
Decolourisation and Biodegradation of Textile Di-azo Dye Congo Red by Chryseobacterium geocarposphaerae DD3



Abstract
:1. Introduction
2. Materials and Methods
2.1. Wastewater Collection and Characteristics
2.2. Polyphasic Characterization of the Bacterial Isolate
2.2.1. Bacteriological Analysis
2.2.2. Molecular Characterization by 16S rRNA Amplification and Phylogenetic Analysis
2.3. Congo Red Decolourisation Study
2.4. Analysis of Biodegradation Using Different Spectroscopic Methods
2.5. Phytotoxicity Study
3. Results
3.1. Wastewater Collection and Characteristics
3.2. Polyphasic Characterisation and Identification of Chryseobacterium Geocarpopshaerae DD3
3.3. Congo Red Decolourisation Study
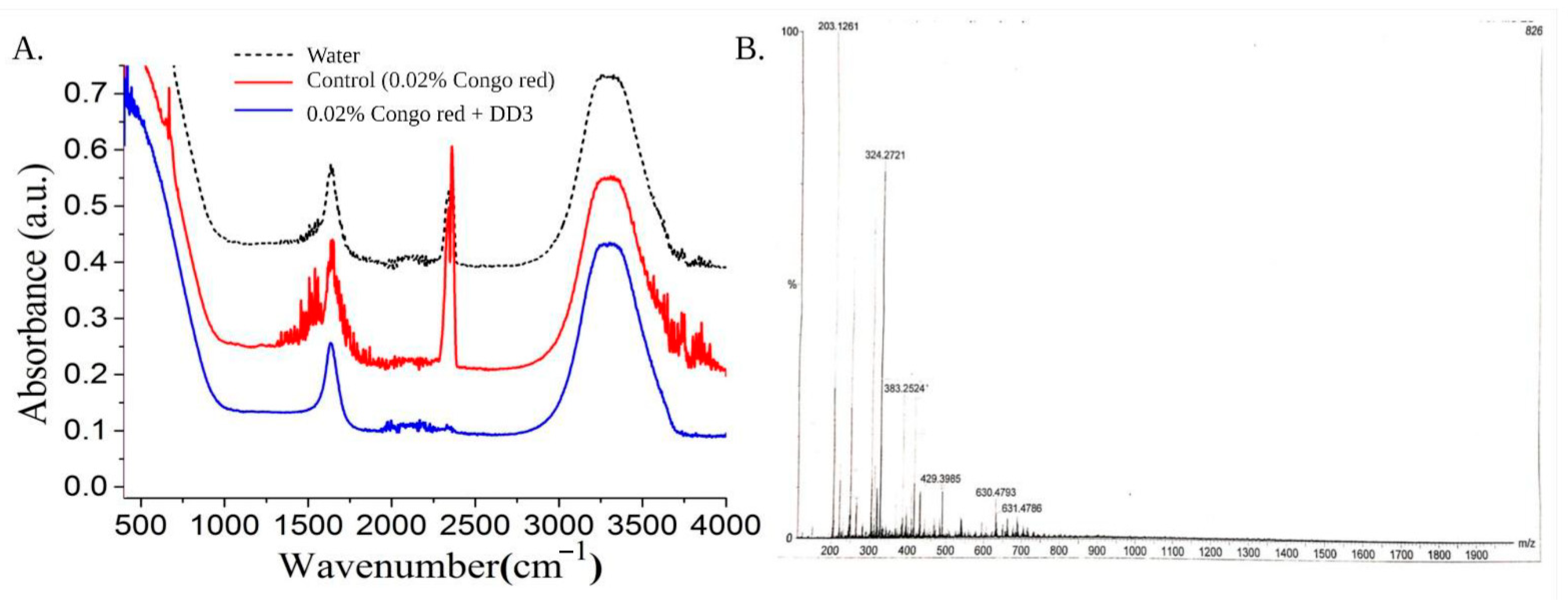
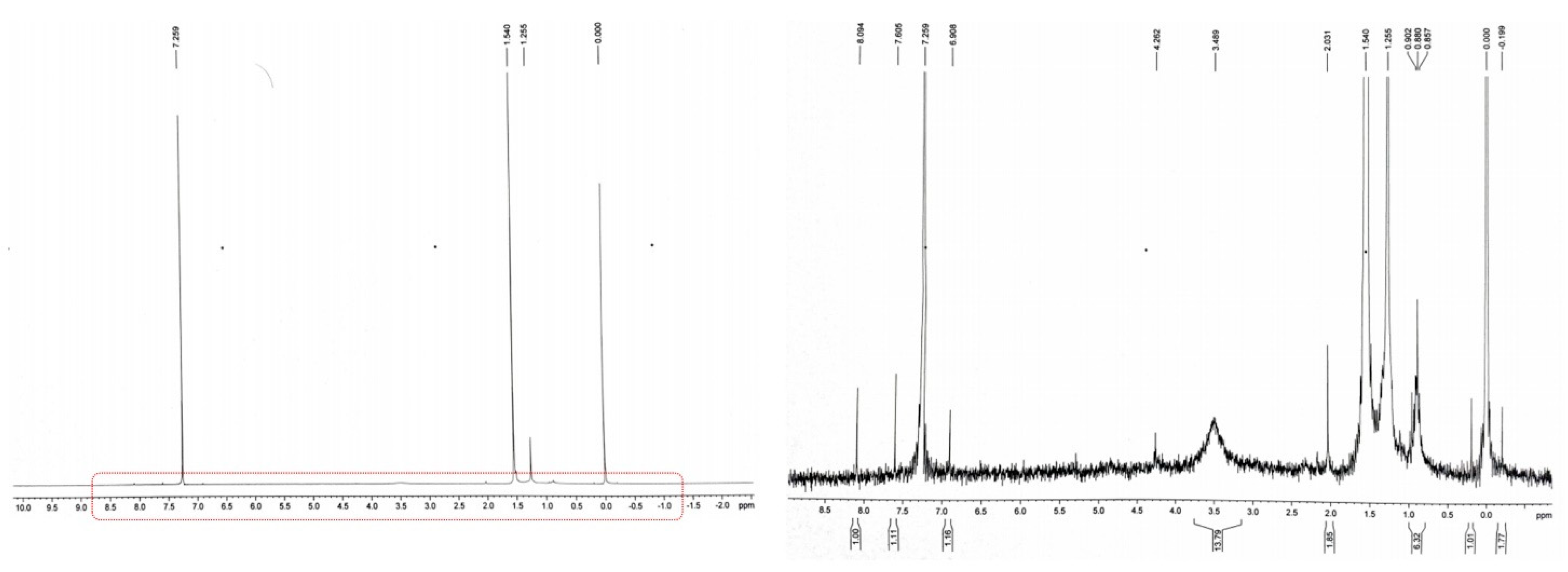
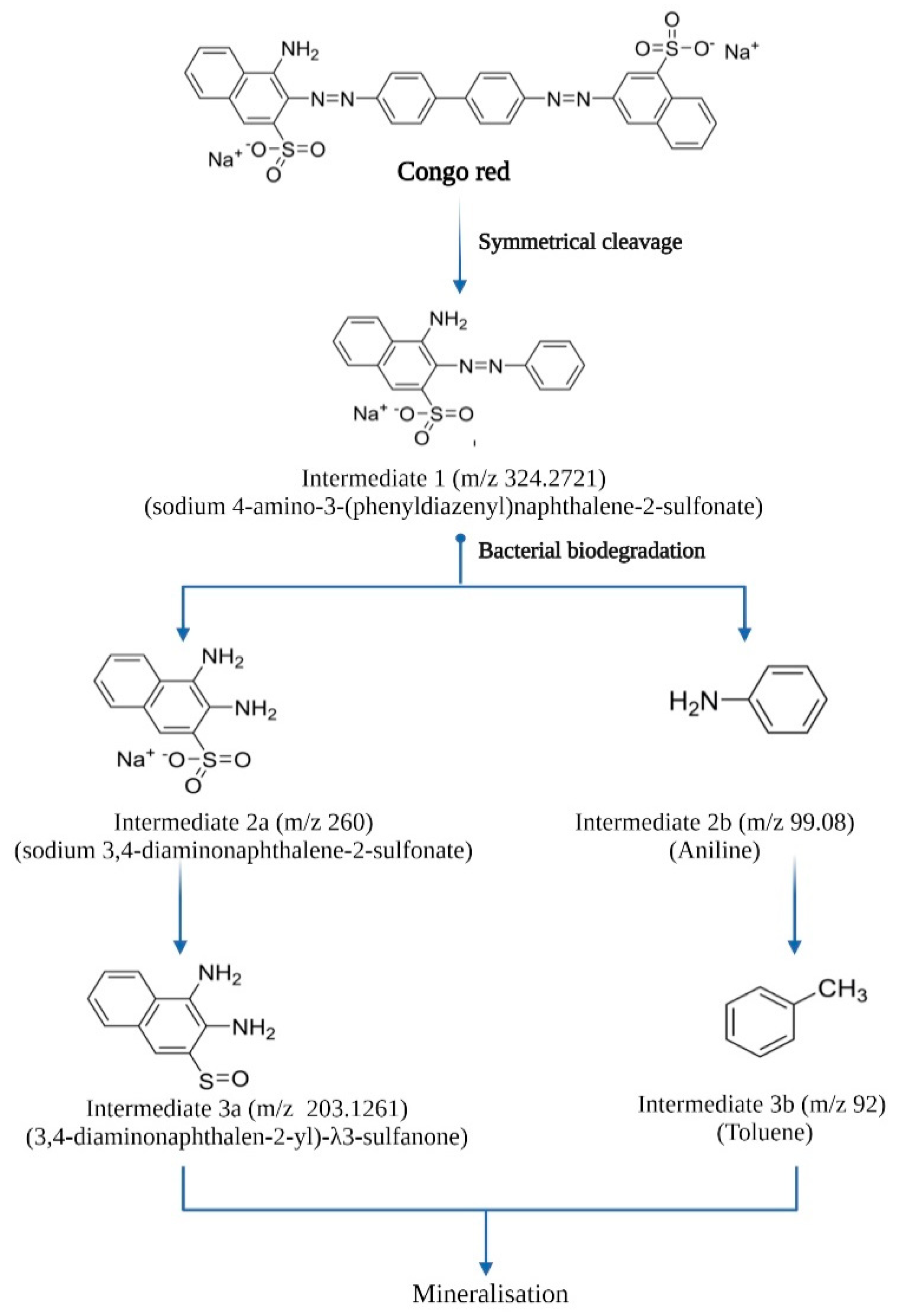
3.4. Analysis of Biodegradation Using Different Spectroscopic Methods
3.5. Phytotoxicity Study
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sarkar, S.; Ponce, N.T.; Banerjee, A.; Bandopadhyay, R.; Rajendran, S.; Lichtfouse, E. Green polymeric nanomaterials for the photocatalytic degradation of dyes: A review. Environ. Chem. Lett. 2020, 18, 1569–1580. [Google Scholar] [CrossRef]
- Agha, Y.Y.; Bahjat, S.A.; Thanoon, M.F. Assessment of Bacterial Pigments as Textile Colorants. Indian J. Public Health Res. Dev. 2019, 10, 1565–1569. [Google Scholar] [CrossRef]
- Carneiro, P.A.; Umbuzeiro, G.A.; Oliveira, D.P.; Zanoni, M.V.B. Assessment of water contamination caused by a mutagenic textile effluent/dyehouse effluent bearing disperse dyes. J. Hazard. Mater. 2010, 174, 694–699. [Google Scholar] [CrossRef] [PubMed]
- Baban, A.; Yediler, A.; Lienert, D.; Kemerdere, N.; Kettrup, A. Ozonation of high strength segregated effluents from a woollen textile dyeing and finishing plant. Dye. Pigment. 2003, 58, 93–98. [Google Scholar] [CrossRef]
- Sudha, M.; Saranya, A.; Selvakumar, G.; Sivakumar, N. Microbial degradation of azo dyes: A review. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 670–690. [Google Scholar]
- Tkaczyk, A.; Mitrowska, K.; Posyniak, A. Synthetic organic dyes as contaminants of the aquatic environment and their implications for ecosystems: A review. Sci. Total Environ. 2020, 717, 137222. [Google Scholar] [CrossRef] [PubMed]
