Use of Dry Dairy Manure Pellets as Nutrient Source for Tomato (Solanum lycopersicum var. cerasiforme) Growth in Soilless Media

1
Department of Biological and Agricultural Engineering, Texas A&M University, College Station, TX 77843, USA
2
Department of Soil and Crop Sciences, Texas A&M University, College Station, TX 77843, USA
3
Elevate Credit, Inc. Fort Worth, TX 76109, USA
4
Department of Biological Systems Engineering, College of Agricultural and Life Sciences, University of Wisconsin-Madison, Madison, WI 53706, USA
*
Author to whom correspondence should be addressed.
Sustainability 2019, 11(3), 811; https://doi.org/10.3390/su11030811
Submission received: 29 November 2018
/
Revised: 28 December 2018
/
Accepted: 18 January 2019
/
Published: 4 February 2019
(This article belongs to the Special Issue Sustainable Crop Production Systems)
Abstract
:A sustainable dairy manure amendment for soilless crop growth systems was evaluated for its ability to provide nutrients and serve as a major component of the growing media. After manure liquid/solid separation, the solids stream containing organic N and P was pelletized and used as a nutrient source for cherry tomato (Solanum lycopersicum var. cerasiforme) culture in soilless media. The pellets are low in moisture, odor, and pathogens, and they can be hauled at lower cost over longer distances and more easily stored than raw or composted manure. Manure pellet additions to soilless media were evaluated at 0%, 2.5%, 5%, 10%, 15%, 20%, 25%, 30%, 40%, and 50% by volume. Manure pellets had a total N content of 3.7%. Fruit size, ripeness, and biomass, plant height, nutrients value in tissue/pellets/media, and time to complete growth cycle were analyzed. Overall, manure pellet treatments improved plant height and growth rate compared to the negative control, especially when pellets were 15% to 50% of the soilless media. This indicates that the nutrients in the manure were being mineralized, and plants were able to utilize the manure-based nutrients for growth. Leaf tissue nutrient analysis revealed that N, K, Zn, and Fe in leaf tissue were not at sufficiency levels at any level of manure pellet addition. Phosphorus and Cu reached sufficiency levels with 10% or greater manure pellet additions. Calcium, Mg, S, Mn, and B were sufficient in all plants, regardless of fertilizer or manure pellet treatment. Manure pellets demonstrate the potential to be used as a substrate and partial growth medium to reduce synthetic fertilizer use for more sustainable soilless container culture.
1. Introduction
Over-application of animal manure to agricultural fields in recent decades is one of the leading contributors to environmental problems such as eutrophication [1,2]. Manure collection and treatment/separation systems are crucial on large dairy farms. The need to export nutrients, especially N and P, out of watersheds [3] has encouraged manure processing and liquid/solid separations. These processes generate a nutrient-rich solid stream that can be processed into a solid fertilizer with a formulated N, P, and K ratio for specific crops. In addition, it also generates a nutrient-reduced liquid stream that can be irrigated locally. After manure liquid/solid separation, the high-solid stream with elevated nutrients, especially N and P, can be pelletized, hauled at lower cost over long distances, and stored more easily than raw or composted manure [4,5]. Furthermore, the pellets are low in odor and pathogens, which makes them feasible for use in indoor and urban agriculture applications.
As we continue to face growing demands for food production, especially with an emphasis on environmental sustainability, it is critical to develop food production systems that can meet these demands [6]. Organic production systems strive to use natural nutrient cycles, such as integrating animal waste into crop production. Pelletized dairy manure is an excellent organic source of nutrients. Organic N and P in manure are mineralized into plant-available inorganic forms (i.e., nitrate—N, ammonium—N, and phosphate—P) as a result of microbial activity. Unlike synthetic fertilizers that are immediately available for plant uptake, mineralization results in a slow release of N and P to the environment, allowing time for plant uptake and reducing environmental impact.
Slow-release nutrients are a desirable characteristic for many agricultural applications ranging from container culture to turfgrass production to crop production. In addition, the organic substrate contributes to organic matter improving the nutrient retention properties in soil and soilless media. Organic matter improves the water holding capacity of soils and can reduce irrigation demands. Additionally, organic production has the potential to be more environmentally sustainable and better able to command a market premium [7]. Our objective was to evaluate the potential use of pelletized animal manure as the sole nutrient source in a soilless potting-media-based contained environment agriculture (CEA) system. In order for manure pellets to be used as a nutrient source, mineralization of the organic nutrients must occur, and the pellet additions cannot be toxic.
2. Materials and Methods
2.1. Manure Pellets
The dairy manure pellets were manufactured by FEECO international (Green Bay, WI, USA). In this manure pelletization process, freshly anaerobic-digested dairy manure was first processed through a liquid/solid separation system including dissolved air floatation and screw press separations. The separated manure solid was then mixed with pre-dried digested manure. The product was pelletized using a disc pelletizer. Next, the wet manure pellets were fed into a rotary dryer (inlet and outlet air temperatures were 288 °C and 121 °C, respectively). The final manure pellet product was screened to a size of 2–4 mm with a moisture content of about 8.5% and a bulk density of 0.56 g·cm−3.
The manure pellets were air-dried and analyzed for nutrient content at Marshfield Agriculture Research Station at University of Wisconsin–Madison. The nutrient assessment included total Kjeldahl N, moisture content, and elements (P, S, K, Ca, Mg, Zn, B, Mn, Fe, Cu, Al, and Na) following standard protocols as described by Peters et al. [8]. Manure pellets were also examined for bacteria indicators according to Liu et al. [9].
The manure pellets used in this experiment were low in moisture (13.7 ± 0.3%) with low to no unpleasant odors (observation). The manure pellets were also low in both bacteria indicators—total coliforms and E. coli—which were below detection limits (100 most probable number method). The dry manure pellets contained higher N and P than typical composted dairy manure products with 0.7% N, 0.4% P, and 1.6% K (wet weight basis) (Table 1) [10]. The higher N and P values of the manure pellet and low odor are both desirable attributes for organic nutrient sources in CEA systems. In addition, the N:P ratio is greater than that of composts (2.05:1 vs. 1.75:1). This is beneficial as the N demand of plants is higher than their P demand, and the greater ratio reduces the potential for over-application of P if pellets are applied on an N basis. The K content of the manure pellets is lower than that of typical composted dairy products due to the manure liquid/solid separation process used to create the pellets. The separation process diverts the aqueous fraction, containing much of the K, away from the solids that are used for pellet production.
