

Designing a Seed Health Strategy for Organic Cropping Systems, Based on a Dynamic Perspective on Seed and Plant Health

1
Organic Food and Farming Institute (ITAB)—149, Rue de Bercy, 75595 Paris, France
2
Socio-Economie, Environnement, Développement (SEED), Département Sciences et Gestion Environnement, Université de Liège Campus Arlon, 185 Avenue de Longwy, B-6700 Arlon, Belgium
3
Wageningen Plant Research, Wageningen University & Research, Droevendaalsesteeg 1, 6708 PB Wageningen, The Netherlands
*
Author to whom correspondence should be addressed.
Sustainability 2022, 14(17), 10903; https://doi.org/10.3390/su141710903
Submission received: 30 June 2022
/
Revised: 11 August 2022
/
Accepted: 22 August 2022
/
Published: 31 August 2022
(This article belongs to the Special Issue Breeding and Seed Sector Innovations for Organic Food Systems)
Abstract
:Seed germination and seedling establishment are the main factors for a successful start of a crop, determining crop health, weed suppression and overall crop performance. They are critical, but sensitive phases in crop production. High seed vigour and appropriate seed microbiota significantly contribute to the resilience of seedlings. This opinion paper is based on a review of relevant literature, two case studies with wheat and carrot and a subsequent stakeholder consultation in the framework of the European project LIVESEED. It presents a seed health strategy for organic farming systems that accounts for dynamic processes behind seed and plant health, taking advantage of the contributions of seed vigour and seed microbiota. Seed and plant health are understood as a continuum. Benefits of seed vigour and microbiota are showcased. Recommendations are given for organic seed production, seed processing and storage, as well as organic plant breeding and future research. In conclusion, organic farming systems, in particular, have the opportunity, both to develop this perspective on seed and plant health and to benefit from it. Adopting this perspective may require a shift in how crop plants and their seeds are cared for, as well as adapted regulatory frameworks.
1. Introduction
Crop production starts with preparing the soil and sowing of the seed. Farmers’ seed choice is mainly based on the cultivar best suited for producing the crop under the given environmental conditions and may include resistance against pests or diseases. But for a successful start of the crop, the physical, physiological, and health quality of the seeds is at least as important. Seed germination and seedling establishment are critical but also sensitive phases in crop production. Healthy, vigorous seeds will emerge rapidly and uniformly, provide fast growing root systems to retrieve water and nutrients, as well as a fast-growing shoot to capture sunlight and produce energy, related to the exponential growth rate of seedlings [1]. This will both give the seedling more tolerance to abiotic and biotic stresses in the field including more capacity to compete with weeds [2]. Thereby, the quality of the seeds affects how well a crop will establish, influencing crop yield, farmers’ incomes and, ultimately, food security. European and national regulations demand minimal qualifications of seed quality in purity, germination and health (e.g., for vegetable seed [3] and cereal seed [4]). To assess seed quality, routine seed testing evaluates seed germination rates under optimal laboratory conditions and detects potential seedborne pathogens, according to crop species. The common perspective on seed health thus focusses on keeping out seedborne plant diseases. If a problematic level of seedborne pathogen is detected, seed treatments can be used for disease control.
In organic farming, to comply with the European organic regulation [5], these treatments range from natural compounds (e.g., vinegar or plant extracts) and physical treatments (e.g., hot water, steam, or brush cleaning) to the application of so-called ‘biologicals’ (e.g., antagonistic micro-organisms such as Pseudomonas chlororaphis strains). Health in general, and plant health in particular, is not defined in the same way by everyone involved in organic crop production. While some are satisfied with curative measures to sustain plant health, others prefer to base plant health on more self-sustaining ecological and biological interactions [6,7]. Both practical experience and recent scientific findings indicate that considering additional aspects to seed health would benefit organic agriculture and other forms of agroecological agriculture [8,9], that refrain from the use of synthetic chemical inputs. In this aspect, seed vigour and seed microbiota should receive growing attention as factors contributing to the performance of seeds and seedlings.
Seed vigour is defined by the International Seed Testing Association as the sum of those properties that determine the activity and performance of seed lots of acceptable germination in a wide range of environments [10]. It describes the resilience of seeds and seedlings, i.e., their ability to germinate and emerge under field stresses. Despite all the care taken by farmers before and during sowing, field conditions can vary, which can put a strong restraint on the establishment of seedlings. Such uncertainties and unpredictable stress factors are expected to increase with global climate change.
The plant microbiota is understood as the microbial ecosystem, composed of bacteria, fungi, and other micro-organisms, associated with plants. The composition of the microbiota is determined by sequencing their DNA, providing information on all microbial genomes present, called the microbiome. However, in practice the term microbiome is frequently used to refer to the community of micro-organisms [11]. Microbiota have been found to impact the growth and survival of plants harbouring them, leading to the perception of plants not as stand-alone entities, but as holobionts [12]. Plants transfer part of their microbiota to the next generation as the seed microbiota [13]. Until the present, attention has almost exclusively been paid to pathogens among these seed microbiota, called seed borne pathogens. But their pathogenicity seems to depend on the concentration and on the general composition of the seed microbiome [14,15]. Seed can also contain micro-organisms that aid the seedling against pathogens or abiotic stresses to support seedling establishment, as described in the next section.
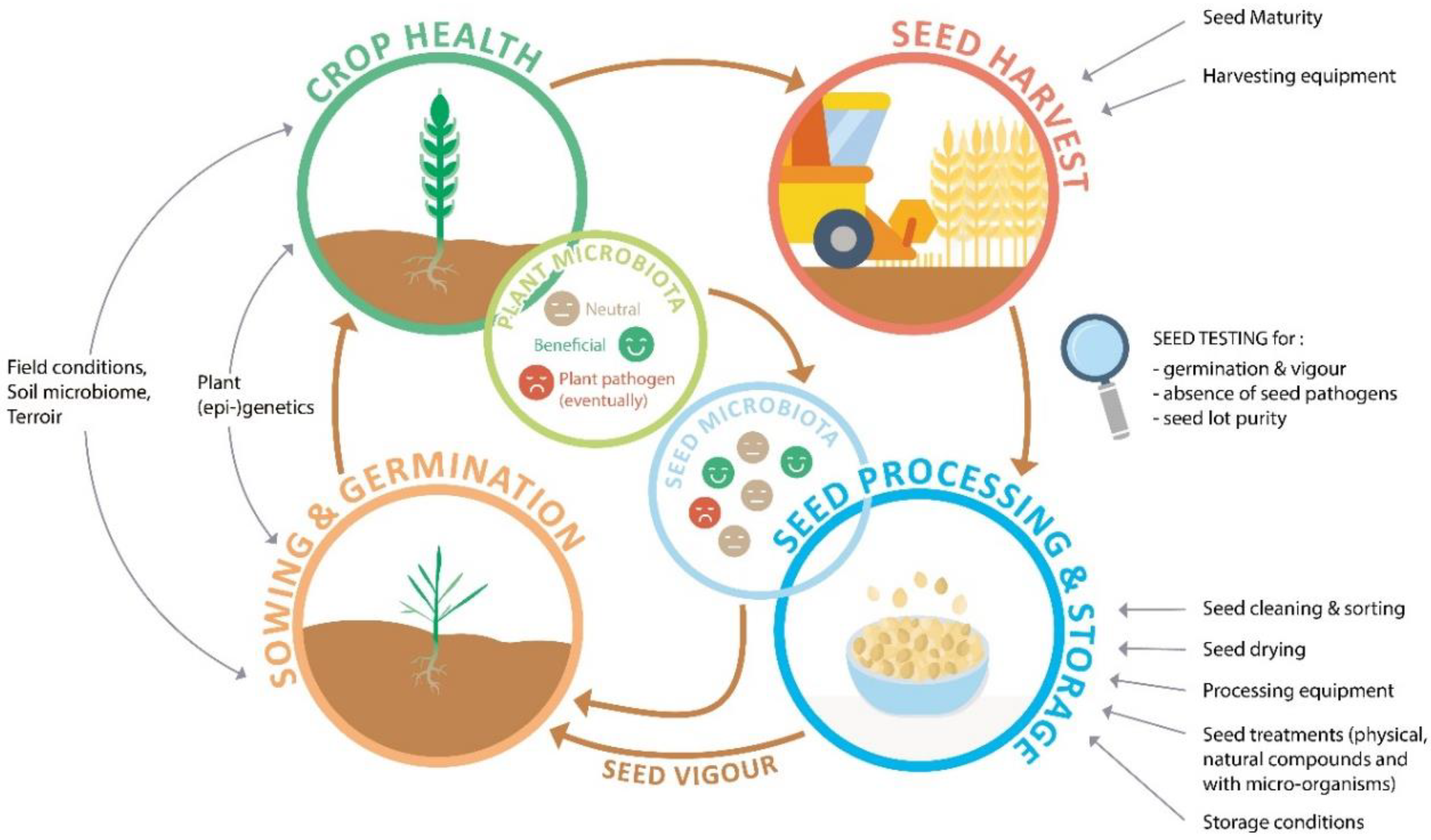
Seed quality and seed health are characteristics commonly considered separately, but they blend into each other as both are influenced by seed vigour and the microbiota. Pathogenicity also depends on the tolerance or resistance of the of the host, the vigour of the seed and seedling and environmental factors [14]. These considerations broaden the picture on seed and plant health beyond the mere detection of pathogens. Seeds are the outcome of a seed production process in which a multitude of abiotic and biotic (including human) factors are at work, that can interfere with seedling and plant health, both in positive and negative ways. Figure 1 represents seed and plant health as a cyclical continuum, highlighting how they are affected by environmental factors and management practices all along the plant life cycle.
