Selection Criteria for Drought-Tolerant Bread Wheat Genotypes at Seedling Stage

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
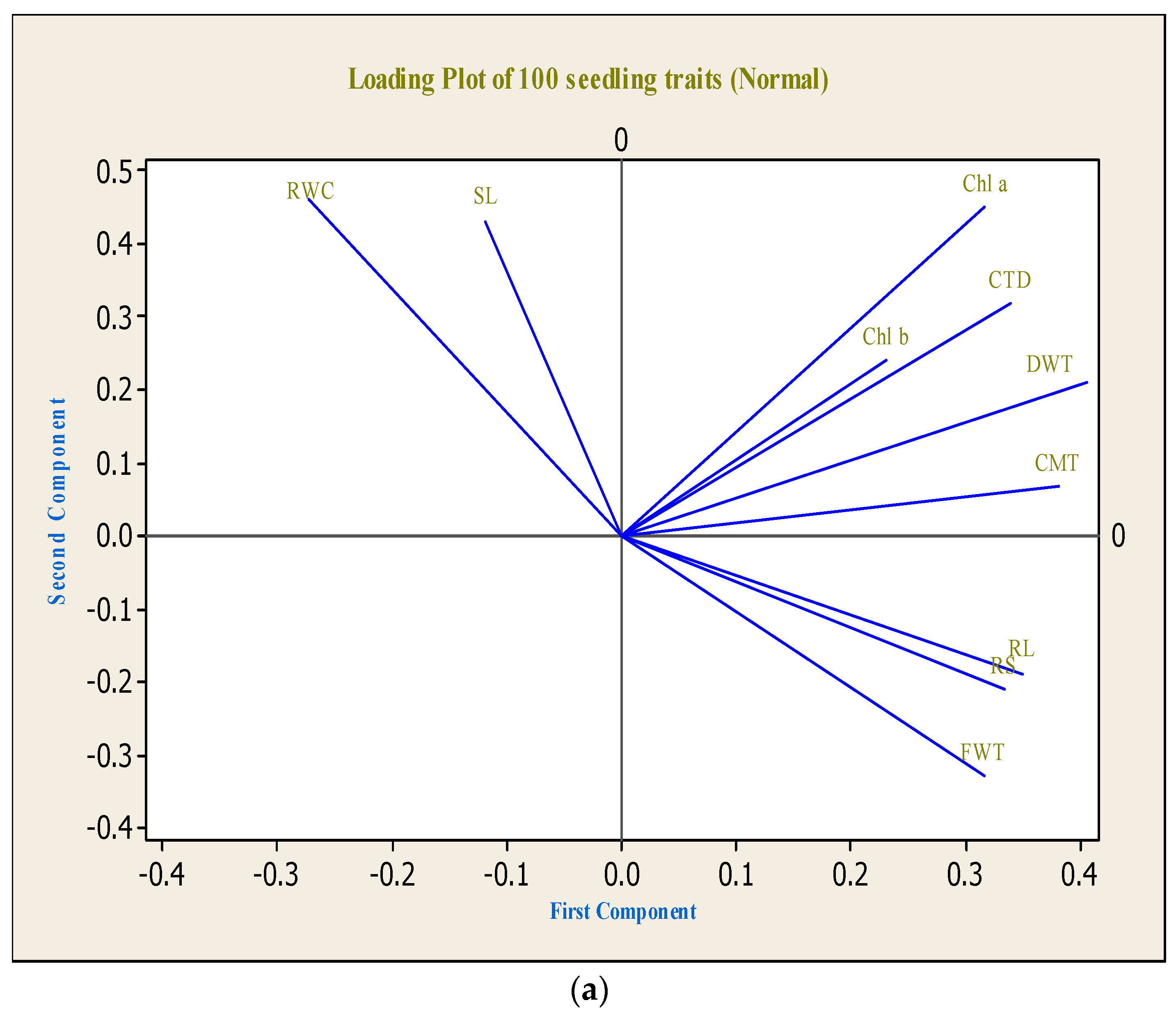
3. Results and Discussion
3.1. Mean Variability
3.2. Correlation Analysis
3.3. Principal Component Analysis (PCA)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Dixon, J.; Braun, H.-J.; Kosina, P.; Crouch, J.H. Wheat Facts and Futures 2009; Cimmyt: Mexico City, Mexico, 2009. [Google Scholar]
- Ahmed, H.G.M.-D.; Khan, A.S.; Kashif, M.; Khan, S. Genetic analysis of yield and physical traits of spring wheat grain. J. Natl. Sci. Found. Sri Lanka 2018, 46, 23–30. [Google Scholar] [CrossRef]
- Abbas, M.; Sheikh, A.; Sabir, H.M.; Nighat, S. Factors responsible for low wheat productivity in Central Punjab. Pak. J. Agric. Sci. 2005, 42, 3–4. [Google Scholar]
- Bashir, A.; Mahmood, M.A.; Sheikh, A.; Kashif, M. Causes of wheat yield decline in the irrigated Punjab. J. Agric. Res. 2006, 44, 71–83. [Google Scholar]
- Yagdi, E.A.C.K. Study of genetic diversity in wheat (Triticum aestivum) varities using Random Amplified Polymorphic DNA (RAPD) analysis. Turk. J. Field Crops 2012, 17, 91–95. [Google Scholar]
- Toker, C.; Canci, H.; Yildirim, T. Evaluation of perennial wild Cicer species for drought resistance. Genet. Resour. Crop Evol. 2007, 54, 1781–1786. [Google Scholar] [CrossRef]
- Ali, A.; Ali, N.; Ullah, N.; Ullah, F.; Adnan, M.; Ahmed, Z. Effect of drought stress on the physiology and yield of the Pakistani wheat germplasms. Int. J. Adv. Res. Technol. 2013, 2, 419–430. [Google Scholar]
- Batool, A.; Noorka, I.R.; Afzal, M.; Syed, A.H. Estimation of heterosis, heterobeltiosis and potence ratio over environments among pre and post Green Revolution Spring wheat in Pakistan. J. Basic Appl. Sci. 2013, 9, 36–43. [Google Scholar]
- Mir, R.R.; Zaman-Allah, M.; Sreenivasulu, N.; Trethowan, R.; Varshney, R.K. Integrated genomics, physiology and breeding approaches for improving drought tolerance in crops. Theor. Appl. Genet. 2012, 125, 625–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, I.; Khaliq, I.; Khan, A.S.; Farooq, M. Screening of spring wheat (Triticum aestivum L.) genotypes for drought tolerance on the basis of seedling traits. Pak. J. Agric. Sci. 2014, 51, 367–372. [Google Scholar]
- Ahmed, H.; Khan, A.S.; Khan, S.H.; Kashif, M. Genome wide allelic pattern and genetic diversity of spring wheat genotypes through SSR markers. Int. J. Agric. Biol. 2017, 19, 1559–1565. [Google Scholar]
- Ashfaq, W.; Ul-Allah, S.; Kashif, M.; Sattar, A.; Nabi, H.G. Genetic variability study among wheat genotypes under normal and drought conditions. J. Glob. Innov. Agric. Soc. Sci. 2016, 4, 111–116. [Google Scholar] [CrossRef]
- Noorka, I.R.; Batool, A.; Rauf, S.; Teixeira da Silva, J.; Ashraf, E. Estimation of heterosis in wheat (Triticum aestivum L.) under contrasting water regimes. Int. J. Plant Breed. 2013, 7, 55–60. [Google Scholar]
- Zhang, H.; Wang, H. Evaluation of drought tolerance from a wheat recombination inbred line population at the early seedling growth stage. Afr. J. Agric. Res. 2012, 7, 6167–6172. [Google Scholar]