2.2. Soilless Media Treatments
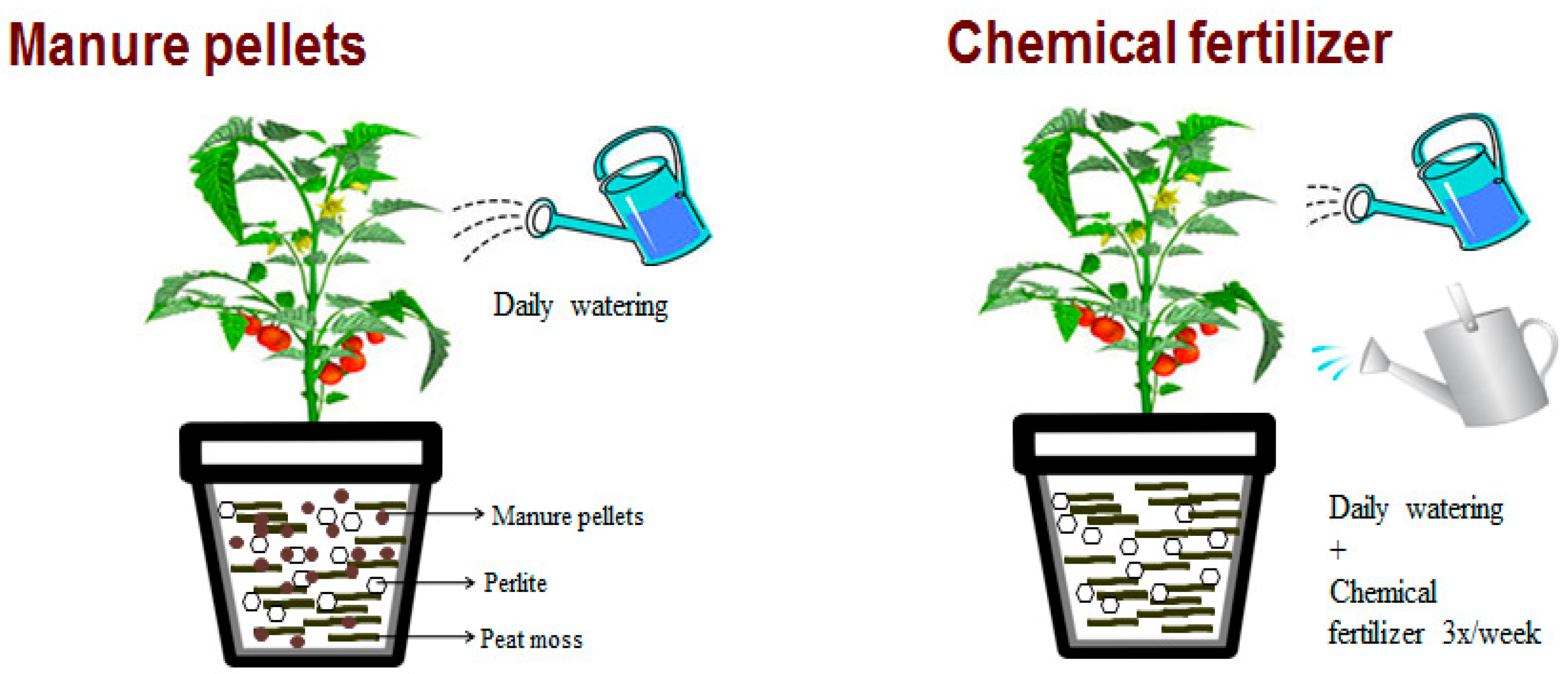
The pellets were evaluated in a soilless container system to isolate the nutrient delivery properties of the pellets. In order to retain water, keep the pH between 5.5 and 6.5, and supply aeration, a soilless medium of 70% (by volume) peat moss (Fafard, Agawam, MA, USA) and 30% (by volume) coarse-sized perlite (Midwest Perlite, Appleton, WI) was made. The total volume of the plant culture media was 1.5 L·pot−1. Treatments included manure pellets mixed into the soilless media at 0%, 2.5%, 5%, 10%, 15%, 20%, 25%, 30%, 40%, and 50% by volume. Nutrient contents were calculated on a g·pot−1 and kg·ha−1 basis (Table 2). A 0% manure pellet treatment with no fertilization was used as a negative control, and a 0% manure pellet treatment fertilized with Peat-Lite Special 20-10-20 liquid synthetic fertilizer (Peters Professionals, Everris, Netherlands) was used as a positive control. This synthetic fertilizer included chelated micronutrients and was applied at 380 mg·N·L−1 at each watering/fertilization 3 times a week (Figure 1). All the pots were watered daily with tap water (Madison Water Utility, Madison, WI, USA) as needed. Triplicates of each treatment were conducted in this experiment for a total of 33 pots.
2.3. Tomato Plant Culture
In order to explore the application of the processed dairy manure pellets, cherry tomato (Solanum lycopersicum var. cerasiforme), one of the most popular garden vegetables, was selected. In general, tomatoes require greater N, P, K, and Ca than many other leafy vegetables. Sweet ‘n Neat Scarlet F1 untreated treated seeds were purchased from Harris Seeds (Rochester, NY, USA). These tomato plants are compact determinant tomatoes. The seeds were germinated and grown at the West Madison Agricultural Research Station greenhouse, University of Wisconsin-Madison. The seedlings were transplanted in April 2016, one plant per pot, 3 weeks after germination and grown for 11 weeks following transplanting. The experiment was conducted in the same greenhouse under conditions of 27/18 °C (day/night) temperatures. In addition to sunlight, 1000 watt high-pressure sodium lights were used as supplemental lighting (600 μmol light at bench height).
Seedling heights were measured biweekly after planting. Soluble salts and pH in the media mixes were measured at Weeks 2, 6, and 10. The soluble salts were measured by electrical conductivity (EC) using a Kelway salinity tester (Wyckoff, NJ, USA). The pH was analyzed using a 1:1 soil-to-water ratio. New mature leaves were analyzed for nutrients at Week 10, a week prior to harvest. The leaf tissue samples were oven-dried at 60 °C for 3 days and analyzed for N, P, K, Ca, Mg, Zn, S, Zn, Mn, B, Fe, and Cu following standard protocols as described by reference [8] at Marshfield Agriculture Research Station at the University of Wisconsin–Madison.
A sample of the potting mixture (100 g) from each pot after harvesting was collected for nutrients analysis at Marshfield Agriculture Research Station at the University of Wisconsin–Madison. Nutrients were extracted using the modified saturated media extract method (DTPA) [8,11].
Tomato fruits and aboveground shoot biomass were harvested during Week 11. The total plant weight (fresh and dry) and fruit weight of each plant were measured right after harvesting. The tomato fruits were separated into ripe (red, marketable) and unripe (non-marketable, green) fruits. The numbers of ripe and unripe tomatoes, average tomato fruit weight per plant, and percentages of ripe and unripe tomatoes per potting mixture were recorded and calculated. The harvested cherry tomatoes from each potting mixture were separated into three sizes according to fruit diameters: small (<1.8 cm), medium (between 1.8 and 2.7 cm), and large (>2.7 cm). The number of fruits from each category was recorded. The tomato shoot biomass was oven-dried at 60 °C for 7 days and evaluated for dry weight.