In the framework of the European research project LIVESEED [16], we sought to devise a seed health strategy for organic farming systems that fully accounts for the dynamic processes behind seed and plant health, and which takes advantage of the contributions of seed vigour and seed microbiomes. The European organic regulation [5] requires that seeds used in organic agriculture are produced under organic conditions for at least one generation. Such seeds are referred to as ’organic seeds’. For the seed health strategy, we based our reflections on a review of relevant literature, complemented by two main case studies: damping-off with carrot (Daucus carota) seedlings caused by Alternaria radicina and the management of common bunt caused by Tilletia spp. on wheat (Triticum aestivum) in organic farming systems. Methods used in these case studies ranged from laboratory analysis over field experiments to farmers’ workshops [17]. Feedback on the proposed seed health strategy was then collected from seed growers, seed companies, breeders, associations concerned with organic seed and researchers at an online workshop (nine participants from six European countries), as well as an online consultation in English (14 responses from seven European countries and the USA) and French (four responses from three European countries), all held in 2021.
In the following Section 2, we will show how seed vigour and microbiomes contribute to seed and plant health. Then (Section 3), implications for plant breeding, seed production and seed sanitation are described, as well as numerous open questions. Strategic next steps to improve seed health are listed in Section 4, including research topics to be addressed and key actions to put into practice. Main conclusions are drawn in Section 5.
2. Contributions of Seed Vigour and Microbiota to Seed and Plant Health
In the past decades, plant pathologists have extended the basic “disease triangle” composed of a plant pathogen, its host, and an environment favourable to the respective disease [18]. Authors have added a range of additional factors [19] including time [20,21], humans [22] and vectors [23], thus representing the basic parameters of plant disease as a tetrahedron or pyramid. As regards the management of seedborne diseases, a closer look at seed vigour and microbiota allows for a more comprehensive understanding of the dynamic processes that lead to seedling and crop health.
2.1. Seed Vigour
High seed vigour enhances the resilience of seeds and young seedlings to environmental stress conditions [24,25]. Biotic stress can be due to pathogens or competing plants, whereas abiotic stress can occur for instance from ageing during seed storage, too low or high temperatures during germination, drought, salinity, or flooding. As resilience can differ between stresses, it is best to define seed vigour with an indication of the type of stress concerned, e.g., cold or salinity during germination.
Seed vigour is determined by a combination of factors: conditions in which the seed was produced and harvested, storage, seed treatments and plant genetics. During the last phase of seed development, several protection mechanisms are imposed to limit damage induced during storage [26]. This protection includes antioxidants and production of so-called late embryogenesis abundant proteins, that can act as chaperones for structural proteins or for condensed packaging of the DNA [27]. Finishing this maturation process is very important for optimal seed vigour, not only in relation to seed ageing, but also for tolerance towards physical sanitation treatments [28] and for a rapid germination and field emergence. Most of our crop seeds are desiccation tolerant. Seed quality nevertheless declines during storage, mainly due to oxidative damage to membranes, proteins, RNA and DNA [26]. Seed ageing is stimulated at higher humidity and temperatures and by oxygen. For that reason, gene banks are recommended to store seeds vacuum sealed at −20 °C after drying at around 15–20% RH. Seed companies store relatively expensive horticultural seeds in conditioned warehouses at 15 or 20 °C and 30% RH. More bulky cereal seed lots are dried and subsequently often stored in non-conditioned warehouses. Damage that has accumulated during storage can only be repaired enzymatically upon rewetting of the seed at sowing, but this takes energy and time. Therefore, aged seeds will germinate more slowly or not at all, depending on the amount of damage accumulated.
Seed vigour can be influenced by genetic components. Generally known is the poor vigour of sweet corn seeds because of their lack of starch, needed to provide energy for the emerging seedling. With kale crops (Brassica oleracea), genes have been identified that influence the speed of germination or upward shoot growth in soil of increasing impedance [29]. Genetic loci for seed vigour have also been identified with rice (Oryza sativa) and wheat [30,31]. Legumes often show genetic variation for seed coat colours. The flavonoids responsible for this can play a role as antioxidants and in the water permeability of the seed coat. As such they can influence the tolerance against ageing [32].
Seed vigour plays a role in biotic stress tolerance. Almost 50 years ago it was shown with wheat, that in contrast to low vigour seeds, high vigour seeds had little need for fungicide treatment to maintain yield levels [33]. Although the effect may depend on the crop, the pathogen pressure and other field conditions, this demonstrated the potential advantage of high vigour seeds for farming. A study with spinach (Spinacia oleracea) seeds showed that smaller seeds with a thinner pericarp are less sensitive to high soil moisture conditions that limit oxygen diffusion [34], whereas less mature spinach seeds are more sensitive to the soil borne pathogen Pythium ultimum [35]. In the frame of the LIVESEED project we studied the effect of vigour on biotic stress tolerance using carrot seeds and the pathogen Alternaria radicina, which can cause seedling ‘damping-off’ by infecting and damaging the root system. Lower vigour had been obtained by storing seeds at 40 °C and 85% relative humidity for four days. The study showed that high vigour carrot seeds were more tolerant to the pathogen compared to the seeds with reduced vigour (Figure 2). Optimisation of seed harvesting and storage, in combination with seed sorting and breeding for higher seed vigour, therefore, offers possibilities to create more resilient cropping systems.
2.2. Seed Microbiota
In recent years it has been shown that many seed-associated bacteria or fungi can aid the seed and seedling in the defence against pathogens, in germination, seedling development and nutrient acquisition [36,37]. A part of the seed microbiome originates from the mother plant, while micro-organisms from the external environment can also enter the seed or adhere to its surface. With rice for example, Sphingomonas melonis bacteria can be transmitted through the seeds and provide the seedling with resistance to damping off caused by the pathogen Burkholderia plantarii, while transmission of the beneficial S. melonis bacteria varies with seed production conditions [38]. Bacteria isolated from wild cotton (Gossypium hirsutum) seed fibres can promote germination, root growth and alleviate salt stress of cultivated cotton seedlings grown from seeds where the fibres were removed (delinted) using an acid [39]. For tomato (Solanum lycopersicum), seed-associated microbiota can protect seedlings against Pseudomonas syringae pv. tomato establishment and disease [40]. With wheat, bacteria isolated from the seed microbiome from drought tolerant wheat lines can induce drought tolerance in sensitive wheat lines by soaking their seeds in a suspension of these supporting bacteria, a treatment known as biopriming [41]. Interestingly, this study also showed that several different genera of seed microbiome bacteria could induce this drought tolerance.
When not present in foundation seeds, a beneficial seed microbiome may be introduced during seed production, e.g., by spraying the seed crop with inoculum during flowering. This has successfully been demonstrated at an experimental scale with soybean (Glycine max), pepper (Capsicum annuum) and wheat [42]. If and how this could be applied on a large scale has to be studied. When used directly as seed dressing (as so-called biologicals) for supporting the seedling, seed endophytes have the advantage, by definition, of tolerating dry storage. The observation that for instance drought stress tolerance can be induced by different taxa of bacteria from the seed microbiome [41], indicates that it may be possible to isolate candidate microbial taxa from the crop species itself and potentially from locally grown (wild) plant populations. This would avoid the introduction of micro-organisms and symbioses, that do not occur naturally in a given region.
