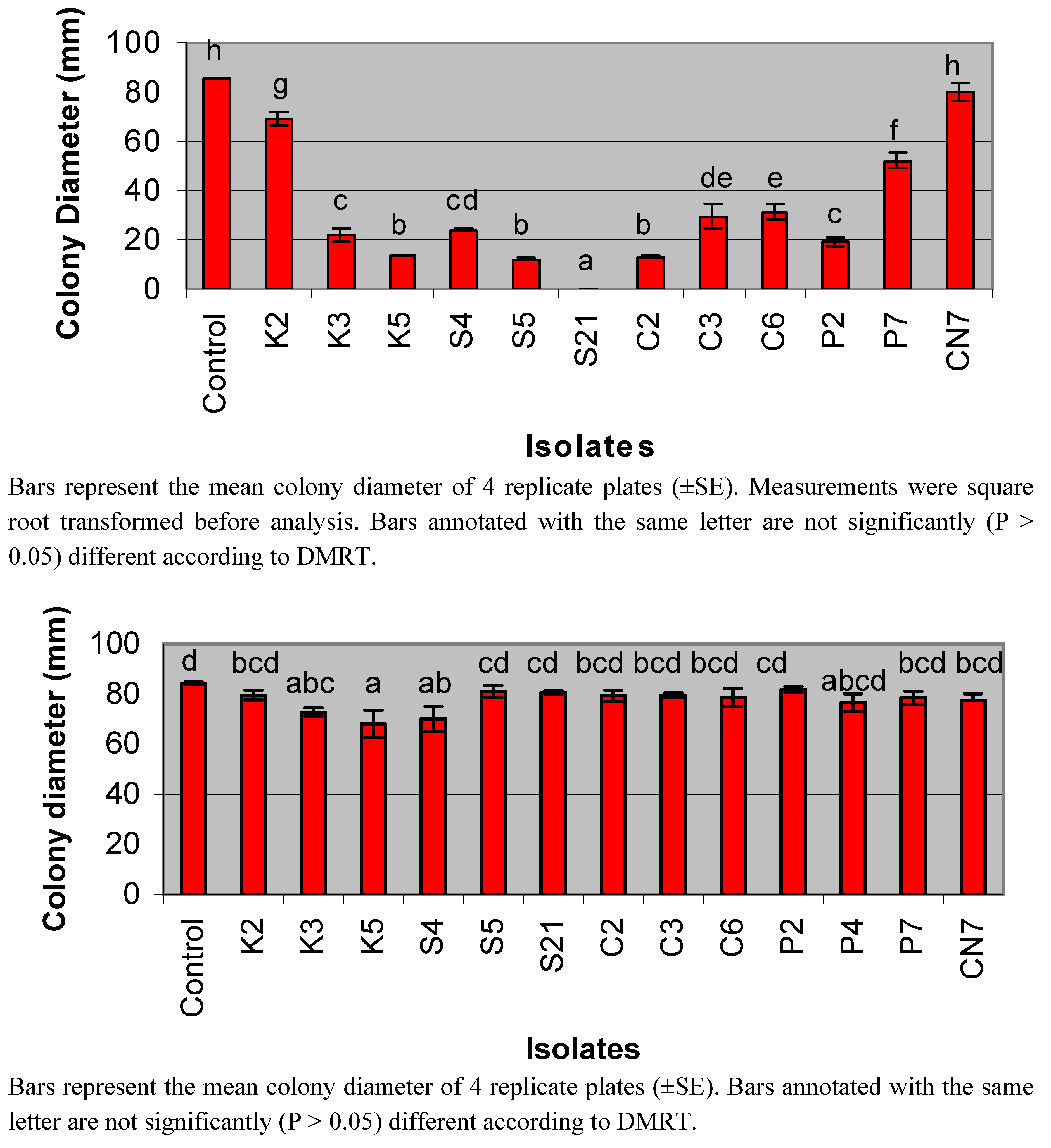
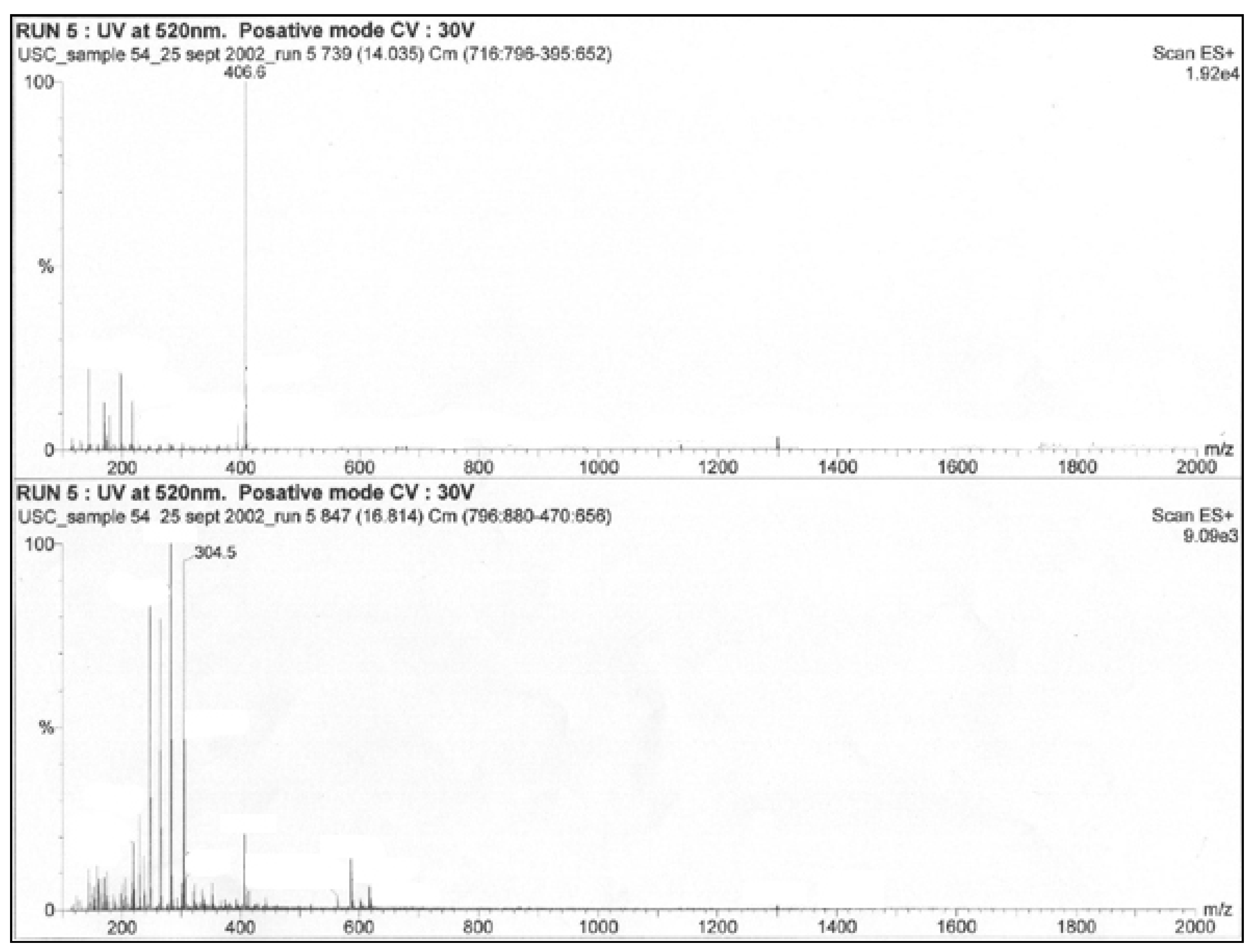
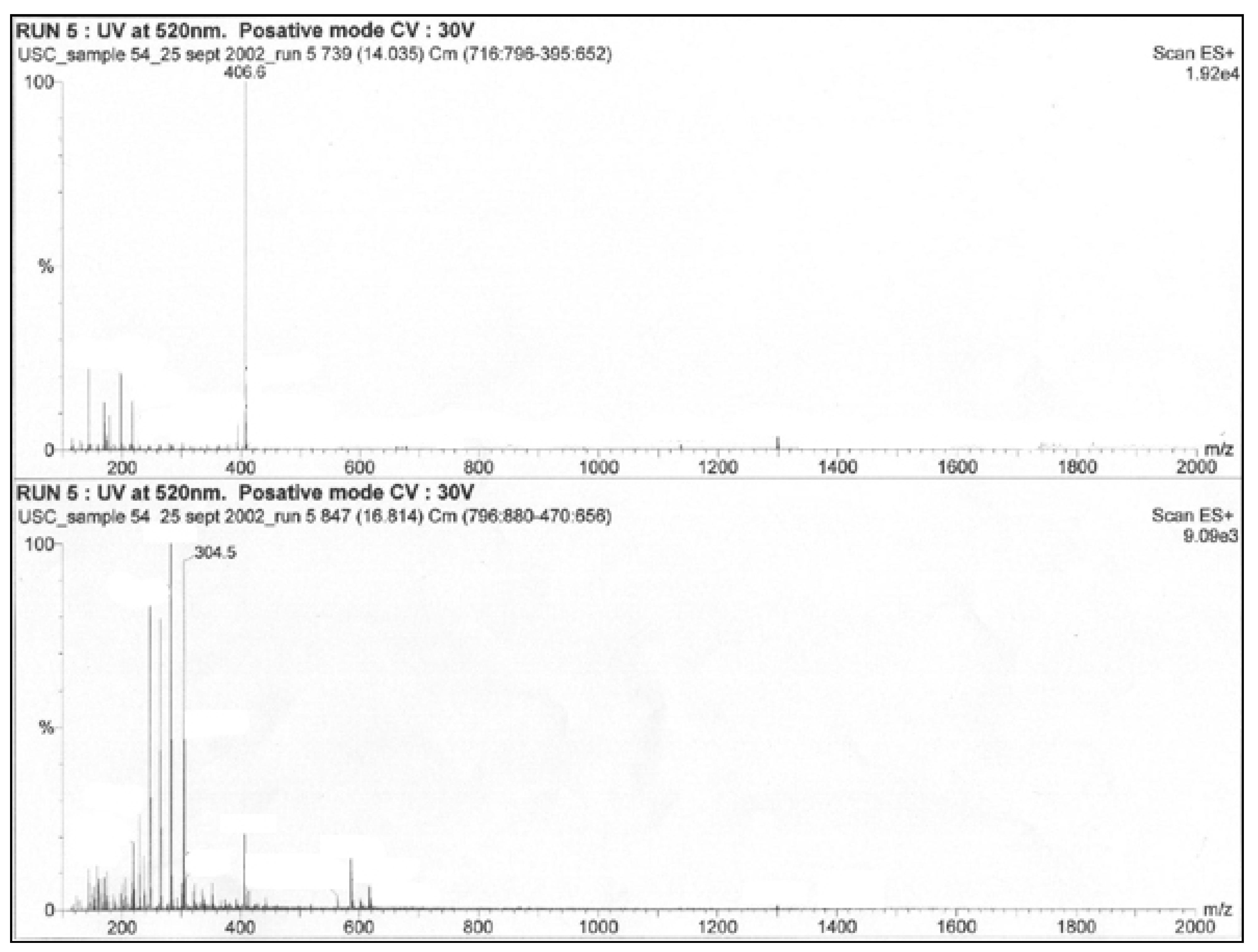
3.2. Extract Assays
| Extract | Inhibition Zone (mm) |
| S21-MeOH | 8.3 |
| S21-EtOAc | 11.0 |
| S4-MeOH | 9.0 |