2.4. Statistical Analysis
Statistical analysis was performed using a GLIMMIX model in SAS 9.4 (SAS Institute, Cary, NC, USA), and all interactions were evaluated. Fruit size, ripeness, biomass, and height were considered fixed effects, while replication was considered a random effect. A probability level of 95% was employed to determine statistical differences. Plant growth rate was determined during the linear growth phase (i.e., Weeks 2 to 8) to determine the slope (i.e., growth rate in cm·week−1).
3. Results and Discussion
3.1. Fruit
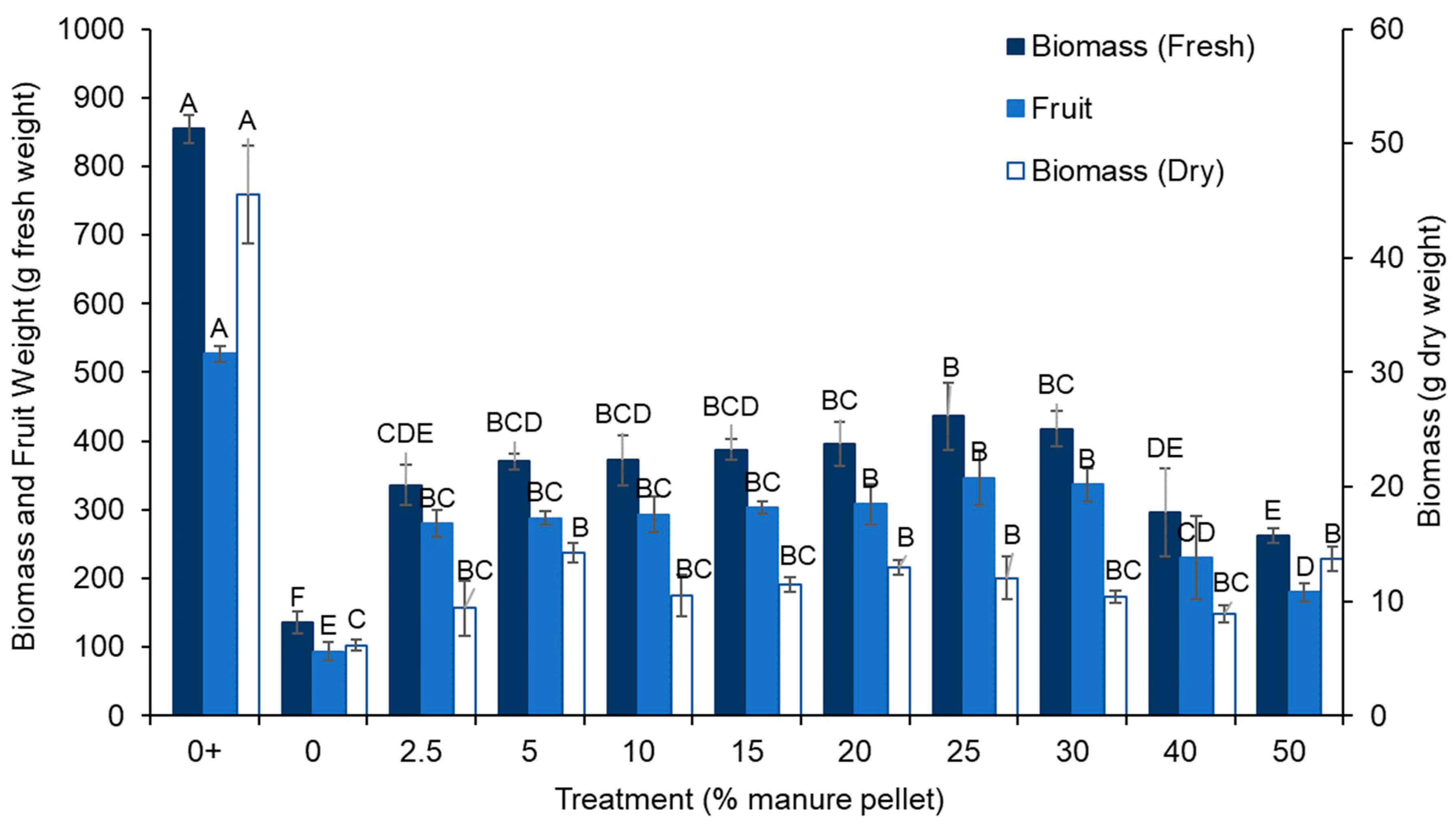
The fertilized control treatment had the highest cherry tomato fruit yield (i.e., total fresh weight of harvested tomatoes) and the unfertilized control had the lowest yield (p < 0.0001; Figure 2). The manure pellets fell in between the fertilized and unfertilized control treatments with the 25% manure pellet treatment being numerically highest, which is consistent with the fresh and dry biomass data.
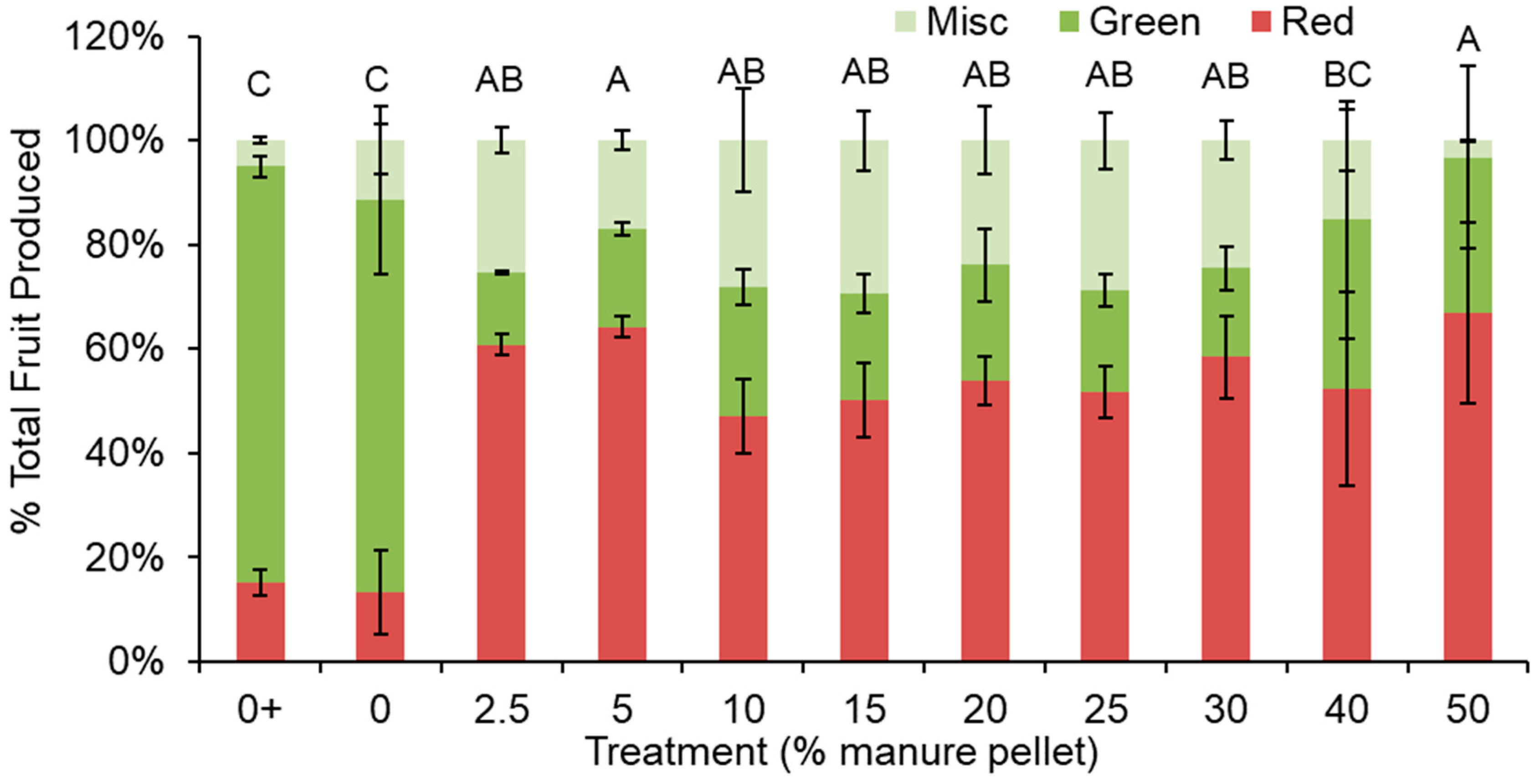
Manure pellet treatments had significantly higher proportions of red fruit and lower proportions of green fruit than the fertilized and unfertilized 0% manure pellet treatments (i.e., positive and negative controls) (p = 0.007; Figure 3). Although the fertilized control had the greatest number of tomatoes, only 15% were ripe at harvest. The majority of tomatoes produced from all treatments were medium in size (i.e., between 1.8 and 2.7 cm in diameter; Figure 4). The fertilized control treatment produced the most large tomatoes (p < 0.0001), which represented 20% of the tomatoes harvested from this treatment. All other treatments produced similar numbers of large tomatoes ranging from 0% to 7% of production for each treatment group. Medium tomatoes were greatest in the fertilized control, lowest in the unfertilized