- Shahbazi, H.; Bihamta, M.R.; Taeb, M.; Darvish, F. Germination characters of wheat under osmotic stress: Heritability and relation with drought tolerance. Int. J. Agric. Res. Rev. 2012, 2, 689–698. [Google Scholar]
- Bilal, M.; Rashid, R.; Rehman, S.; Iqbal, F.; Ahmed, J.; Abid, M.; Ahmed, Z.; Hayat, A. Evaluation of wheat genotypes for drought tolerance. J. Green Physiol. Genet. Genom. 2015, 1, 11–21. [Google Scholar]
- Almeselmani, M.; Abdullah, F.; Hareri, F.; Naaesan, M.; Ammar, M.A.; ZuherKanbar, O.; Saud, A.A. Effect of drought on different physiological characters and yield component in different varieties of Syrian durum wheat. J. Agric. Sci. 2011, 3, 127. [Google Scholar] [CrossRef]
- Arjenaki, F.G.; Jabbari, R.; Morshedi, A. Evaluation of drought stress on relative water content, chlorophyll content and mineral elements of wheat (Triticum aestivum L.) varieties. Int. J. Agric. Crop Sci. 2012, 4, 726–729. [Google Scholar]
- Datta, J.; Mondal, T.; Banerjee, A.; Mondal, N. Assessment of drought tolerance of selected wheat cultivars under laboratory condition. J. Agric. Technol. 2011, 7, 383–393. [Google Scholar]
- Jaleel, C.A.; Manivannan, P.; Wahid, A.; Farooq, M.; Al-Juburi, H.J.; Somasundaram, R.; Panneerselvam, R. Drought stress in plants: A review on morphological characteristics and pigments composition. Int. J. Agric. Biol. 2009, 11, 100–105. [Google Scholar]
- Anjum, S.A.; Xie, X.-Y.; Wang, L.-C.; Saleem, M.F.; Man, C.; Lei, W. Morphological, physiological and biochemical responses of plants to drought stress. Afr. J. Agric. Res. 2011, 6, 2026–2032. [Google Scholar]
- Bojović, B.; Stojanović, J. Chlorophyll and carotenoid content in wheat cultivars as a function of mineral nutrition. Arch. Biol. Sci. 2005, 57, 283–290. [Google Scholar] [CrossRef] [Green Version]
- Gugino, B.K.; Abawi, G.S.; Idowu, O.J.; Schindelbeck, R.R.; Smith, L.L.; Thies, J.E.; Wolfe, D.W.; Van Es, H.M. Cornell Soil Health Assessment Training Manual; Cornell University College of Agriculture and Life Sciences: Ithaca, NY, USA, 2009. [Google Scholar]
- Moebius-Clune, B.; Moebius-Clune, D.; Gugino, B.; Idowu, O.; Schindelbeck, R.; Ristow, A. Comprehensive Assessment of Soil Health: The Cornell Framework Manual, Edition 3.1; Cornell University: Ithaca, NY, USA, 2016. [Google Scholar]
- Dhanda, S.; Sethi, G. Inheritance of excised-leaf water loss and relative water content in bread wheat (Triticum aestivum). Euphytica 1998, 104, 39–47. [Google Scholar] [CrossRef]
- Blum, A.; Ebercon, A. Cell membrane stability as a measure of drought and heat tolerance in wheat 1. Crop Sci. 1981, 21, 43–47. [Google Scholar] [CrossRef]
- Lohithaswa, H.; Desai, S.; Hanchinal, R.; Patil, B.; Math, K.; Kalappanavar, I.; Bandivadder, T.; Chandrashekhara, C. Combining ability in tetraploid wheat for yield, yield attributing traits, quality and rust resistance over environments. Karnataka J. Agric. Sci. 2014, 26, 190–193. [Google Scholar]