| S4-EtOAc | 13.0 |
| C3-EtOAc | 7.0 |
| C3-MeOH | 0.0 |
| C3-LP-EtOAc | 6.6 |
| K5-MeOH | 0.0 |
| K5-EtOAc | 1.6 |
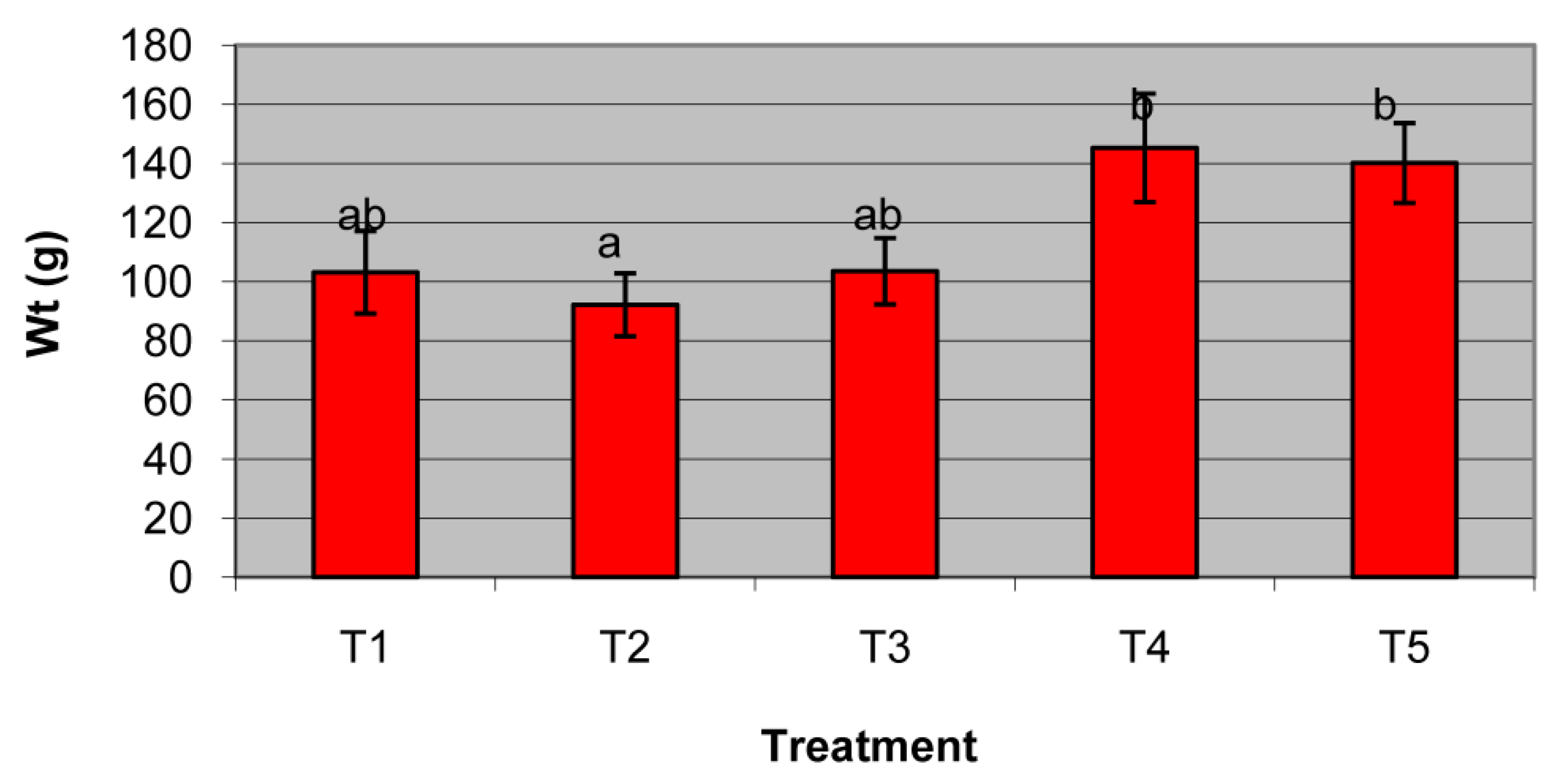
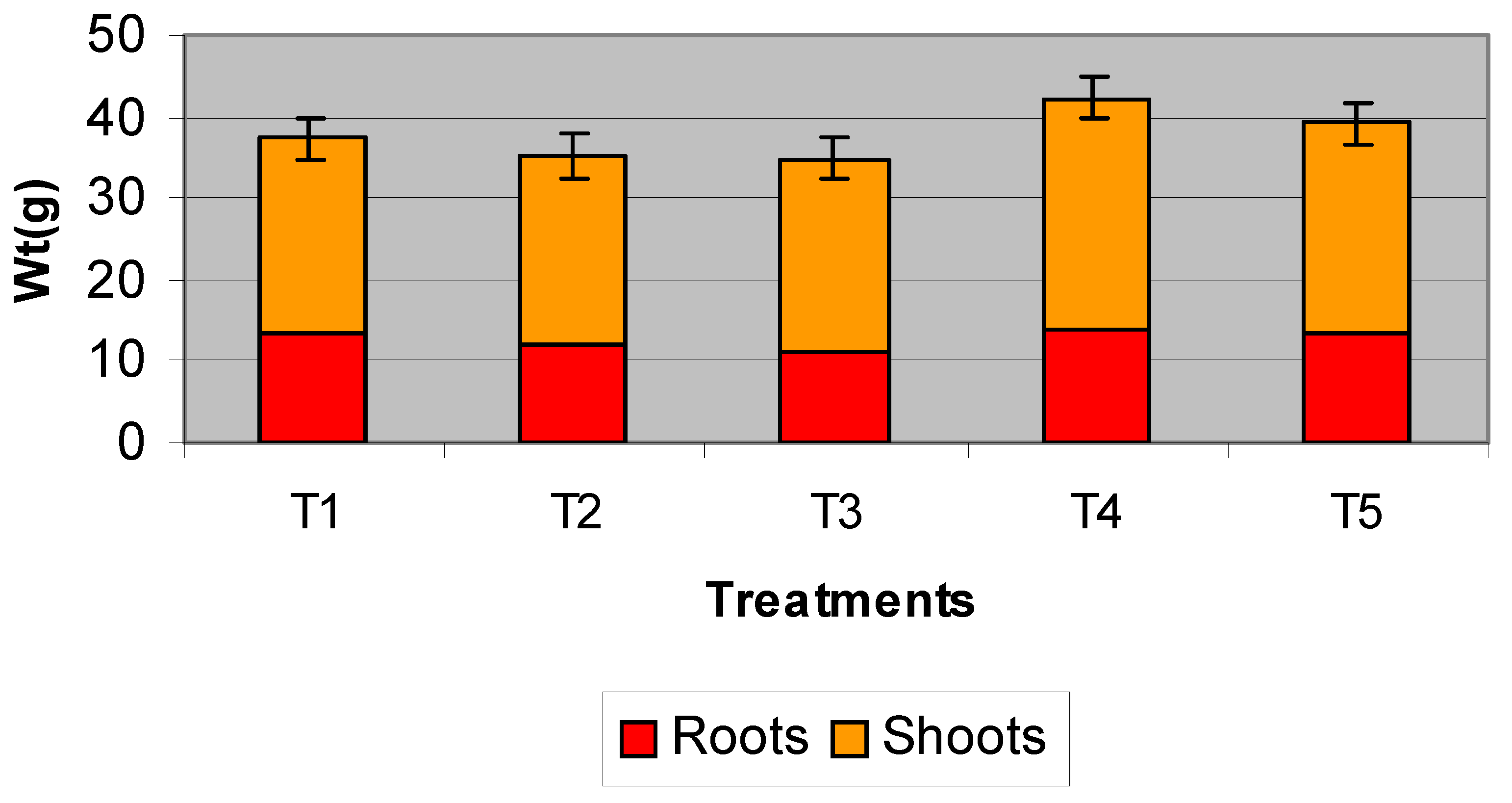
3.3. Strawberry Pot Trials