control, and intermediate among the manure pellet treatments (p < 0.0001; Figure 4). Among the manure pellet treatments, 25% manure pellets had the greatest number of medium-sized fruit, but it did not differ from other manure treatments statistically, except for the 2.5% and 50% manure pellet treatments. The number of small-sized fruit did not differ among any of the treatments with or without manure pellets (p = 0.291).
3.2. Plant Growth
Plant height differed by treatment at each week of growth (Week 2, p < 0.001; Week 4, p < 0.001; Week 6, p = 0.030; Week 8, p = 0.005; Week 10, p = 0.009; Figure 5). Plant growth reached its maximum height between Weeks 8 and 10. The tomatoes in the 0% manure pellet treatment (i.e., negative control) were consistently shorter than those in other treatments, which was expected for plants grown in soilless media without added nutrients. In general, the plant heights of manure pellet treatments did not differ from the fertilized control, except at Week 2, where plants for manure pellet treatments between 2.5% and 30% were taller than the fertilized control, and plants for 40% and 50% manure pellet treatments were equivalent to the fertilized control. Another exception occurred at Weeks 8 and 10 when the plants for the 10% manure pellet treatment (Weeks 8 and 10) and the 50% manure pellet treatment (Week 10 only) were shorter than the fertilized control.
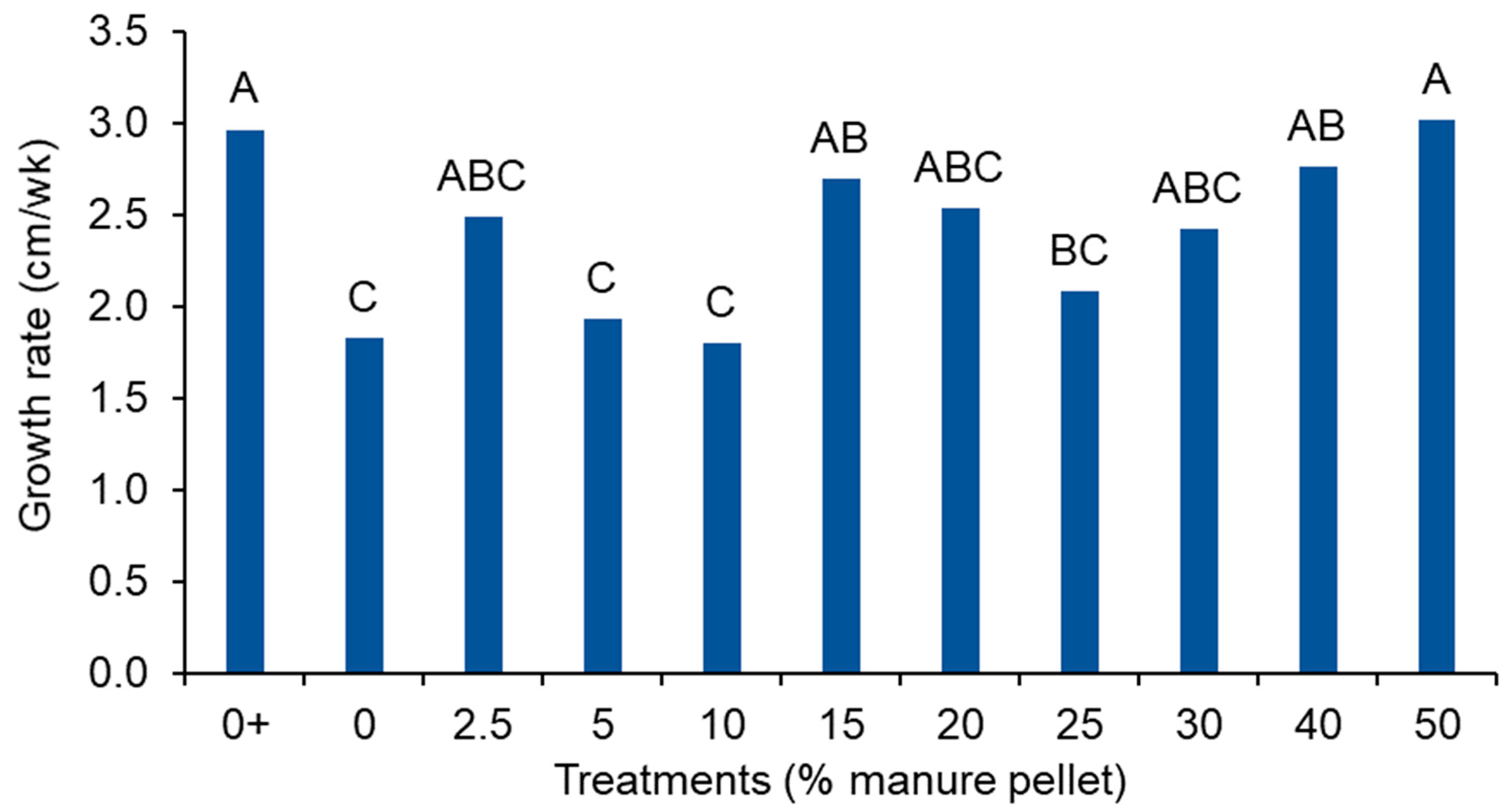
The plant growth rate ranged from 1.8 to 3 cm·week−1 during the linear growth phase of the plant (i.e., Weeks 2–8) and differed among treatments (p = 0.015; Figure 6). The growth rate was highest in the fertilized control and 50% manure pellet treatments at 3 cm·week−1. Except for the 5%, 10%, and 25% manure pellet treatments, the manure pellet treatments did not differ from the fertilized control.
