
Stand Structure and Local Landscape Variables Are the Dominant Factors Explaining Shrub and Tree Diversity in Mediterranean Forests

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
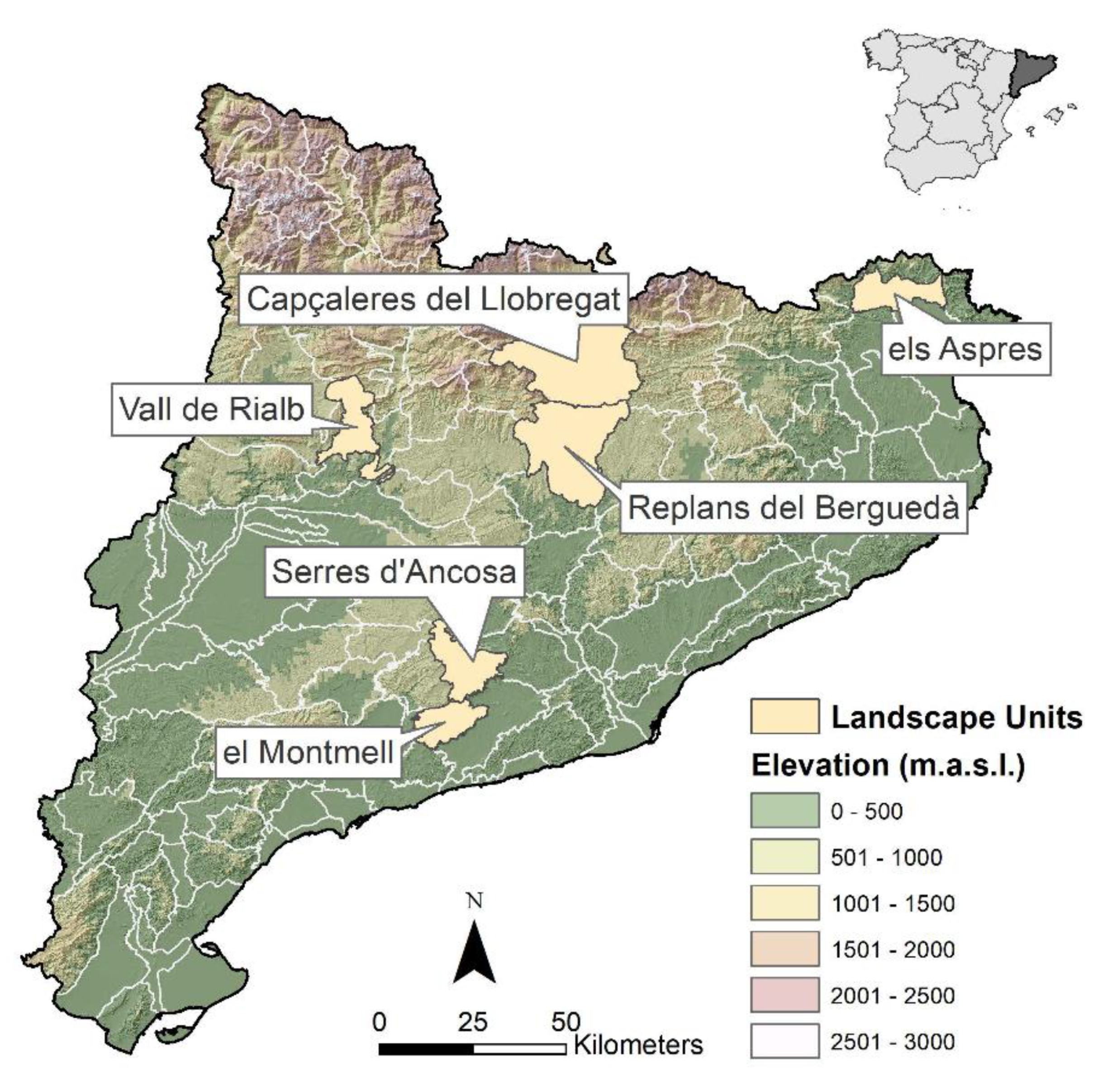
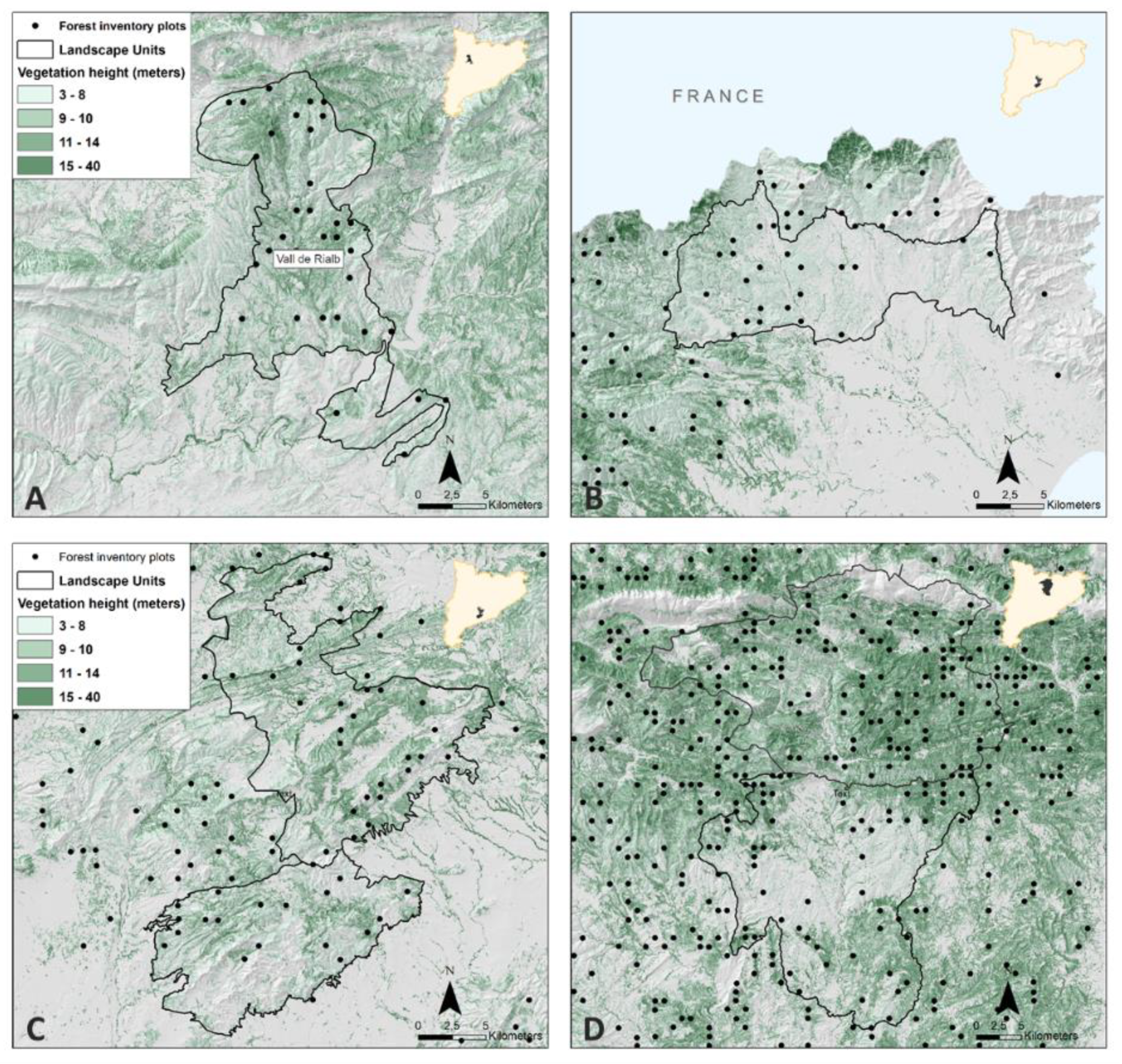
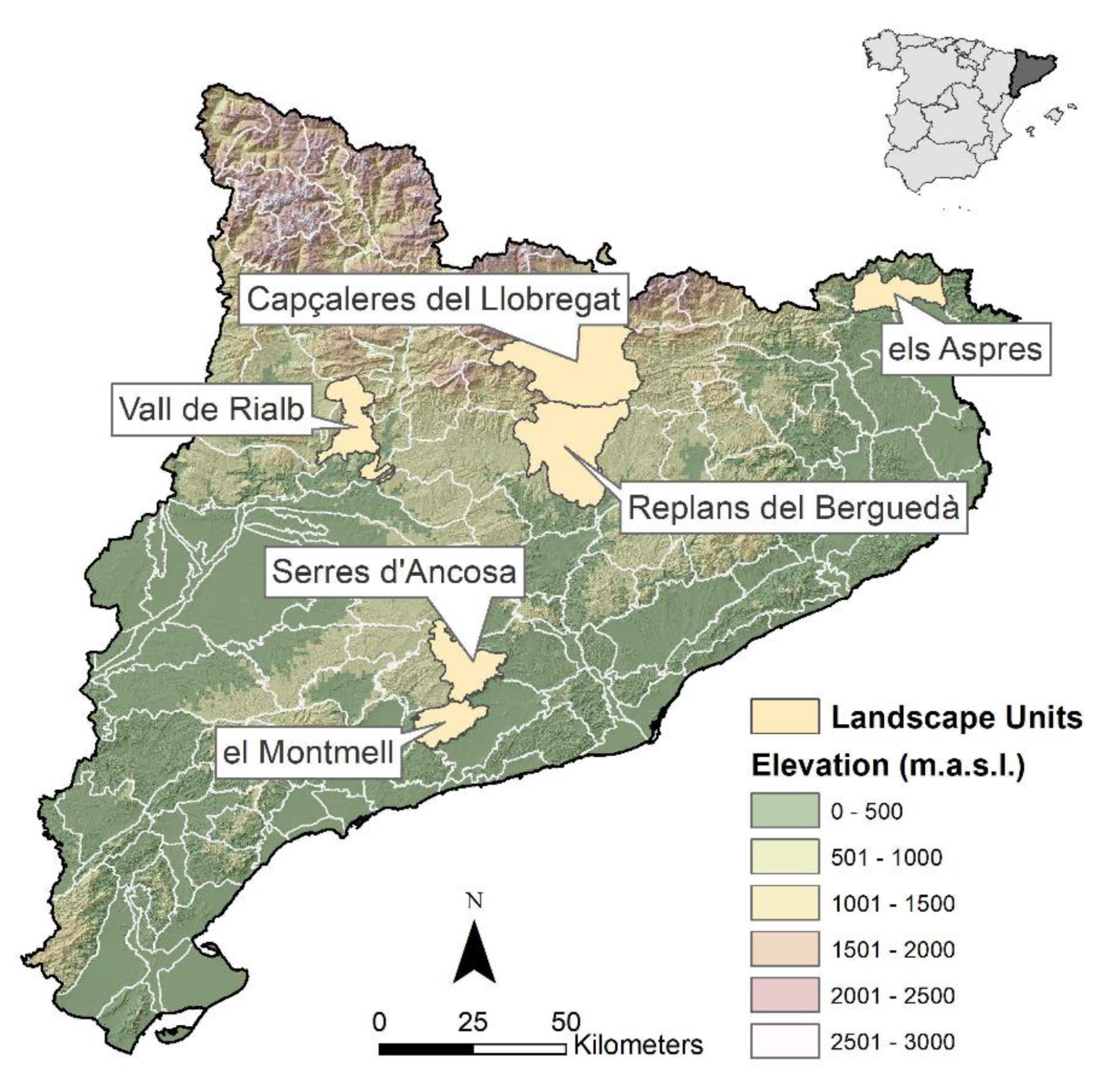
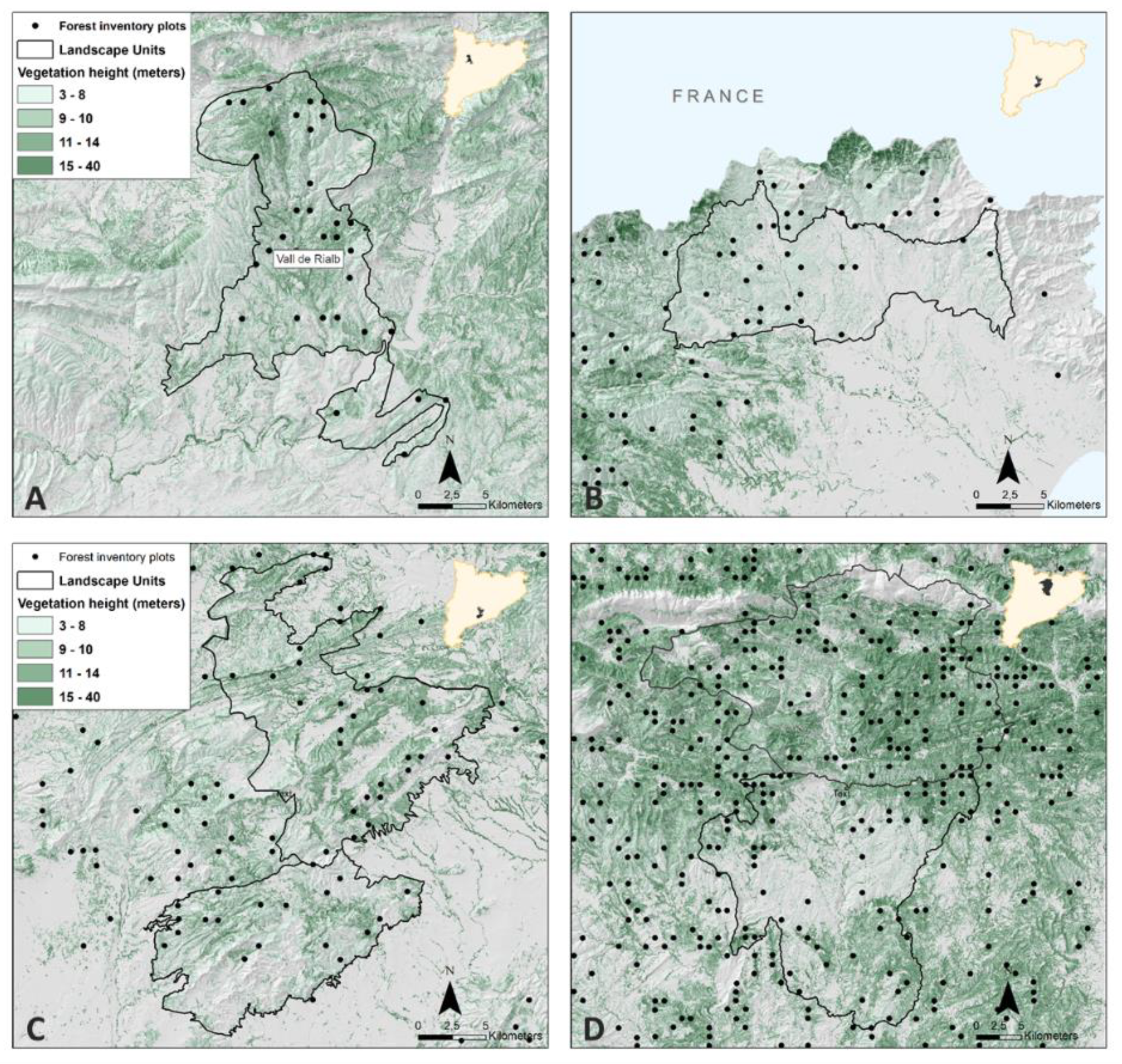
2.1. Study Area and Data Source
2.2. Stand-Level Diversity and Structure Variables
2.3. Local Landscape Variables
2.4. Model Building with Artificial Neural Networks
3. Results
4. Discussion
4.1. Structural and Landscape Predictors of Tree and Shrub Diversity
4.2. Structural and Landscape Target Characteristics for Tree and Shrub Diversity in Specific Mediterranean Forest Types
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


References
- Pitkanen, S. Correlation between stand structure and ground vegetation: An analytical approach. Plant Ecol. 1997, 131, 109–126. [Google Scholar] [CrossRef]
- Pereira, H.M.; Leadley, P.W.; Proença, V.; Alkemade, R.; Scharlemann, J.; Fernandez-Manjarrés, J.F.; Araújo, M.B.; Balvanera, P.; Biggs, R.; Cheung, W.W.L.; et al. Scenarios for Global Biodiversity in the 21st Century. Science 2010, 330, 1496–1501. [Google Scholar] [CrossRef] [Green Version]
- Fady-Welterlen, B. Is there really more biodiversity in Mediterranean forest ecosystems? Taxon 2005, 54, 905–910. [Google Scholar] [CrossRef]
- Mittermeier, R.A.; Turner, W.R.; Larsen, F.W.; Brooks, T.M.; Gascon, C. Global Biodiversity Conservation: The Critical Role of Hotspots. In Biodiversity Hotspots; Zachos, F., Habel, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar] [CrossRef]
- Bellard, C.; Leclerc, C.; Leroy, B.; Bakkenes, M.; Veloz, S.; Thuiller, W.; Courchamp, F. Biodiversity hotspots and global change. Glob. Ecol. Biogeogr. 2014, 23, 1376–1386. [Google Scholar] [CrossRef]
- Beilin, R.; Lindborg, R.; Stenseke, M.; Pereira, H.M.; Llausàs, A.; Slätmo, E.; Cerqueira, Y.; Navarro, L.; Rodrigues, P.; Reichelt, N.; et al. Analysing How Drivers of Agricultural Land Abandonment Affect Biodiversity and Cultural Landscapes Using Case Studies from Scandinavia, Iberia and Oceania. Land Use Policy 2014, 36, 60–72. [Google Scholar] [CrossRef]
- MedECC. Climate and Environmental Change in the Mediterranean Basin–Current Situation and Risks for the Future; First Mediterranean Assessment Report; Cramer, W., Guiot, J., Marini, K., Eds.; Union for the Mediterranean, Plan Bleu, UNEP/MAP: Marseille, France, 2020; 600p, in press. [Google Scholar]
- Ameztegui, A.; Brotons, L.; Coll, L. Land-Use Changes as Major Drivers of Mountain Pine (Pinus uncinata Ram.) Expansion in the Pyrenees. Glob. Ecol. Biogeogr. 2010, 19, 632–641. [Google Scholar] [CrossRef] [Green Version]
- Cervera, T.; Pino, J.; Marull, J.; Padró, R.; Tello, E. Understanding the Long-Term Dynamics of Forest Transition: From Deforestation to Afforestation in a Mediterranean Landscape (Catalonia, 1868–2005). Land Use Policy 2019, 80, 318–331. [Google Scholar] [CrossRef]
- Poyatos, R.; Latron, J.; Llorens, P. Land Use and Land Cover Change after Agricultural Abandonment: The Case of a Mediterranean Mountain Area (Catalan Pre-Pyrenees). Mt. Res. Dev. 2003, 23, 362–368. [Google Scholar] [CrossRef] [Green Version]
- Amici, V.; Santi, E.; Filibeck, G.; Diekmann, M.; Geri, F.; Landi, S.; Scoppola, A.; Chiarucci, A. Influence of secondary forest succession on plant diversity patterns in a Mediterranean landscape. J. Biogeogr. 2013, 40, 2335–2347. [Google Scholar] [CrossRef]
