
Macroecology of North European Wet Grassland Landscapes: Habitat Quality, Waders, Avian Predators and Nest Predation



Abstract
:1. Introduction
2. Materials and Methods
2.1. A Macroecological Approach with Multiple Case Studies
2.2. Northern Europe as a Landscape Laboratory
2.3. Six Case Studies in a Gradient of Landscape Change
2.3.1. Sweden
2.3.2. Lithuania and Belarus
2.3.3. Iceland
2.4. Waders, Avian Predators and Predation Pressure
2.5. Artificial Nest Predation as Proxy of Predation Pressure
3. Results
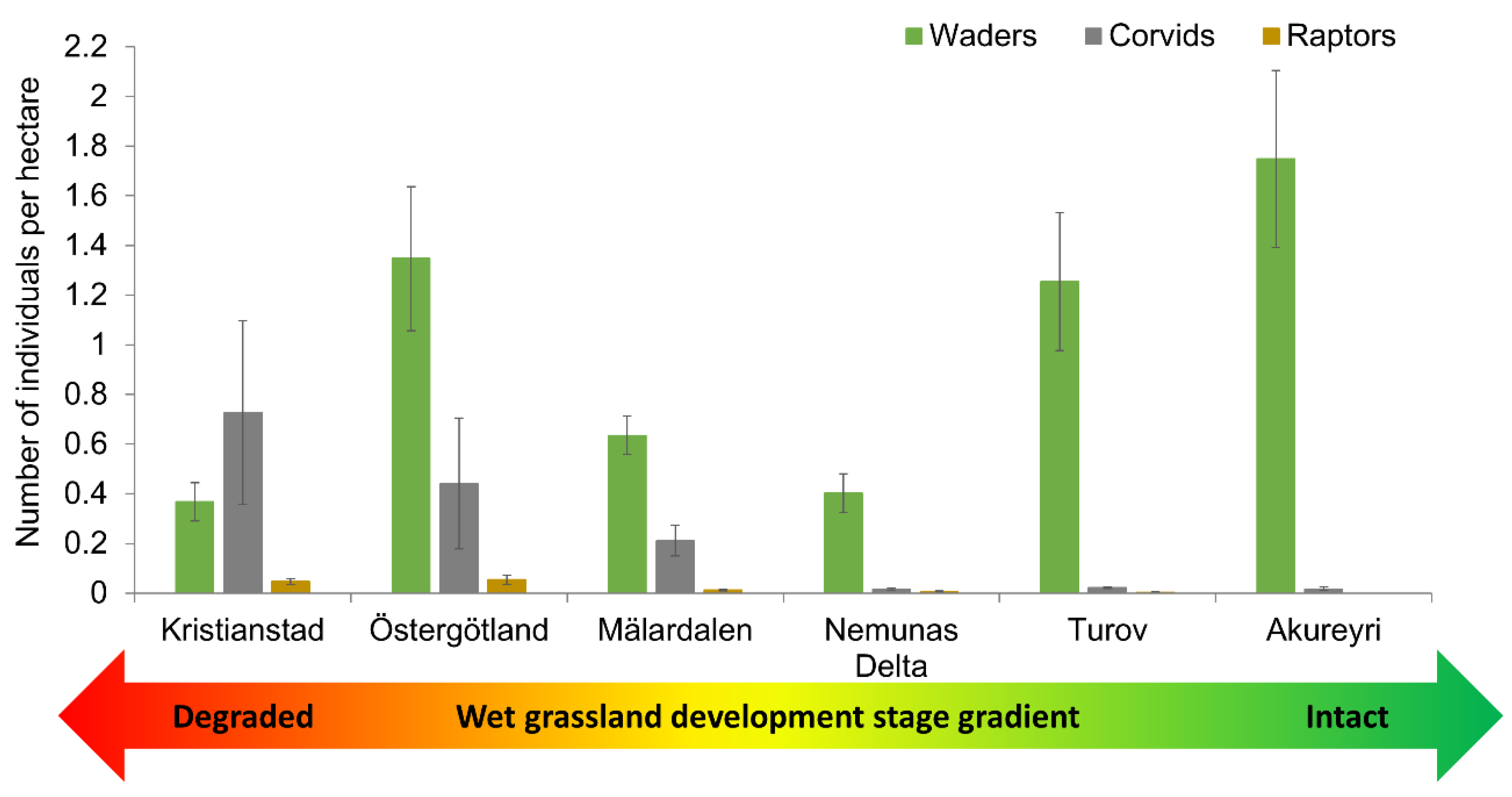
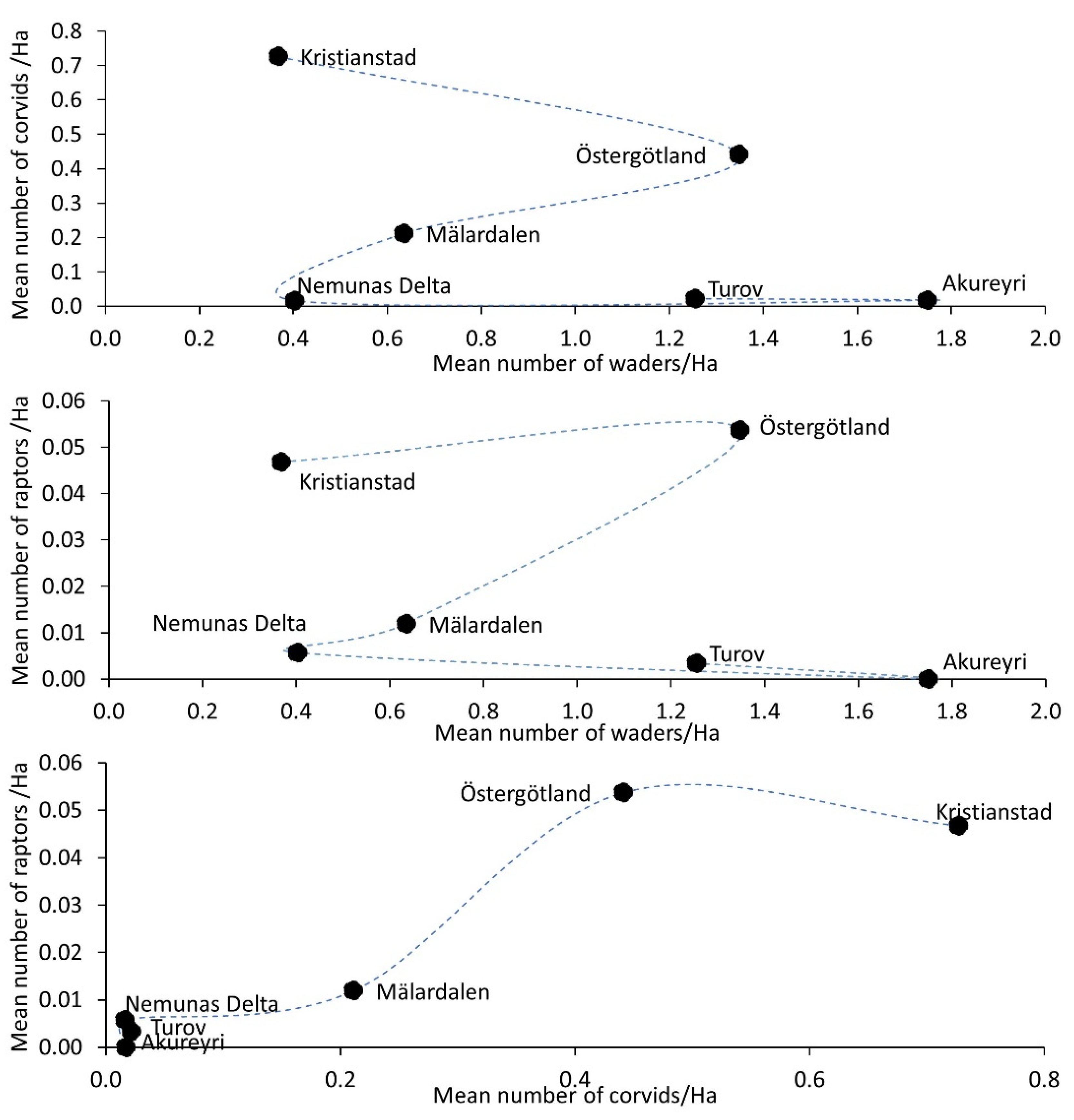
3.1. Waders and Predators in the Macroecological Gradient
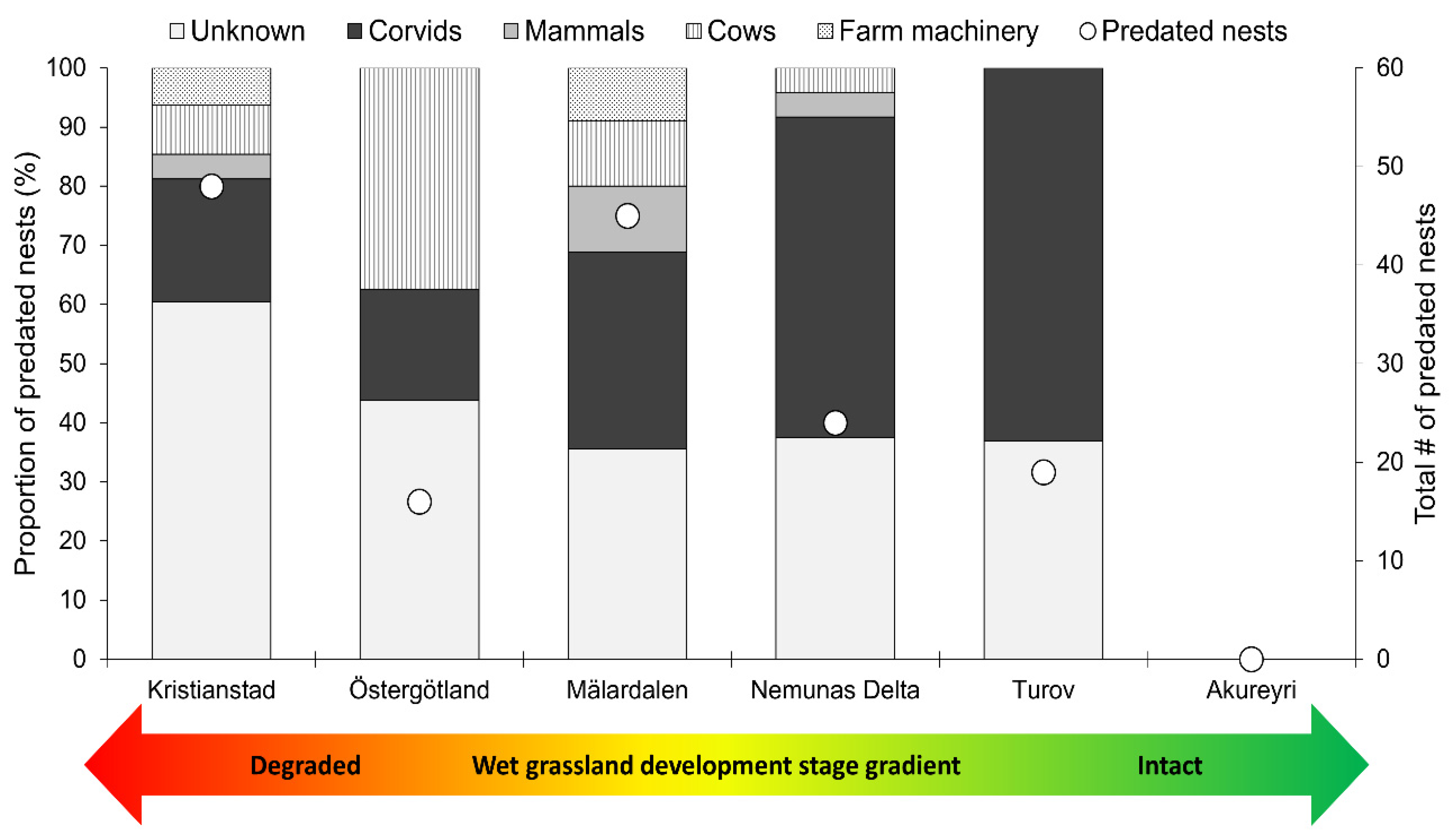
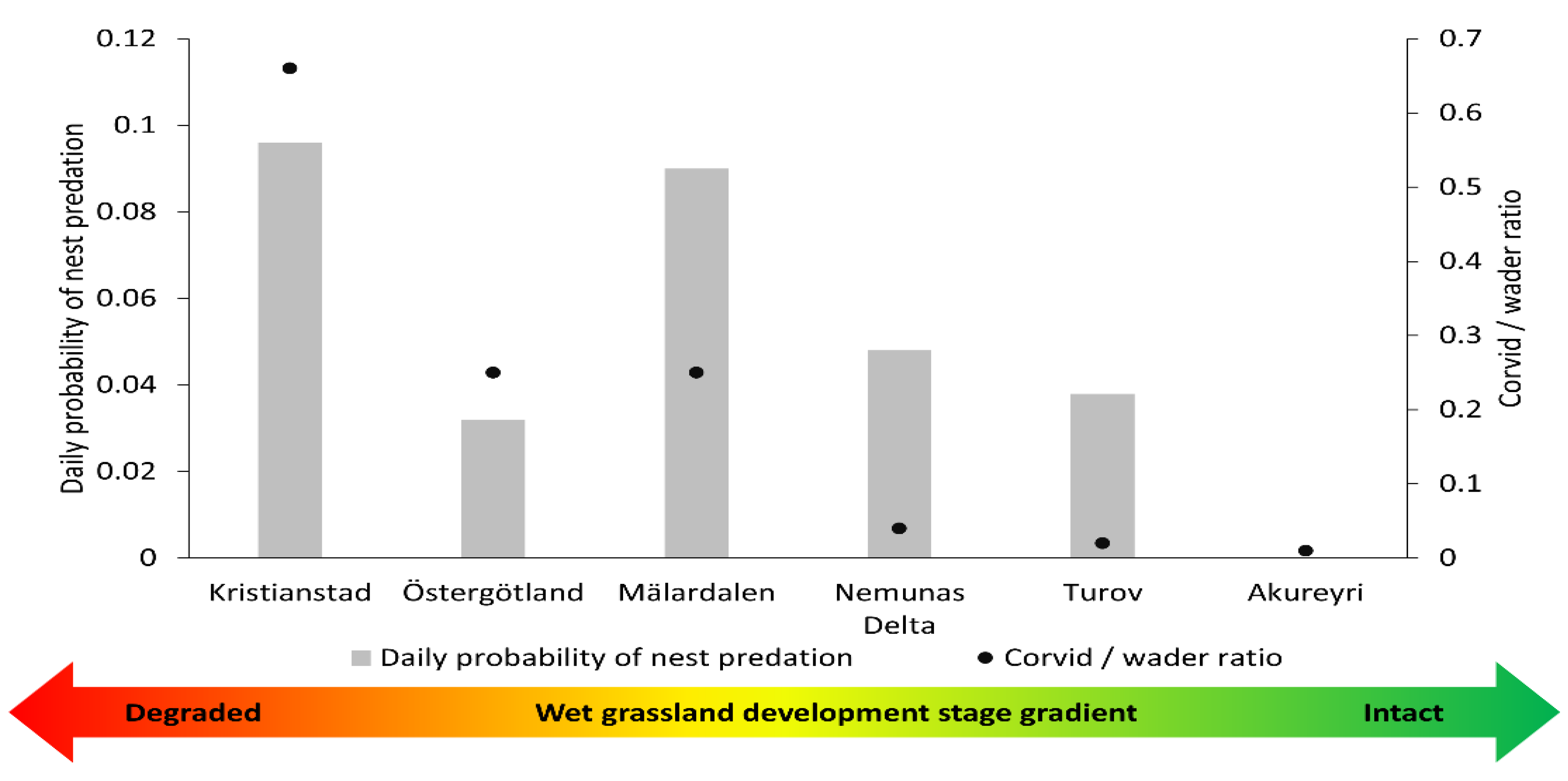
3.2. Artificial Nest Predation
4. Discussion
4.1. Clear Macroecological Patterns across Landscapes
4.2. Small and Broad Scale Perspectives in Space and Time
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Joyce, C.; Wade, P. Wet Grasslands: A European Perspective; John Wiley: Chichester, UK, 1998; p. 340. [Google Scholar]
- Benstead, P.; Drake, M.; Jose, P.; Mountford, O.; Newbold, C.; Treweek, J. The Wet Grassland Guide; Royal Society for the Protection of Birds: Sandy, UK, 1997. [Google Scholar]
- Price, E. Lowland Grassland and Heathland Habitats; Routledge: London, UK, 2003. [Google Scholar]
- Antrop, M. The transformation of the Mediterranean landscapes: An experience of 25 years of observations. Landsc. Urban Plan. 1993, 24, 3–13. [Google Scholar] [CrossRef]
- Hartel, T.; Plieninger, T. European Wood-Pastures in Transition: A Social-Ecological Approach; Routledge: London, UK, 2014. [Google Scholar]
- Benstead, P.; Jose, P.; Joyce, C.; Wade, P. European Wet Grassland: Guidelines for Management and Restoration; RSPB: Sandy, UK, 1999. [Google Scholar]
- Newton, I. The recent declines of farmland bird populations in Britain: An appraisal of causal factors and conservation actions. IBIS 2004, 146, 579–600. [Google Scholar] [CrossRef]
- Schekkerman, H.; Teunissen, W.; Oosterveld, E. The effect of ‘mosaic management’ on the demography of Black-tailed Godwit Limosa limosa on farmland. J. Appl. Ecol. 2008, 45, 1067–1075. [Google Scholar] [CrossRef]
- Beintema, A.J. Man-made polders in the netherlands: A traditional habitat for shorebirds. Colonial Waterbirds 1986, 9, 196–202. [Google Scholar] [CrossRef]
- Brinson, M.M.; Malvárez, A. Temperate freshwater wetlands: Types, status, and threats. Environ. Conserv. 2002, 29, 115–133. [Google Scholar] [CrossRef]
- Illyés, E.; Botta-Dukát, Z.; Molnár, Z. Patch and landscape factors affecting the naturalness-based quality of three model grassland habitats in Hungary. Acta Bot. Hung. 2008, 50, 179–197. [Google Scholar] [CrossRef]
- Durant, D.; Tichit, M.; Kernéïs, E.; Fritz, H. Management of agricultural wet grasslands for breeding waders: Integrating ecological and livestock system perspectives—A review. Biodivers. Conserv. 2008, 17, 2275–2295. [Google Scholar] [CrossRef]
- Catry, T.; Alves, J.A.; Andrade, J.; Costa, H.; Dias, M.P.; Fernandes, P.; Leal, A.; Lourenco, P.M.; Martins, R.C.; Moniz, F.; et al. Long-term declines of wader populations at the Tagus estuary, Portugal: A response to global or local factors? Bird Conserv. Int. 2011, 21, 438–453. [Google Scholar] [CrossRef] [Green Version]