Overall, the manure pellet treatments improved plant height and growth rate compared to the negative control, especially when pellets were 15% to 50% of the soilless media. This indicates that the manure was being mineralized and plants were able to utilize the manure-based nutrients for growth. Interestingly, growth rates were similar for 50% manure pellets and the positive control.
3.3. Biomass
The fertilized control treatment had significantly higher fresh and dry biomass compared to all other treatments, while the unfertilized control was lower in those parameters than all other treatments (p < 0.0001 for all; Figure 2). This data is consistent with the yield and plant height data that suggest that manure pellets were intermediate between fertilized and unfertilized control treatments. Fresh and dry biomass from the manure pellet treatments fell between those in the fertilized and unfertilized control treatments with the 25% manure pellet treatment being numerically highest in fresh and dry biomass.
The biomass was analyzed for nutrients, but because individual samples were composited, they cannot be statistically evaluated. Trends and general observations are noted. Numerically, N was highest in the leaves of the fertilized control (3.90% N), followed by manure treatments (1.30% to 1.91% N), then the unfertilized control (1.15% N; Table 3). Excluding the 40% manure pellet treatment, there was a strong linear trend between percent manure pellet and N in the leaf tissue (R2 = 0.87). The North Carolina Department of Agriculture and Consumer Services Agronomic Division (2001) suggests a sufficiency range of 3.5% to 5.0% N in tomato leaves [12]. Le Bot et al. suggests that 2.5% N in the aerial dry matter is the critical N concentration for tomato grown in soilless culture [13]. Using either of these criteria, the fertilized control was the only treatment to reach these levels.
The P concentrations in the leaf tissue of treatments with > 10% manure pellets were similar to that of the fertilized control (0.61% P), while the treatments with < 10% manure pellets had leaf P concentrations similar to that of the unfertilized control (0.18% P). Excluding the 40% and 50% manure pellet treatments, there was a strong linear trend between % manure pellet and P in the plant tissue (R2 = 0.92). The North Carolina Department of Agriculture and Consumer Services Agronomic Division (2001) suggests a sufficiency range of 0.3% to 0.65% P in tomato leaves [12]. Juarez-Maldonado et al. (2017) reported sufficient concentrations of P in leaf to be between 0.42% and 0.72% [14]. Treatments with 10% or more manure pellets added were considered sufficient using these criteria.
The K concentration in the leaf tissue was numerically lower in manure pellet treatments (0.35% to 0.76% K) compared to the fertilized (3.15% K) or unfertilized (1.02% K) control. There was no linear trend between manure pellet additions and K in the leaf tissue (R2 = 0.03). The concentration of K in the leaf tissue of plants in manure pellet treatments was 34% to 74% that of the unfertilized control. Accounting for differences in biomass from the groups, the total K was similar among the 0% to 50% manure pellet treatments. It is likely that the seemingly lower concentration in pellet treatments is a dilution effect. However, it does suggest that the K in manure pellets is not sufficient to contribute to plant growth greater than that of the unfertilized control. This is further supported by the below sufficiency concentrations of K in leaf tissue of pellet treatments (Table 4) [12]. Manure compost is typically higher in K (i.e., 1.6%) than manure pellets (i.e., 0.9%) because liquid/solid separation was performed before the pelletizing process. This may be something that needs to be adjusted and addressed in the pelleting procedure.
The leaf Ca and Mg concentrations appeared to be greater in manure treatments, except for Ca in the 50% manure pellet treatment, which was lower than in both the fertilized and unfertilized controls. The values are considered to be within the sufficiency range for tomato [12]. All treatments were relatively similar in leaf tissue S concentrations and considered sufficient [12]. The zinc and Cu concentrations in the leaf tissue tended to increase linearly from 0% to 50% manure pellets (R2 = 0.85 and 0.80, respectively), but were lower numerically than in the fertilized control. The zinc concentrations were below sufficiency levels for tomato, while Cu was sufficient when pellet additions were 10% or greater [12]. The leaf tissue concentrations of Fe, Mn, and B were considerably higher in the fertilized control compared to all other treatments. Overall Fe in the leaf tissue decreased with increasing manure pellet additions. Manganese did not follow a trend, and B peaked at 25% to 30% manure pellets. Iron concentrations in the leaf tissue were below sufficiency levels, while Mn and B concentrations were in the sufficient range [12].
The nutrient concentrations in the leaf tissue of the fertilized control fell within the sufficiency range established for greenhouse tomatoes [12]. Nitrogen, K, Zn, and Fe in the leaf tissue were not at sufficiency levels at any level of manure pellet addition. Phosphorus and Cu reached sufficiency levels with 10% or greater manure pellet additions. Calcium, Mg, S, Mn, and B were sufficient in all plants, regardless of fertilizer or manure pellet treatment. Nutrient deficiencies, particularly with macronutrients such as N, P, and K, are known to affect growth, maturity, and yield. However, most nutrients were numerically higher in manure pellet treatments compared to the unfertilized control, especially when the manure pellet rate was >10%. This suggests that manure pellets were able to provide some nutrients to the plant. The increased amount in manure pellet treatments over the unfertilized control is likely attributable to nutrients in the manure pellets; however, the nutrients are either not mineralizing quickly enough to meet the plant’s demand or the quantity of nutrients in the pellets in not enough to support tomato growth.
3.4. Soilless Media Mix Analysis Post-Harvest
Potting media analysis (Table 4) was conducted post-harvest to evaluate the nutrients remaining from the manure in the potting media. Although a direct comparison of treatments cannot be performed because of the compositing of media samples from each treatment, trends among the manure pellet rates can be evaluated. Nitrogen in the manure pellets must be mineralized to either NH4+ or NO3− prior to uptake by the plant. Nitrate was low in the potting media, even in the fertilized control. Nitrate levels considered to be at an acceptable level, based on this analysis method, would be > 40 mg· N· L−1 (Warnke, 1997). However, NO3− concentrations increased linearly with manure pellet additions (R2 = 0.71). Pellets were created following the separation of manure solids and liquids, which likely removes most of the nitrate and ammonium prior to pelletization. Thus, the mineralization of organic forms of N from the manure pellet is likely responsible for plant-available N. Nitrification results in the formation of ammonium that is subsequently nitrified by soil organisms. The presence of nitrate indicates that both of these processes are occurring.