- Vega-Garcia, C.; Chuvieco, E. Applying local measures of spatial heterogeneity to Landsat-TM images for predicting wildfire occurrence in Mediterranean landscapes. Landsc. Ecol. 2006, 21, 595–605. [Google Scholar] [CrossRef]
- Fabbio, G.; Merlo, M.; Tosi, V. Silvicultural management in maintaining biodiversity and resistance of forests in Europe—The Mediterranean region. J. Environ. Manag. 2003, 67, 67–76. [Google Scholar] [CrossRef]
- Sluiter, R.; de Jong, S.M. Spatial Patterns of Mediterranean Land Abandonment and Related Land Cover Transitions. Landsc. Ecol. 2007, 22, 559–576. [Google Scholar] [CrossRef]
- Otero, I.; Marull, J.; Tello, E.; Diana, G.L.; Pons, M.; Coll, F.; Boada, M. Land abandonment, landscape, and biodiversity: Questioning the restorative character of the forest transition in the Mediterranean. Ecol. Soc. 2015, 20, 7. [Google Scholar] [CrossRef] [Green Version]
- Van Leeuwen, C.E.; Cammeraat, E.L.H.; de Vente, J.; Boix-Fayos, C. The evolution of soil conservation policies targeting land abandonment and soil erosion in Spain: A review. Land Use Policy 2019, 83, 174–186. [Google Scholar] [CrossRef]
- Liang, J.; Crowther, T.W.; Picard, N.; Wiser, S.; Zhou, M.; Alberti, G.; Schulze, E.-D.; McGuire, A.D.; Bozzato, F.; Pretzsch, H.; et al. Positive biodiversity–productivity relationship predominant in global forests. Science 2016, 354, aaf8957. [Google Scholar] [CrossRef] [Green Version]
- Morin, X.; Fahse, L.; Mazancourt, C.; Scherer-Lorenzen, M.; Bugmann, H. Temporal stability in forest productivity increases with tree diversity due to asynchrony in species dynamics. Ecol. Lett. 2014, 17, 1526–1535. [Google Scholar] [CrossRef]
- Schnabel, F.; Schwarz, J.A.; Dănescu, A.; Fichtner, A.; Nock, C.A.; Bauhus, J.; Potvin, C. Drivers of productivity and its temporal stability in a tropical tree diversity experiment. Glob. Chang. Biol. 2019, 25, 4257–4272. [Google Scholar] [CrossRef]
- Jactel, H.; Bauhus, J.; Boberg, J.; Bonal, D.; Castagneyrol, B.; Gardiner, B.; Gonzalez-Olabarria, J.R.; Koricheva, J.; Meurisse, N.; Brockerhoff, E.G. Tree Diversity Drives Forest Stand Resistance to Natural Disturbances. Curr. For. Rep. 2017, 3, 223–243. [Google Scholar] [CrossRef]
- Sánchez-Pinillos, M.; Coll, L.; Cáceres, M.D.; Ameztegui, A. Assessing the persistence capacity of communities facing natural disturbances on the basis of species response traits. Ecol. Indic. 2016, 66, 76–85. [Google Scholar] [CrossRef] [Green Version]
- Coll, L.; Ameztegui, A.; Collet, C.; Löf, M.; Mason, B.; Pach, M.; Verheyen, K.; Abrudan, I.; Barbati, A.; Barreiro, S.; et al. Knowledge gaps about mixed forests: What do European forest managers want to know and what answers can science provide? For. Ecol. Manag. 2018, 407, 106–115. [Google Scholar] [CrossRef] [Green Version]
- Torras, O.; Saura, S. Effects of silvicultural treatments on forest biodiversity indicators in the Mediterranean. For. Ecol. Manag. 2008, 255, 3322–3330. [Google Scholar] [CrossRef]
- Ferris, R.; Peace, A.J.; Humphrey, J.W.; Broome, A.C. Relationships between vegetation, site type and stand structure in coniferous plantations in Britain. For. Ecol. Manag. 2000, 136, 35–51. [Google Scholar] [CrossRef]
- Lowman, M.D.; Nadkarni, N.M. Forest Canopies; Academic Press: San Diego, CA, USA, 1995; ISBN 0-12-457650-8. [Google Scholar]