- Roodbergen, M.; Werf, B.; Hötker, H. Revealing the contributions of reproduction and survival to the Europe-wide decline in meadow birds: Review and meta-analysis. J. Ornithol. 2011, 153, 53–74. [Google Scholar] [CrossRef]
- Gill, J.A.; Langston, R.H.W.; Alves, J.A.; Atkinson, P.W.; Bocher, P.; Cidraes Vieira, N.; Crockford, N.J.; Gélinaud, G.; Groen, N.; Gunnarsson, T.G.; et al. Contrasting trends in two Black-tailed Godwit populations: A review of causes and recommendations. Wader Study Group Bull. 2007, 114, 43–50. [Google Scholar]
- Schekkerman, H.; Teunissen, W.; Oosterveld, E. Mortality of Black-tailed Godwit Limosa limosa and Northern Lapwing Vanellus vanellus chicks in wet grasslands: Influence of predation and agriculture. J. Ornithol. 2009, 150, 133–145. [Google Scholar] [CrossRef]
- Thorup, O. Breeding waders in Europe 2000. In International Wader Studies; Wader Study Group: Thetford, UK, 2005; Volume 14. [Google Scholar]
- Ottvall, R.; Edenius, L.; Elmberg, J.; Engström, H.; Green, M.; Holmqvist, N.; Lindström, Å.; Pärt, T.; Tjernberg, M. Population trends for Swedish breeding birds. Ornis Svec. 2009, 19, 117–192. [Google Scholar]
- Lindström, Å.; Green, M. Monitoring Population Changes of Birds in Sweden. Annual Report for 2012; Department of Biology, Lund University: Lund, Sweden, 2013; 80p. [Google Scholar]
- Manton, M.; Angelstam, P.; Milberg, P.; Elbakidze, M. Wet grasslands as a green infrastructure for ecological sustainability: Wader conservation in Southern Sweden as a case study. Sustainability 2016, 8, 340. [Google Scholar] [CrossRef] [Green Version]
- Bergner, A. Fåglar på Strandängar i Östergötland—Inventeringar 1996–2013; Länsstyrelsen Östergötland: Linköping, Sweden, 2013. [Google Scholar]
- Nilsson, S. Skandinavisk Fauna. Foglarna. Första Bandet; Gleerups: Lund, Sweden, 1858. [Google Scholar]
- Turner, M.G. Landscape ecology: The effect of pattern on process. Annu. Rev. Ecol. Syst. 1989, 20, 171–197. [Google Scholar] [CrossRef]
- Gustafson, E.J. Quantifying landscape spatial pattern: What is the state of the art? Ecosystems 1998, 1, 143–156. [Google Scholar] [CrossRef]
- Ward, J.V.; Malard, F.; Tockner, K. Landscape ecology: A framework for integrating pattern and process in river corridors. Landsc. Ecol. 2002, 17, 35–45. [Google Scholar] [CrossRef]
- Forman, R.T. Some general principles of landscape and regional ecology. Landsc. Ecol. 1995, 10, 133–142. [Google Scholar] [CrossRef]
- Nagendra, H.; Munroe, D.K.; Southworth, J. From pattern to process: Landscape fragmentation and the analysis of land use/land cover change. Agric. Ecosyst. Environ. 2004, 101, 111–115. [Google Scholar] [CrossRef]
- Tuvendal, M.; Elmqvist, T. Ecosystem services linking social and ecological systems: River brownification and the response of downstream stakeholders. Ecol. Soc. 2011, 16, 21. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Fischer, J. Habitat Fragmentation and Landscape Change: An Ecological and Conservation Synthesis; Island Press: Washington, DC, USA, 2006; p. 329. [Google Scholar]
- Ives, A.R.; Cardinale, B.J.; Snyder, W.E. A synthesis of subdisciplines: Predator–prey interactions, and biodiversity and ecosystem functioning. Ecol. Lett. 2005, 8, 102–116. [Google Scholar] [CrossRef]
- Andrén, H.; Angelstam, P.; Lindström, E.; Widén, P. Differences in predation pressure in relation to habitat fragmentation: An experiment. Oikos 1985, 45, 273–277. [Google Scholar] [CrossRef]
