
Blue Carbon: Comparison of Chronosequences from Avicennia marina Plantation and Proteresia coarctata Dominated Mudflat, at the World’s Largest Mangrove Wetland




Abstract
:1. Introduction
2. Material and Method
2.1. Study Site
2.2. Community Participation in the Restoration Activity
2.3. Plantation, Monitoring and Assessment Strategy
2.4. Soil Sampling and Analyses
2.5. Assessing Health of Mangrove Ecosystem
2.6. Statistical Tests
3. Results and Discussion
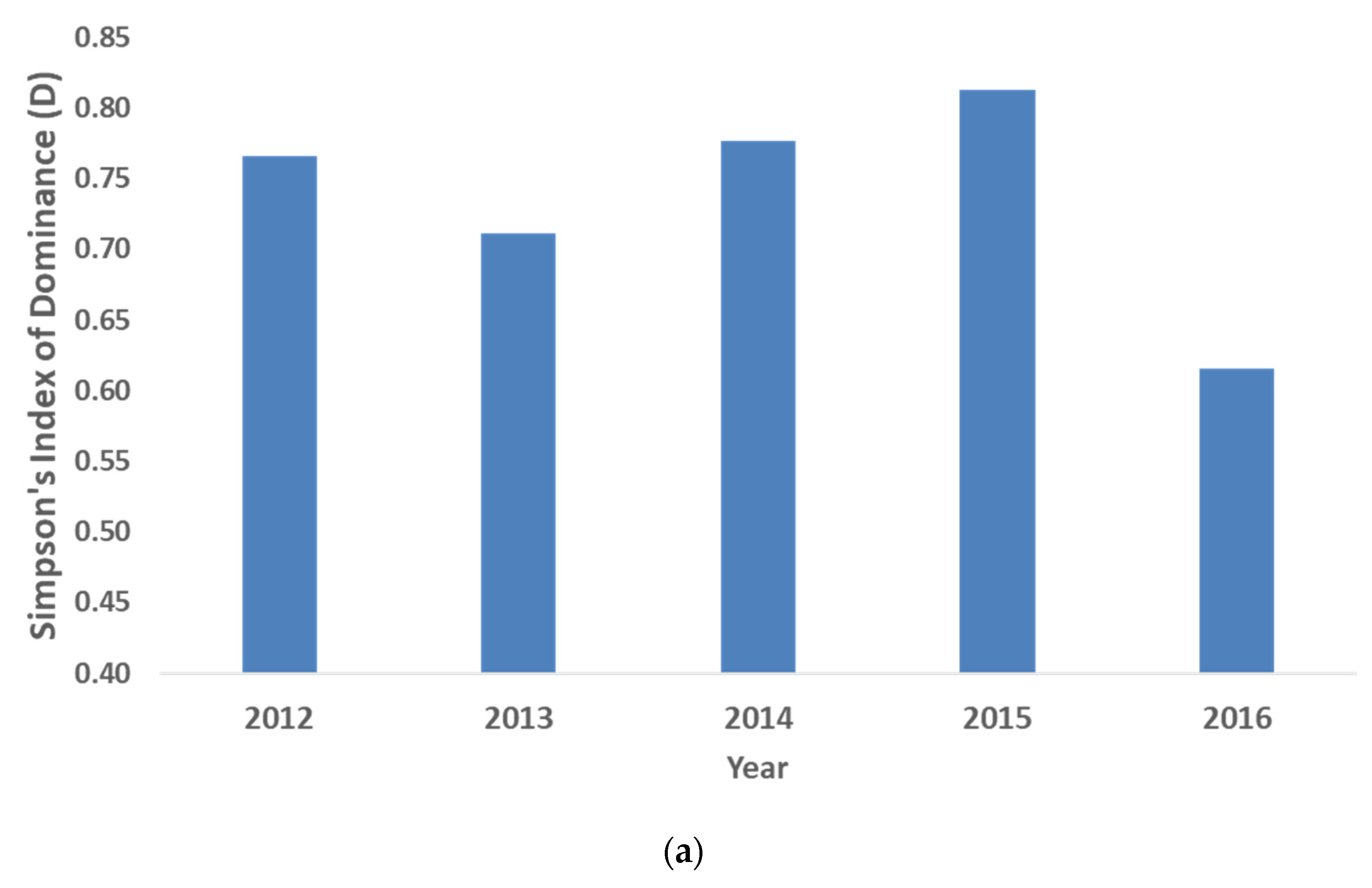
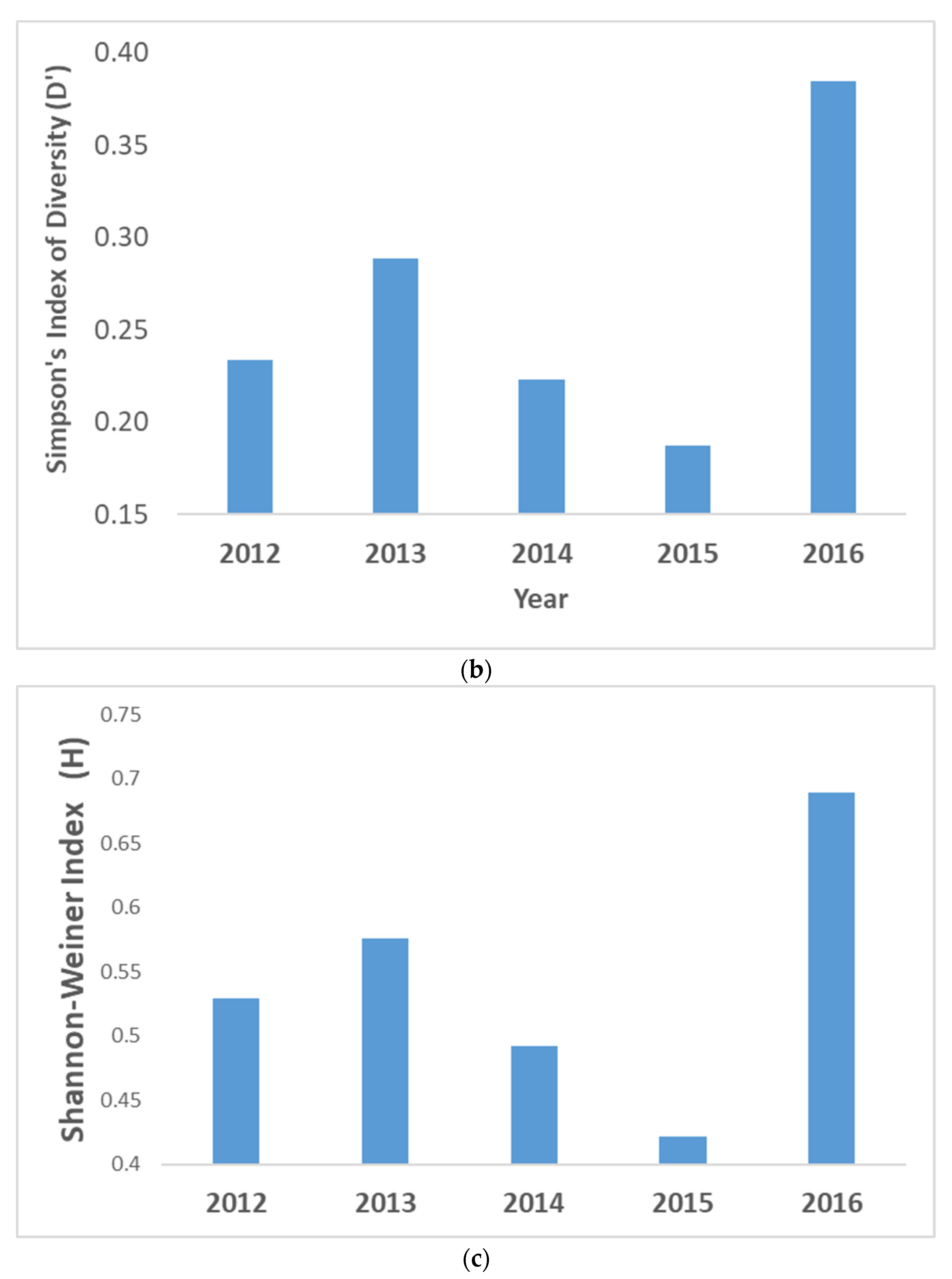
3.1. Changes in Mangrove Community Ecology
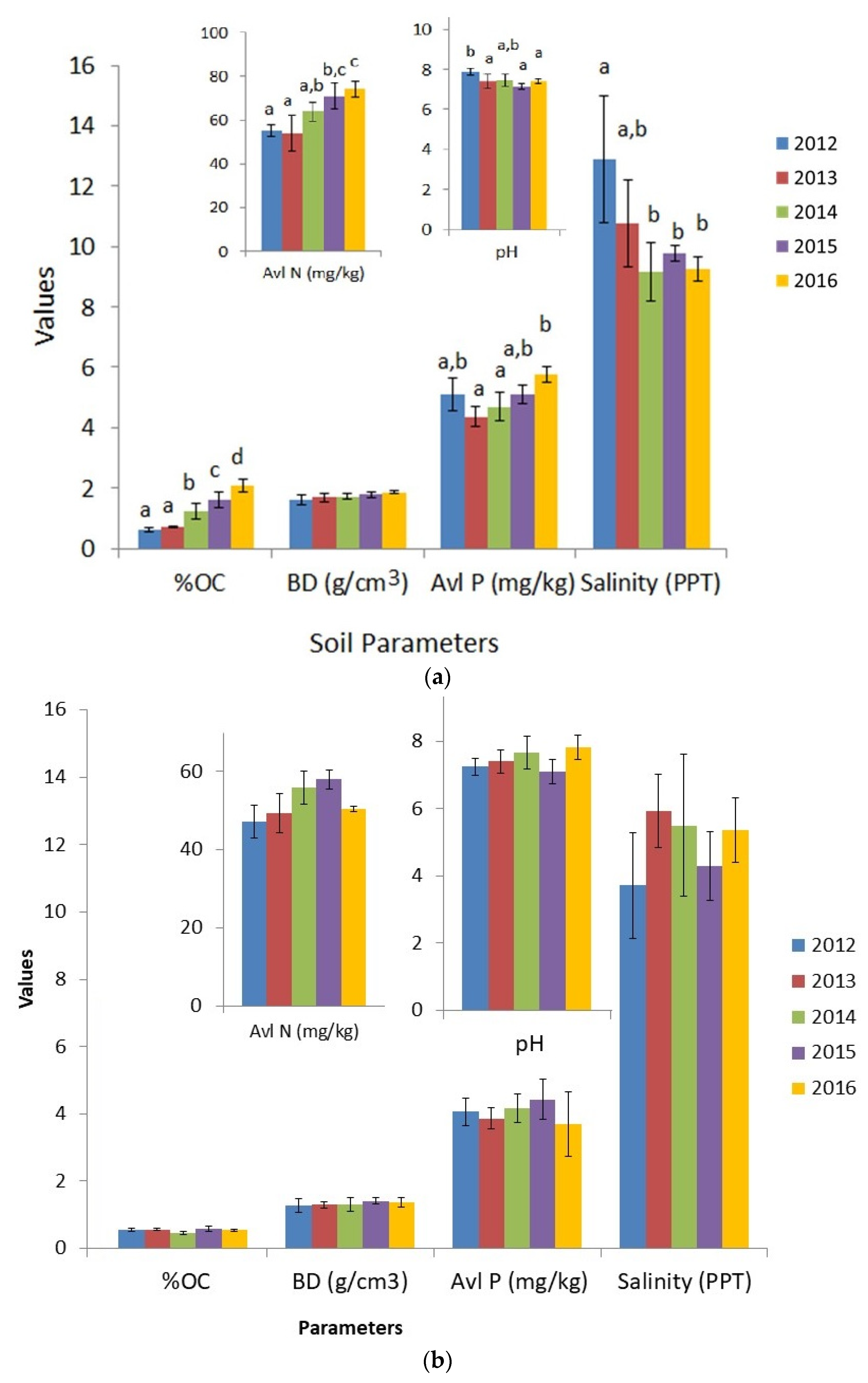
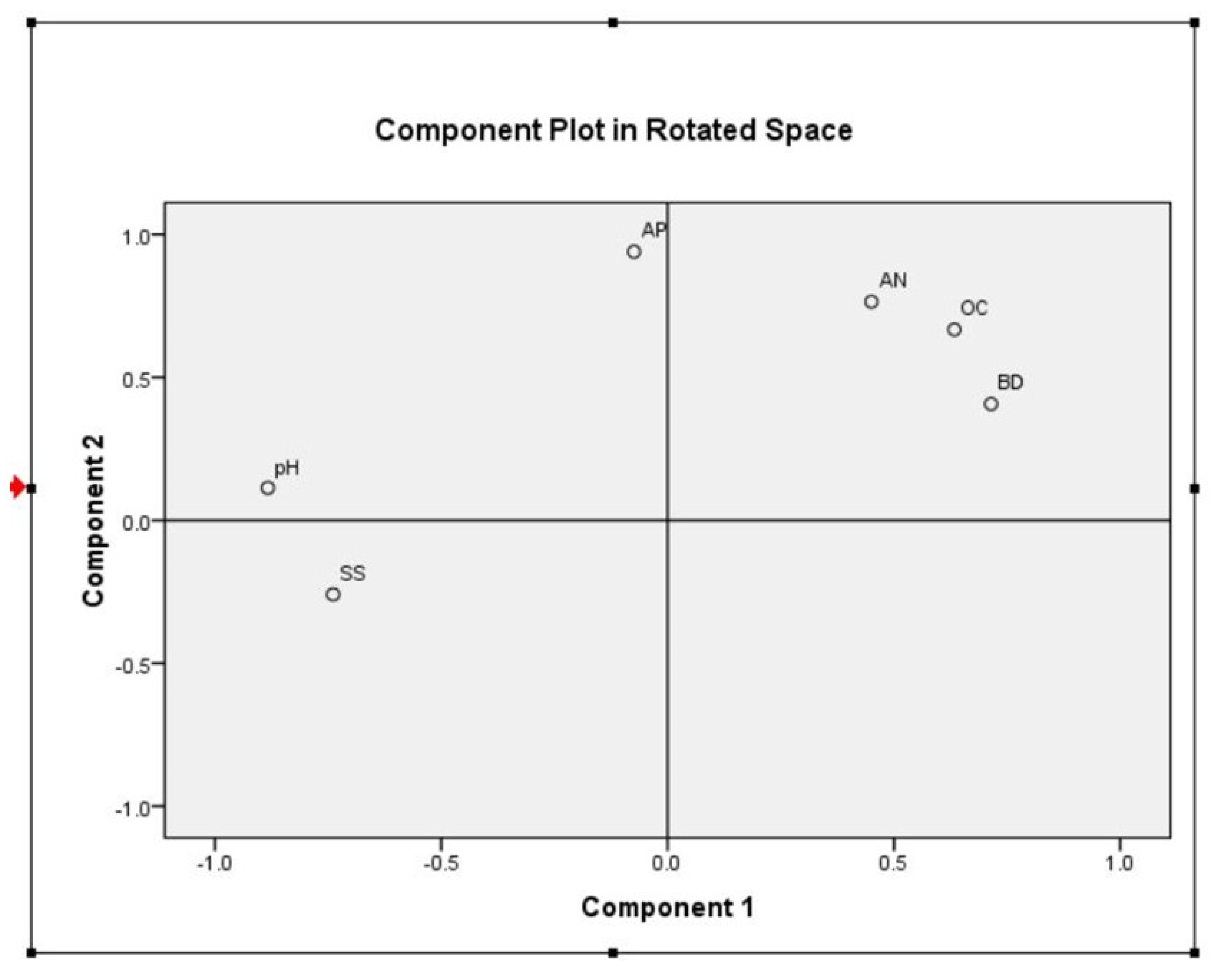
3.2. Variations in Soil Parameters between 2012 and 2016
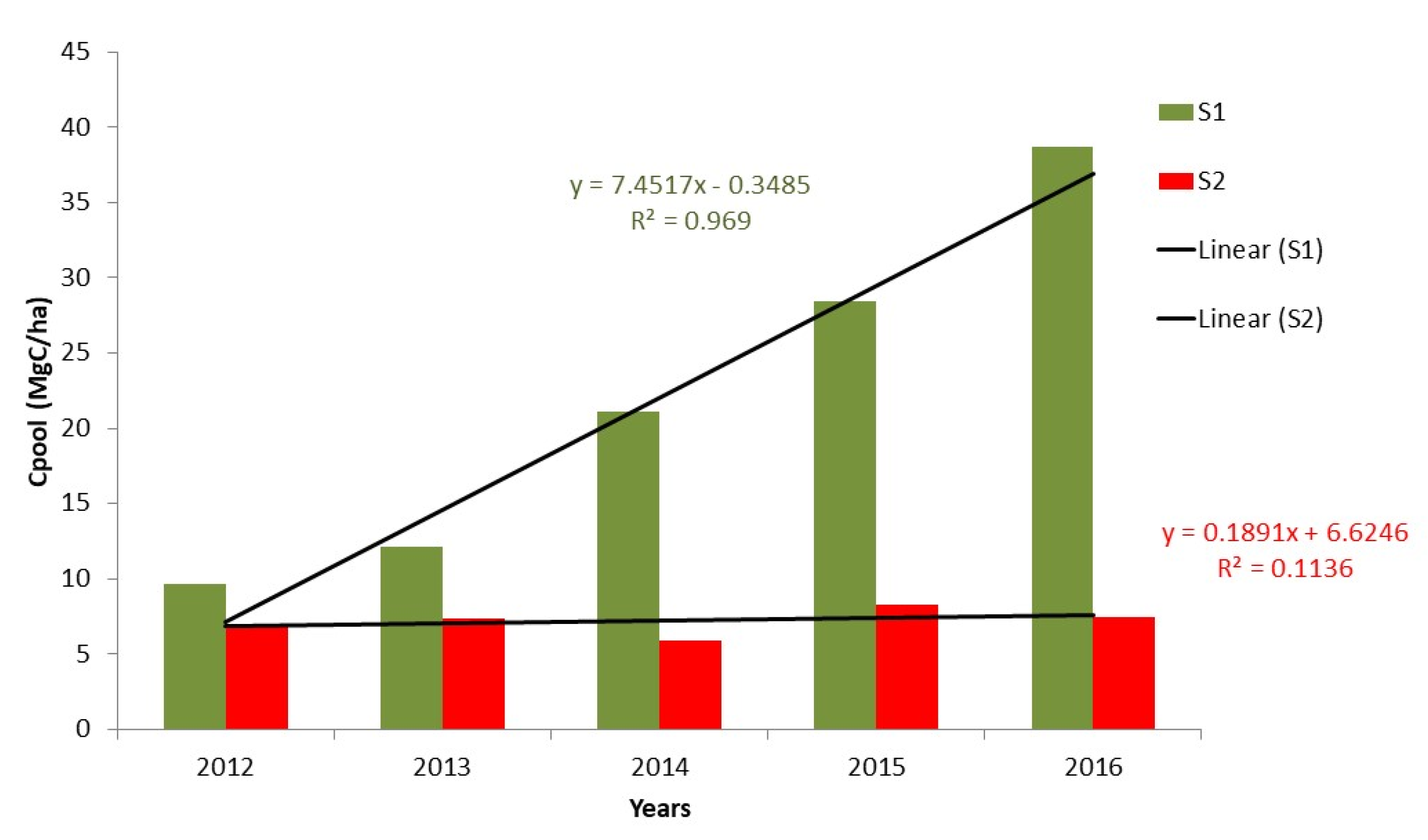
3.3. Blue Carbon Sequestration
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chowdhury, A.; Sanyal, P.; Maiti, S.K. Dynamics of mangrove diversity influenced by climate change and consequent accelerated sea level rise at Indian Sundarbans. Int. J. Global Warm. 2016, 9, 486–506. [Google Scholar] [CrossRef]
- Dasgupta, R.; Hashimoto, S.; Basu, M.; Okuro, T.; Johnson, B.A.; Kumar, P.; Dhyani, S. Spatial characterization of non-material values across multiple coastal production landscapes in the Indian Sundarban delta. Sustain. Sci. 2022, 17, 725–738. [Google Scholar] [CrossRef]
- Donato, D.C.; Kauffman, J.B.; Murdiyarso, D.; Kurnianto, S.; Stidham, M.; Kanninen, M. Mangroves among the most carbon-rich forests in the tropics. Nat. Geosci. 2011, 4, 293–297. [Google Scholar] [CrossRef]