For P, the remaining concentrations increased from 2.5% to 20% manure pellets but decreased from 20% to 50%. It is not clear why this trend occurred except that, potentially, plants were taking up all mineralizable forms when nutrients were very scarce in the low-manure-pellet treatments. However, plants’ uptake mechanisms may be less efficient when nutrients are more readily available. Phosphorus is known to have both high-affinity and low-affinity transporters of P that are induced by low and high external concentrations [15]. Based on the extraction method, values above 3 mg·L−1, which were found in all the manure pellet treatments, are considered adequate for soilless container culture [10].
Except for slightly elevated levels at 25% manure pellets, the K concentration was relatively consistent. The concentrations of K in all media, including the fertilized control, were lower than the 60 mg·L−1, which is the critical level in soilless media [10]. This is not surprising as K+ is highly leachable. Sodium followed the same trend, although the 50% manure pellet rate appeared higher than the other treatments. Similar levels to higher levels of Na compared to K may also have contributed to the lower biomass and fruit yield of the 50% manure pellet treatment as Na and K are both monovalent cations that can compete for uptake [16].
Zinc and S concentrations in potting had a strong trend of increasing with increasing manure pellet addition to the soilless media (R2 = 0.92 and 0.86, respectively), while Fe decreased (R2 = 0.71). The increasingly high pH of the pellet-amended potting media may have reduced the solubility of Fe following mineralization from the manure pellet. However, Zn should have followed the same trend. Another explanation would be the increasing concentrations of organic chelators (e.g., organic acids) from the pellets could have bound Fe, increasing its leachability. The calcium and Mg concentrations were considered above the optimum concentrations [10].
The pH and EC of the potting media increased with increasing manure pellets (Table 4), which is expected when mixing soilless mixture (pH ~ 6.1) with an organic amendment (such as manure pellets, pH ~ 8) containing salts. However, increases were largely between 0% and 10% manure pellet additions and relatively consistent with further additions of manure pellets. A pH of 8.0 is considered too high for tomato production under optimal conditions [17]. All EC levels were considered low using this method [10].
The results of potting media with manure pellet additions post-harvest remain adequate for P, low in K, and potentially low in Fe. Nitrogen is more difficult to assess as a complete analysis of N was not performed. However, the experiments clearly showed that the manure-based N is mineralized, taken up by plants, and nitrified to NO3− in the container.
4. Conclusions and Implications
Overall, our assessment of manure pellets as a fertilization agent in CEA suggests that dried dairy manure pellets are feasible for use. The results for biomass and fruit production in this study are relatively consistent, suggesting that 25% to 30% manure pellet addition is the optimum range for manure pellet culture. Belda et al. observed similar results growing ornamentals in mixtures of manure composts and peat: they found that 25% manure compost produced good-quality plants and that higher rates tended to be phytotoxic [18]. One potential reason for the reduced productivity is a lack of adequate N and K in the manure pellet treatments. None of the manure pellet treatments reached adequate N or K in the leaf tissue. These two elements are particularly in demand while the plant is growing, as well as during flowering and fruiting. Tomato, in particular, is known to demand more N and K than P from potting media [14]. In addition, N is known to affect tomato yield and size [19,20]. This likely explains the similarly in growth rates between the fertilized control and the 50% manure pellet treatment with differences in overall yield.
Culture of tomatoes with a mixture of manure pellets and synthetic fertilizer may be a practical compromise as a promising sustainable crop production system. Fertilizers can be used to supplement manure pellets (or be incorporated into the palletization process) for an overall reduction in total fertilizer used. This would reduce the demand for synthetic fertilizers, which typically utilize fossil fuels as a high-energy source for their production. In addition, animal wastes are recycled in a manner with less odor and nutrient loss than raw manures.
Author Contributions
Conceptualization, Z.L.; methodology, Z.L., J.H. and X.W.; validation, Z.L., J.H. and X.W.; statistical analysis, J.H., X.L.; investigation, Z.L.; writing—original draft preparation, Z.L.; writing—review and editing, J.H.; project administration, T.R.; funding acquisition, T.R.
Acknowledgments
We gratefully acknowledge the United States Department of Agriculture—National Institute of Food and Agriculture for its financial support (USDA BRDI Grant number 2012-10006-19423). We also gratefully acknowledge Lynn Hummel’s help and support from University of Wisconsin–Madison Greenhouses and Joseph Burke’s help with statistical programming.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pachepsky, Y.A.; Sadeghi, A.; Bradford, S.; Shelton, D.; Guber, A.; Dao, T. Transport and fate of manure-borne pathogens: Modeling perspective. Agric. Water Manag. 2006, 86, 81–92. [Google Scholar] [CrossRef]
- Sharpley, A.N.; McDowell, R.W.; Kleinman, P.J. Amounts, forms, and solubility of phosphorus in soils receiving manure. Soil Sci. Soc. Am. J. 2004, 68, 2048–2057. [Google Scholar] [CrossRef]
- Szögi, A.A.; Vanotti, M.B.; Hunt, P.G. Dewatering of phosphorus extracted from liquid swine waste. Bioresour. Technol. 2006, 97, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Møller, H.B.; Sommer, S.G.; Ahring, B.K. Separation efficiency and particle size distribution in relation to manure type and storage conditions. Bioresour. Technol. 2002, 85, 189–196. [Google Scholar] [CrossRef]
- Weiss, W.; Willett, L.; St-Pierre, N.; Borger, D.; McKelvey, T.; Wyatt, D. Varying forage type, metabolizable protein concentration, and carbohydrate source affects manure excretion, manure ammonia, and nitrogen metabolism of dairy cows. J. Dairy Sci. 2009, 92, 5607–5619. [Google Scholar] [CrossRef] [PubMed]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D.; Cassman, K.G.; Matson, P.A.; Naylor, R.; Polasky, S. Agricultural sustainability and intensive production practices. Nature 2002, 418, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Peters, J.B.; Combs, S.; Hoskins, B.; Jarman, J.; Kovar, J.; Watson, M.; Wolf, A.; Wolf, N. Recommended Methods of Manure Analysis; University of Wisconsin Cooperative Extension Publishing: Madison, WI, USA, 2003. [Google Scholar]
- Liu, Z.; Carroll, Z.S.; Long, S.C.; Gunasekaran, S.; Runge, T. Use of cationic polymers to reduce pathogen levels during dairy manure separation. J. Environ. Manag. 2016, 166, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Warncke, D. Greenhouse root media. In Recommended Chemical Soil Test Procedures for the North Central Region, 1st ed.; NCR Publication: Kansas City, MO, USA, 1998; Chapter 14; Volume 221, pp. 61–64. [Google Scholar]
- Butler, T.; Muir, J.P. Dairy manure compost improves soil and increases tall wheatgrass yield. Agron. J. 2006, 98, 1090–1096. [Google Scholar] [CrossRef]
- NCDA&CS. Tissue Sampling & Analysis for Greenhouse Tomatoes; North Carolina Department of Agriculture and Consumer Services Agronomic Division: Raleigh, NC, USA, 2011. Available online: http://www.ncagr.gov/agronomi/pdffiles/gtomato.pdf (accessed on 17 October 2018).