- Angelstam, P. Predation on Ground-Nesting Birds’ Nests in Relation to Predator Densities and Habitat Edge. Oikos 1986, 47, 365–373. [Google Scholar] [CrossRef]
- Kentie, R.; Both, C.; Hooijmeijer, J.C.E.W.; Piersma, T. Management of modern agricultural landscapes increases nest predation rates in Black-tailed Godwits Limosa limosa. IBIS 2015, 157, 614–625. [Google Scholar] [CrossRef]
- Amar, A.; Thirgood, S.; Pearce-Higgins, J.; Redpath, S. The impact of raptors on the abundance of upland passerines and waders. Oikos 2008, 117, 1143–1152. [Google Scholar] [CrossRef]
- Loman, J.; Göransson, G. Egg shell dumps and Crow Corvus cornix predation on simulated birds’ nests. Oikos 1978, 30, 461–466. [Google Scholar] [CrossRef] [Green Version]
- Cronert, H. Strandängsfåglar i Vattenriket. Häckfågelkarteringarna och simultanräkningar. Spoven 2014, 1, 1–19. [Google Scholar]
- Pehlak, H.; Lõhmus, A. An artificial nest experiment indicates equal nesting success of waders in coastal meadows and mires. Ornis Fenn. 2008, 85, 66–71. [Google Scholar]
- Martin, T.E. Processes organizing open-nesting bird assemblages: Competition or nest predation? Evol. Ecol. 1988, 2, 37–50. [Google Scholar] [CrossRef]
- Angelstam, P.; Axelsson, R.; Elbakidze, M.; Laestadius, L.; Lazdinis, M.; Nordberg, M.; Pătru-Stupariu, I.; Smith, M. Knowledge production and learning for sustainable forest management on the ground: Pan-European landscapes as a time machine. Forestry 2011, 84, 581–596. [Google Scholar] [CrossRef] [Green Version]
- Angelstam, P.; Manton, M.; Yamelynets, T.; Fedoriak, M.; Albulescu, A.-C.; Bravo, F.; Cruz, F.; Jaroszewicz, B.; Kavtarishvili, M.; Muñoz-Rojas, J.; et al. Maintaining natural and traditional cultural green infrastructures across Europe: Learning from historic and current landscape transformations. Landsc. Ecol. 2021, 36, 637–663. [Google Scholar] [CrossRef]
- Angelstam, P.; Fedoriak, M.; Cruz, F.; Muñoz-Rojas, J.; Yamelynets, T.; Manton, M.; Washbourne, C.-L.; Dobrynin, D.; Izakovicova, Z.; Jansson, N.; et al. Meeting places and social capital supporting rural landscape stewardship: A Pan-European horizon scanning. Ecol. Soc. 2021, 26. [Google Scholar] [CrossRef]
- Manton, M.; Angelstam, P.; Naumov, V. Effects of Land Use Intensification on Avian Predator Assemblages: A Comparison of Landscapes with Different Histories in Northern Europe. Diversity 2019, 11, 70. [Google Scholar] [CrossRef] [Green Version]
- Papworth, S.; Rist, J.; Coad, L.; Milner-Gulland, E. Evidence for shifting baseline syndrome in conservation. Conserv. Lett. 2009, 2, 93–100. [Google Scholar] [CrossRef]
- Pauly, D. Anecdotes and the shifting baseline syndrome of fisheries. Trends Ecol. Evol. 1995, 10, 430. [Google Scholar] [CrossRef]
- Jongman, R.H.G.; Bouwma, I.M.; Griffioen, A.; Jones-Walters, L.; Van Doorn, A.M. The Pan European Ecological Network: PEEN. Landsc. Ecol. 2011, 26, 311–326. [Google Scholar] [CrossRef]
- Wiens, J.A. Spatial scaling in ecology. Funct. Ecol. 1989, 3, 385–397. [Google Scholar] [CrossRef]
- Hall, L.S.; Krausman, P.R.; Morrison, M.L. The habitat concept and a plea for standard terminology. Wildl. Soc. Bull. 1997, 25, 173–182. [Google Scholar]
- Eglington, S.M.; Gill, J.A.; Smart, M.A.; Sutherland, W.J.; Watkinson, A.R.; Bolton, M. Habitat management and patterns of predation of Northern Lapwings on wet grasslands: The influence of linear habitat structures at different spatial scales. Biol. Conserv. 2009, 142, 314–324. [Google Scholar] [CrossRef]