- Yaseen, D.A.; Scholz, M. Textile dye wastewater characteristics and constituents of synthetic effluents: A critical review. Int. J. Environ. Sci. Technol. 2019, 16, 1193–1226. [Google Scholar] [CrossRef] [Green Version]
- Mishra, S.; Nayak, J.K.; Maiti, A. Bacteria-mediated bio-degradation of reactive azo dyes coupled with bio-energy generation from model wastewater. Clean Technol. Environ. Policy 2020, 22, 651–667. [Google Scholar] [CrossRef]
- Singh, R.L.; Singh, P.K.; Singh, R.P. Enzymatic decolorization and degradation of azo dyes—A review. Int. Biodeterior. Biodegrad. 2015, 104, 21–31. [Google Scholar] [CrossRef]
- Lade, H.; Govindwar, S.; Paul, D. Low-cost biodegradation and detoxification of textile azo dye CI reactive blue 172 by Providencia rettgeri strain HSL1. J. Chem. 2015, 2015, 894109. [Google Scholar] [CrossRef] [Green Version]
- Solís, M.; Solís, A.; Pérez, H.I.; Manjarrez, N.; Flores, M. Microbial decolouration of azo dyes: A review. Process Biochem. 2012, 47, 1723–1748. [Google Scholar] [CrossRef]
- Ito, T.; Adachi, Y.; Yamanashi, Y.; Shimada, Y. Long–term natural remediation process in textile dye–polluted river sediment driven by bacterial community changes. Water Res. 2016, 100, 458–465. [Google Scholar] [CrossRef]
- Yu, J.; Wang, X.; Yue, P.L. Optimal decolorization and kinetic modeling of synthetic dyes by Pseudomonas strains. Water Res. 2001, 35, 3579–3586. [Google Scholar] [CrossRef]
- Paul, S.A.; Chavan, S.K.; Khambe, S.D. Studies on characterization of textile industrial waste water in Solapur city. Int. J. Chem. Sci. 2012, 10, 635–642. [Google Scholar]
- Guadie, A.; Tizazu, S.; Melese, M.; Guo, W.; Ngo, H.H.; Xia, S. Biodecolorization of textile azo dye using Bacillus sp. strain CH12 isolated from alkaline lake. Biotechnol. Rep. 2017, 15, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Yaseen, D.A.; Scholz, M. Shallow pond systems planted with Lemna minor treating azo dyes. Ecol. Eng. 2016, 94, 295–305. [Google Scholar] [CrossRef]
- Sekomo, C.B.; Rousseau, D.P.; Saleh, S.A.; Lens, P.N. Heavy metal removal in duckweed and algae ponds as a polishing step for textile wastewater treatment. Ecol. Eng. 2012, 44, 102–110. [Google Scholar] [CrossRef]