- Hashemi, S.A. Evaluating plant species diversity and physiographical factors in natural broad leaf forest. Am. J. Environ. Sci. 2010, 6, 20–25. [Google Scholar] [CrossRef]
- Hashemi, S.A. Biodiversity Indices of natural Hornbeam stands in relation to stand volume in management area. Middle East J. Sci. Res. 2011, 10, 64–69. [Google Scholar]
- Rago, M.M.; Urretavizcaya, M.F.; Lederer, N.S.; Defossé, G.E. Plant Community Response to Forest Fuel Management in Patagonian Pine Plantations. Front. For. Glob. Chang. 2020, 3, 55. [Google Scholar] [CrossRef]
- Nieppola, J.J.; Carleton, T.J. Relations between Understory Vegetation, Site Productivity, and Environmental Factors in Pinus Sylvestris L. Stands in Southern Finland. Vegetatio 1991, 93, 57–72. [Google Scholar]
- Guitton, J.L.; Ruchaud, F. Conséquences Écologiques de L’éclaircie des Peuplements de réSineux (Ecological Consequences of Thinning in Coniferous Plantations). Recherch Eisidore Website. 1996. Available online: http://www.rechercheisidore.fr/search/resource/?uri=10670/1.sjmcpq (accessed on 10 May 2021).
- Rameau, J.C. Aménagement Forestier. Importance de L’écologie, Prise en Compte de la Biodiversité. 1999. Available online: http://documents.irevues.inist.fr/bitstream/handle/2042/5507/87_101.pdf?sequence=1 (accessed on 10 May 2021).
- Van Pelt, R.; Franklin, J. Influence of canopy structure on the understory environment in tall, old growth, conifer forests. Can. J. For. Res. 2000, 30, 1231–1245. [Google Scholar] [CrossRef]
- Shater, Z. Studying the Effect of Stand Characteristics on Plant Species diversity in the Median Part of Natural Pinus brutia Ten. (Brutia pine) Forests in Al-Bayer and Al-Bassit Regions-Syria. Tishreen Univ. J. Res. Sci. Stud. Biol. Sci. Ser. 2013, 35, 72–84. [Google Scholar]
- Bohn, F.J.; May, F.; Huth, A. Species composition and forest structure explain the temperature sensitivity patterns of productivity in temperate forests. Biogeosciences 2018, 15, 1795–1813. [Google Scholar] [CrossRef] [Green Version]
- Wei, L.; Archaux, F.; Hulin, F.; Bilger, I.; Gosselin, F. Stand attributes or soil micro-environment exert greater influence than management type on understory plant diversity in even-aged oak high forests. For. Ecol. Manag. 2020, 460, 117897. [Google Scholar] [CrossRef]
- Amici, V.; Rocchini, D.; Filibeck, G.; Bacaro, G.; Santi, E.; Geri, F.; Landi, S.; Scoppola, A.; Chiarucci, A. Landscape structure effects on forest plant diversity at local scale: Exploring the role of spatial extent. Ecol. Complex. 2015, 21, 44–55. [Google Scholar] [CrossRef]
- Malavasi, M.; Bartak, V.; Carranza, M.L.; Simova, P.; Acosta, A.T.R. Landscape pattern and plant biodiversity in Mediterranean coastal dune ecosystems: Do habitat loss and fragmentation really matter? J. Biogeogr. 2018, 45, 1367–1377. [Google Scholar] [CrossRef]
- Robert, K.; Jones, D.O.B.; Huvenne, V.A.I. Megafaunal distribution and biodiversity in a heterogeneous landscape: The iceberg-scoured Rockall Bank, NE Atlantic. Mar. Ecol. Prog. Ser. 2014, 501, 67–88. [Google Scholar] [CrossRef] [Green Version]
- Katayama, N.; Amano, T.; Naoe, S.; Yamakita, T.; Komatsu, I.; Takagawa, S.; Sato, N.; Ueta, M.; Miyashita, T. Landscape Heterogeneity–Biodiversity Relationship: Effect of Range Size. PLoS ONE 2014, 9, e93359. [Google Scholar] [CrossRef]