The seed production environment influences the composition of the seed microbiome. When plants are exposed to biotic or abiotic stress, they can restructure their microbiome as a ‘cry for help’ [43]. Invasion by phytopathogenic micro-organisms can modify seed microbial assemblages [44] and the opposite is also true: seeds may carry endophytic taxa which act as antagonists to pathogenic fungi [45,46]. With tomato it has been shown that plants preferentially transmit beneficial micro-organisms to the seed microbiome [47]. If this turns out to be broadly applicable, steering the seed microbiome during seed production may be possible. While in general for the seedling shoot the microbiome mainly originates from the seed [48], the soil seems the main source from which older plants recruit their microbiome [47]. Diversity of the soil microbiome is therefore expected to be important for plants in recruiting their helpers and ultimately for the composition of the seed microbiome. In that sense, the higher microbial diversity associated with organic farming practices [49,50,51] may also aid in establishing more diverse seed microbiomes.
The mentioned examples illustrate how both seed vigour and seed microbiota contribute to the health of seeds, seedlings and, ultimately, crops. As the seed microbiota are intrinsically linked with seed vigour, more research is needed to understand how the seed as organism and its microbiota mutually affect each other. In our carrot seed vigour experiments for instance, it is not clear if the reduced tolerance of partly deteriorated carrot seeds towards the Alternaria pathogen (Figure 2) is due to damage induced to the seed itself, e.g., its mitochondria or membranes, or due to damage to its microbiota.
3. Implications—And Open Questions—For Plant Breeding, Seed Production and Seed Sanitation
The extended view on seed and plant health presented above has a range of implications in all the domains that concern seeds—from plant breeding over seed production and sanitation, including their legal frameworks. These implications come with their share of open questions.
- Plant breeding
To date, considerations for plant and seed health in plant breeding have mainly led to breeding programs for resistance based on plant genetics to facilitate the control of plant diseases in crops. Holobiont breeding [52] has the potential to integrate the role of seed-transmitted microbiota [53] and genetic effects on seed vigour to improve seed and plant health in the future, as a complement to resistance breeding. Domestication, modern plant breeding and cultivation practices have unintentionally reduced the general capacity of crop plants to recruit and host beneficial micro-organisms [54,55]. By embracing the holobiont concept, such tendencies might be reversed, at least partly [56,57]. For instance, organic breeding programmes have selected for plant genotypes with an enhanced ability to recruit endemic beneficial microbial communities from the crop environment in the maize (Zea mays) rhizosphere [58] and taproot endophyte composition in carrot [59].
Others have suggested to propagate plant cultivars in combination with associated micro-organisms, by providing a starter microbiome culture of keystone plant-beneficial microbiota from target soils, adapted to a given plant genotype [60]. However, inoculating soils or plants with individual microbial taxa or communities faces challenges as concerns the survival of introduced taxa and their vertical transmission to the next generation. As mentioned, selected microbiota can be introduced into the seed microbiome and transferred to the next plant generation, e.g., by inoculating flowers of the seed crop [42] or by incubating the seeds in a buffer with the microbiota, but this may ensure transmission only over one plant generation. This only provides a solution when seeds are produced by specialised companies [52]. Holobiont breeding and seed production strategies relying on vertical transmission of microbiota via the seeds would be a durable approach to deliver resilient cropping systems. Research has provided evidence that crop genetics can influence the seed microbiome, at least on seed endophyte composition (e.g., [61]). Putting these findings into practice in breeding programmes yet requires a more comprehensive understanding of the composition and effects of the seed microbiota and of factors influencing their transmission.
Selection for plant genotypes that contribute to the production of high-vigour seeds could be strengthened in organic breeding programs. Especially in systems with low soil fertility, negative effects of stress factors on seedlings may thereby be mitigated. Organic nitrogen fertilization leads to lower protein content in harvested grain as compared to systems with mineral nitrogen fertilizer [62], possibly putting a limitation on seed vigour. With wheat and maize seeds, the absolute content of protein is highly correlated with seedling dry weight [63]. This highlights an additional reason to breed for genotypes with a high nitrogen use efficiency [64]. One may hypothesize that breeding under high-input conventional agriculture, with the use of mineral fertilizers and fungicide seed treatments has not allowed to select for plant genotypes providing seedlings with a high tolerance to biotic and abiotic stress, for instance with a fast and more extensively growing root system. In that case, breeding under organic conditions may well present an opportunity to improve seed and seedling vigour under low to moderate nutrient conditions [65]. In this also a genetic interaction with a supportive microbiome may be helpful.
Plant epigenetics—i.e., processes that affect transcriptional regulation and expression levels of plant genes, conferring phenotypic variation in response to environmental changes—are an additional factor of plant health that can be mediated by seeds. Challenged by pathogenic viruses, fungi, bacteria, host plants can go through epigenetic modifications inducing defence mechanisms [66,67]. These modification patterns can be transmitted to subsequent generations by inheritance, thereby equipping the progeny with heightened resistance. This has been demonstrated for several crop species, especially rice and wheat, opening new perspectives on the potential of harnessing epigenetic variations for crop disease resistance or tolerance in an approach also called “epi-breeding” [68].
Consequently, breeding plants under organic farming conditions does not only allow to select for the most appropriate crop genotypes within target environments presenting abiotic and biotic constraints. It also offers the opportunity to benefit from plant-environment interactions at microbial and epigenetic levels that strengthen the resilience of seedlings and to develop crops that efficiently draw from these interactions, provided that further research yields knowledge and recommendations that can be put into breeding practice.
- Seed production
As also indicated in Section 2 and Figure 1, healthy, well-nurtured and timely harvested seed crops are likely to produce the most vigorous seedlings [69]. Therefore, fertile plots with low disease pressure are more appropriate to produce well-filled, healthy, and vigorous seeds. In on-farm, collaborative breeding, crops are sometimes willingly exposed to low soil fertility, drought, or other stress factors to select for stress tolerance and general robustness in open-pollinated cultivars [70,71]. While this may present advantages to select for locally most adapted genotypes, it does not favour optimal seed filling and seed vigour. In such a context, it may be preferable to dissociate on-farm selection from seed production, conducting the latter under fertile and favourable conditions.
Integrating the microbiome and epigenetics in plant breeding (see previous subsection), would also require appropriate seed multiplication environments and practices to ensure that epigenetic and microbial properties are transferred reliably. Research efforts are needed to find methods to steer the seed microbiome toward support of the seed and subsequent seedling against certain environmental stresses. As referred to in Section 2, there are indications that the mother plant recruits specific microbiota into the seed microbiome, in relation to the stress it experiences. Inoculating seed crops or seeds with supporting micro-organisms may be an alternative, as was demonstrated with drought tolerant microbiota that were isolated from wheat seeds [41,42]. Research is needed to find a balance between providing stress during seed production to obtain an optimised seed microbiome on the one hand and preventing too much loss of other aspects of seed vigour, on the other.
In the long run, integrating the seed and plant microbiomes in how we produce seeds may also affect recommendations on the geographical scale of seed production. Currently, most vegetable seeds are produced and transported globally, for reasons of optimal seed production climates, counter season seed production and optimized production costs. Research with common bean (Phaseolus vulgaris) has shown that local environmental factors described as “terroir” shape the composition of seed microbiomes [72]. Berg and Raaijmaakers [73] warned that the “centralized production and global trade of seeds may contribute to homogeneity of the plant microbiome at global scale but may also impact on soil microbial diversity and health”. They argue to “save seed microbes”. Local seed systems and community seed banks worldwide have long since been recognized as major actors of crop genetic diversity preservation [74,75,76]. Cultivation and use of locally adapted cultivars and local, small-scale seed production may also contribute to the preservation of diversified, locally adapted seed microbial communities.
As yet, it remains widely unknown how the structure of plant microbial communities varies at different geographical scales and whether or not locally produced seeds provide some advantage in the form of microbial adaptation to local conditions. Achieving such a level of comprehension still requires considerable research both into the factors shaping seed and plant microbiota and into their functional attributes. This includes developing an understanding of the effects of moving seeds across geographic regions on seed microbial communities and, consequently, on plant health. According to the findings of such future research, gene banks that are currently responsible for the ex-situ conservation of genetic crop diversity, could in future embrace the mission of conserving not only plant genetic diversity, but also the diversity of microbial communities associated with the seeds they store [73,77].
- Seed treatments
When seeds are infected with pathogens, sanitation treatments may be necessary. It is likely that treatments with some physical methods and natural compounds authorised in organic agriculture also reduce a significant part of the seed microbiome (e.g., [78]). Moreover, treatments aimed to destroy pathogens often have negative influences on seed vigour. Therefore, the use of treatments to eradicate pathogenic micro-organisms may be perceived as contradictory with the agroecological transformation [8,9] of cropping systems based on resilient ecosystem interactions and thriving microbial. We argue that both strategies may be complementary: bio-diverse, resilient cropping systems (in combination with sound organic cropping and seed production practices) may strongly reduce the need for intervention with seed sanitation treatments. Nevertheless, seed sanitation is expected to remain sometimes necessary, at least for quarantine or other regulated pathogens (e.g., Clavibacter michiganensis ssp. michiganensis, the causal agent of bacterial canker disease with tomato), or as occasional support or safety net (e.g., for management of common bunt in wheat).