- Shah, A.H.; Manikandan, E.; Ahmed, M.B.; Ganesan, V. Enhanced bioactivity of Ag/ZnO nanorods-A comparative antibacterial study. J. Nanomed. Nanotechol. 2013, 4, 3. [Google Scholar] [CrossRef]
- Sarkar, S.; Banerjee, A.; Halder, U.; Biswas, R.; Bandopadhyay, R. Degradation of synthetic azo dyes of textile industry: A sustainable approach using microbial enzymes. Water Conserv. Sci. Eng. 2017, 2, 121–131. [Google Scholar] [CrossRef] [Green Version]
- Zerva, I.; Remmas, N.; Kagalou, I.; Melidis, P.; Ariantsi, M.; Sylaios, G.; Ntougias, S. Effect of Chlorination on Microbiological Quality of Effluent of a Full-Scale Wastewater Treatment Plant. Life 2021, 11, 68. [Google Scholar] [CrossRef]
- Wuhrmann, K.; Mechsner, K.L.; Kappeler, T.H. Investigation on rate—Determining factors in the microbial reduction of azo dyes. Appl. Microbiol. Biotechnol. 1980, 9, 325–338. [Google Scholar] [CrossRef]
- Dave, S.R.; Patel, T.L.; Tipre, D.R. Bacterial Degradation of Azo Dye Containing Wastes. In Microbial Degradation of Synthetic Dyes in Wastewaters; Singh, S., Ed.; Springer: Cham, Switzerland, 2015; pp. 57–83. [Google Scholar] [CrossRef]
- Manjarrez Paba, G.; Baldiris Ávila, R.; Baena Baldiris, D. Application of environmental bacteria as potential methods of azo dye degradation systems. Glob. J. Environ. Sci. Manag. 2021, 7, 131–154. [Google Scholar] [CrossRef]
- Gharbani, P.; Tabatabaii, S.M.; Mehrizad, A. Removal of Congo red from textile wastewater by ozonation. Int. J. Environ. Sci. Technol. 2008, 5, 495–500. [Google Scholar] [CrossRef] [Green Version]
- Gopinath, K.P.; Murugesan, S.; Abraham, J.; Muthukumar, K. Bacillus sp. mutant for improved biodegradation of Congo red: Random mutagenesis approach. Bioresour. Technol. 2009, 100, 6295–6300. [Google Scholar] [CrossRef]
- Hsueh, C.C.; Chen, B.Y. Comparative study on reaction selectivity of azo dye decolorization by Pseudomonas luteola. J. Hazard. Mater. 2007, 141, 842–849. [Google Scholar] [CrossRef] [PubMed]
- Ng, I.S.; Chen, T.; Lin, R.; Zhang, X.; Ni, C.; Sun, D. Decolorization of textile azo dye and Congo red by an isolated strain of the dissimilatory manganese-reducing bacterium Shewanella xiamenensis BC01. Appl. Microbiol. Biotechnol. 2014, 98, 2297–2308. [Google Scholar] [CrossRef] [PubMed]