- Turner, M.G. Landscape ecology: What is the state of the science? Annu. Rev. Ecol. Evol. Syst. 2005, 36, 319–344. [Google Scholar] [CrossRef]
- Austin, M.P. Spatial prediction of species distribution: An interface between ecological theory and statistical modeling. Ecol. Model. 2002, 157, 101–118. [Google Scholar] [CrossRef] [Green Version]
- McElhinny, C.H. Forest and Woodland Structure as an Index of Biodiversity: A Review. A Literature Review Commissioned by NSW NPWS; Department of Forestry, Australian National University: Canberra, Australia, 2002. [Google Scholar]
- Curzon, M.T.; D’Amato, A.W.; Fraver, S.; Palik, B.J.; Bottero, A.; Foster, J.R.; Gleason, K.E. Harvesting influences functional identity and diversity over time in forests of the northeastern U.S.A. For. Ecol. Manag. 2017, 400, 93–99. [Google Scholar] [CrossRef]
- Vidal, C.; Alberdi, I.; Redmond, J.; Vestman, M.; Lanz, A.; Schadauer, K. The role of European National Forest Inventories for international forestry reporting. Ann. For. Sci. 2016, 73, 793–806. [Google Scholar] [CrossRef] [Green Version]
- Corona, P.; Chirici, G.; McRoberts, R.E.; Winter, S.; Barbati, A. Contribution of large-scale forest inventories to biodiversity assessment and monitoring. For. Ecol. Manag. 2011, 262, 2061–2069. [Google Scholar] [CrossRef] [Green Version]
- Alberdi, I.; Vallejo, R.; Álvarez-González, J.G.; Condés, S.; González-Ferreiro, E.; Guerrero, S.; Hernández, L.; Martínez-Jauregui, M.; Montes, F.; Oliveira, N.; et al. The multi-objective Spanish National Forest Inventory. For. Syst. 2017, 26, e04S. [Google Scholar] [CrossRef]
- Larsson, T.-B.; Angelstam, P.; Balent, G.; Barbati, A.; Bijlsma, R.J.; Boncina, A.; Bradshaw, R.; Bücking, W.; Ciancio, O.; Corona, P.; et al. Biodiversity Evaluation Tools for European Forests; Blackwell Science: Oxford, UK, 2001; Volume 50, p. 237. [Google Scholar]
- Barrett, T.M.; Gray, A.N. Potential of a national monitoring program for forests to assess change in high-latitude ecosystems. Biol. Conserv. 2011, 144, 1285–1294. [Google Scholar] [CrossRef]
- Alberdi, I.; Sandoval, V.; Condés, S.; Cañellas, I.; Vallejo, R. El Inventario Forestal Nacional español, una herramienta para el conocimiento, la gestión y la conservación de los ecosistemas forestales arbolados. Ecosistemas 2016, 25, 88–97. [Google Scholar] [CrossRef]
- Alberdi, I.; Cañellas, I.; Condes, S. A long-scale biodiversity monitoring methodology for Spanish national forest inventory. Application to Álava region. For. Syst. 2014, 23, 93–110. [Google Scholar] [CrossRef]
- Saura, S.; Martín-Queller, E.; Hunter, M.L. Forest landscape change and biodiversity conservation. In Forest landscapes and Global Change; Springer: New York, NY, USA, 2014; pp. 167–198. [Google Scholar]
- Chiarucci, A.; Bacaro, G.; Filibeck, G.; Landi, S.; Maccherini, S.; Scoppola, A. Scale dependence of plant species richness in a network of protected areas. Biodivers. Conserv. 2012, 21, 503–516. [Google Scholar] [CrossRef]
- Torras, O.; Martín-Queller, E.; Saura, S. Relating landscape structure, environment and management to biodiversity indicators estimated from forest inventory data in Catalonia (NE Spain). Investig. ÓN Agrar. Sist. Y Recur. For. 2009, 18, 322–337. [Google Scholar] [CrossRef] [Green Version]
- Dauber, J.; Hirsch, M.; Simmering, D.; Waldhardt, R.; Otte, A.; Wolters, V. Landscape structure as an indicator of biodiversity: Matrix effects on species richness. Agric. Ecosyst. Environ. 2003, 98, 321–329. [Google Scholar] [CrossRef]