- Manton, M.; Angelstam, P. Defining Benchmarks for Restoration of Green Infrastructure: A Case Study Combining the Historical Range of Variability of Habitat and Species’ Requirements. Sustainability 2018, 10, 326. [Google Scholar] [CrossRef] [Green Version]
- Dunning, J.B.; Danielson, B.J.; Pulliam, H.R. Ecological processes that affect populations in complex landscapes. Oikos 1992, 65, 169–175. [Google Scholar] [CrossRef] [Green Version]
- Roberge, J.-M.; Angelstam, P.; Villard, M.-A. Specialised woodpeckers and naturalness in hemiboreal forests—Deriving quantitative targets for conservation planning. Biol. Conserv. 2008, 141, 997–1012. [Google Scholar] [CrossRef]
- Brown, J.H. Macroecology; University of Chicago Press: Chicago, IL, USA, 1995; p. 284. [Google Scholar]
- Beck, J.; Ballesteros-Mejia, L.; Buchmann, C.M.; Dengler, J.; Fritz, S.A.; Gruber, B.; Hof, C.; Jansen, F.; Knapp, S.; Kreft, H.; et al. What’s on the horizon for macroecology? Ecography 2012, 35, 673–683. [Google Scholar] [CrossRef]
- Plieninger, T.; Kizos, T.; Bieling, C.; Le Dû-Blayo, L.; Budniok, M.-A.; Bürgi, M.; Crumley, C.L.; Girod, G.; Howard, P.; Kolen, J. Exploring ecosystem-change and society through a landscape lens: Recent progress in European landscape research. Ecol. Soc. 2015, 20, 5. [Google Scholar] [CrossRef] [Green Version]
- Eisenhardt, K.M. Building theories from case study research. Acad. Manag. Rev. 1989, 14, 532–550. [Google Scholar] [CrossRef]
- Flyvbjerg, B. Case Study. In The Sage Handbook of Qualative Research, 4th ed.; Denzin, N.K., Lincoln, Y.S., Eds.; Sage: Thousand Oaks, CA, USA, 2011; pp. 301–316. [Google Scholar]
- Stake, R.E. The Art of Case Study Research; Sage: London, UK, 1995. [Google Scholar]
- Yin, R.K. The case study crisis: Some answers. Adm. Sci. Q. 1981, 26, 58–65. [Google Scholar] [CrossRef] [Green Version]
- Angelstam, P.; Dönz-Breuss, M. Measuring forest biodiversity at the stand scale: An evaluation of indicators in European forest history gradients. Ecol. Bull. 2004, 51, 305–332. [Google Scholar] [CrossRef]
- Puumalainen, J.; Kennedy, P.; Folving, S. Monitoring forest biodiversity: A European perspective with reference to temperate and boreal forest zone. J. Environ. Manag. 2003, 67, 5–14. [Google Scholar] [CrossRef]
- Angelstam, P.; Roberge, J.M.; Lõhmus, A.; Bergmanis, M.; Brazaitis, G.; Dönz-Breuss, M.; Edenius, L.; Kosinski, Z.; Kurlavicius, P.; Lārmanis, V.; et al. Habitat modelling as a tool for landscape-scale conservation: A review of parameters for focal forest birds. Ecol. Bull. 2004, 51, 427–453. [Google Scholar] [CrossRef]
- Åberg, J.; Swenson, J.E.; Angelstam, P. The habitat requirements of hazel grouse (Bonasa bonasia) in managed boreal forest and applicability of forest stand descriptions as a tool to identify suitable patches. For. Ecol. Manag. 2003, 175, 437–444. [Google Scholar] [CrossRef]
- Valasiuk, S.; Giergiczny, M.; Żylicz, T.; Klimkowska, A.; Angelstam, P. Conservation of disappearing cultural landscape’s biodiversity: Are people in Belarus willing to pay for wet grassland restoration? Wetl. Ecol. Manag. 2018, 26, 943–960. [Google Scholar] [CrossRef] [Green Version]
- Waldén, B. Den Stora Sjösänkningen; Aktiebolaget Littorin Rydén boktryckerip: Örebro, Sweden, 1940; p. 366. [Google Scholar]