- Nishioka, T.; Elsharkawy, M.M.; Suga, H.; Kageyama, K.; Hyakumachi, M.; Shimizu, M. Development of culture medium for the isolation of Flavobacterium and Chryseobacterium from rhizosphere soil. Microbes Environ. 2016, 31, ME15144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- American Public Health Association. Standard Methods Barrfor the Examination of Water and Waste Water, 20th ed.; American Public Health Association: Washington, DC, USA, 1998. [Google Scholar]
- Pelczar, M.J.; Bard, R.C.; Burnett, G.W.; Conn, H.J.; Demoss, R.D.; Euans, E.E.; Weiss, F.A.; Jennison, M.W.; Meckee, A.P.; Riker, A.J.; et al. Manual of Microbiological Methods. In Society of American Bacteriology; McGraw Hill: New York, NY, USA, 1957. [Google Scholar]
- Rana, S.; Kumar, K. Study of Phytotoxic effect of textile wastewater on seed germination and seedling growth of Triticum aestivum. Int. J. Biosci. Technol. 2017, 10, 58–66. [Google Scholar]
- Balapure, K.; Aghera, P.; Bhatt, N.; Madamwar, D. Community synergism: Degradation of triazine dye reactive black 1 by mixed bacterial cultures KND_PR under microaerophilic and aerobic conditions. Environ. Process. 2019, 6, 713–739. [Google Scholar] [CrossRef]
- Mishra, S.; Mohanty, P.; Maiti, A. Bacterial mediated bio-decolourization of wastewater containing mixed reactive dyes using jack-fruit seed as co-substrate: Process optimization. J. Clean. Prod. 2019, 235, 21–33. [Google Scholar] [CrossRef]
- You, R.; Domínguez, C.; Matamoros, V.; Bayona, J.M.; Díez, S. Chemical characterization and phytotoxicity assessment of peri-urban soils using seed germination and root elongation tests. Environ. Sci. Pollut. R. 2019, 26, 34401–34411. [Google Scholar] [CrossRef]
- Nachiyar, C.V.; Rajkumar, G.S. Degradation of a tannery and textile dye, Navitan Fast Blue S5R by Pseudomonas aeruginosa. World J. Microbiol. Biotechnol. 2003, 19, 609–614. [Google Scholar] [CrossRef]
- Kolekar, Y.M.; Pawar, S.P.; Gawai, K.R.; Lokhande, P.D.; Shouche, Y.S.; Kodam, K.M. Decolorization and degradation of Disperse Blue 79 and Acid Orange 10, by Bacillus fusiformis KMK5 isolated from the textile dye contaminated soil. Bioresour. Technol. 2008, 99, 8999–9003. [Google Scholar] [CrossRef] [PubMed]