- Spellerberg, I.F.; Fedor, P.J. A tribute to Claude Shannon (1916–2001) and a plea for more rigorous use of species richness, species diversity and the ‘Shannon–Wiener’Index. Glob. Ecol. Biogeogr. 2003, 12, 177–179. [Google Scholar] [CrossRef] [Green Version]
- Yeboah, D.; Chen, H.Y.H.; Kingston, S. Tree species richness decreases while species evenness increases with disturbance frequency in a natural boreal forest landscape. Ecol. Evol. 2016, 6, 842–850. [Google Scholar] [CrossRef]
- Magurran, A.E. Diversidad ecológica y su Medición; Vedrà: Barcelona, Spain, 1989. [Google Scholar]
- Margalef, R.; Daget, J. Les modèles mathématiques en écologie. Masson, Paris. 172 p. 90F. Limnol. Oceanogr. 1978, 23, 575–576. [Google Scholar] [CrossRef]
- Vericat, P.; Piqué, M.; Koua, O.; Pla, M. Mapa de Formacions Forestals Pures i Mixtes de Catalunya a partir del Mapa Forestal de España 1:50.000 digitalitzat; Centre de Ciència i Tecnologia Forestal de Catalunya: Solsona, Spain, 2010. [Google Scholar]
- Piqué, M.; Vericat, P.; Cervera, T.; Baiges, T.; Farriol, R. Tipologies Forestals Arbrades. Sèrie: Orientacions de Gestió Forestal Sostenible per a Catalunya (ORGEST); Centre de la Propietat Forestal, Departament d’Agricultura, Ramaderia, Pesca, Alimentació i Medi Natural, Generalitat de Catalunya: Barcelona, Spain, 2011. [Google Scholar]
- McGarigal, K.; Cushman, S.A.; Ene, E. FRAGSTATS v4: Spatial Pattern Analysis Program for Categorical and Continuous Maps. Computer Software Program Produced by the Authors at the University of Massachusetts, Amherst. 2012. Available online: http://www.umass.edu/landeco/research/fragstats/fragstats.html (accessed on 18 October 2021).
- Hasenauer, H.; Merkl, D.; Weingartner, M. Estimating tree mortality of Norway spruce stands with neural networks. Adv. Environ. Res. 2001, 5, 405–414. [Google Scholar] [CrossRef]
- Soares da Rocha, S.J.S.; Eleto Torres, C.M.M.; Jacovine, G.; Leite, H.; Gelcer, E.; Neves, K.; Schettini, B.; Villanova, P.H.; Silva, L.; Reis, L.; et al. Artificial neural networks: Modeling tree survival and mortality in the Atlantic Forest biome in Brazil. Sci. Total Environ. 2018, 645, 655–661. [Google Scholar] [CrossRef]
- Özçelik, R.; Diamantopoulou, M.J.; Brooks, J.R.; Wiant, H.V. Estimating tree bole volume using artificial neural network models for four species in Turkey. J. Environ. Manag. 2010, 91, 742–753. [Google Scholar] [CrossRef]
- Correia Vieira, G.; Ribeiro de Mendonça, A.; Fernandes da Silva, G.; Zanetti, S.S.; Marques da Silva, M.; Rosa dos Santos, A. Prognoses of diameter and height of trees of eucalyptus using artificial intelligence. Sci. Total Environ. 2018, 619–620, 1473–1481. [Google Scholar] [CrossRef]
- Vega-Garcia, C.; Lee, B.S.; Woodard, P.M.; Titus, S.J. Applying neural network technology to human-caused wildfire occurrence prediction. AI Appl. 1996, 10, 9–18. [Google Scholar]
- Bisquert, M.; Caselles, E.; Sánchez, J.M.; Caselles, V. Application of artificial neural networks and logistic regression to the prediction of forest fire danger in Galicia using MODIS data. Int. J. Wildland Fire 2012, 21, 1025–1029. [Google Scholar] [CrossRef]
- NeuralWare, 2001–2014. NeuralSIM, the Complete Solution for Neural Data Modelling. In User Guide; NeuralWare: Pittsburgh, PA, USA, 2001–2014.