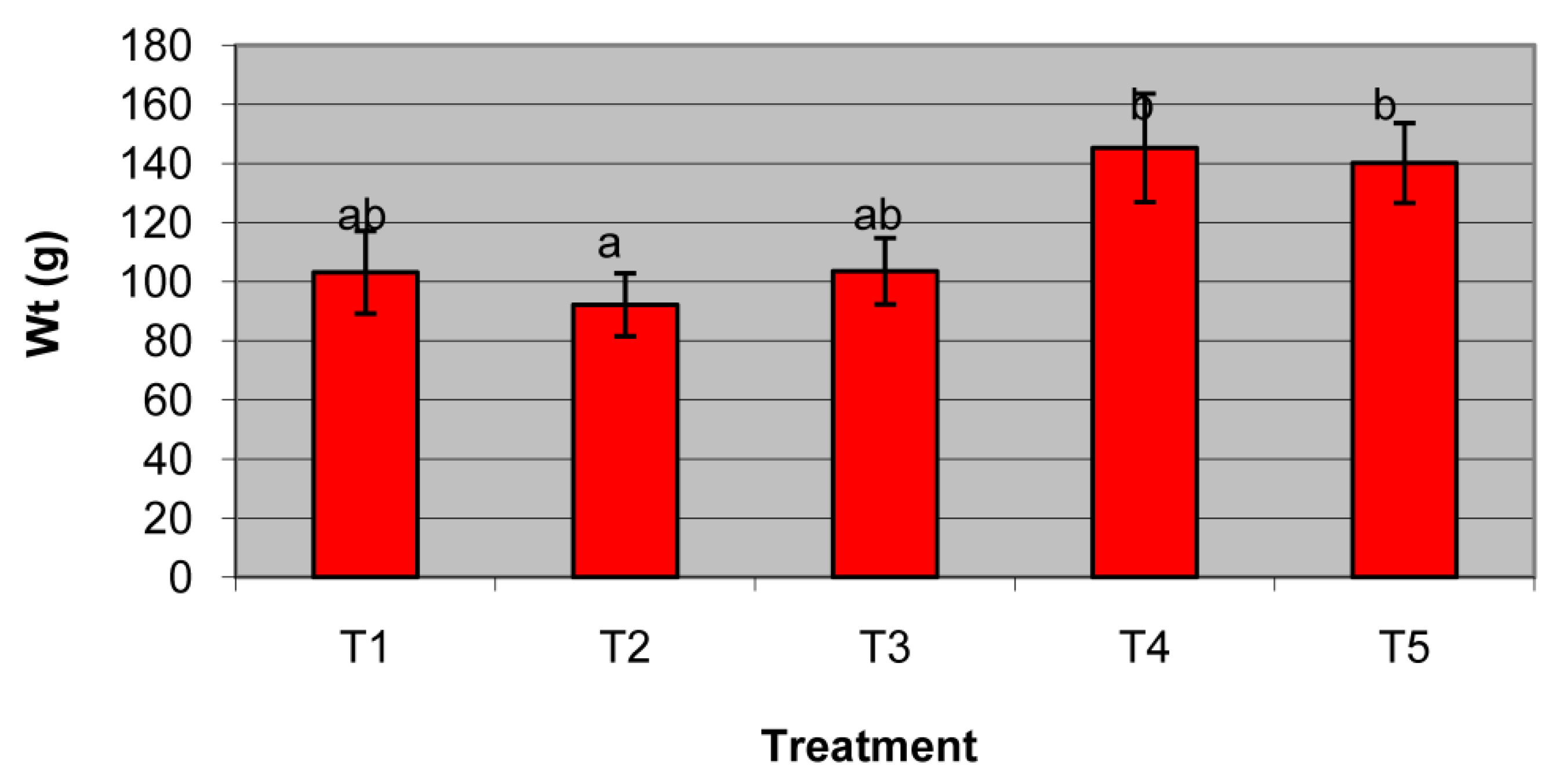
| Pot Treatment | Disease Index | Incidence (%) |
|---|---|---|
| Treatment-1 | 1 | 91% |
| 2 | 2% | |
| 3 | 0% | |
| 4 | 2% | |
| 5 | 5% | |
| Treatment-2 | 1 | 75% |
| 2 | 9% | |
| 3 | 2% | |