Seed surface disinfection, by physical methods or natural compounds, can create an empty niche for opportunistic pathogens in the soil, e.g., several pathogens causing damping-off of seedlings. According to the experience of one of the seed companies participating in the above-mentioned workshops (see introduction), physically disinfected seeds are less sensitive to soil borne pathogens when sown in organic compared to conventionally managed soil. Indeed, disinfected seeds will largely lack micro-organisms on their surface offering an open space for re-colonisation upon sowing. A higher biodiversity in soil microbiota [49,50,51] may have a greater capacity to re-colonize the seeds with positive or neutral microbiota and thereby buffer against colonisation by pathogens. Broader studies or a wider range of practical experiences would be needed to confirm and quantify this observation.
When sanitation treatments cannot be avoided, it might be useful to restore the microbiome by either biopriming or coating the seeds with beneficial micro-organisms originating from seed microbiomes. Methods, in particular, when and how to apply biologicals to seeds, still need to be fine-tuned to ensure effectiveness. Specifically, it needs to be clear to what extent applied biologicals are remanent in soils and vertically transmitted to the next plant generation. Biologicals that become remanent in the environment or are vertically transmitted, create the risk of interfering with and altering endemic microbial communities and of consequences for biosecurity [73]. From a legal perspective, formulated products based on micro-organisms require a formal registration. On the one hand, a stringent registration process may ensure effectiveness, safety, and biosecurity of such products. On the other hand, costs, and administrative burdens due to the registration process are the main reason why there is as yet only a rather limited set of biologicals for seed coating on the market.
Outside the scope of registration requirements for commercial, formulated products, seed applications of artisanal and local preparations—such as compost teas, effective micro-organisms, biodynamic preparations, or the Latin-American “organismos de montaña”—are being experimented in farms and gardens worldwide [79,80,81]. These preparations often involve endemic micro-organisms originating from local plants, soils or animals and have mainly been developed to be applied to crops and soils. Seed applications are still marginal. Nevertheless, benefits of such preparations for seed production and for seed and seedling health would be worth investigating using scientific methods.
- Legal framework: Plant Health regulations and pathogen thresholds
At the level of the European Union (EU), seed health is regulated by Plant Health [82] rules and a set of seed marketing directives/regulations (e.g., [3,4]). National laws may further specify these measures. The rules also define thresholds at which certain seed-borne diseases are tolerated, analytical methods by which the respective pathogens should be quantified and applicable measures for control or eradication. There is scientific evidence for greater microbial life in organic soils, compared to conventionally managed soils, as mentioned in previous sections. Both empirical observations from farming practice and scientific findings [83,84] indicate that diversified organic systems may have more buffering capacity against plant pathogens. A beneficial seed microbiome may support resilience against seed-borne pathogens in organic systems and further strengthen that buffering capacity. This implies that organic systems could potentially deal with higher thresholds for certain seedborne plant diseases.
Adapting the threshold standards for organic seeds would in some cases reduce the cost for seed production or increase the availability of organic seeds for crops that are presently difficult to produce organically. For example, in Europe, requirements for the control of common bunt on certified seed of wheat and other cereals are set at the national level and differ between countries. Discussions among specialists at an online workshop on national common bunt requirements, concluded that bunt thresholds on certified seeds could be adapted to the seed sanitation treatments available for organic seed in a given country (Proceedings of the Workshop on European Bunt requirements, Satellite to the XXI International Workshop on Bunt and Smut Diseases 2021, in preparation). Such adapted thresholds ensure that healthy seed is marketed, and that appropriate seed sanitation is applied, when necessary, without constituting unnecessary market barriers for organic seed.
Pathogen thresholds are based on the level of risk that is considered acceptable. Defining thresholds is a delicate task, as certain seed borne diseases can cause high amounts of economic damage under conditions favourable to them, for example by Clavibacter michiganensis subsp. michiganensis (Cmm) that causes bacterial wilt and canker of tomato. Disease control is mainly focussed on prevention to avoid seed infection, as seeds constitute the main source for disease introduction to new areas. A worldwide collaboration between seed companies and seed testing organisations has established a so-called ‘Good Seed and Plant Practices’ chain system (GSPP), with protocols to prevent tomato seed and plant lots from being infected by the Cmm pathogen [85]. On the other hand, small-scale, diversified organic seed companies have criticised this GSPP certification system (personal communications) for being inaccessible to small-scale companies, for economic and technical reasons (in particular, the requirement of totally covered, separated production areas with netted ventilation and, sometimes, disinfected water). Moreover, by systematically applying strict isolation measures to tomato seed and transplant production, this system is not appropriate for producers who aim at resilient, low-input cropping systems, basing plant health on ecosystem and biological interactions (e.g., principles of “Health” and “Ecology” formulated by IFOAM the International Federation of Organic Agriculture Movements [86]).
Regarding another regulated bacterial disease, common bacterial blight with beans (Xanthomonas axonopodis pv. phaseoli), some small-scale artisanal seed companies also criticised EU legal requirements for bean seeds as a disproportionate market barrier, as they were subject to the same sampling and analysis obligations as multi-national companies growing and marketing seed all over the globe [87]. These critiques can be interpreted as a call for requirements adapted to small-scale, artisanal seed production enterprises and local seed networks, based on the observation that they are important drivers of crop diversity and represent a limited risk of spreading diseases [88,89]. Extensive research on the resilience of organic, small-scale, and diversified seed systems is needed to provide the necessary background knowledge to respond to these requests, while limiting the risk of spreading seed-borne diseases.
4. Strategy to Improve Seed Health from a Dynamic Perspective: Next Steps
For the aim of leveraging the potential of this broadened perspective on seed and plant health for the benefit of organic seed production, the above-mentioned implications and open questions need to be addressed and integrated, from local on farm seed production to entire seed systems. The following strategy is a set of recommendations for future research and development, in plant breeding and seed production.
A more comprehensive understanding is needed of the factors affecting the composition and transmission of seed microbiota, as well of the functions and effects of microbial taxa associated to seeds. This will also require integrating ecological and reductionist approaches to microbiome research [90]. In time, this may give indications on how seed production, harvesting, processing and storage can favour the composition of optimal seed microbiota. It would also allow to draw conclusions on how to compensate for damaged seed microbiota after sanitation treatments and to know when microbially diverse organic soils can buffer negative effects of seed disinfection. Seed treatments by inoculation with biologicals could be made more effective and risks for biosecurity assessed, in particular by utilizing endemic seed microbial diversity and taking risks of disturbing endemic microbial communities into account.
Further insights into the geographic distribution of seed microbial communities and their role in the local adaptation and resilience of crops are necessary to understand the contribution of local seed systems and community seed banks in the preservation of endemic seed microbes. More generally, these insights would also provide information about the implications of (globalized) seed movements. Accordingly, gene banks could develop appropriate conservation strategies, considering the microbial communities associated to crop seeds.
Exploring how seed production conditions and treatments affect seed microbiota, seed vigour and crop epigenetics may allow to formulate recommendations for seed production from a holistic perspective, considering eventual trade-offs. Furthermore, understanding the role of crop and overall biological diversity in interaction with seed microbiota, vigour and epigenetics would enable breeding programmes to incorporate these aspects. This understanding may also shed new light on the potential of local seed systems and community seed banks. Overarching approaches to plant breeding and seed production may be required, where plant breeding targets specific environments and seed production unravels the full (epi-)genetic and microbial potential of crop plants. Encompassing all these factors may be overwhelmingly multi-factorial, strongly context-dependant and might pose some analytical challenges. Transdisciplinary approaches will help tackle those challenges, by combining scientific findings with the know-how and locally embedded knowledge of organic seed producers.
A first, practical step for breeding programs is to develop methods to screen and select for plants that favour high seed vigour. At the stage of variety evaluation, when breeders or farmers’ groups compare different selections or cultivars to identify the ones best adapted and performing in their conditions, it is also useful to account for the vigour of the seed used for trials. Differences in performance may sometimes not be explained by differences between cultivars, but by differences in initial seed vigour, due to diverging seed production or seed storage conditions. Further, the role of the seed and seedling microbiome should be considered, as potential sources of resilience, in the frame of holobiont breeding.