- Fahlman, S.E.; Lebiere, C. The cascade-correlation learning architecture. In Advances in Neural Information Processing Systems; Morgan Kaufmann Publishers Inc.: San Francisco, CA, USA, 1990; Volume 2, pp. 524–532. [Google Scholar]
- Alcázar, J.; Palau, A.; Vega-Garcia, C. A neural net model for environmental flow estimation at the Ebro River Basin, Spain. J. Hydrol. 2008, 349, 44–55. [Google Scholar] [CrossRef]
- Vega-Garcia, C.; Decuyper, M.; Alcázar, J. Applying cascade-correlation neural networks to in-fill gaps in Mediterranean daily flow data series. Water 2019, 11, 1691. [Google Scholar] [CrossRef] [Green Version]
- Werbos, P.J. The Roots of Backpropagation: From Ordered Derivatives to Neural Networks and Political Forecasting; John Wiley & Sons, Inc.: New York, NY, USA, 1994; 319p. [Google Scholar]
- Torras, O.; Gil-Tena, A.; Saura, S. How does forest landscape structure explain tree species richness in a Mediterranean context? Biodivers. Conserv. 2008, 17, 1227–1240. [Google Scholar] [CrossRef]
- Özçelik, R.; Gul, A.U.; Merganic, J.; Merganicova, K.J. Tree species diversity and its relationship to stand parameters and geomorphology features in the eastern Black Sea region forests of Turkey. J. Environ. Biol. 2008, 29, 291–298. [Google Scholar]
- Neumann, M.; Starlinger, F. The significance of different indices for stand structure and diversity in forests. For. Ecol. Manag. 2001, 145, 91–106. [Google Scholar] [CrossRef]
- Zenner, E.K. Do residual trees increase structural complexity in Pacific Northwest coniferous forests? Ecol. Appl. 2000, 10, 800–810. [Google Scholar] [CrossRef]
- Shater, Z. Diversité Végétale et Sylviculture: Effet de la Plantation et de la Gestion D’espèces Forestières Introduites sur la Diversité Végétale. Etude du cas D’anciennes Châtaigneraies des Cévennes, Midi de la France. Ph.D. Thesis, Faculté des Sciences et Techniques de Saint Jérôme, Marseille, France, 2001. [Google Scholar]
- Blondel, J. The ‘Design’ of Mediterranean Landscapes: A Millennial Story of Humans and Ecological Systems during the Historic Period. Hum. Ecol. 2006, 34, 713–729. [Google Scholar] [CrossRef]
- Tiscar-Oliver, P.A. Patterns of shrub diversity and tree regeneration across topographic and stand-structural gradients in a Mediterranean forest. For. Syst. 2015, 24, 11. [Google Scholar] [CrossRef] [Green Version]
- Olarieta, J.R.; Bargués Tobella, A.; Rodríguez-Ochoa, R.; Antúnez, M. Soil control over the distribution of Mediterranean oak forests in the Montsec mountains (notheastern Spain). Geoderma 2017, 291, 11–20. [Google Scholar] [CrossRef]
- Pons, J.; Pausas, J.G. Oak regeneration in heterogeneous landscapes: The case of fragmented Quercus suber forests in the eastern Iberian Peninsula. For. Ecol. Manag. 2006, 231, 196–204. [Google Scholar] [CrossRef]
- Riegel, G.M.; Miller, R.F.; Krueger, W.C. Competition for resources between understory vegetation and overstory Pinus ponderosa in northeastern Oregon. Ecol. Appl. 1992, 2, 71–85. [Google Scholar] [CrossRef]
- Nilsson, M.C.; Wardle, D.A. Understory vegetation as a forest ecosystem driver: Evidence from the northern Swedish boreal forest. Front. Ecol. Environ. 2005, 3, 421–428. [Google Scholar] [CrossRef]
- Shater, Z.; Gondard, H.; Amorini, E.; Romane, F. Effects of afforestation by introduced species in the old sweet chestnut (Castanea sativa Miller) groves of Cevennes, southern France, on plant species diversity. Zb. Gozdarstva Lesar. 2002, 68, 149–169. [Google Scholar]
- Vilà, M.; Vayreda, J.; Comas, L.; Ibáñez, J.J.; Mata, T.; Obón, B. Species richness and wood production: A positive association in Mediterranean forests. Ecol. Lett. 2007, 10, 241–250. [Google Scholar] [CrossRef]
- Smee, D. Species with a large impact on community structure. Nat. Educ. Knowl. 2012, 3, 40. [Google Scholar]
- Coll, L.; González-Olabarría, J.R.; Mola-Yudego, B.; Pukkala, T.; Messier, C. Predicting understory maximum shrubs cover using altitude and overstory basal area in different Mediterranean forests. Eur. J. For. Res. 2011, 11, 55. [Google Scholar] [CrossRef] [Green Version]