| 4 | 7% | |
| 5 | 7% | |
| Treatment-3 | 1 | 70% |
| 2 | 6% | |
| 3 | 8% | |
| 4 | 5% | |
| 5 | 11% | |
| Treatment-4 | 1 | 62% |
| 2 | 9% | |
| 3 | 12% | |
| 4 | 8% | |
| 5 | 9% | |
| Treatment-5 | 1 | 70% |
| 2 | 8% | |
| 3 | 6% | |
| 4 | 6% | |
| 5 | 10% |
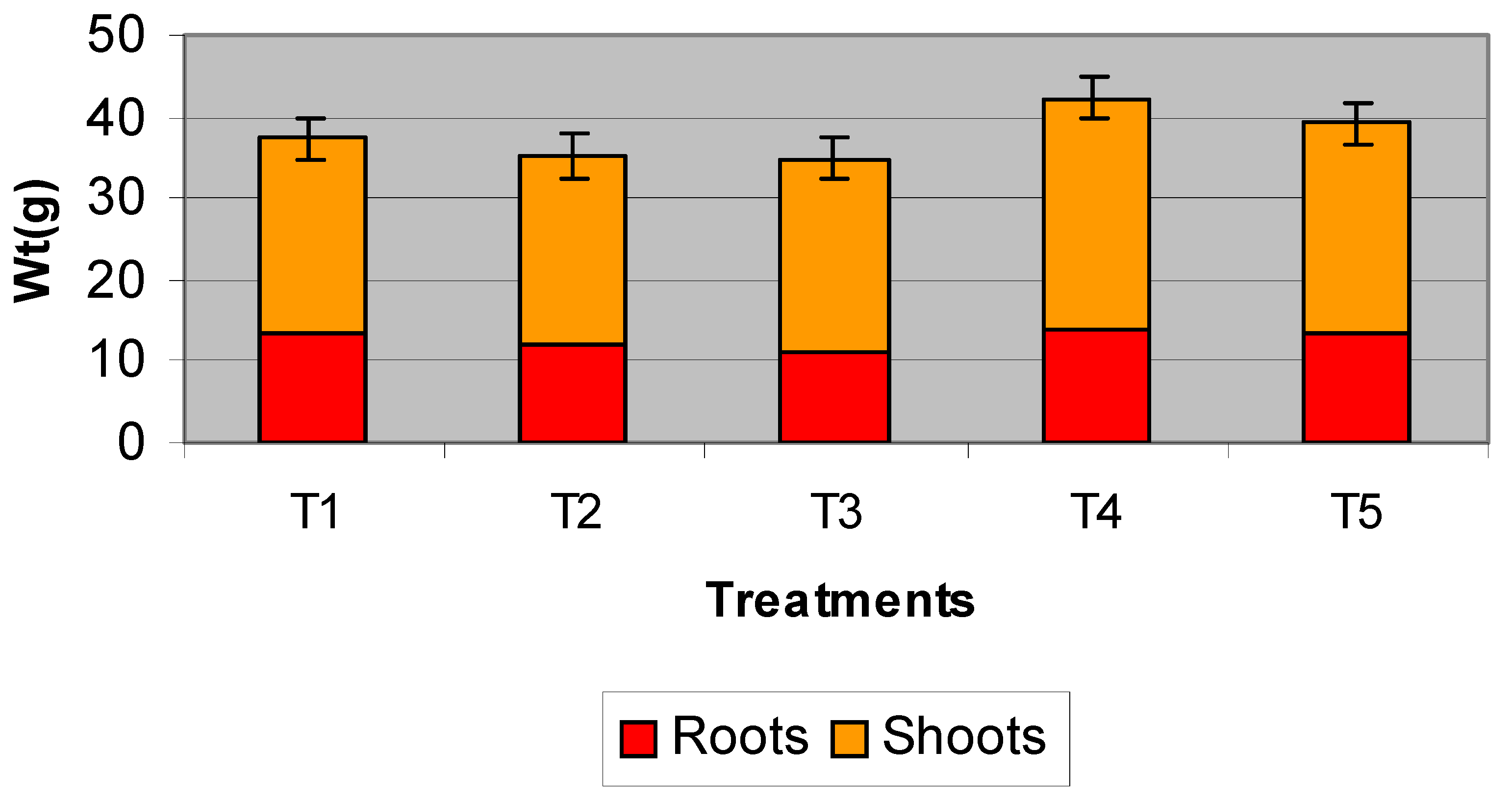
4. Conclusions
Acknowledgements
References
- Queensland Strawberry Industry. Available online: http://www.qldstrawberries.com.au/links.htm (accessed on 1 December 2010).
- Strawberry R&D Update; Department of Primary Industries and Fisheries: Nambour, Australia, 2007. Available online: http://www.qldstrawberries.com.au/pdf/strawberry_update_2007.pdf (accessed on 1 December 2010).