For seed production, educating seed producers, seed companies and farmers on the role of the seed microbiota and seed vigour would be worthwhile, with a particular focus on seed maturation, optimal harvest time and conditions, seed drying and storage. Here, too, simple and affordable indicators or tests would be useful. With the aimed increase of organically managed agricultural land and growing incitements to use organic seed, more organic seed producers and companies will be needed in future. Proper training and support will aid them to produce high-quality seed.
In general, more emphasis could be placed on producing and maintaining high seed vigour to further improve stress resilience of seedlings. Integrating seed vigour in seed quality assessment would be a first step, although this is more labour-intensive than simple germination testing. In future, the role of the seed microbiome in seed quality could also be integrated.
Overall, taking the proposed multi-factorial and dynamic perspective on seed health leads to a shift in how we view and care for seed and plant health. From a stance of plant pathology, focussing mainly on plant pathogens and disease, we may evolve to a stance better described as plant salutology—the science of plant health—where the focus lies on the processes sustaining health. Regulations as presently in place for seed quality, plant health and the application of biologicals may severely restrict the implementation of a seed microbiome strategy and the associated shift in perspective. Research and public discourse [6] are therefore needed to create a legal framework for steering the seed and plant health, in particular seed and plant microbiomes. Concerning the reintroduction of seed microbiome components through inoculation, benefits and risks need to be assessed. Inoculation with microbiota isolated from the crop itself or wild relatives with a regional perspective may be a way to benefit from seed microbiomes without standardising specific microbial communities at a global level.
Plant viruses, archaea, protists and nematodes are not mentioned in this strategy. Indeed, research on plant and seed associated microbiomes have mainly focussed on bacteria and fungi. The role and—sometimes beneficial—effects of viruses in plants have hardly been elucidated [91]. There is a knowledge gap concerning how viruses interact with the fungal and bacterial fractions of the microbiome. The fungal [92] and bacterial [93,94] taxa may induce resistance of plants to viral diseases. Viruses can also affect the overall microbiome [95]. Elucidating these interactions would allow integrating viral components of the microbiome and viral diseases in a more comprehensive strategy.
Key recommended actions towards the strategy are summarized in Table 1.
5. Conclusions
Plants and micro-organisms have co-evolved into symbiotic relationships. Modern agriculture based on synthetic pesticides and mineral fertilisers has unintentionally disrupted this relation to a large extend. Organic farming and breeding can greatly benefit from taking this relation into account to (re)create resilient cropping systems. It is likely that the use of synthetic pesticides and mineral fertilisers has also limited the selection for and benefits of high seed vigour, as lacking vigour in seedlings was often compensated by these inputs. Seed vigour and the seed microbiome need to be included in a strategy to provide organic farmers with higher quality propagation material, that can produce seedlings with a higher resilience against biotic and abiotic stresses. On the one hand, the contributions of healthy, vigorous seeds are expected to be of particular importance for crop health, weed suppression and overall crop performance in organic cropping systems, where “quick fixes” to compensate for an unsatisfactory start to a crop are scarce. On the other hand, organic farming holds a particular potential to develop these aspects of seed health and quality, especially regarding microbial life associated to seeds. Organic farming seeks inspiration from the resilience of natural ecosystems. Additionally, in this sense, we expect that seed vigour and microbiomes will receive increasing attention in the future.
Author Contributions
Conceptualization, S.M.K., F.R. and S.P.C.G.; methodology, S.M.K. and S.P.C.G.; validation, S.M.K. and S.P.C.G.; writing—original draft preparation, S.M.K. and S.P.C.G.; writing—review and editing, S.M.K., F.R. and S.P.C.G.; visualization, S.M.K. and S.P.C.G.; funding acquisition, F.R. and S.P.C.G. All authors have read and agreed to the published version of the manuscript.
Funding
This article is an outcome of the LIVESEED project, funded by the European Union’s Horizon 2020 research and innovation programme under grant agreement No 727230 and by the Swiss State Secretariat for Education, Research and Innovation (SERI) under contract number 17.00090. The contribution from Wageningen Research has also been funded by the Dutch Topsector Horticulture & Propagation material (grant TU EU-2017-05).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors thank the seed producers, seed companies, researchers and organisations that have accepted to share their knowledge and experiences. Their feedback and food for thought on the seed health strategy have been important contributions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Storkey, J. Modelling Seedling Growth Rates of 18 Temperate Arable Weed Species as a Function of the Environment and Plant Traits. Ann. Bot. 2004, 93, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, J. Barley Seed Vigour and Mechanical Weed Control. Weed Res. 2008, 40, 219–230. [Google Scholar] [CrossRef]
- European Council. Council Directive 2002/55/EC on the Marketing of Vegetable Seed; European Council: Brussels, Belgium, 2002. [Google Scholar]
- European Council. Council Directive 66/402/EEC of 14 June 1966 on the Marketing of Cereal Seed; European Council: Brussels, Belgium, 1966. [Google Scholar]
- European Parliament and Council. Regulation (EU) 2018/848 of the European Parliament and of the Council of 30 May 2018 on Organic Production and Labelling of Organic Products and Repealing Council Regulation (EC) No 834/2007; European Parliament and Council: Brussels, Belgium, 2018. [Google Scholar]
- Döring, T.F.; Pautasso, M.; Finckh, M.R.; Wolfe, M.S. Concepts of Plant Health—Reviewing and Challenging the Foundations of Plant Protection. Plant Pathol. 2012, 61, 1–15. [Google Scholar] [CrossRef]
- Klaedtke, S.; Mélard, F.; Chable, V.; Stassart, P.M. Les artisans semenciers, les haricots et leurs agents pathogènes. La biodiversité cultivée et la santé des plantes au cœur d’une identité professionnelle. Etudes Rural. 2018, 202, 36–55. [Google Scholar] [CrossRef]
- Altieri, M.A. Agroecology: The Science of Sustainable Agriculture, 2nd ed.; Westview Press: Boulder, CO, USA; IT Publications: London, UK, 1995; ISBN 978-0-8133-1717-5. [Google Scholar]
- Stassart, P.M.; Baret, P.V.; Grégoire, J.-C.; Hance, T.; Mormont, M.; Reheul, D.; Stilmant, D.; Vanloqueren, G. Visser L’agroécologie: Trajectoire et potentiel—Pour une transition vers des systèmes alimentaires durables. In Agroécologie Entre Pratiques et Sciences Sociales; Educagri éditions: Dijon, France, 2012; ISBN 978-2-84444-876-7. [Google Scholar]
- ISTA. International Rules for Seed Testing. Full Issue Engl. Fr. 2022, 2022, i-19–8(8). [Google Scholar] [CrossRef]
- Song, C.; Zhu, F.; Carrión, V.J.; Cordovez, V. Beyond Plant Microbiome Composition: Exploiting Microbial Functions and Plant Traits via Integrated Approaches. Front. Bioeng. Biotechnol. 2020, 8, 896. [Google Scholar] [CrossRef]
- Vandenkoornhuyse, P.; Quaiser, A.; Duhamel, M.; Le Van, A.; Dufresne, A. The Importance of the Microbiome of the Plant Holobiont. New Phytol. 2015, 206, 1196–1206. [Google Scholar] [CrossRef]
- Shade, A.; Jacques, M.-A.; Barret, M. Ecological Patterns of Seed Microbiome Diversity, Transmission, and Assembly. Curr. Opin. Microbiol. 2017, 37, 15–22. [Google Scholar] [CrossRef]
- Vayssier-Taussat, M.; Albina, E.; Citti, C.; Cosson, J.-F.; Jacques, M.-A.; Lebrun, M.-H.; Le Loir, Y.; Ogliastro, M.; Petit, M.-A.; Roumagnac, P.; et al. Shifting the Paradigm from Pathogens to Pathobiome: New Concepts in the Light of Meta-Omics. Front. Cell. Infect. Microbiol. 2014, 4, 29. [Google Scholar] [CrossRef]
- Wassermann, B.; Cernava, T.; Müller, H.; Berg, C.; Berg, G. Seeds of Native Alpine Plants Host Unique Microbial Communities Embedded in Cross-Kingdom Networks. Microbiome 2019, 7, 108. [Google Scholar] [CrossRef] [Green Version]
- LIVESEED Project. Boosting Organic Seed and Plant Breeding across Europe, European Union Horizon 2020 Research and Innovation Programme Grant Agreement No 727230; Swiss State Secretariat for Education, Research and Innovation Contract Number 17.00090. 2017. Available online: https://www.liveseed.eu/ (accessed on 30 June 2022).
- Groot, S.P.C.; Klaedtke, S.; Messmer, M.; Rey, F. Organic Seed Health. An Inventory of Issues and a Report on Case Studies. LIVESEED Project. 2020. Available online: https://orgprints.org/id/eprint/39423 (accessed on 29 August 2022).