- Bera, B.; Bhattacharjee, S.; Sengupta, N.; Shit, P.K.; Adhikary, P.P.; Sengupta, D.; Saha, S. Significant reduction of carbon stocks and changes of ecosystem service valuation of Indian Sundarban. Sci. Rep. 2022, 12, 7809. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, A.; Naz, A.; Bhattacharyya, S.; Sanyal, P. Cost–benefit analysis of ‘Blue Carbon’ sequestration by plantation of few key mangrove species at Sundarban Biosphere Reserve, India. Carbon Manag. 2018, 9, 575–586. [Google Scholar] [CrossRef]
- Islam, S.M.; Bhuiyan, M.A.H. Sundarbans mangrove forest of Bangladesh: Causes of degradation and sustainable management options. Environ. Sustain. 2018, 1, 113–131. [Google Scholar] [CrossRef]
- Das, S.; Vincent, J.R. Mangrove protected villages and reduced death toll during Indiansuper cyclone’. Proc. Natl. Acad. Sci. USA 2009, 106, 7357–7360. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M. Impact of increased salinity on the plant community of the Sundarbans Mangrove of Bangladesh. Community Ecol. 2020, 21, 273–284. [Google Scholar] [CrossRef]
- Takahashi, Y.; Park, K.J.; Natori, Y.; Dublin, D.; Dasgupta, R.; Miwa, K. Enhancing synergies in nature’s contributions to people in socio-ecological production landscapes and seascapes: Lessons learnt from ten site-based projects in biodiversity hotspots. Sustain. Sci. 2022, 17, 823–836. [Google Scholar] [CrossRef]
- Naskar, S.; Palit, P.K. Anatomical and physiological adaptations of mangroves. Wetl. Ecol. Manag. 2015, 23, 357–370. [Google Scholar] [CrossRef]
- Cano-Ortiz, A.; Musarella, C.M.; Piñar, J.C.; Pinto Gomes, C.J.; del Río González, S.; Cano, E. Diversity and conservation status of mangrove communities in two areas of Mesocaribea biogeographic region. Curr. Sci. 2018, 115, 534–540. [Google Scholar] [CrossRef]