- Bugalho, M.N.; Caldeira, M.C.; Pereira, J.S.; Aronson, J.; Pausas, J.G. Mediterranean cork oak savannas require human use to sustain biodiversity and ecosystem services. Front. Ecol. Environ. 2011, 9, 278–286. [Google Scholar] [CrossRef] [Green Version]
- Salis, M.; Arca, B.; Alcasena-Urdiroz, F.; Massaiu, A.; Bacciu, V.; Bosseur, F.; Caramelle, P.; Dettori, S.; De Oliveira, A.S.F.; Molina-Terren, D.; et al. Analyzing the recent dynamics of wildland fires in Quercus suber L. woodlands in Sardinia (Italy), Corsica (France) and Catalonia (Spain). Eur. J. For. Res. 2019, 138, 415–431. [Google Scholar] [CrossRef] [Green Version]
- Aronson, J.; Pereira, J.S.; Pausas, J.G. Cork Oak Woodlands on the Edge: Ecology, Adaptive Management, and Restoration; Island Press: Washington, DC, USA, 2009. [Google Scholar]

{kind=link}
{kind=link}
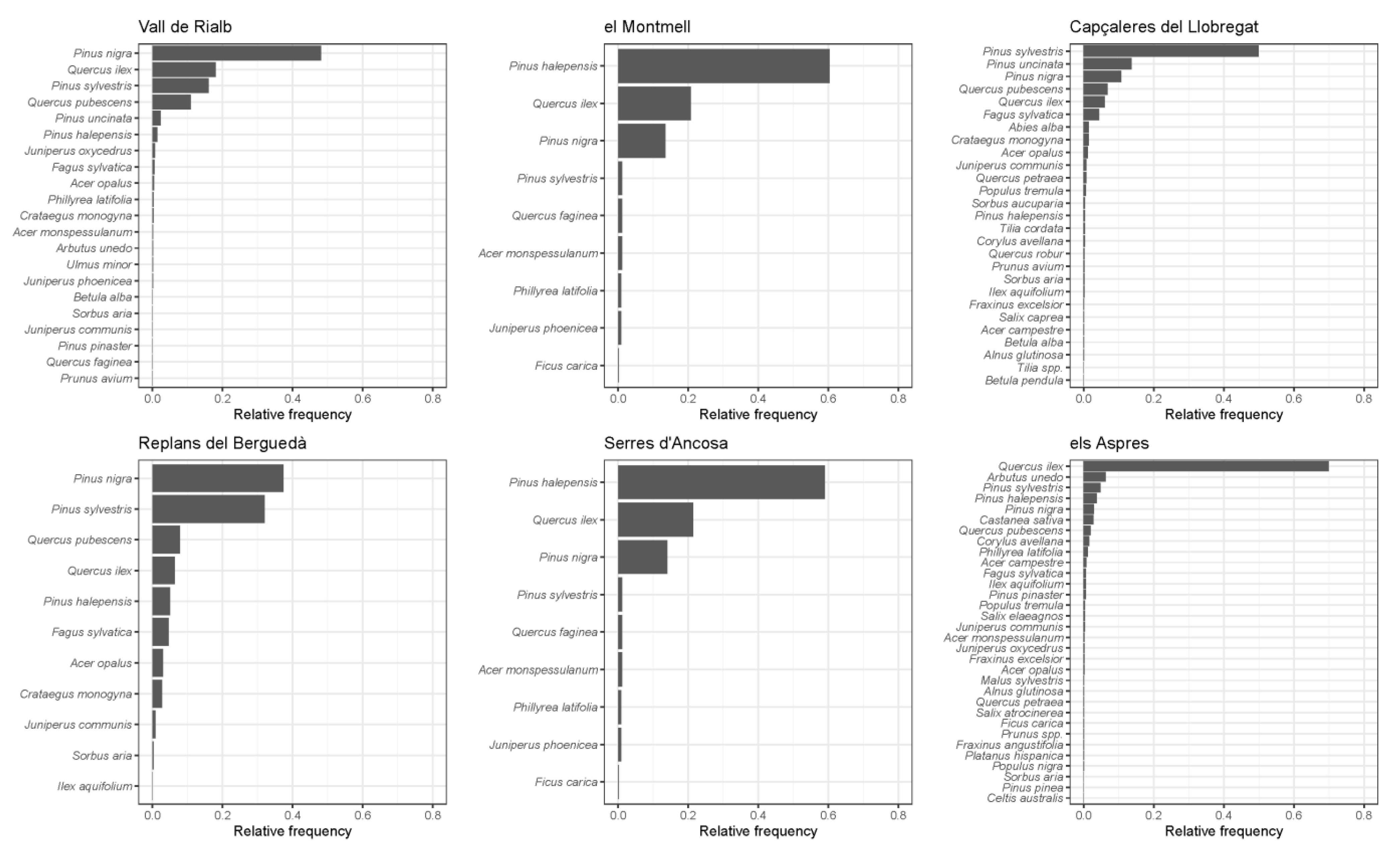{kind=link}
{kind=link}
| Abbreviation of Variable | Variable Description | Type of Variable |
|---|---|---|
| Dominant tree species data | ||
| Develop | Development status of the dominant species (seedling, sapling, thicket, pole, mature stand) | Categorical |
| Even/Uneven | Coetaneous, even-aged, mixed-aged, uneven-aged | Categorical |
| Origin | Natural, artificial, naturalized | Categorical |
| Shrubs data | ||
| Mean.HShrub | Mean shrub height | Interval |
| Std.HShrub | Standard deviation of shrub height | Interval |
| Trees data | ||
| D.Max | Maximum tree diameter | Interval |
| D.Min | Minimum tree diameter | Interval |
| Mean.D | Diameter mean | Interval |
| Std.D | Diameter standard deviation | Interval |
| Mean.HTrees | Mean tree canopy height | Interval |
| Std.HTrees | Standard deviation of tree canopy height | Interval |
| Cover and site data | ||
| Fcc.Tot | Fraction cover of total vegetation | Interval |
| Fcc.Trees | Fraction cover of tree canopy | Interval |
| Texture | Soil texture of the plot (1, sand; 2, mixed; 3, clay) | Categorical |
| MatOrg | Organic matter present in the plot soil (1, high humus content; 2, moderate humus; 3, low humus) | Categorical |
| pHSoil | pH level of the soil in the plot (1–14) | Interval |
| RegFelling | Evidence of regeneration fellings applied (dummy; no, 0; yes, 1) | Categorical |
| Class Metrics (Clas.) | Description | Landscape Metrics (Land.) | Description |
|---|---|---|---|
| CA | Total (Class) Area | TA | Total Area |
| PLAND | Percentage of Landscape | ||
| NP | Number of Patches | NP | Number of Patches |
| LPI | Largest Patch Index | LPI | Largest Patch Index |
| TE | Total Edge | TE | Total Edge |
| AREA_MN | Mean Patch Area | AREA_MN | Mean Patch Area |
| AREA_SD | Standard Deviation in Patch Area | AREA_SD | Standard Deviation in Patch Area |
| SHAPE_MN | Mean patch area | SHAPE_MN | Mean Patch Area |
| SHAPE_SD | Standard deviation in patch area | SHAPE_SD | Standard Deviation in Patch Area |
| ENN_MN | Euclidean Nearest-Neighbor Distance—mean | ENN_MN | Euclidean Nearest-Neighbor Distance—mean |
| ENN_SD | Euclidean Nearest-Neighbor Distance—standard deviation | ENN_SD | Euclidean Nearest-Neighbor Distance—standard deviation |
| CLUMPY | Clumpiness Index | CONTAG | Contagion |
| PLADJ | Percentage of Like Adjacencies | PLADJ | Percentage of Like Adjacencies |
| IJI | Interspersion and Juxtaposition Index | IJI | Interspersion and Juxtaposition Index |
| COHESION | Patch Cohesion Index | COHESION | Patch Cohesion Index |
| DIVISION | Landscape Division Index | DIVISION | Landscape Division Index |
| SPLIT | Splitting Index | SPLIT | Splitting Index |
| AI | Aggregation Index | AI | Aggregation Index |
| SHDI | Shannon Diversity Index | ||
| SHEI | Shannon Evenness Index |
| Variables | LU | Rich | ShDI | LU | Rich | ShDI | LU | Rich | ShDI | LU | Rich | ShDI | LU | Rich | ShDI | LU | Rich | ShDI | Total times in tree models |
| Models | Net Arch | 8–14–1 | 6–6–1 | Net Arch | 6–3–1 | 7–17–1 | Net Arch | 7–2–1 | 6–17–1 | Net Arch | 8–19–1 | 9–9–1 | Net Arch | 2–12–1 | 7−20−1 | Net Arch | 9−3−1 | 7–12–1 | |
| rTrain | 0.92 | 0.7 | rTrain | 0.8 | 0.88 | rTrain | 0.6 | 0.53 | rTrain | 0.58 | 0.4 | rTrain | 0.47 | 0.55 | rTrain | 0.6 | 0.56 | ||
| rTest | 0.75 | 0.7 | rTest | 0.9 | 0.89 | rTest | 0.7 | 0.55 | rTest | 0.65 | 0.6 | rTest | 0.48 | 0.71 | rTest | 0.6 | 0.56 | ||