- Stevens, R.B. Cultural Practices in Disease Control. In Plant Pathology, an Advanced Treatise; Horsfall, J.G., Dimond, A.E., Eds.; Academic Press: New York, NY, USA, 1960; Volume 3, pp. 357–429. [Google Scholar]
- Francl, L.J. The Disease Triangle: A Plant Pathological Paradigm Revisited. Plant Health Instr. 2001. [Google Scholar] [CrossRef]
- Browning, J.A.; Simons, M.D.; Torres, E. Managing Host Genes: Epidemiologic and Genetic Concepts. In How Disease is Managed; Elsevier: Amsterdam, The Netherlands, 1977; pp. 191–212. ISBN 978-0-12-356401-6. [Google Scholar]
- Agrios, G.N. Plant Pathology, 5th ed.; Elsevier Academic Press: Amsterdam, The Netherlands, 2005; ISBN 978-0-12-044565-3. [Google Scholar]
- Andrade-Piedra, J.L.; Cáceres, P.A.; Forbes, G.A.; Pumisacho, M. Humans: The Neglected Corner of the Disease Tetrahedron—Developing a Training Guide for Resource-Poor Farmers to Control Potato Late Blight. Acta Hortic. 2009, 401, 111–122. [Google Scholar] [CrossRef]
- Hickey, C.; Brannen, P.M.; Blaauw, B.R. Pierce’s Disease of Grape: Identification and Management. UGA Coop. Ext. Bull 2019, 1514, 1–6. Available online: https://extension.uga.edu/publications/detail.html?number=B1514 (accessed on 30 August 2022).
- Finch-Savage, W.E.; Bassel, G.W. Seed Vigour and Crop Establishment: Extending Performance beyond Adaptation. J. Exp. Bot. 2016, 67, 567–591. [Google Scholar] [CrossRef] [PubMed]
- Yamane, K.; Garcia, R.; Imayoshi, K.; Mabesa-Telosa, R.C.; Banayo, N.P.M.C.; Vergara, G.; Yamauchi, A.; Sta Cruz, P.; Kato, Y. Seed Vigour Contributes to Yield Improvement in Dry Direct-Seeded Rainfed Lowland Rice: Seed Vigour under Drought in Rice. Ann. Appl. Biol. 2018, 172, 100–110. [Google Scholar] [CrossRef]
- Bewley, J.D.; Bradford, K.J.; Hilhorst, H.W.M.; Nonogaki, H. Seeds: Physiology of Development, Germination and Dormancy, 3rd ed.; Springer: New York, NY, USA, 2013; ISBN 978-1-4614-4692-7. [Google Scholar]
- Rajjou, L.; Debeaujon, I. Seed Longevity: Survival and Maintenance of High Germination Ability of Dry Seeds. Comptes Rendus. Biol. 2008, 331, 796–805. [Google Scholar] [CrossRef]
- Groot, S.P.C.; Birnbaum, Y.; Rop, N.; Jalink, H.; Forsberg, G.; Kromphardt, C.; Werner, S.; Koch, E. Effect of Seed Maturity on Sensitivity of Seeds towards Physical Sanitation Treatments. Seed Sci. Technol. 2006, 34, 403–413. [Google Scholar] [CrossRef]
- Finch-Savage, W.E.; Clay, H.A.; Lynn, J.R.; Morris, K. Towards a Genetic Understanding of Seed Vigour in Small-Seeded Crops Using Natural Variation in Brassica oleracea. Plant Sci. 2010, 179, 582–589. [Google Scholar] [CrossRef]
- Li, C.S.; Shao, G.S.; Wang, L.; Wang, Z.F.; Mao, Y.J.; Wang, X.Q.; Zhang, X.H.; Liu, S.T.; Zhang, H.S. QTL Identification and Fine Mapping for Seed Storability in Rice (Oryza sativa L.). Euphytica 2017, 213, 127. [Google Scholar] [CrossRef]
- Lee, J.-S.; Velasco-Punzalan, M.; Pacleb, M.; Valdez, R.; Kretzschmar, T.; McNally, K.L.; Ismail, A.M.; Cruz, P.C.S.; Sackville Hamilton, N.R.; Hay, F.R. Variation in Seed Longevity among Diverse Indica Rice Varieties. Ann. Bot. 2019, 124, 447–460. [Google Scholar] [CrossRef]
- Mohamed-Yasseen, Y.; Barringer, S.A.; Splittstoesser, W.E.; Costanza, S. The Role of Seed Coats in Seed Viability. Bot. Rev. 1994, 60, 426–439. [Google Scholar] [CrossRef]
- Das Gupta, P.R.; Austenson, H.M. Analysis of Interrelationships among Seedling Vigor, Field Emergence, and Yield in Wheat. Agron. J. 1973, 65, 417–422. [Google Scholar] [CrossRef]
- Magnée, K.J.H.; Scholten, O.E.; Postma, J.; Lammerts van Bueren, E.T.; Groot, S.P.C. Sensitivity of Spinach Seed Germination to Moisture Is Driven by Oxygen Availability and Influenced by Seed Size and Pericarp. Seed Sci. Technol. 2020, 48, 117–131. [Google Scholar] [CrossRef]
- Magnée, K.J.H. From Seed to Seedling—Damping-off Tolerance in Spinacia oleracea L. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 2022. [Google Scholar]
- White, J.F.; Kingsley, K.L.; Butterworth, S.; Brindisi, L.; Gatei, J.W.; Elmore, M.T.; Verma, S.K.; Yao, X.; Kowalski, K.P. Seed-Vectored Microbes: Their Roles in Improving Seedling Fitness and Competitor Plant Suppression. In Seed Endophytes; Verma, S.K., White, J.F., Jr., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 3–20. ISBN 978-3-030-10503-7. [Google Scholar]
- Nelson, E.B. The Seed Microbiome: Origins, Interactions, and Impacts. Plant Soil 2018, 422, 7–34. [Google Scholar] [CrossRef]
- Matsumoto, H.; Fan, X.; Wang, Y.; Kusstatscher, P.; Duan, J.; Wu, S.; Chen, S.; Qiao, K.; Wang, Y.; Ma, B.; et al. Bacterial Seed Endophyte Shapes Disease Resistance in Rice. Nat. Plants 2021, 7, 60–72. [Google Scholar] [CrossRef]
- Irizarry, I.; White, J.F. Application of Bacteria from Non-Cultivated Plants to Promote Growth, Alter Root Architecture and Alleviate Salt Stress of Cotton. J. Appl. Microbiol. 2017, 122, 1110–1120. [Google Scholar] [CrossRef] [PubMed]
- Morella, N.M.; Zhang, X.; Koskella, B. Tomato Seed-Associated Bacteria Confer Protection of Seedlings Against Foliar Disease Caused by Pseudomonas syringae. Phytobiomes J. 2019, 3, 177–190. [Google Scholar] [CrossRef]
- Hone, H.; Mann, R.; Yang, G.; Kaur, J.; Tannenbaum, I.; Li, T.; Spangenberg, G.; Sawbridge, T. Profiling, Isolation and Characterisation of Beneficial Microbes from the Seed Microbiomes of Drought Tolerant Wheat. Sci. Rep. 2021, 11, 11916. [Google Scholar] [CrossRef] [PubMed]
- Mitter, B.; Pfaffenbichler, N.; Flavell, R.; Compant, S.; Antonielli, L.; Petric, A.; Berninger, T.; Naveed, M.; Sheibani-Tezerji, R.; von Maltzahn, G.; et al. A New Approach to Modify Plant Microbiomes and Traits by Introducing Beneficial Bacteria at Flowering into Progeny Seeds. Front. Microbiol. 2017, 8, 11. [Google Scholar] [CrossRef]
- Song, Y.; Haney, C.H. Drought Dampens Microbiome Development. Nat. Plants 2021, 7, 994–995. [Google Scholar] [CrossRef]
- Rezki, S.; Campion, C.; Iacomi-Vasilescu, B.; Preveaux, A.; Toualbia, Y.; Bonneau, S.; Briand, M.; Laurent, E.; Hunault, G.; Simoneau, P.; et al. Differences in Stability of Seed-Associated Microbial Assemblages in Response to Invasion by Phytopathogenic Microorganisms. PeerJ 2016, 4, e1923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalaf, E.M.; Raizada, M.N. Bacterial Seed Endophytes of Domesticated Cucurbits Antagonize Fungal and Oomycete Pathogens Including Powdery Mildew. Front. Microbiol. 2018, 9, 42. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Zhang, R.; Wu, X.; Xu, T.; Ahmad, S.; Zhang, X.; Zhao, J.; Liu, Y. An Endophytic Strain of the Genus Bacillus Isolated from the Seeds of Maize (Zea mays L.) Has Antagonistic Activity against Maize Pathogenic Strains. Microb. Pathog. 2020, 142, 104074. [Google Scholar] [CrossRef]