- Röbbelen, G. Untersuchungen an strahleninduzierten Blattfarbmutanten von Arabidopsis thaliana (L) Heynh. Zeitschrift für Induktive Abstammungs- und Vererbungslehre 1957, 88, 189–252. [Google Scholar]
- Steel, R.G.D.; Torrie, J.H. Principles and Procedures of Statistics, a Biometrical Approach; McGraw-Hill Kogakusha: Tokyo, Japan, 1980. [Google Scholar]
- Ogunbayo, S.; Ojo, D.; Guei, R.; Oyelakin, O.; Sanni, K.L. Phylogenetic diversity and relationships among 40 rice accessions using morphological and RAPDs techniques. Afr. J. Biotechnol. 2005, 4, 1234–1244. [Google Scholar]
- Leishman, M.R.; Westoby, M. The role of seed size in seedling establishment in dry soil conditions—Experimental evidence from semi-arid species. J. Ecol. 1994, 82, 249–258. [Google Scholar] [CrossRef]
- Dhanda, S.; Sethi, G.; Behl, R. Indices of drought tolerance in wheat genotypes at early stages of plant growth. J. Agron. Crop Sci. 2004, 190, 6–12. [Google Scholar] [CrossRef]
- Tuberosa, R.; Salvi, S. Genomics-based approaches to improve drought tolerance of crops. Trends Plant Sci. 2006, 11, 405–412. [Google Scholar] [CrossRef]
- De Dorlodot, S.; Forster, B.; Pagès, L.; Price, A.; Tuberosa, R.; Draye, X. Root system architecture: opportunities and constraints for genetic improvement of crops. Trends Plant Sci. 2007, 12, 474–481. [Google Scholar] [CrossRef] [PubMed]
- Sehgal, D.; Skot, L.; Singh, R.; Srivastava, R.K.; Das, S.P.; Taunk, J.; Sharma, P.C.; Pal, R.; Raj, B.; Hash, C.T. Exploring potential of pearl millet germplasm association panel for association mapping of drought tolerance traits. PLoS ONE 2015, 10, e0122165. [Google Scholar] [CrossRef]
- Ahmad, M.; Shabbir, G.; Minhas, N.; Shah, M.K.N. Identification of drought tolerant wheat genotypes based on seedling traits. Sarhad J. Agric. 2013, 29, 21–27. [Google Scholar]
- Faisal, S.; Mujtaba, S.; Khan, M.; Mahboob, W. Morpho-Physiological assessment of wheat (Triticum aestivum L.) genotypes for drought stress tolerance at seedling stage. Pak. J. Bot. 2017, 49, 445–452. [Google Scholar]
- Foito, A.; Byrne, S.L.; Shepherd, T.; Stewart, D.; Barth, S. Transcriptional and metabolic profiles of Lolium perenne L. genotypes in response to a PEG—Induced water stress. Plant Biotechnol. J. 2009, 7, 719–732. [Google Scholar] [CrossRef] [PubMed]
- Khodarahmpour, Z. Effect of drought stress induced by polyethylene glycol (PEG) on germination indices in corn (Zea mays L.) hybrids. Afr. J. Biotechnol. 2011, 10, 18222–18227. [Google Scholar] [CrossRef]
- Soltani, A.; Gholipoor, M.; Zeinali, E. Seed reserve utilization and seedling growth of wheat as affected by drought and salinity. Environ. Exp. Bot. 2006, 55, 195–200. [Google Scholar] [CrossRef]
- Sassi, K.; Abid, G.; Jemni, L.; Dridi-Al-Mohandes, B.; Boubaker, M. Comparative study of six varieties of durum wheat (Triticum durum Desf.) Vis-a-vis water stress. JAPS 2012, 15, 2157–2170. [Google Scholar]
- Kamran, M.; Shahbaz, M.; Ashraf, M.; Akram, N.A. Alleviation of drought-induced adverse effects in spring wheat (Triticum aestivum L.) using proline as a pre-sowing seed treatment. Pak. J. Bot. 2009, 41, 621–632. [Google Scholar]