- Palmqvist, L.A. Bruket Av Våtängarna Vid Nötmyran Västerfärnebo Socken: Ängsbruket Och Bebyggelsen i Hedbo by; Nordiska Museet: Stockholm, Sweden, 1980; p. 60. [Google Scholar]
- Food and Agriculture Organization. FAO Statistical Yearbook 2014; FAO Regional Office for Europe and Central Asia: Budapest, Hungary, 2014. [Google Scholar]
- Berg, Å.; Jonsson, M.; Lindberg, T.; Källebrink, K.-G. Population dynamics and reproduction of Northern Lapwings Vanellus vanellus in a meadow restoration area in central Sweden. IBIS 2002, 144, 131–140. [Google Scholar] [CrossRef]
- International, B. Birds of Europe: Population Estimates, Trends and Conservation Status; BirdLife International: Cambridge, UK, 2004. [Google Scholar]
- Ottosson, U.; Ottvall, R.; Elmberg, J.; Green, M.; Gustafsson, R.; Haas, F.; Holmqvist, N.; Lindström, Å.; Nilsson, L.; Svensson, M.; et al. Fåglarna i Sverige—Antal Och Förekomst; SOF: Halmstad, Sweden, 2012. [Google Scholar]
- Dawson, L.; Elbakidze, M.; Angelstam, P.; Gordon, J. Governance and management dynamics of landscape restoration at multiple scales: Learning from successful environmental managers in Sweden. J. Environ. Manag. 2017, 197, 24–40. [Google Scholar] [CrossRef] [PubMed]
- Bergman, K.-O.; Dániel-Ferreira, J.; Milberg, P.; Öckinger, E.; Westerberg, L. Butterflies in Swedish grasslands benefit from forest and respond to landscape composition at different spatial scales. Landsc. Ecol. 2018, 33, 2189–2204. [Google Scholar] [CrossRef] [Green Version]
- Lindborg, R.; Eriksson, O. Historical landscape connectivity affects present plant species diversity. Ecology 2004, 85, 1840–1845. [Google Scholar] [CrossRef]
- Magnusson, S.-E.; Magntorn, K.; Wallsten, E.; Cronert, H.; Thelaus, M. Kristianstads Vattenrike Biosphere Reserve Nomination Form; Kristianstad Kommun: Kristianstad, Sweden, 2004. [Google Scholar]
- Angelstam, P.; Elbakidze, M. Forest landscape stewardship for functional green infrastructures in Europe’s West and East: Diagnosing and treating social-ecological systems. In The Science and Practice of Landscape Stewardship; Bieling, C., Plieninger, T., Eds.; Cambridge University Press: Cambridge, UK, 2017; pp. 124–144. [Google Scholar]
- Stanevičius, V.; Mačiulis, M.; Švažas, S. Breeding ecology of lapwing (Vanellus vanellus) in floodplains of the Nemunas River delta in 2006–2007. Ekologija 2008, 54, 10–16. [Google Scholar] [CrossRef]
- Švažas, S.; Kozulin, A.; Grishanov, G.; Maximenkov, M.; Rašomavičius, V.; Raudonikis, L.; Baichorov, V.; Skuratovich, A.; Balčiauskas, L.; Drobelis, E.; et al. Important Transboundary Belarusian–Lithuanian and Lithuanian–Russian Wetlands; Akstis: Vilnius, Lithuania, 2003; p. 96. [Google Scholar]
- Kurlavičius, P. Lithuanian breeding bird atlas. In Lithuanian Ornithological Society; Lututė: Švenčionys, Lithuania, 2006. [Google Scholar]
- Thorup, O.; Preiksa, Z.; Pehlak, H.; Altemüller, M.; Drews, H. Status of the Baltic Dunlin Calidris alpina in Lithuania. Wader Study Group Bull. 2011, 118, 184–187. [Google Scholar]
- Pinchuk, P.; Karlionova, N.; Zhurauliou, D. Wader ringing at the Turov ornithological station, Pripyat Valley (S Belarus) in 1996–2003. Ring 2005, 27, 101. [Google Scholar] [CrossRef] [Green Version]
- Delany, S.; Scott, D.; Helmink, A. Waterbird population estimates. Wetl. Int. Glob. Ser. 2006, 12, 226. [Google Scholar]