- Basáñez-Muñoz, A.D.J.; Jordán-Garza, A.G.; Serrano, A. Forest Structure and Projections of Avicennia germinans (L.) L. at Three Levels of Perturbation in a Southwestern Gulf of Mexico Mangrove. Forests 2021, 12, 989. [Google Scholar] [CrossRef]
- Macreadie, P.I.; Costa, M.D.; Atwood, T.B.; Friess, D.A.; Kelleway, J.J.; Kennedy, H.; Duarte, C.M. Blue carbon as a natural climate solution. Nat. Rev. Earth Environ. 2021, 2, 826–839. [Google Scholar] [CrossRef]
- Sidik, F.; Supriyanto, B.; Krisnawati, H.; Muttaqin, M.Z. Mangrove conservation for climate change mitigation in Indonesia. Wiley Interdiscip. Rev. Clim. Chang. 2018, 9, e529. [Google Scholar] [CrossRef]
- Gulliver, A.; Carnell, P.E.; Trevathan-Tackett, S.M.; Duarte de Paula Costa, M.; Masqué, P.; Macreadie, P.I. Estimating the potential blue carbon gains from tidal marsh rehabilitation: A case study from south eastern Australia. Front. Mar. Sci. 2020, 7, 403. [Google Scholar] [CrossRef]
- Jimenez, L.C.Z.; Queiroz, H.M.; Otero, X.L.; Nóbrega, G.N.; Ferreira, T.O. Soil Organic Matter Responses to Mangrove Restoration: A Replanting Experience in Northeast Brazil. Int. J. Environ. Res. Public Health 2021, 18, 8981. [Google Scholar] [CrossRef]
- Maiti, S.K. Ecorestoration of Coal Mine Degraded Lands, 1st ed.; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Walkley, A.; Black, I.A. An examination of the Degtjareff method for determining soil organic matter and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Subbiah, B.V.; Asija, G.L. A rapid procedure for the determination of available nitrogen in soils. Curr. Sci. 1956, 25, 259–260. [Google Scholar]