- Marcińska, I.; Czyczyło-Mysza, I.; Skrzypek, E.; Filek, M.; Grzesiak, S.; Grzesiak, M.T.; Janowiak, F.; Hura, T.; Dziurka, M.; Dziurka, K. Impact of osmotic stress on physiological and biochemical characteristics in drought-susceptible and drought-resistant wheat genotypes. Acta Physiol. Planta. 2013, 35, 451–461. [Google Scholar] [CrossRef]
- Esfandiari, E.; Shakiba, M.R.; Mahboob, S.A.; Alyari, H.; Toorchi, M. Water stress, antioxidant enzyme activity and lipid peroxidation in wheat seedling. J. Food Agric. Environ. 2007, 5, 149–153. [Google Scholar]
- Selote, D.S.; Bharti, S.; Khanna-Chopra, R. Drought acclimation reduces O2− accumulation and lipid peroxidation in wheat seedlings. Biochem. Biophys. Res. Commun. 2004, 314, 724–729. [Google Scholar] [CrossRef] [PubMed]
- Subrahmanyam, D.; Subash, N.; Haris, A.; Sikka, A. Influence of water stress on leaf photosynthetic characteristics in wheat cultivars differing in their susceptibility to drought. Photosynthetica 2006, 44, 125–129. [Google Scholar] [CrossRef]
- Pour-Aboughadareh, A.; Ahmadi, J.; Mehrabi, A.A.; Etminan, A.; Moghaddam, M.; Siddique, K.H. Physiological responses to drought stress in wild relatives of wheat: Implications for wheat improvement. Acta Physiol. Planta 2017, 39, 106–114. [Google Scholar] [CrossRef]
- Ramani, H.; Mandavia, M.; Dave, R.; Bambharolia, R.; Silungwe, H.; Garaniya, N. Biochemical and physiological constituents and their correlation in wheat (Triticum aestivum L.) genotypes under high temperature at different development stages. Int. J. Plant Physiol. Biochem. 2017, 9, 1–8. [Google Scholar]
- Keyvan, S. The effects of drought stress on yield, relative water content, proline, soluble carbohydrates and chlorophyll of bread wheat cultivars. J. Anim. Plant Sci. 2010, 8, 1051–1060. [Google Scholar]
- Van Heerden, P.; De Villiers, O. Evaluation of the relative water content and the reduction of 2, 3, 5-triphenyltetrazoliumchloride as indicators of drought tolerance in spring wheat cultivars. S. Afr. J. Plant Soil 1996, 13, 131–135. [Google Scholar] [CrossRef]
- Ritchie, S.W.; Nguyen, H.T.; Holaday, A.S. Leaf water content and gas-exchange parameters of two wheat genotypes differing in drought resistance. Crop Sci. 1990, 30, 105–111. [Google Scholar] [CrossRef]
- Ahmadizadeh, M. Physiological and agro-morphological response to drought stress. MiddleEast J. Sci. Res. 2013, 13, 998–1009. [Google Scholar]
- Hussain, M.; Askandar, H.S.; Hassan, Z.A. Selecting high yielding wheat hybrids from a restricted factorial mating design. Sarhad J. Agric. 2013, 29, 173–179. [Google Scholar]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Łukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Planta 2016, 38, 102. [Google Scholar] [CrossRef]
- Bala, P.; Sikder, S. Evaluation of Heat Tolerance of Wheat Genotypes Through Membrane Thermostability Test. MAYFEB J. Agric. Sci. 2017, 2, 1–6. [Google Scholar]
- Zhang, M.; Lv, D.; Ge, P.; Bian, Y.; Chen, G.; Zhu, G.; Li, X.; Yan, Y. Phosphoproteome analysis reveals new drought response and defense mechanisms of seedling leaves in bread wheat (Triticum aestivum L.). J. Proteomics 2014, 109, 290–308. [Google Scholar] [CrossRef]