- Gunnarsson, T.G.; Gill, J.A.; Appleton, G.F.; Gíslason, H.; Gardarsson, A.; Watkinson, A.R.; Sutherland, W.J. Large-scale habitat associations of birds in lowland Iceland: Implications for conservation. Biol. Conserv. 2006, 128, 265–275. [Google Scholar] [CrossRef]
- Gunnarsson, T.G.; Gill, J.A.; Potts, P.M.; Atkinson, P.W.; Croger, R.E.; Gélinaud, G.; Gardarsson, A.; Sutherland, W.J. Estimating population size in Black-tailed Godwits Limosa limosa islandica by colour-marking. Bird Study 2005, 52, 153–158. [Google Scholar] [CrossRef] [Green Version]
- Jóhannesdóttir, L.; Alves, J.A.; Gill, J.A.; Gunnarsson, T.G. Reconciling biodiversity conservation and agricultural expansion in the subarctic environment of Iceland. Ecol. Soc. 2017, 22. [Google Scholar] [CrossRef] [Green Version]
- Robinson, W.D.; Styrsky, J.N.; Brawn, J.D. Are Artificial Bird Nests Effective Surrogates for Estimating Predation on Real Bird Nests? A Test With Tropical Birds. Auk 2005, 122, 843–852. [Google Scholar] [CrossRef]
- Faaborg, J. Truly artificial nest studies. Conserv. Biol. 2004, 18, 369–370. [Google Scholar] [CrossRef]
- Zanette, L. What do artificial nests tells us about nest predation? Biol. Conserv. 2002, 103, 323–329. [Google Scholar] [CrossRef]
- Major, R.E.; Kendal, C.E. The contribution of artificial nest experiments to understanding avian reproductive success: A review of methods and conclusions. IBIS 1996, 138, 298–307. [Google Scholar] [CrossRef]
- Andrén, H.; Angelstam, P. Elevated predation rates as an edge effect in habitat islands: Experimental evidence. Ecology 1988, 69, 544–547. [Google Scholar] [CrossRef]
- Davison, W.B.; Bollinger, E. Predation Rates on Real and Artificial Nests of Grassland Birds. Auk 2000, 117, 147–153. [Google Scholar] [CrossRef]
- Villard, M.-A.; Part, T. Don’t put all your eggs in real nests: A sequel to Faaborg. Conserv. Biol. 2004, 18, 371–372. [Google Scholar] [CrossRef]
- Mandema, F.S.; Tinbergen, J.M.; Ens, B.J.; Bakker, J.P. Livestock grazing and trampling of birds’ nests: An experiment using artificial nests. J. Coast. Conserv. 2013, 17, 409–416. [Google Scholar] [CrossRef] [Green Version]
- Macdonald, M.A.; Bolton, M. Predation on wader nests in Europe. IBIS 2008, 150, 54–73. [Google Scholar] [CrossRef]
- Mason, L.R.; Smart, J.; Drewitt, A.L. Tracking day and night provides insights into the relative importance of different wader chick predators. IBIS 2017, 160, 71–88. [Google Scholar] [CrossRef]
- Green, R.E.; Hawell, J.; Johnson, T.H. Identification of predators of wader eggs from egg remains. Bird Study 1987, 34, 87–91. [Google Scholar] [CrossRef]
- Mayfield, H.F. Suggestions for calculating nest success. Wilson Bull. 1975, 87, 456–466. [Google Scholar]
- Andrén, H. Corvid density and nest predation in relation to forest fragmentation: A landscape perspective. Ecology 1992, 73, 794–804. [Google Scholar] [CrossRef]
- Laidlaw, R.A.; Gunnarsson, T.G.; Méndez, V.; Carneiro, C.; Þórisson, B.; Wentworth, A.; Gill, J.A.; Alves, J.A. Vegetation structure influences predation rates of early nests in subarctic breeding waders. IBIS 2020, 162, 1225–1236. [Google Scholar] [CrossRef] [Green Version]
- Groen, N.M.; Kentie, R.; Goeij, P.d.; Verheijen, B.; Hooijmeijer, J.C.E.W.; Piersma, T. A modern landscape ecology of Black-Tailed Godwits: Habitat selection in Southwest Friesland, the Netherlands. Ardea 2012, 100, 19–28. [Google Scholar] [CrossRef] [Green Version]