- Olsen, S.R. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate (No. 939); US Department of Agriculture: Washington, DC, USA, 1954. [Google Scholar]
- Tue, N.T.; Dung, L.V.; Nhuan, M.T.; Omori, K. Carbon storage of a tropical mangrove forest in Mui Ca Mau National Park, Vietnam. Catena 2014, 121, 119–126. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Laude, R. Statistics and partitioning of species diversity, and similarity among multiple communities. Oikos 1996, 76, 5–13. [Google Scholar]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–656. [Google Scholar] [CrossRef]
- Acharya, S.; Patra, D.K.; Mahalik, G.; Mohapatra, P.K. Quantitative Ecological Study of Rhizophoraceae Mangroves of Bhitarkanika Wildlife Sanctuary Regions of Odisha Coast, India. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2021, 91, 897–908. [Google Scholar] [CrossRef]
- Agraz-Hernández, C.M.; Chan-Keb, C.A.; Muñiz-Salazar, R.; Pérez-Balan, R.A.; Vanegas, G.P.; Manzanilla, H.G.; del Río Rodríguez, R. Pore Water Chemical Variability and Its Effect on Phenological Production in Three Mangrove Species under Drought Conditions in Southeastern Mexico. Diversity 2022, 14, 668. [Google Scholar] [CrossRef]
- Singh, J.K. Structural characteristics of mangrove forest in different coastal habitats of Gulf of Khambhat arid region of Gujarat, west coast of India. Heliyon 2020, 6, e04685. [Google Scholar] [CrossRef]
- Khan, M.A.; Aziz, I. Salinity tolerance in some mangrove species from Pakistan. Wetl. Ecol. Manag. 2001, 9, 229–233. [Google Scholar] [CrossRef]
- Patel, N.T.; Gupta, A.; Pandey, A.N. Salinity tolerance of Avicennia marina (Forssk.) Vierh. from Gujarat coasts of India. Aquat. Bot. 2010, 93, 9–16. [Google Scholar] [CrossRef]