- Epée Missé, P.T. Wheat Seedling Physiological Adaptation to Overcome Water Stress. SSRN 2018, 2–11. [Google Scholar] [CrossRef]
- Farshadfar, E.; Amiri, R. Genetic analysis of physiological indicators of drought tolerance in bread wheat using diallel technique. Genetika 2015, 47, 107–118. [Google Scholar] [CrossRef]
- Saeidi, M.; Zabihi-e-Mahmoodabad, R. Evaluation of drought stress on relative water content and chlorophyll content of sesame (Sesamum indicum L.) genotypes at early flowering stage. Res. J. Environ. Sci. 2009, 3, 345–350. [Google Scholar]
- Khalilzadeh, R.; Seyed Sharifi, R.; Jalilian, J. Antioxidant status and physiological responses of wheat (Triticum aestivum L.) to cycocel application and bio fertilizers under water limitation condition. J. Plant Interact. 2016, 11, 130–137. [Google Scholar] [CrossRef]
- Schlemmer, M.R.; Francis, D.D.; Shanahan, J.; Schepers, J.S. Remotely measuring chlorophyll content in corn leaves with differing nitrogen levels and relative water content. Agron. J. 2005, 97, 106–112. [Google Scholar] [CrossRef]
- Livingston, D.P.; Hincha, D.K.; Heyer, A.G. Fructan and its relationship to abiotic stress tolerance in plants. Cell. Mol. Life Sci. 2009, 66, 2007–2023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Liu, B.; An, G.; Gao, X. Morpho-physiological responses of Alhagi sparsifolia Shap.(leguminosae) seedlings to progressive drought stress. Pak. J. Bot. 2016, 48, 429–438. [Google Scholar]
- Ghafoor, G.; Hassan, G.; Ahmad, I.; Khan, S.N.; Suliman, S. Correlation analysis for different parameters of F2 bread wheat population. Pure Appl. Biol. 2013, 2, 28–31. [Google Scholar] [CrossRef]
- Ali, Y.; Atta, B.M.; Akhter, J.; Monneveux, P.; Lateef, Z. Genetic variability, association and diversity studies in wheat (Triticum aestivum L.) germplasm. Pak. J. Bot. 2008, 40, 2087–2097. [Google Scholar]
- Khan, M.; Shabbir, G.; Akram, Z.; Shah, M.; Ansar, M.; Cheema, N. Character association studies of seedling traits in different wheat genotypes under moisture stress conditions. SABRAO J. Breed. Genet. 2011, 45, 458–467. [Google Scholar]
- Saidi, A.; Ookawa, T.; Hirasawa, T. Responses of root growth to moderate soil water deficit in wheat seedlings. Plant Prod. Sci. 2010, 13, 261–268. [Google Scholar] [CrossRef]
- Soleimani, Z.; Ramshini, H.; Mortazavian, S.M.M.; Fazelnajafabadi, M.; Foughi, B. Screening for drought tolerance in Iranian wheat genotypes (Triticum aestivumL.) using physiological traits evaluated under drought stress and normal condition. Aust. J. Crop Sci. 2014, 8, 200–208. [Google Scholar]
- Naroui Rad, M.R.; Kadir, M.A.; Yusop, M.R. Genetic behaviour for plant capacity to produce chlorophyll in wheat (Triticum aestivum) under drought stress. Aust. J. Crop Sci. 2012, 6, 415. [Google Scholar]
- Oraki, H.; Aghaalikhana, M. Effect of water deficit stress on proline contents, soluble sugars, chlorophyll and grain yield of sunflower (Helianthus annuus L.) hybrids. Afr. J. Biotechnol. 2012, 11, 164–168. [Google Scholar]