- Helbig, J. Biological control of Botrytis cinerea Pers. ex Fr. in strawberry by Paenibacillus polymyxa (isolate 18191). J. Phytopathol. 2001, 149, 265–273. [Google Scholar] [CrossRef]
- Kalamarakis, A.E.; Petsikos-Panagiotarou, N.; Mavroidis, B.; Ziogas, B.N. Activity of fluazinam against strains of Botrytis cinerea resistant to benzimidazoles and/or dicarboximides and to a benzimidazole-phenylcarbamate mixture. J. Phytopathol. 2000, 148, 449–455. [Google Scholar] [CrossRef]
- Dianez, F.; Santos, M.; Blanco, P.; Tello, J.C. Fungicide resistance in Botrytis cinerea isolates from strawberry crops in Huelva (southwestern Spain). Phytoparasitica 2002, 30, 529–534. [Google Scholar] [CrossRef]
- Freeman, S.; Minz, D.; Kolesnik, I.; Barbul, O.; Zveibil, A.; Mayon, M.; Nitzani, Y.; Kirshner, B.; Rav-David, D.; Bilu, A.; Dag, A.; Shafir, S.; Elad, Y. Trichoderma biocontrol of Colletotrichum acutatum and Botrytis cinerea and survival in strawberry. Eur. J. Plant Pathol. 2004, 110, 361–370. [Google Scholar] [CrossRef]
- Vagelas, I.; Papachatzis, A.; Kalorizou, H.; Wogiatzi, E. Biological control of botrytis fruit rot (gray mold) on strawberry and red pepper fruits by olive oil mill wastewater. Biotechnol. Biotechnol. Eq. 2009, 23, 1489–1491. [Google Scholar] [CrossRef]
- Rosslenbroich, H.-J.; Stuebler, D. Botrytis cinerea—History of chemical control and novel fungicides for its management. Crop Prot. 2000, 19, 557–561. [Google Scholar]
- Szandala, E.S.; Backhouse, D. Suppression of sporulation of Botrytis cinerea by antagonists applied after infection. Australas. Plant Pathol. 2000, 30, 165–170. [Google Scholar] [CrossRef]
- Elad, Y. Mechanisms involved in biological control of Botrytis cinerea incited diseases. Eur. J. Plant Pathol. 1996, 102, 719–732. [Google Scholar] [CrossRef]
- Mommaerts, V.; Platteau, G.; Boulet, J.; Sterk, G.; Smagghe, G. Trichoderma-based biological control agents are compatible with the pollinator Bombus terrestris: A laboratory study. Biol. Control. 2008, 46, 463–466. [Google Scholar] [CrossRef]
- Atlas, R.M. Handbook of Microbiological Media, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2006. [Google Scholar]
- Kuster, E.; Williams, S.T. Selection of media for isolation of streptomycetes. Nature 1964, 202, 928–929. [Google Scholar] [CrossRef] [PubMed]
- Williams, S.T.; Wellington, E.M.H. Actinomycetes. In Methods of Soil Analysis, Part 2. Chemical and Microbiological Properties, 2nd ed.; Miller, R.H., Keeney, O.R., Eds.; American Society of Agronomy: Madison, DC, USA, 1982; pp. 969–987. [Google Scholar]
- Cross, T. Growth and examination of actinomycetes. In Bergey’s Manual of Systematic Bacteriology; Williams, S.T., Sharpe, M.E., Holt, J.G., Eds.; Williams and Wilkins: Baltimore, MD, USA, 1989; Volume 4, pp. 2340–2344. [Google Scholar]
- Shirling, E.B.; Gottlieb, D. Methods for characterization of Streptomyces species. Int. J. Syst. Bacteriol. 1966, 16, 313–340. [Google Scholar] [CrossRef]
- Wellington, E.M.H.; Williams, S.T. Preservation of actinomycetes inoculum in frozen glycerol. Microbios Lett. 1978, 6, 151–157. [Google Scholar]
- Dhingra, O.D.; Sinclair, J.B. Basic Plant Pathology Methods, 2nd ed.; CRC Press: Boca Raton, FL, USA, 1995. [Google Scholar]
- Gibbs, J.N. A study of the epiphytic growth habit of Fomes annosus. Ann. Bot. 1967, 31, 755–774. [Google Scholar]
- Dennis, C.; Webster, J. Antagonistic properties of species-groups of Trichoderma II production of volatile antibiotics. T. Brit. Mycol. Soc. 1971, 57, 41–48. [Google Scholar] [CrossRef]
- Shimizu, M.; Nakagawa, Y.; Sato, Y.; Furumai, T.; Igarashi, Y.; Onaka, H.; Yoshida, R.; Kunoh, H. Studies on endophytic Actinomycetes (I) Streptomyces sp. isolated from rhododendron and its antifungal activity. J. Gen. Plant. Pathol. 2000, 66, 360–366. [Google Scholar] [CrossRef]
- Edwards, S.G.; Seddon, B. Mode of antagonism of Brevibacillus brevis against Botrytis cinerea in vitro. J. Appl. Microbiol. 2001, 91, 652–659. [Google Scholar] [CrossRef] [PubMed]
- Kurtboke, D.I.; Cardillo, R.; Nasini, G.; Petrolini, B.; Sardi, P.; Locci, R. Anti-candida metabolites produced by streptomycetes isolated from the atmosphere of agricultural environments. In Actinomycetes; Lechevalier, H., Ensign, J.C., Kalakoutskii, L.V., Locci, R., Eds.; Waksman Institute of Microbiology—Rutgers The State University of New Jersey: New Brunswick, NJ, USA, 1986; pp. 223–231. [Google Scholar]
- Kurtboke, D.I.; Shankar, M.; Rowland, C.Y.; Sivasithamparam, K. Responses of a sterile red fungus to soil types, wheat varieties and the presence of certain isolates of Streptomyces. Plant Soil 1993, 157, 35–40. [Google Scholar]
- Agricultural Field Experiments; Petersen, R.G. (Ed.) Marcel Dekker, Inc.: New York, NY, USA, 1994.