- Thivakaran, G.A.; Sharma, S.B.; Chowdhury, A.; Murugan, A. Status, structure and environmental variations in semi-arid mangroves of India. J. For. Res. 2020, 31, 163–173. [Google Scholar] [CrossRef]
- Mohanraj, R.; Akil Prasath, R.V.; Rajasekaran, A. Assessment of vegetation, soil nutrient dynamics and heavy metals in the Prosopis juliflora invaded lands at semi-arid regions of Southern India. Catena 2022, 216, 106374. [Google Scholar] [CrossRef]
- Ellison, A.M.; Mukherjee, B.B.; Karim, A. Testing patterns of zonation in mangroves: Scale dependence and environmental correlates in the Sundarbans of Bangladesh. J. Ecol. 2000, 88, 813–824. [Google Scholar] [CrossRef]
- Fang, X.M.; Wang, G.G.; Xu, Z.J.; Zong, Y.Y.; Zhang, X.L.; Li, J.J.; Chen, F.S. Litter addition and understory removal influenced soil organic carbon quality and mineral nitrogen supply in a subtropical plantation forest. Plant Soil 2021, 460, 527–540. [Google Scholar] [CrossRef]
- AboEl-Nil, M.M. Growth and establishment of mangrove (Avicennia marina) on the coastlines of Kuwait. Wetl Ecol Manag. 2001, 9, 421–428. [Google Scholar] [CrossRef]
- Feng, J.; Cui, X.; Zhou, J.; Wang, L.; Zhu, X.; Lin, G. Effects of exotic and native mangrove forests plantation on soil organic carbon, nitrogen, and phosphorus contents and pools in Leizhou, China. Catena 2019, 180, 1–7. [Google Scholar] [CrossRef]
- Muñoz-Rojas, M. Soil quality indicators: Critical tools in ecosystem restoration. Curr. Opin. Environ. Sci. Health 2018, 5, 47–52. [Google Scholar] [CrossRef]
- Salmo, S.G.; Lovelock, C.; Duke, N.C. Vegetation and soil characteristics as indicators of restoration trajectories in restored mangroves. Hydrobiologia 2013, 720, 1–18. [Google Scholar] [CrossRef]