- Kaiser, H.F. The application of electronic computers to factor analysis. Educ. Psychol. Meas. 1960, 20, 141–151. [Google Scholar] [CrossRef]
- El-Deeb, A.; Mohamed, N. Factor and cluster analysis for some quantitative characters in sesame (Sesamum indicum L.). In Proceedings of the Annual Conference ISSR, Cairo University, Cairo, Egypt, 4–6 December 1999; Volume 34. [Google Scholar]
- Jaynes, D.; Kaspar, T.; Colvin, T.; James, D. Cluster analysis of spatiotemporal corn yield patterns in an Iowa field. Agron. J. 2003, 95, 574–586. [Google Scholar] [CrossRef]
- Sisodia, B.; Rai, V. An application of principal component analysis for pre-harvest forecast model for wheat crop based on biometrical characters. Int. Res. J. Agric. Econ. Stat. 2017, 8, 83–87. [Google Scholar]
- Ali, M.; Zulkiffal, M.; Anwar, J.; Hussain, M.; Farooq, J.; Khan, S. Morpho-physiological diversity in advanced lines of bread wheat under drought conditions at post-anthesis stage. J. Anim. Plant Sci. 2015, 25, 431–441. [Google Scholar]
- Wang, J.; Li, S.; Li, K. Effects of water limited deficit stress during different growth stages on leaf protective enzymes of winter wheat. Acta Bot. Boreali Occident. Sin. 2001, 21, 47–52. [Google Scholar]
- Khodadadi, M.; Fotokian, M.H.; Miransari, M. Genetic diversity of wheat (Triticum aestivum L.) genotypes based on cluster and principal component analyses for breeding strategies. Aust. J. Crop Sci. 2011, 5, 17–24. [Google Scholar]
- Saima, G.; Khan, S.; Munawar, S.; Muhammad, A.; Muhammad, S. Genetic evaluation of spring wheat (Triticum aestivum) germplasm for yield and seedling vigor traits. J. Agric. Soc. Sci. 2012, 8, 123–128. [Google Scholar]
- Sajjad, M.; Khan, S.; Khan, A.S. Exploitation of germplasm for grain yield improvement in spring wheat (Triticum aestivum). Int. J. Agric. Biol. 2011, 13, 695–700. [Google Scholar]
- Bowne, J.B.; Erwin, T.A.; Juttner, J.; Schnurbusch, T.; Langridge, P.; Bacic, A.; Roessner, U. Drought responses of leaf tissues from wheat cultivars of differing drought tolerance at the metabolite level. Mol. Plant 2012, 5, 418–429. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Genotype(G) | Conditions(C) | G×C | Error | Total | |
|---|---|---|---|---|---|
| SoV/df | 104 | 1 | 104 | 420 | 629 |
| RL (cm) | 61.15** | 1749.07 ** | 6.86 ** | 3.13 | |
| SL (cm) | 39.51 ** | 3792.76 ** | 8.55 ** | 2.1 | |
| R/S | 0.12351 ** | 0.29618 ** | 0.01561 ** | 0.00841 | |
| FWT (g) | 0.0185 ** | 18.6482 ** | 0.0010 * | 0.0008 | |
| DWT (g) | 0.01035 ** | 2.07891 ** | 0.00253 ** | 0.00057 | |
| RWC | 97.9 ** | 14377.6 ** | 1.9 ns | 5.6 | |
| CMT | 85.2 ** | 17463.7 ** | 0.4 ns | 3.4 | |
| CTD (mg/g Fw) | 0.00669 ** | 0.77175** | 0.00010 ns | 0.00062 | |
| Chl.a(mg/g Fw) | 0.02483 ** | 0.01939 ** | 0.00074 ns | 0.00151 | |
| Chl.b(mg/g Fw) | 0.00701 ** | 0.76442 | 0.00007 ** | 0.0005 |
| Traits | Conditions | Minimum | Maximum | Mean | SD |
|---|---|---|---|---|---|
| RL (cm) | Normal | 7.22 | 23.00 | 11.66 | 4.005 |
| Drought | 5.31 | 17.65 | 8.32 | 2.542 | |