- Hamad, M.T.; Saied, M.S. Kinetic studies of Congo red dye adsorption by immobilized Aspergillus niger on alginate. Appl. Water Sci. 2021, 11, 35. [Google Scholar] [CrossRef]
- Kapdan, I.K.; Kargia, F.; McMullan, G.; Marchant, R. Effect of environmental conditions on biological decolorization of textile dyestuff by C. versicolor. Enzyme Microb. Technol. 2000, 26, 381–387. [Google Scholar] [CrossRef]
- Chen, K.C.; Wu, J.Y.; Liou, D.J.; Hwang, S.C.J. Decolorization of the textile dyes by newly isolated bacterial strains. J. Biotechnol. 2003, 101, 57–68. [Google Scholar] [CrossRef]
- Asad, S.; Amoozegar, M.A.; Pourbabaee, A.; Sarbolouki, M.N.; Dastgheib, S.M.M. Decolorization of textile azo dyes by newly isolated halophilic and halotolerant bacteria. Bioresour. Technol. 2007, 98, 2082–2088. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Singh, P.K.; Singh, R.L. Bacterial Decolorization of Textile Azo Dye Acid Orange by Staphylococcus hominis RMLRT03. Toxicol. Int. 2014, 21, 160–166. [Google Scholar] [CrossRef] [Green Version]
- Sari, I.P.; Simarani, K. Decolorization of selected azo dye by Lysinibacillus fusiformis W1B6: Biodegradation optimization, isotherm, and kinetic study biosorption mechanism. Adsorpt. Sci. Technol. 2019, 37, 492–508. [Google Scholar] [CrossRef] [Green Version]
- Kishor, R.; Purchase, D.; Saratale, G.D.; Ferreira, L.F.R.; Bilal, M.; Iqbal, H.M.; Bharagava, R.N. Environment friendly degradation and detoxification of Congo red dye and textile industry wastewater by a newly isolated Bacillus cohnni (RKS9). Environ. Technol. Innov. 2021, 22, 101425. [Google Scholar] [CrossRef]
- Asses, N.; Ayed, L.; Hkiri, N.; Hamdi, M. Congo red decolorization and detoxification by Aspergillus niger: Removal mechanisms and dye degradation pathway. BioMed Res. Int. 2018, 2018, 3049686. [Google Scholar] [CrossRef] [Green Version]
- D’Souza, E.; Fulke, A.B.; Mulani, N.; Ram, A.; Asodekar, M.; Narkhede, N.; Gajbhiye, S.N. Decolorization of Congo red mediated by marine Alcaligenes species isolated from Indian West coast sediments. Environ. Earth Sci. 2017, 76, 721. [Google Scholar] [CrossRef]
- Barragán, B.E.; Costa, C.; Marquez, M.C. Biodegradation of azo dyes by bacteria inoculated on solid media. Dye. Pigment. 2007, 75, 73–81. [Google Scholar] [CrossRef]