- Cardona, P.; Botero, L. Soil characteristics and vegetation structure in a heavily deteriorated mangrove forest in the Caribbean coast of Colombia. Biotropica 1998, 30, 24–34. [Google Scholar] [CrossRef]
- Begam, M.; Sutradhar, T.; Chowdhury, R.; Mukherjee, C.; Basak, S.K.; Ray, K. Native salt-tolerant grass species for habitat restoration, their acclimation and contribution to improving edaphic conditions: A study from a degraded mangrove in the Indian Sundarbans. Hydrobiologia 2017, 803, 373–387. [Google Scholar] [CrossRef]
- Mandal, A.K.; Nandi, N.C. Fauna of Sundarban Mangrove Ecosystem, West Bengal, India; Zoological Survey of India: Kolkata, India, 1989; Volume 3. [Google Scholar]
- Preston, J.C.; Fjellheim, S. Understanding past, and predicting future, niche transitions based on grass flowering time variation. Plant Physiol. 2020, 183, 822–839. [Google Scholar] [CrossRef]
- Ellison, A.M. Mangrove restoration: Do we know enough? Restor. Ecol. 2000, 8, 219–229. [Google Scholar] [CrossRef]
- Zhang, J.; Shen, C.; Ren, H.; Wang, J.; Han, W. Estimating change in sedimentary organic carbon content during mangrove restoration in southern china using carbon isotopic measurements. Pedosphere 2012, 22, 58–66. [Google Scholar] [CrossRef]
- Lee, S.Y.; Hamilton, S.; Barbier, E.B.; Primavera, J.; Lewis, R.R. Better restoration policies are needed to conserve mangrove ecosystems. Nature Ecology & Evolution. Nat. Ecol. Evol. 2019; 3, 870–872. [Google Scholar]