- Kentie, R. Spatial Demography of Black-Tailed Godwits: Metapopulation Dynamics in a Fragmented Agricultural Landscape. Ph.D. Thesis, University of Groningen, Groningen, The Netherlands, 2015. [Google Scholar]
- Treinys, R.; Dementavičius, D.; Rumbutis, S.; Švažas, S.; Butkauskas, D.; Sruoga, A.; Dagys, M. Settlement, habitat preference, reproduction, and genetic diversity in recovering the white-tailed eagle Haliaeetus albicilla population. J. Ornithol. 2016, 157, 311–323. [Google Scholar] [CrossRef]
- Hipfner, M.J.; Blight, L.K.; Lowe, R.W.; Wilhelm, S.I.; Robertson, G.J.; Barrett, R.T.; Anker-Nilssen, T.; Good, T.P. Unintended consequences: How the recovery of sea eagle Haliaeetus spp. populations in the northern hemisphere is affecting seabirds. Mar. Ornithol. 2012, 40, 39–52. [Google Scholar]
- Dawson, L.; Elbakidze, M.; Schellensac, M.; Shkarubad, A.; Angelstam, P. Bogs, Birds and Berries in Belarus: The governance and management dynamics of wetland restoration in a state-centric, top-down context. Ecol. Soc. 2021, 26, 8. [Google Scholar] [CrossRef]
- Teunissen, W.; Schekkerman, H.; Willems, F.; Majoor, F. Identifying predators of eggs and chicks of Lapwing Vanellus vanellus and Black-tailed Godwit Limosa limosa in the Netherlands and the importance of predation on wader reproductive output. IBIS 2008, 150, 74–85. [Google Scholar] [CrossRef]
- Laidlaw, R.A.; Smart, J.; Smart, M.A.; Gill, J.A. Managing a food web: Impacts on small mammals of managing grasslands for breeding waders. Anim. Conserv. 2013, 16, 207–215. [Google Scholar] [CrossRef]
- Rickenbach, O.; Grüebler, M.U.; Schaub, M.; Koller, A.; Naef-Daenzer, B.; Schifferli, L. Exclusion of ground predators improves Northern Lapwing Vanellus vanellus chick survival. IBIS 2011, 153, 531–542. [Google Scholar] [CrossRef]
- Laidlaw, R.A.; Smart, J.; Smart, M.A.; Gill, J.A. The influence of landscape features on nest predation rates of grassland-breeding waders. IBIS 2015, 157, 700–712. [Google Scholar] [CrossRef] [Green Version]
- Jackson, D.B. Experimental removal of introduced hedgehogs improves wader nest success in the Western Isles, Scotland. J. Appl. Ecol. 2001, 38, 802–812. [Google Scholar] [CrossRef]
- Wilson, G.R.; Brittingham, M.C.; Goodrich, L.J. How Well do Artificial Nests Estimate Success of Real Nests? Condor 1998, 100, 357–364. [Google Scholar] [CrossRef] [Green Version]
- Fuller, R.J. Avian Responses to Transitional Habitats in Temperate Cultural Landscapes: Woodland Edges and Young-Growth. Birds and Habitat. Relationships in Changing Landscapes; Fuller, R.J., Ed.; Cambridge University Press: Cambridge, UK, 2012. [Google Scholar]
- Berg, Å.; Wretenberg, J.; Żmihorski, M.; Hiron, M.; Pärt, T. Linking occurrence and changes in local abundance of farmland bird species to landscape composition and land-use changes. Agric. Ecosyst. Environ. 2015, 204, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Żmihorski, M.; Krupiński, D.; Kotowska, D.; Knape, J.; Pärt, T.; Obłoza, P.; Berg, Å. Habitat characteristics associated with occupancy of declining waders in Polish wet grasslands. Agric. Ecosyst. Environ. 2018, 251, 236–243. [Google Scholar] [CrossRef]
- Rönkä, A. Distribution, status and population trends in the Temminck’s Stint Calidris temminckii in the Finnish Bothnian Bay. Ornis Fenn. 1996, 73, 1–11. [Google Scholar]