- Masarbo, R.S.; Niranjana, S.R.; Monisha, T.R.; Nayak, A.S.; Karegoudar, T.B. Efficient decolorization and detoxification of sulphonated azo dye Ponceau 4R by using single and mixed bacterial consortia. Biocatal. Biotransform. 2019, 37, 367–376. [Google Scholar] [CrossRef]
- Haque, M.M.; Haque, M.A.; Mosharaf, M.K.; Marcus, P.K. Decolorization, degradation and detoxification of carcinogenic sulfonated azo dye methyl orange by newly developed biofilm consortia. Saudi J. Biol. Sci. 2021, 28, 793–804. [Google Scholar] [CrossRef] [PubMed]
- Telke, A.A.; Joshi, S.M.; Jadhav, S.U.; Tamboli, D.P.; Govindwar, S.P. Decolorization and detoxification of Congo red and textile industry effluent by an isolated bacterium Pseudomonas sp. SU-EBT. Biodegradation 2010, 21, 283–296. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Control | DD3 Treated Water | CR Solution |
|---|---|---|---|
| Plumule length | 3 cm | 1.9 cm | - |
| Radicle length | 5.6 cm | 5.4 cm | - |
| Root length | 2.4 cm | 3.4 cm | 1.1 cm |
| Number of roots | 9 | 11 | 2 |
| Shoot length | 6.0 cm | 4.4 cm | 1.3 cm |
| Number of leaves | 2 | 2 | - |
| Length of leaf | 1.9 cm | 1.5 cm | - |
| Width of leaf | 0.8 cm | 0.8 cm | - |
| Germination rate | 90% | 80% | 50% |
| Relative germination | 100% | 88.89% | 55.56% |
| % toxicity | 0% | 3.57% | 100% |
| Tolerance index | - | 1.41 | 0.45 |
| Vigour index | 756 | 624 | 120 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarkar, S.; Echeverría-Vega, A.; Banerjee, A.; Bandopadhyay, R. Decolourisation and Biodegradation of Textile Di-azo Dye Congo Red by Chryseobacterium geocarposphaerae DD3. Sustainability 2021, 13, 10850. https://doi.org/10.3390/su131910850
Sarkar S, Echeverría-Vega A, Banerjee A, Bandopadhyay R. Decolourisation and Biodegradation of Textile Di-azo Dye Congo Red by Chryseobacterium geocarposphaerae DD3. Sustainability. 2021; 13(19):10850. https://doi.org/10.3390/su131910850
Chicago/Turabian StyleSarkar, Shrabana, Alex Echeverría-Vega, Aparna Banerjee, and Rajib Bandopadhyay. 2021. "Decolourisation and Biodegradation of Textile Di-azo Dye Congo Red by Chryseobacterium geocarposphaerae DD3" Sustainability 13, no. 19: 10850. https://doi.org/10.3390/su131910850