- Su, J.; Friess, D.A.; Gasparatos, A. A meta-analysis of the ecological and economic outcomes of mangrove restoration. Nat. Commun. 2021, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Saderne, V.; Geraldi, N.R.; Macreadie, P.I.; Maher, D.T.; Middelburg, J.J.; Serrano, O.; Almahasheer, H.; Arias-Ortiz, A.; Cusack, M.; Eyre, B.D.; et al. Role of carbonate burial in Blue Carbon budgets. Nat. Commun. 2019, 10, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Cuellar-Martinez, T.; Ruiz-Fernández, A.C.; Sanchez-Cabeza, J.A.; Perez-Bernal, L.H.; Sandoval-Gil, J. Relevance of carbon burial and storage in two contrasting blue carbon ecosystems of a north-east Pacific coastal lagoon. Sci. Total Environ. 2019, 675, 581–593. [Google Scholar] [CrossRef] [PubMed]
- Hasibuan, A.; Supriatna, A.K.; Carnia, E. Local stability analysis of two density-dependent semelparous species in two age classes. Front. Appl. Math. Stat. 2022, 8, 953223. [Google Scholar] [CrossRef]
- Lundgren, M.R.; Des Marais, D.L. Life history variation as a model for understanding trade-offs in plant–environment interactions. Curr Biol. 2020, 30, 180–189. [Google Scholar] [CrossRef]
- Akhand, A.; Chanda, A.; Dasgupta, R. Advancement in Measurement and Estimation Methods of Blue Carbon Studies. Assessing, Mapping and Modelling of Mangrove Ecosystem Services in the Asia-Pacific Region; Springer: Berlin/Heidelberg, Germany, 2022; pp. 127–142. [Google Scholar]
- Kadaverugu, R.; Dhyani, S.; Dasgupta, R.; Kumar, P.; Hashimoto, S.; Pujari, P. Multiple values of Bhitarkanika mangroves for human well-being: Synthesis of contemporary scientific knowledge for mainstreaming ecosystem services in policy planning. J. Coast. Conserv. 2021, 25, 1–15. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sand | Silt | Clay | |
|---|---|---|---|
| S1 | 1.60 ± 0.17 | 79.57 ± 1.27 | 18.83 ± 1.31 |