| SL(cm) | Normal | 13.16 | 29.17 | 22.69 | 2.965 |
| Drought | 11.00 | 25.19 | 17.79 | 2.755 | |
| R/S | Normal | 0.27 | 0.94 | 0.51 | 0.170 |
| Drought | 0.29 | 0.92 | 0.47 | 0.132 | |
| FWT (g) | Normal | 0.75 | 1.11 | 0.94 | 0.048 |
| Drought | 0.48 | 0.91 | 0.59 | 0.065 | |
| DWT(g) | Normal | 0.18 | 0.49 | 0.26 | 0.059 |
| Drought | 0.10 | 0.27 | 0.15 | 0.028 | |
| RWC (%) | Normal | 61.00 | 82.33 | 71.22 | 4.099 |
| Drought | 51.45 | 72.78 | 61.67 | 4.099 | |
| CMT (%) | Normal | 55.00 | 74.00 | 64.13 | 3.789 |
| Drought | 44.47 | 63.47 | 53.60 | 3.789 | |
| CTD (mg/g Fw) | Normal | 0.32 | 0.49 | 0.40 | 0.034 |
| Drought | 0.24 | 0.41 | 0.33 | 0.034 | |
| Chl.a (mg/g Fw) | Normal | 1.36 | 1.65 | 1.48 | 0.066 |
| Drought | 1.34 | 1.61 | 1.47 | 0.065 | |
| Chl.b (mg/g Fw) | Normal | 0.46 | 0.64 | 0.54 | 0.034 |
| Drought | 0.39 | 0.57 | 0.47 | 0.034 |
| Traits | Best-Performing Genotypes: Names and Mean Values (Drought-Tolerant) | Worst-Performing Genotypes: Names and Mean Values (Drought-Susceptible) |
|---|---|---|
| SL (cm) | 9508 (25.19) followed by Sunstar (23.86) | BARS-2009 (11) followed by MOOMAL-2002 (12) |
| RL (cm) | CHAKWAL-86 (17.65) followed by 10117 (15.65) and DPW-621-50 (14) | MOOMAL-2002 (4.98) followed by BARS-2009 (5.31) and MILLAT-2011(5.33) |
| R/S | CHAKWAL-86 (0.92) followed by 10117 (0.90) and BWL-812 (0.71) | 9508(0.29) followed by UFAQ-2002(0.30) and G40 (0.31) |
| FWT (g) | 9797 (0.91) followed by 9493(0.83) and 9930 (0.78) | MOOMAL-2002 (0.48) followed by UFAQ-2002(0.48) and BARS-2009 (0.49) |
| DWT (g) | 9493(0.27) followed by9797 (0.21) and 10115(0.20) | BARS-2009 (0.10) followed by UFAQ-2002(0.11) and MILLAT-2011(0.11) |
| RWC (%) | 9618 (72.78) followed by 10111 (70.45) and 10117 (69.78) | BARS-2009 (51.45) followed by UFAQ-2002(54.12) and MOOMAL-2002 (54.43) |
| CMT (%) | 9493(63.47) followed by G11 (62.47) and CHAKWAL-86 (59.47) | 9508(44.47) followed by MILLAT-2011(49.14) and MILLAT-2011(49.14) |
| CTD (mg/g Fw) | 9618 (0.41) followed by BWL-812 (0.40) and 9930 (0.39) | MILLAT-2011(0.24) followed by MOOMAL-2002 (0.25) and UFAQ-2002(0.27) |
| Chl.a (mg/g Fw) | CHAKWAL-86 (1.61) followed by 10115(1.60) and 9493(1.59) | BARS-2009 (1.34) followed by MILLAT-2011(1.37) and UFAQ-2002(1.38) |
| Chl.b (mg/g Fw) | CHAKWAL-86 (0.57) followed by BWL-812 (0.55) and 9618 (0.54) | MOOMAL-2002 (0.39) followed by BARS-2009 (0.40) and 9508(0.42) |
| Traits | RL | SL | RS | FWT | DWT | RWC | CMT | CTD | Chl a | |
|---|---|---|---|---|---|---|---|---|---|---|
| SL | N | 0.04 ns | ||||||||
| D | −0.19 ns | |||||||||
| RS | N | −0.01 ns | −0.17 ns | |||||||
| D | 0.93 ** | −0.20 * | ||||||||
| FWT | N | 0.19 * | −0.14 ns | 0.15m ns | ||||||
| D | 0.48 ** | −0.18 ns | 0.41 ** | |||||||
| DWT | N | 0.33 ** | −0.16 ns | 0.03 ns | 0.88 ** | |||||
| D | 0.62 ** | −0.05 ns | 0.53 ** | 0.52 ** | ||||||
| RWC | N | 0.30 ** | −0.19 ns | −0.01 ns | 0.86 ** | 0.92 ** | ||||
| D | −0.33 ** | 0.39 ** | −0.30 * | −0.70 ** | −0.33 ** | |||||
| CMT | N | 0.26 ** | −0.17 ns | −0.02 ns | 0.86 ** | 0.87 ** | 0.92 ** | |||