- Bergna, A.; Cernava, T.; Rändler, M.; Grosch, R.; Zachow, C.; Berg, G. Tomato Seeds Preferably Transmit Plant Beneficial Endophytes. Phytobiomes J. 2018, 2, 183–193. [Google Scholar] [CrossRef]
- Johnston-Monje, D.; Gutiérrez, J.P.; Lopez-Lavalle, L.A.B. Seed-Transmitted Bacteria and Fungi Dominate Juvenile Plant Microbiomes. Front. Microbiol. 2021, 12, 737616. [Google Scholar] [CrossRef] [PubMed]
- Maeder, P.; Fliessbach, A.; Dubois, D.; Gunst, L.; Fried, P.; Niggli, U. Soil Fertility and Biodiversity in Organic Farming. Science 2002, 296, 1694–1697. [Google Scholar] [CrossRef] [PubMed]
- Tahat, M.M.; Alananbeh, K.M.; Othman, Y.A.; Leskovar, D.I. Soil Health and Sustainable Agriculture. Sustainability 2020, 12, 4859. [Google Scholar] [CrossRef]
- Moeskops, B.; Sukristiyonubowo; Buchan, D.; Sleutel, S.; Herawaty, L.; Husen, E.; Saraswati, R.; Setyorini, D.; De Neve, S. Soil Microbial Communities and Activities under Intensive Organic and Conventional Vegetable Farming in West Java, Indonesia. Appl. Soil Ecol. 2010, 45, 112–120. [Google Scholar] [CrossRef]
- Wei, Z.; Jousset, A. Plant Breeding Goes Microbial. Trends Plant Sci. 2017, 22, 555–558. [Google Scholar] [CrossRef]
- Vujanovic, V.; Germida, J. Seed Endosymbiosis: A Vital Relationship in Providing Prenatal Care to Plants. Can. J. Plant Sci. 2017, 97, 972–981. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; van Themaat, E.V.L.; Schulze-Lefert, P. Structure and Functions of the Bacterial Microbiota of Plants. Annu. Rev. Plant Biol. 2013, 64, 807–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Romero, E.; Aguirre-Noyola, J.L.; Taco-Taype, N.; Martínez-Romero, J.; Zuñiga-Dávila, D. Plant Microbiota Modified by Plant Domestication. Syst. Appl. Microbiol. 2020, 43, 126106. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Jin, K.; Raaijmakers, J.M. Designing a Home for Beneficial Plant Microbiomes. Curr. Opin. Plant Biol. 2021, 62, 102025. [Google Scholar] [CrossRef] [PubMed]
- Tosi, M.; Mitter, E.K.; Gaiero, J.; Dunfield, K. It Takes Three to Tango: The Importance of Microbes, Host Plant, and Soil Management to Elucidate Manipulation Strategies for the Plant Microbiome. Can. J. Microbiol. 2020, 66, 413–433. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, W.; Jaradat, A.A.; Hurburgh, C.; Pollak, L.M.; Goodman, M. Breeding Maize under Biodynamic-Organic Conditions for Nutritional Value and N Efficiency/N2 Fixation. Open Agric. 2019, 4, 322–345. [Google Scholar] [CrossRef]
- Abdelrazek, S.; Simon, P.; Colley, M.; Mengiste, T.; Hoagland, L. Crop Management System and Carrot Genotype Affect Endophyte Composition and Alternaria dauci Suppression. PLoS ONE 2020, 15, e0233783. [Google Scholar] [CrossRef] [PubMed]
- Gopal, M.; Gupta, A. Microbiome Selection Could Spur Next-Generation Plant Breeding Strategies. Front. Microbiol. 2016, 7, 1971. [Google Scholar] [CrossRef]
- Rybakova, D.; Mancinelli, R.; Wikström, M.; Birch-Jensen, A.-S.; Postma, J.; Ehlers, R.-U.; Goertz, S.; Berg, G. The Structure of the Brassica napus Seed Microbiome Is Cultivar-Dependent and Affects the Interactions of Symbionts and Pathogens. Microbiome 2017, 5, 104. [Google Scholar] [CrossRef]
- Mayer, J.; Gunst, L.; Mäder, P.; Samson, M.-F.; Carcea, M.; Narducci, V.; Thomsen, I.K.; Dubois, D. Productivity, Quality and Sustainability of Winter Wheat under Long-Term Conventional and Organic Management in Switzerland. Eur. J. Agron. 2015, 65, 27–39. [Google Scholar] [CrossRef]
- Wen, D.; Hou, H.; Meng, A.; Meng, J.; Xie, L.; Zhang, C. Rapid Evaluation of Seed Vigor by the Absolute Content of Protein in Seed within the Same Crop. Sci. Rep. 2018, 8, 5569. [Google Scholar] [CrossRef]
- Li, W.; He, X.; Chen, Y.; Jing, Y.; Shen, C.; Yang, J.; Teng, W.; Zhao, X.; Hu, W.; Hu, M.; et al. A Wheat Transcription Factor Positively Sets Seed Vigour by Regulating the Grain Nitrate Signal. New Phytol. 2020, 225, 1667–1680. [Google Scholar] [CrossRef]
- Lammerts van Bueren, E.T.; Østergård, H.; Goldringer, I.; Scholten, O. Plant Breeding for Organic and Sustainable, Low-Input Agriculture: Dealing with Genotype–Environment Interactions. Euphytica 2008, 163, 321–322. [Google Scholar] [CrossRef]
- Roberts, M.R.; López Sánchez, A. Plant Epigenetic Mechanisms in Response to Biotic Stress. In Epigenetics in Plants of Agronomic Importance: Fundamentals and Applications; Alvarez-Venegas, R., De-la-Peña, C., Casas-Mollano, J.A., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 65–113. ISBN 978-3-030-14759-4. [Google Scholar]
- Zhu, Q.-H.; Shan, W.-X.; Ayliffe, M.A.; Wang, M.-B. Epigenetic Mechanisms: An Emerging Player in Plant-Microbe Interactions. Mol. Plant-Microbe Interact. 2016, 29, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Zhi, P.; Chang, C. Exploiting Epigenetic Variations for Crop Disease Resistance Improvement. Front. Plant Sci. 2021, 12, 692328. [Google Scholar] [CrossRef]
- Dasgupta, P.R.; Austenson, H.M. Relations between Estimates of Seed Vigor and Field Performance in Wheat. Can. J. Plant Sci. 1973, 53, 43–46. [Google Scholar] [CrossRef]
- Ceccarelli, S.; Grando, S. Decentralized-Participatory Plant Breeding: An Example of Demand Driven Research. Euphytica 2007, 155, 349–360. [Google Scholar] [CrossRef]
- Noël, R. Etude de Schémas de Sélection Paysanne. Oral Presentation at the Final Meeting of the Project “Covalience” on 6 June 2021. Available online: https://maison-de-la-semence-paysanne-dordogne.netlify.app/post/mds-gc/publication/reponse_selection/#presentation-et-resultats (accessed on 30 August 2022).
- Klaedtke, S.; Jacques, M.-A.; Raggi, L.; Préveaux, A.; Bonneau, S.; Negri, V.; Chable, V.; Barret, M. Terroir Is a Key Driver of Seed-Associated Microbial Assemblages: Terroir Shapes the Seed Microbiota. Environ. Microbiol. 2016, 18, 1792–1804. [Google Scholar] [CrossRef]
- Berg, G.; Raaijmakers, J.M. Saving Seed Microbiomes. ISME J. 2018, 12, 1167–1170. [Google Scholar] [CrossRef]
- Brush, S.B. Genes in the Field: On-Farm Conservation of Crop Diversity; International Plant Genetic Resources Institute: Rome, Italy ; International Development Research Centre: Ottawa, ON, Canada; Lewis Publishers: Boca Raton, FL, USA, 2000; ISBN 978-0-88936-884-2. [Google Scholar]
- Andersen, R.; Shrestha, P.; Otieno, G.; Nishikawa, Y.; Kasasa, P.; Mushita, A. Community Seed Banks: Sharing Experiences from North and South. Report from a Side Event Held 1 November 2017, during the Seventh Session of the Governing Body of the International Treaty on Plant Genetic Resources for Food and Agriculture; Kigali, Rwanda. 2017. Available online: https://cgspace.cgiar.org/bitstream/handle/10568/92510/Community_Andersen_2018.pdf?sequence=1&isAllowed=y (accessed on 6 June 2022).