| rValid | 0.86 | 0.6 | rValid | 0.8 | 0.85 | rValid | 0.6 | 0.54 | rValid | 0.60 | 0.5 | rValid | 0.47 | 0.58 | rValid | 0.60 | 0.55 | ||
| Develop | Serres d’Ancosa Mature P.halepensis | 1 | 1 | Montmell Young P. halepensis | Replans de BerguedàYoung and mature P. sylvestris | Capçaleres del LlobregatMature P. sylvestris | 1 | 1 | Aspres Young and adult Q. suber | −1 | Vall de Rialbmature P. nigra mixed with Q. faginea, Q ilex | 5 | |||||||
| Even/Uneven | −1 | 1 | 2 | ||||||||||||||||
| Mean.Hshrub | 2 | 1 | 3 | ||||||||||||||||
| Std.Hshrub | 1 | 1 | 1 | 2 | 1 | 6 | |||||||||||||
| D.Max | 1 | −1 | 1 | 1 | 2 | 1 | −1 | 8 | |||||||||||
| D.Min | −2 | −1 | −1 | −1 | 1 | 6 | |||||||||||||
| Mean.D | −1 | −1 | −1 | −1 | −1 | −1 | 6 | ||||||||||||
| Std.D | 1 | 1 | 1 | 1 | 2 | 2 | 8 | ||||||||||||
| Mean.HTrees | −1 | −1 | 1 | −2 | 5 | ||||||||||||||
| Std.HTrees | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 8 | ||||||||||
| Fcc.Tot | −1 | −1 | −1 | −1 | 4 | ||||||||||||||
| Fcc.Trees | 1 | 1 | 2 | ||||||||||||||||
| Texture | −1 | 1 | |||||||||||||||||
| MatOrg | −1 | 1 | −1 | 3 | |||||||||||||||
| PhSoil | −1 | 2 | 1 | 4 | |||||||||||||||
| RegFelling | −1 | 1 | |||||||||||||||||
| Class.PLAND | −1 | −1 | −1 | 3 | |||||||||||||||
| Class.SPLIT | 1 | 1 | |||||||||||||||||
| Land.SHDI | 1 | 1 | −1 | 3 | |||||||||||||||
| Land.NP | 1 | 1 | 2 |
| Variables | LU | Rich | ShDI | LU | Rich | ShDI | LU | Rich | ShDI | LU | Rich | ShDI | LU | Rich | ShDI | LU | Rich | ShDI | Total times in shrub models |
| Models | Net Arch | 9–15–1 | 3–19–1 | Net Arch | 3–23–1 | 6–0–1 | Net Arch | 7–15–1 | 6–13–1 | Net Arch | 8–3–1 | 7–19–1 | Net Arch | 8–7–1 | 8–9–1 | Net Arch | 5–23–1 | 7–17–1 | |
| rTrain | 0.66 | 0.57 | rTrain | 0.62 | 0.5 | rTrain | 0.62 | 0.55 | rTrain | 0.5 | 0.53 | rTrain | 0.5 | 0.6 | rTrain | 0.52 | 0.5 | ||
| rTest | 0.58 | 0.73 | rTest | 0.87 | 0.9 | rTest | 0.57 | 0.54 | rTest | 0.6 | 0.52 | rTest | 0.5 | 0.8 | rTest | 0.57 | 0.53 | ||
| rValid | 0.62 | 0.62 | rValid | 0.72 | 0.6 | rValid | 0.6 | 0.55 | rValid | 0.54 | 0.52 | rValid | 0.5 | 0.6 | rValid | 0.54 | 0.51 | ||
| Develop | Serres d’AncosaMature P.halepensis | Montmell Young P. halepensis | 1 | Replans de BerguedàYoung and mature P. sylvestris | Capçaleres del LlobregatMature P. sylvestris | Aspres Young and adult Q. suber | Vall de Rialbmature P. nigra mixed with Q. faginea, Q ilex | 1 | |||||||||||
| Even/Uneven | −1 | −1 | 1 | 3 | |||||||||||||||
| Mean.Hshrub | 2 | −1 | −1 | −2 | 1 | 1 | 8 | ||||||||||||
| Std.Hshrub | −1 | 1 | −1 | 2 | −1 | 6 | |||||||||||||
| D.Max | −1 | −2 | −1 | −1 | 1 | −1 | 7 | ||||||||||||
| D.Min | 1 | 1 | 1 | 3 | |||||||||||||||
| Mean.D | −1 | 1 | |||||||||||||||||
| Std.D | −1 | 1 | 2 | ||||||||||||||||
| Mean.HTrees | 3 | 3 | |||||||||||||||||
| Std.HTrees | 1 | 1 | 1 | 3 | |||||||||||||||
| Fcc.Tot | −1 | −2 | 3 | ||||||||||||||||
| Fcc.Trees | −1 | −1 | −1 | −2 | −1 | −1 | 2 | 1 | 10 | ||||||||||
| Texture | −1 | 1 | 2 | ||||||||||||||||
| MatOrg | 1 | 1 | 2 | ||||||||||||||||
| PhSoil | 1 | −1 | −1 | 2 | 1 | 6 | |||||||||||||
| RegFelling | 1 | 1 | |||||||||||||||||
| Class.PLAND | −1 | −1 | −1 | 3 | |||||||||||||||
| Class.SPLIT | −1 | −1 | −1 | −1 | 4 | ||||||||||||||
| Land.SHDI | 3 | −1 | 1 | 5 | |||||||||||||||
| Land.NP | 1 | −1 | −1 | 1 | 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghadban, S.; Ameztegui, A.; Rodrigues, M.; Chocarro, C.; Alcasena, F.; Vega-Garcia, C. Stand Structure and Local Landscape Variables Are the Dominant Factors Explaining Shrub and Tree Diversity in Mediterranean Forests. Sustainability 2021, 13, 11658. https://doi.org/10.3390/su132111658
Ghadban S, Ameztegui A, Rodrigues M, Chocarro C, Alcasena F, Vega-Garcia C. Stand Structure and Local Landscape Variables Are the Dominant Factors Explaining Shrub and Tree Diversity in Mediterranean Forests. Sustainability. 2021; 13(21):11658. https://doi.org/10.3390/su132111658
Chicago/Turabian StyleGhadban, Siba, Aitor Ameztegui, Marcos Rodrigues, Cristina Chocarro, Fermín Alcasena, and Cristina Vega-Garcia. 2021. "Stand Structure and Local Landscape Variables Are the Dominant Factors Explaining Shrub and Tree Diversity in Mediterranean Forests" Sustainability 13, no. 21: 11658. https://doi.org/10.3390/su132111658