- Bot, J.L.; Jeannequin, B.; Fabre, R. Growth and nitrogen status of soilless tomato plants following nitrate withdrawal from the nutrient solution. Ann. Bot. 2001, 88, 361–370. [Google Scholar] [CrossRef]
- Juárez-Maldonado, A.; de-Alba-Romenus, K.; Morales-Díaz, A.B.; Benavides-Mendoza, A. Macro-nutrient uptake dynamics in greenhouse tomato crop. J. Plant Nutr. 2017, 40, 1908–1919. [Google Scholar] [CrossRef]
- Liu, C.; Muchhal, U.S.; Uthappa, M.; Kononowicz, A.K.; Raghothama, K.G. Tomato phosphate transporter genes are differentially regulated in plant tissues by phosphorus. Plant Physiol. 1998, 116, 91–99. [Google Scholar] [CrossRef] [PubMed]
- White, P.J. Ion uptake mechanisms of individual cells and roots: Short-distance transport. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2012; Chapter 2; pp. 7–47. [Google Scholar]
- Sainju, U.M.; Dris, R.; Singh, B. Mineral nutrition of tomato. J. Food. Agric. Environ. 2003, 1, 176–183. [Google Scholar]
- Belda, R.M.; Mendoza-Hernández, D.; Fornes, F. Nutrient-rich compost versus nutrient-poor vermicompost as growth media for ornamental-plant production. J. Plant Nutr. Soil Sci. 2013, 176, 827–835. [Google Scholar] [CrossRef]
- Elia, A.; Conversa, G.; La Rotonda, P.; Montemurrow, F. Nitrogen level effect on yield and quality of fertigated processing tomato in Southern Italy. Acta Hortic. 2007, 235–240. [Google Scholar] [CrossRef]
- Ozores-Hampton, M.; Simonne, E.; Roka, F.; Morgan, K.; Sargent, S.; Snodgrass, C.; McAvoy, E. Nitrogen rates effects on the yield nutritional status, fruit quality, and the profitability of tomato grown in the spring with subsurface irrigation. Hortscience 2012, 57, 1129–1135. [Google Scholar] [CrossRef]
Figure 1.
Experimental setup for cherry tomato growth in soilless media.

Figure 2.
Fresh biomass, total fruit yield, and dry biomass of cherry tomatoes at harvest. The 0% manure pellet treatment with fertilizer (i.e., positive control) is indicated with +. Letters that differ among treatments indicate differences in height at that given week of plant growth. Error bars indicate standard error.
Figure 2.
Fresh biomass, total fruit yield, and dry biomass of cherry tomatoes at harvest. The 0% manure pellet treatment with fertilizer (i.e., positive control) is indicated with +. Letters that differ among treatments indicate differences in height at that given week of plant growth. Error bars indicate standard error.

Figure 3.
Percentage of harvested cherry tomatoes as red, green, and miscellaneous (Misc). “Miscellaneous” includes damaged fruit. The 0% manure pellet treatment with fertilizer (i.e., positive control) is indicated with +. Letters that differ among treatments indicate differences in red fruit production. Error bars indicate standard error.
Figure 3.
Percentage of harvested cherry tomatoes as red, green, and miscellaneous (Misc). “Miscellaneous” includes damaged fruit. The 0% manure pellet treatment with fertilizer (i.e., positive control) is indicated with +. Letters that differ among treatments indicate differences in red fruit production. Error bars indicate standard error.

Figure 4.
Number of tomatoes grouped by size as small (<1.8 cm), medium (between 1.8 and 2.7 cm), and large (>2.7 cm). The 0% manure pellet treatment with fertilizer (i.e., positive control) is indicated with +. Letters that differ among treatments indicate differences within each tomato size grouping (i.e., small, medium, and large). Error bars indicate standard error.
Figure 4.
Number of tomatoes grouped by size as small (<1.8 cm), medium (between 1.8 and 2.7 cm), and large (>2.7 cm). The 0% manure pellet treatment with fertilizer (i.e., positive control) is indicated with +. Letters that differ among treatments indicate differences within each tomato size grouping (i.e., small, medium, and large). Error bars indicate standard error.

Figure 5.
Cherry tomato plant height at 2, 4, 6, 8, and 10 weeks of growth. The 0% manure pellet treatment with fertilizer (i.e., positive control) is indicated with +. Letters that differ among treatments indicate differences in height at that given week of plant growth. Error bars indicate standard error.
Figure 5.
Cherry tomato plant height at 2, 4, 6, 8, and 10 weeks of growth. The 0% manure pellet treatment with fertilizer (i.e., positive control) is indicated with +. Letters that differ among treatments indicate differences in height at that given week of plant growth. Error bars indicate standard error.

Figure 6.
Cherry tomato growth rate in Weeks 2 to 8. The 0% manure pellet treatment with fertilizer (i.e., positive control) is indicated with +. Letters that differ among treatments indicate differences in height at that given week of plant growth.
Figure 6.
Cherry tomato growth rate in Weeks 2 to 8. The 0% manure pellet treatment with fertilizer (i.e., positive control) is indicated with +. Letters that differ among treatments indicate differences in height at that given week of plant growth.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Nutrient content in dry manure pellets (dry weight basis).
| Nutrient | N | P | K | Ca | Mg | S | Zn | B | Mn | Fe | Cu | Al | Na |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| % | mg·kg−1 | ||||||||||||
| Pellets | 3.7 | 1.8 | 0.90 | 2.9 | 1.3 | 0.50 | 529 | 27 | 290 | 1000 | 170 | 634 | 3000 |
Table 2.