- Koivula, K.; Rönkä, A. Habitat deterioration and efficiency of antipredator strategy in a meadow-breeding wader, Temminck’s stint (Calidris temminckii). Oecologia 1998, 116, 348–355. [Google Scholar] [CrossRef]
- Tucker, G.M.; Davies, S.; Fuller, R. The Ecology and Conservation of Lapwings Vanellus Vanellus; Joint Nature Conservation Committee: Peterborough, UK, 1994.
- Seymour, A.S.; Harris, S.; Ralston, C.; White, P.C.L. Factors influencing the nesting success of Lapwings Vanellus vanellus and behaviour of Red Fox Vulpes vulpes in Lapwing nesting sites. Bird Study 2003, 50, 39–46. [Google Scholar] [CrossRef] [Green Version]
- Lank, D.B.; Butler, R.W.; Ireland, J.; Ydenberg, R.C. Effects of predation danger on migration strategies of sandpipers. Oikos 2003, 103, 303–319. [Google Scholar] [CrossRef] [Green Version]
- Červinka, J.; Šálek, M.; Pavluvčík, P.; Kreisinger, J. The fine-scale utilization of forest edges by mammalian mesopredators related to patch size and conservation issues in Central European farmland. Biodivers. Conserv. 2011, 20, 3459–3475. [Google Scholar] [CrossRef]
- Meltofte, H.; Amstrup, O. Forekomsten af rovfugle på Tipperne 1930-2011. Dan. Orn. Foren. Tidsskr. 2013, 107, 229–238. [Google Scholar]
- Herzon, I.; Raatikainen, K.J.; Wehn, S.; Rūsiņa, S.; Helm, A.; Cousins, S.A.O.; Rašomavičius, V. Semi-natural habitats in boreal Europe: A rise of a social-ecological research agenda. Ecol. Soc. 2021, 26. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Case Study Landscapes | Kristianstad | Östergötland | Mälardalen | Nemunas Delta | Turov | Akureyri |
|---|---|---|---|---|---|---|
| Country | Sweden | Sweden | Sweden | Lithuania | Belarus | Iceland |
| Development index | 5 | 4 | 4 | 3 | 2 | 1 |
| Trend in grassland habitat | Strong decline [49] | Decline [21] | Decline [64,65] | Slight decline [66] | Stable [66] | Increase [66] |
| Wader trends | Negative [20] | Negative [21] | Negative [67] | Declining [68] | Stable [68] | Increase [15] |
| # of wet grassland patches | 21 | 22 | 27 | 37 | 30 | 31 |
| Total area (ha) | 1288 | 726 | 1486 | 2698 | 2135 | 1196 |
| Mean Patch size (ha) | 61 | 33 | 55 | 73 | 71 | 39 |
| Patch size range (ha) | 15–168 | 6–153 | 18–134 | 24–133 | 23–219 | 7–174 |
| Standard Deviation | 36 | 33 | 32 | 29 | 43 | 34 |
| Case Study Landscapes | Kristianstad | Östergötland | Mälardalen | Nemunas Delta | Turov | Akureyri |
|---|---|---|---|---|---|---|
| Waders | 14.33 | 23.95 | 27.04 | 28.08 | 62.80 | 35.6 |
| Corvids | 25.00 | 5.77 | 9.00 | 0.97 | 1.30 | 0.35 |
| Raptors | 2.48 | 1.18 | 0.67 | 0.41 | 0.30 | 0.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manton, M.; Angelstam, P. Macroecology of North European Wet Grassland Landscapes: Habitat Quality, Waders, Avian Predators and Nest Predation. Sustainability 2021, 13, 8138. https://doi.org/10.3390/su13158138
Manton M, Angelstam P. Macroecology of North European Wet Grassland Landscapes: Habitat Quality, Waders, Avian Predators and Nest Predation. Sustainability. 2021; 13(15):8138. https://doi.org/10.3390/su13158138
Chicago/Turabian StyleManton, Michael, and Per Angelstam. 2021. "Macroecology of North European Wet Grassland Landscapes: Habitat Quality, Waders, Avian Predators and Nest Predation" Sustainability 13, no. 15: 8138. https://doi.org/10.3390/su13158138