- Vernooy, R.; Sthapit, B.; Otieno, G.; Shrestha, P.; Gupta, A. The Roles of Community Seed Banks in Climate Change Adaption. Dev. Pract. 2017, 27, 316–327. [Google Scholar] [CrossRef]
- Chandel, A.; Mann, R.; Kaur, J.; Norton, S.; Edwards, J.; Spangenberg, G.; Sawbridge, T. Implications of Seed Vault Storage Strategies for Conservation of Seed Bacterial Microbiomes. Front. Microbiol. 2021, 12, 784796. [Google Scholar] [CrossRef]
- van der Wolf, J.M.; Birnbaum, Y.; van der Zouwen, P.S.; Groot, S.P.C. Disinfection of Vegetable Seed by Treatment with Essential Oils, Organic Acids and Plant Extracts. Seed Sci. Technol. 2008, 36, 76–88. [Google Scholar] [CrossRef]
- Scheuerell, S.; Mahaffee, W. Compost Tea: Principles and Prospects For Plant Disease Control. Compost Sci. Util. 2002, 10, 313–338. [Google Scholar] [CrossRef]
- Vaish, S.; Garg, N.; Ahmad, I.Z. Microbial Basis of Organic Farming Systems with Special Reference to Biodynamic Preparations. Indian J. Agric. Sci. 2020, 90, 1219–1225. [Google Scholar]
- Acosta Almanzar, H.A. Microorganismos Eficientes de Montaña: Evaluación de su Potencial Bajo Manejo Agroecológico de Tomate en Costa Rica. Master′s Thesis, Centro Agronómico Tropical de Investigación y Enseñanza, Turrialba, Costa Rica, 2012. [Google Scholar]
- European Parliament and Council. Regulation (EU) 2016/2031 of the European Parliament and of the Council of 26 October 2016 on Protective Measures against Pests of Plants, Amending Regulations (EU) No 228/2013, (EU) No 652/2014 and (EU) No 1143/2014 of the European Parliament and of the Council and Repealing Council Directives 69/464/EEC, 74/647/EEC, 93/85/EEC, 98/57/EC, 2000/29/EC, 2006/91/EC and 2007/33/EC; European Parliament and Council: Brussels, Belgium, 2016. [Google Scholar]
- van Bruggen, A.H.C.; Goss, E.M.; Havelaar, A.; van Diepeningen, A.D.; Finckh, M.R.; Morris, J.G. One Health—Cycling of Diverse Microbial Communities as a Connecting Force for Soil, Plant, Animal, Human and Ecosystem Health. Sci. Total Environ. 2019, 664, 927–937. [Google Scholar] [CrossRef]
- van Bruggen, A.H.C.; Finckh, M.R. Plant Diseases and Management Approaches in Organic Farming Systems. Annu. Rev. Phytopathol. 2016, 54, 25–54. [Google Scholar] [CrossRef]
- Good Seed and Plant Practices. Available online: https://gspp.eu/ (accessed on 6 June 2022).
- IFOAM Organics International Principles of Organic Agriculture. 2005. Available online: https://www.ifoam.bio/principles-organic-agriculture-brochure (accessed on 6 June 2022).
- Klaedtke, S. Governance of Plant Health and Management of Crop Diversity—The Case of Common Bean Health Management among Members of the Association Croqueurs de Carottes; Université de Liège: Liège, Belgium; Agrocampus Ouest: Rennes, France, 2017. [Google Scholar]
- Dachverband Kulturpflanzen—Und Nutztiervielfalt e.V., Board. Evaluation of Article 79 (Plant Passport) of the Plant Health Regulation 2016/2031/EU. 2021. Available online: https://kulturpflanzen-nutztiervielfalt.org/sites/kulturpflanzen-nutztiervielfalt.org/files/Dachverband_Plant_Health_Art79.pdf (accessed on 3 June 2022).
- Meißner, U. Gut zu wissen: Neues im Pflanzengesundheitsrecht. Specie Rara Polit. Int. 2022. Available online: https://www.prospecierara.de/news/news-detail/news/gut-zu-wissen-neues-im-pflanzengesundheitsrecht.html (accessed on 3 June 2022).
- Fitzpatrick, C.R.; Salas-González, I.; Conway, J.M.; Finkel, O.M.; Gilbert, S.; Russ, D.; Teixeira, P.J.P.L.; Dangl, J.L. The Plant Microbiome: From Ecology to Reductionism and Beyond. Annu. Rev. Microbiol. 2020, 74, 81–100. [Google Scholar] [CrossRef]
- Roossinck, M.J. A New Look at Plant Viruses and Their Potential Beneficial Roles in Crops: Beneficial Viruses for Crops. Mol. Plant Pathol. 2015, 16, 331–333. [Google Scholar] [CrossRef]
- Elsharkawy, M.M.; Shimizu, M.; Takahashi, H.; Ozaki, K.; Hyakumachi, M. Induction of Systemic Resistance against Cucumber Mosaic Virus in Arabidopsis thaliana by Trichoderma asperellum SKT-1. Plant Pathol. J. 2013, 29, 193–200. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Behiry, S.I.; Al-Askar, A.A. Bacillus velezensis PEA1 Inhibits Fusarium oxysporum Growth and Induces Systemic Resistance to Cucumber Mosaic Virus. Agronomy 2020, 10, 1312. [Google Scholar] [CrossRef]
- Abo-Zaid, G.A.; Matar, S.M.; Abdelkhalek, A. Induction of Plant Resistance against Tobacco Mosaic Virus Using the Biocontrol Agent Streptomyces cellulosae Isolate Actino 48. Agronomy 2020, 10, 1620. [Google Scholar] [CrossRef]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant–Microbiome Interactions: From Community Assembly to Plant Health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Seed and plant health can be seen as a continuum, based on the iterative process of seed production, involving interactions between various factors affecting germination and early plant growth, general crop health, harvest conditions and seed processing and storage. These factors affect the seed microbiota and seed vigour, thereby affecting germination and seedling health.
Figure 1.
Seed and plant health can be seen as a continuum, based on the iterative process of seed production, involving interactions between various factors affecting germination and early plant growth, general crop health, harvest conditions and seed processing and storage. These factors affect the seed microbiota and seed vigour, thereby affecting germination and seedling health.

Figure 2.
Sensitivity of germinating carrot seeds to the fungal pathogen Alternaria radicina. The vigour from the control seeds (in the left) was declined by storing them four days at 40 °C and 85% relative humidity, resulting in seeds with reduced vigour (right). The seeds were sown on filter paper with either addition of water or a suspension of fungal spores diluted in water. Seedling were imaged seven days after sowing.
Figure 2.
Sensitivity of germinating carrot seeds to the fungal pathogen Alternaria radicina. The vigour from the control seeds (in the left) was declined by storing them four days at 40 °C and 85% relative humidity, resulting in seeds with reduced vigour (right). The seeds were sown on filter paper with either addition of water or a suspension of fungal spores diluted in water. Seedling were imaged seven days after sowing.


{kind=link}
{kind=link}
Table 1.
Key actions for the implementation of the proposed organic seed health strategy.
| Actions | Aimed Outcome |
|---|---|
| Higher organic seed vigour. |
| Knowledge on how to favour optimised seed microbiota; recommendations for an effective preservation and conservation of endemic seed microbiota. |
| Knowledge on essential consortia of seed microbiota for resilient seedlings and crops; knowledge on the role of microbial consortia in the local adaptation of crops. |
| Reintroduction of natural seed microbiota into depleted environments, in particular after seed disinfection. |
| Recommendations and methodologies for organic breeding programmes which preserve and make the most of biological diversity at all levels, including crops, cropping systems and microbial ecology. |
| More precision when comparing the performance of different plant cultivars by accounting for differences in initial seed vigour; improved organic seed quality and seedling resilience. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Klaedtke, S.M.; Rey, F.; Groot, S.P.C. Designing a Seed Health Strategy for Organic Cropping Systems, Based on a Dynamic Perspective on Seed and Plant Health. Sustainability 2022, 14, 10903. https://doi.org/10.3390/su141710903
AMA Style
Klaedtke SM, Rey F, Groot SPC. Designing a Seed Health Strategy for Organic Cropping Systems, Based on a Dynamic Perspective on Seed and Plant Health. Sustainability. 2022; 14(17):10903. https://doi.org/10.3390/su141710903
Chicago/Turabian StyleKlaedtke, Stephanie M., Frédéric Rey, and Steven P. C. Groot. 2022. "Designing a Seed Health Strategy for Organic Cropping Systems, Based on a Dynamic Perspective on Seed and Plant Health" Sustainability 14, no. 17: 10903. https://doi.org/10.3390/su141710903
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.