Pellet and nutrient contents of manure pellet treatments.
| Treatment | ||||||||
|---|---|---|---|---|---|---|---|---|
| % by Volume | g Pellets pot−1 | Mg Pellets ha−1 | g N pot−1 | kg N ha−1 | g P pot−1 | kg P ha−1 | g K pot−1 | kg K ha−1 |
| 2.5% | 21 | 0.21 | 0.8 | 777 | 0.4 | 378 | 0.2 | 189 |
| 5% | 42 | 0.42 | 1.6 | 1554 | 0.8 | 756 | 0.4 | 378 |
| 10% | 84 | 0.84 | 3.1 | 3108 | 1.5 | 1512 | 0.8 | 756 |
| 15% | 126 | 1.26 | 4.7 | 4662 | 2.3 | 2268 | 1.1 | 1134 |
| 20% | 168 | 1.68 | 6.2 | 6216 | 3.0 | 3024 | 1.5 | 1512 |
| 25% | 210 | 2.10 | 7.8 | 7770 | 3.8 | 3780 | 1.9 | 1890 |
| 30% | 252 | 2.52 | 9.3 | 9324 | 4.5 | 4536 | 2.3 | 2268 |
| 40% | 336 | 3.36 | 12.4 | 12432 | 6.0 | 6048 | 3.0 | 3024 |
| 50% | 420 | 4.20 | 15.5 | 15540 | 7.6 | 7560 | 3.8 | 3780 |
Table 3.
Nutrient concentrations in cherry tomato leaf tissue at 10 weeks. Treatments include the percentage of manure added to the soilless media. The 0+ treatment indicates 0% manure pellets with fertilizer (i.e., fertilizer control).
Table 3.
Nutrient concentrations in cherry tomato leaf tissue at 10 weeks. Treatments include the percentage of manure added to the soilless media. The 0+ treatment indicates 0% manure pellets with fertilizer (i.e., fertilizer control).
| Treatment | N | P | K | Ca | Mg | S | Zn | Mn | B | Fe | Cu |
|---|---|---|---|---|---|---|---|---|---|---|---|
| % Manure | % | mg·kg−1 | |||||||||
| Sufficiency range * | 3.5–5.0 | 0.3-0.65 | 2.8-4.0 | 1.0–3.0 | 0.35–1.0 | 0.2–1.0 | 18–80 | 25–200 | 30–75 | 50–300 | 5–35 |
| 0+ | 3.90 | 0.61 | 3.15 | 2.19 | 1.04 | 0.36 | 37 | 166 | 141 | 94 | 8.8 |
| 0 | 1.15 | 0.18 | 1.02 | 1.76 | 1.14 | 0.60 | 3.8 | 71 | 41 | 46 | 2.5 |
| 2.5 | 1.30 | 0.12 | 0.64 | 1.89 | 1.27 | 0.65 | 3.1 | 101 | 37 | 46 | 3.4 |
| 5 | 1.27 | 0.15 | 0.46 | 2.11 | 1.46 | 0.64 | 4.0 | 69 | 41 | 42 | 4.4 |
| 10 | 1.49 | 0.35 | 0.38 | 2.88 | 1.80 | 0.73 | 5.6 | 38 | 53 | 45 | 5.0 |
| 15 | 1.61 | 0.58 | 0.40 | 2.98 | 2.07 | 0.70 | 6.3 | 36 | 84 | 36 | 6.6 |
| 20 | 1.63 | 0.50 | 0.35 | 2.63 | 2.09 | 0.69 | 5.0 | 38 | 86 | 39 | 6.9 |
| 25 | 1.62 | 0.63 | 0.38 | 3.05 | 2.60 | 0.76 | 6.2 | 49 | 108 | 35 | 7.0 |
| 30 | 1.62 | 0.73 | 0.38 | 3.43 | 2.69 | 0.83 | 9.0 | 65 | 102 | 31 | 7.7 |
| 40 | 1.41 | 0.54 | 0.43 | 2.58 | 2.58 | 0.74 | 8.6 | 55 | 95 | 28 | 7.7 |
| 50 | 1.91 | 0.47 | 0.76 | 1.59 | 2.40 | 0.58 | 15 | 47 | 77 | 35 | 7.9 |
* Sufficiency ranges are reported from the North Carolina Dept. of Agriculture & Consumer Services Agronomic Division [12].
Table 4.
Soilless media analysis of pH, electrical conductivity (EC), and nutrients post-harvest. Treatments indicate the percentage of manure pellets added to the media. The treatment indicated with a + represents the fertilized control. (DTPA: diethylenetriaminepentaacetic acid).
Table 4.
Soilless media analysis of pH, electrical conductivity (EC), and nutrients post-harvest. Treatments indicate the percentage of manure pellets added to the media. The treatment indicated with a + represents the fertilized control. (DTPA: diethylenetriaminepentaacetic acid).
| Treatment | pH | EC | Nitrate | P | K | Ca | Mg | Fe | Na | Zn | S |
|---|---|---|---|---|---|---|---|---|---|---|---|
| dS m−1 | mg·L−1 DTPA Saturated Media Extract Method | ||||||||||
| 0%+ | 5.7 | 0.14 | 13 | 20 | 20 | 129 | 88 | 33 | 27 | 44 | 5 |
| 0% | 6.1 | 0.09 | 0 | 5 | 6 | 142 | 90 | 38 | 11 | 9 | 8 |
| 2.5% | 7.3 | 0.10 | 0 | 3 | 2 | 168 | 82 | 29 | 11 | 63 | 5 |
| 5% | 7.6 | 0.12 | 1 | 4 | 5 | 190 | 82 | 23 | 11 | 92 | 5 |
| 10% | 7.9 | 0.15 | 6 | 21 | 4 | 220 | 86 | 16 | 10 | 163 | 9 |
| 15% | 7.8 | 0.21 | 5 | 49 | 5 | 226 | 101 | 12 | 11 | 236 | 9 |
| 20% | 7.9 | 0.26 | 4 | 53 | 5 | 223 | 111 | 9 | 12 | 259 | 11 |
| 25% | 7.9 | 0.25 | 5 | 46 | 27 | 233 | 131 | 6 | 19 | 252 | 16 |
| 30% | 7.9 | 0.27 | 12 | 43 | 9 | 217 | 121 | 7 | 12 | 311 | 17 |
| 40% | 8.0 | 0.36 | 8 | 35 | 14 | 206 | 140 | 5 | 15 | 337 | 22 |
| 50% | 8.2 | 0.42 | 22 | 26 | 14 | 222 | 185 | 5 | 41 | 401 | 39 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liu, Z.; Howe, J.; Wang, X.; Liang, X.; Runge, T. Use of Dry Dairy Manure Pellets as Nutrient Source for Tomato (Solanum lycopersicum var. cerasiforme) Growth in Soilless Media. Sustainability 2019, 11, 811. https://doi.org/10.3390/su11030811
AMA Style
Liu Z, Howe J, Wang X, Liang X, Runge T. Use of Dry Dairy Manure Pellets as Nutrient Source for Tomato (Solanum lycopersicum var. cerasiforme) Growth in Soilless Media. Sustainability. 2019; 11(3):811. https://doi.org/10.3390/su11030811
Chicago/Turabian StyleLiu, Zong, Julie Howe, Xiao Wang, Xiao Liang, and Troy Runge. 2019. "Use of Dry Dairy Manure Pellets as Nutrient Source for Tomato (Solanum lycopersicum var. cerasiforme) Growth in Soilless Media" Sustainability 11, no. 3: 811. https://doi.org/10.3390/su11030811
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.