| D | 0.49 ** | −0.18 ns | 0.48 ** | 0.47 ** | 0.68 ** | 0.50 ** | ||||
| CTD | N | −0.48 ** | 0.16 ns | 0.32 ** | 0.00 ns | −0.15 ns | −0.24 ns | −0.22 * | ||
| D | 0.47 ** | −0.04 ns | 0.41 ** | 0.34 ** | 0.71 ** | −0.18 ns | 0.55 ** | |||
| Chl.a | N | 0.14 ns | 0.02 ns | −0.07 ns | 0.49 ** | 0.65 ** | 0.63 ** | 0.68 ** | −0.20 * | |
| D | 0.29 ** | 0.007 ns | 0.27 ** | 0.28 ** | 0.69 ** | −0.14 ns | 0.62 ** | 0.58 ** | ||
| Chl.b | N | 0.07 ns | 0.03 ns | 0.002 ns | 0.49 ** | 0.63 ** | 0.55 ** | 0.58 ** | −0.03 ns | 0.82 ** |
| D | 0.08 ns | −0.24 * | 0.135 ns | 0.16 ns | 0.43 ** | −0.28 ** | 0.38 ** | 0.38 ** | 0.48 * |
| Environments | PC1 | PC2 | PC3 | PC4 | PC5 | PC6 | PC7 | PC8 | PC9 | PC10 | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Eigenvalue | Normal | 4.65 | 1.49 | 1.32 | 0.86 | 0.49 | 0.44 | 0.34 | 0.2 | 0.17 | 0.05 |
| Drought | 4.81 | 1.55 | 1.24 | 0.91 | 0.74 | 0.37 | 0.16 | 0.09 | 0.08 | 0.04 | |
| Variability (%) | Normal | 46.5 | 14.9 | 13.2 | 8.6 | 4.9 | 4.4 | 3.4 | 2 | 1.7 | 0.5 |
| Drought | 48.1 | 15.5 | 12.4 | 9.1 | 7.4 | 3.7 | 1.6 | 0.9 | 0.8 | 0.4 | |
| Cumulative (%) | Normal | 46.5 | 61.4 | 74.6 | 83.3 | 88.1 | 92.5 | 95.9 | 97.8 | 99.5 | 100 |
| Drought | 48.1 | 63.6 | 75.9 | 85.1 | 92.4 | 96.2 | 97.8 | 98.7 | 99.6 | 100 |
| Variables | Environments | PC1 | PC2 | PC3 |
|---|---|---|---|---|
| Root length | Normal | 0.349 | −0.188 | −0.480 |
| Drought | 0.155 | −0.486 | 0.247 | |
| Shoot length | Normal | −0.118 | 0.432 | −0.410 |
| Drought | −0.066 | −0.055 | −0.668 | |
| Root/shoot | Normal | 0.334 | −0.211 | −0.440 |
| Drought | 0.002 | 0.491 | 0.434 | |
| Fresh weight | Normal | 0.316 | −0.329 | 0.060 |
| Drought | 0.397 | 0.200 | 0.138 | |
| Dry weight | Normal | 0.406 | 0.212 | −0.062 |
| Drought | 0.435 | 0.050 | 0.071 | |
| Relative water content | Normal | −0.272 | 0.463 | −0.323 |
| Drought | 0.432 | −0.006 | 0.101 | |
| Cell membrane thermo-stability | Normal | 0.381 | 0.067 | 0.102 |
| Drought | 0.430 | 0.011 | 0.020 | |
| Carotenoid | Normal | 0.339 | 0.318 | −0.050 |
| Drought | −0.115 | 0.678 | −0.164 | |
| Chlorophyll a | Normal | 0.317 | 0.450 | 0.138 |
| Drought | 0.359 | 0.000 | −0.333 | |
| Chlorophyll b | Normal | 0.231 | 0.240 | 0.514 |
| Drought | 0.334 | 0.135 | −0.363 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, H.G.M.-D.; Sajjad, M.; Li, M.; Azmat, M.A.; Rizwan, M.; Maqsood, R.H.; Khan, S.H. Selection Criteria for Drought-Tolerant Bread Wheat Genotypes at Seedling Stage. Sustainability 2019, 11, 2584. https://doi.org/10.3390/su11092584
Ahmed HGM-D, Sajjad M, Li M, Azmat MA, Rizwan M, Maqsood RH, Khan SH. Selection Criteria for Drought-Tolerant Bread Wheat Genotypes at Seedling Stage. Sustainability. 2019; 11(9):2584. https://doi.org/10.3390/su11092584
Chicago/Turabian StyleAhmed, Hafiz Ghulam Muhu-Din, Muhammad Sajjad, Mingju Li, Muhammad Abubakkar Azmat, Muhammad Rizwan, Rana Haroon Maqsood, and Sultan Habibullah Khan. 2019. "Selection Criteria for Drought-Tolerant Bread Wheat Genotypes at Seedling Stage" Sustainability 11, no. 9: 2584. https://doi.org/10.3390/su11092584