- Swadling, I.R.; Jeffries, P. Antagonistic properties of two bacterial biocontrol agents of grey mould diseases. Biocontrol. Sci. Tech. 1998, 8, 439–448. [Google Scholar] [CrossRef]
- Archbold, D.D.; Hamilton-Kemp, T.R.; Barth, M.M.; Langlois, B.E. Identifying natural volatile compounds that control gray mould (Botrytis cinerea) during postharvest storage of strawberry, blackberry, and grape. J. Agr. Food. Chem. 1997, 45, 4032–4037. [Google Scholar] [CrossRef]
- Srihuttagum, M.; Sivasithamparam, K. The influence of fertilizers on root rot of field peas caused by Fusarium oxysporum, Pythium vexans and Rhizoctonia solani inoculated singly or in combination. Plant Soil 1991, 132, 21–27. [Google Scholar] [CrossRef]
- Synder, L.R. Classification of the solvent properties of common liquid. J. Chromatogr. 1974, 92, 223–230. [Google Scholar] [CrossRef]
- Sutton, J.C. Evaluation of micro-organisms for biocontrol: Botrytis cinerea and strawberry. In Advances in Plant Pathology; Andrews, J.H., Tommerup, I.C., Eds.; Academic Press: London, UK, 1995; Volume 11, pp. 173–190. [Google Scholar]
- Trejo-Estrada, S.R.; Rivas Sepulveda, I.; Crawford, D.L. In vitro and in vivo Antagonism of Streptomyces violaceusniger YCED9 against fungal pathogens of turfgrass. World J. Microb. Biot. 1998, 14, 865–872. [Google Scholar] [CrossRef]
- Tokala, R.K.; Strap, J.L.; Jung, C.M.; Crawford, D.L.; Hamby Salove, M.; Deobald, L.A.; Bailey, J.F.; Morra, M.J. Novel plant-microbe rhizosphere interaction involving Streptomyces lydicus WYEC108 and the pea plant (Pisum sativum). Appl. Environ. Microbiol. 2002, 68, 2161–2171. [Google Scholar] [CrossRef] [PubMed]
- Whipps, J. Microbial interactions and biocontrol in the rhizosphere. J. Exp. Bot. 2001, 52, 487–511. [Google Scholar] [CrossRef] [PubMed]
- Berg, G. Plant-microbe interactions promoting plant growth and health: Perspectives for controlled use of microorganisms in agriculture. Appl. Microbiol. Biotechnol. 2009, 84, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Schrey, S.D.; Tarkka, M.T. Friends and foes: Streptomycetes as modulators of plant disease and symbiosis. Antonie Leeuwenhoek 2008, 94, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Compant, S.; Duffy, B.; Nowak, J.; Clement, C.; Barka, E.A. Use of plant growth-promoting bacteria for biocontrol of plant diseases: Principles, mechanisms of action, and future prospects. Appl. Environ. Microbiol. 2005, 71, 4951–4959. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Cornejo, H.A.; Macias-Rodriguez, L.; Cortes-Penagos, C.; Lopez-Bucio, J. Trichoderma virens, a plant beneficial fungus, enhances biomass production and promotes lateral root growth through an auxin-dependent mechanism in Arabidopsis. Plant Physiol. 2009, 149, 1579–1592. [Google Scholar] [CrossRef] [PubMed]
- Morrissey, J.P.; Dow, J.M.; Mark, L.; O’Gara, F. Are microbes at the root of a solution to world food production? EMBO Rep. 2004, 5, 922–926. [Google Scholar] [CrossRef] [PubMed]
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Eccleston, K.L.; Brooks, P.R.; Kurtböke, D.İ. Assessment of the Role of Local Strawberry Rhizosphere—Associated Streptomycetes on the Bacterially—Induced Growth and Botrytis cinerea Infection Resistance of the Fruit. Sustainability 2010, 2, 3831-3845. https://doi.org/10.3390/su2123831
Eccleston KL, Brooks PR, Kurtböke Dİ. Assessment of the Role of Local Strawberry Rhizosphere—Associated Streptomycetes on the Bacterially—Induced Growth and Botrytis cinerea Infection Resistance of the Fruit. Sustainability. 2010; 2(12):3831-3845. https://doi.org/10.3390/su2123831
Chicago/Turabian StyleEccleston, Karen L., Peter R. Brooks, and D. İpek Kurtböke. 2010. "Assessment of the Role of Local Strawberry Rhizosphere—Associated Streptomycetes on the Bacterially—Induced Growth and Botrytis cinerea Infection Resistance of the Fruit" Sustainability 2, no. 12: 3831-3845. https://doi.org/10.3390/su2123831
APA StyleEccleston, K. L., Brooks, P. R., & Kurtböke, D. İ. (2010). Assessment of the Role of Local Strawberry Rhizosphere—Associated Streptomycetes on the Bacterially—Induced Growth and Botrytis cinerea Infection Resistance of the Fruit. Sustainability, 2(12), 3831-3845. https://doi.org/10.3390/su2123831