| S2 | 1.83 ± 0.15 | 80.77 ± 1.72 | 17.40 ± 1.83 |
| 2012 | 2013 | 2014 | 2015 | 2016 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Number | Species | Family | RD | F | RD | F | RD | F | RD | F | RD | F |
| 1 | Acanthus ilicifolius L. | Acanthaceae | 7 | 100 | 4 | 100 | 12 | 60 | 3 | 60 | 2 | 60 |
| 2 | Avicennia marina (Forssk.) Vierh. | Acanthaceae | 2 | 60 | 11 | 80 | 4 | 100 | 5 | 100 | 20 | 100 |
| 3 | Ceriops tagal (Perr.) C.B.Rob | Rhizophoraceae | 0 | 40 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 4 | Phoenix paludosa Roxb. | Arecaceae | 2 | 40 | 1 | 40 | 5 | 60 | 1 | 60 | 2 | 80 |
| 5 | Proteresia coarctata (Roxb.) Tateoka | Poaceae | 87 | 100 | 84 | 100 | 78 | 100 | 90 | 100 | 76 | 100 |
| 6 | Sonneratia caseolaris (L.) Engl. | Lythraceae | 1 | 20 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chowdhury, A.; Naz, A.; Dasgupta, R.; Maiti, S.K. Blue Carbon: Comparison of Chronosequences from Avicennia marina Plantation and Proteresia coarctata Dominated Mudflat, at the World’s Largest Mangrove Wetland. Sustainability 2023, 15, 368. https://doi.org/10.3390/su15010368
Chowdhury A, Naz A, Dasgupta R, Maiti SK. Blue Carbon: Comparison of Chronosequences from Avicennia marina Plantation and Proteresia coarctata Dominated Mudflat, at the World’s Largest Mangrove Wetland. Sustainability. 2023; 15(1):368. https://doi.org/10.3390/su15010368
Chicago/Turabian StyleChowdhury, Abhiroop, Aliya Naz, Rajarshi Dasgupta, and Subodh Kumar Maiti. 2023. "Blue Carbon: Comparison of Chronosequences from Avicennia marina Plantation and Proteresia coarctata Dominated Mudflat, at the World’s Largest Mangrove Wetland" Sustainability 15, no. 1: 368. https://doi.org/10.3390/su15010368