Zonation and Directional Dynamics of Mangrove Forests Derived from Time-Series Satellite Imagery in Mai Po, Hong Kong



Abstract
1. Introduction
2. Study Area and Data Sets
3. Methods
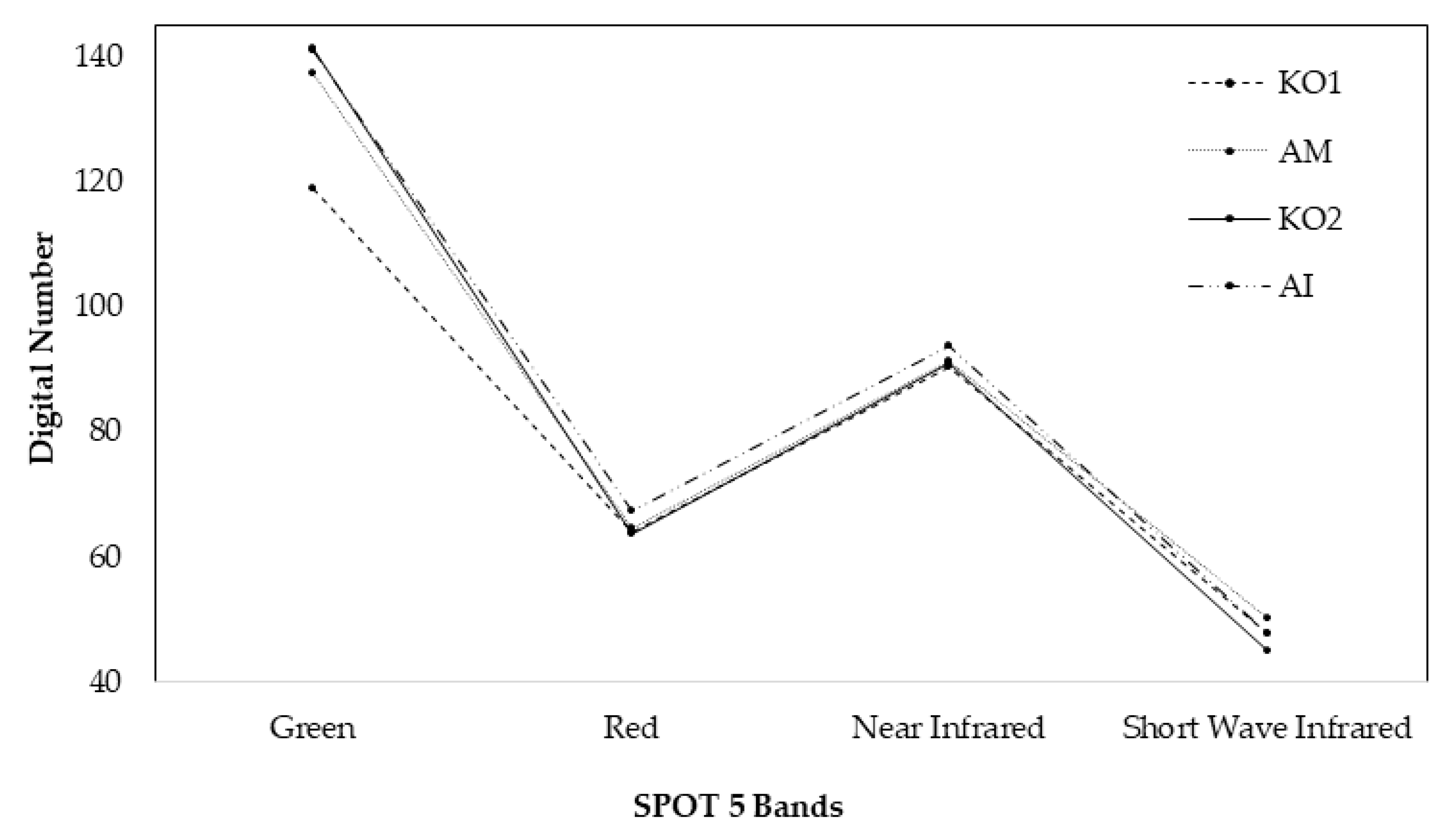
3.1. Mangrove Forest Classification on Species Level
3.2. Directional Analysis Using Standard Deviational Ellipse
3.3. Validation and Accuracy Assessment
4. Results
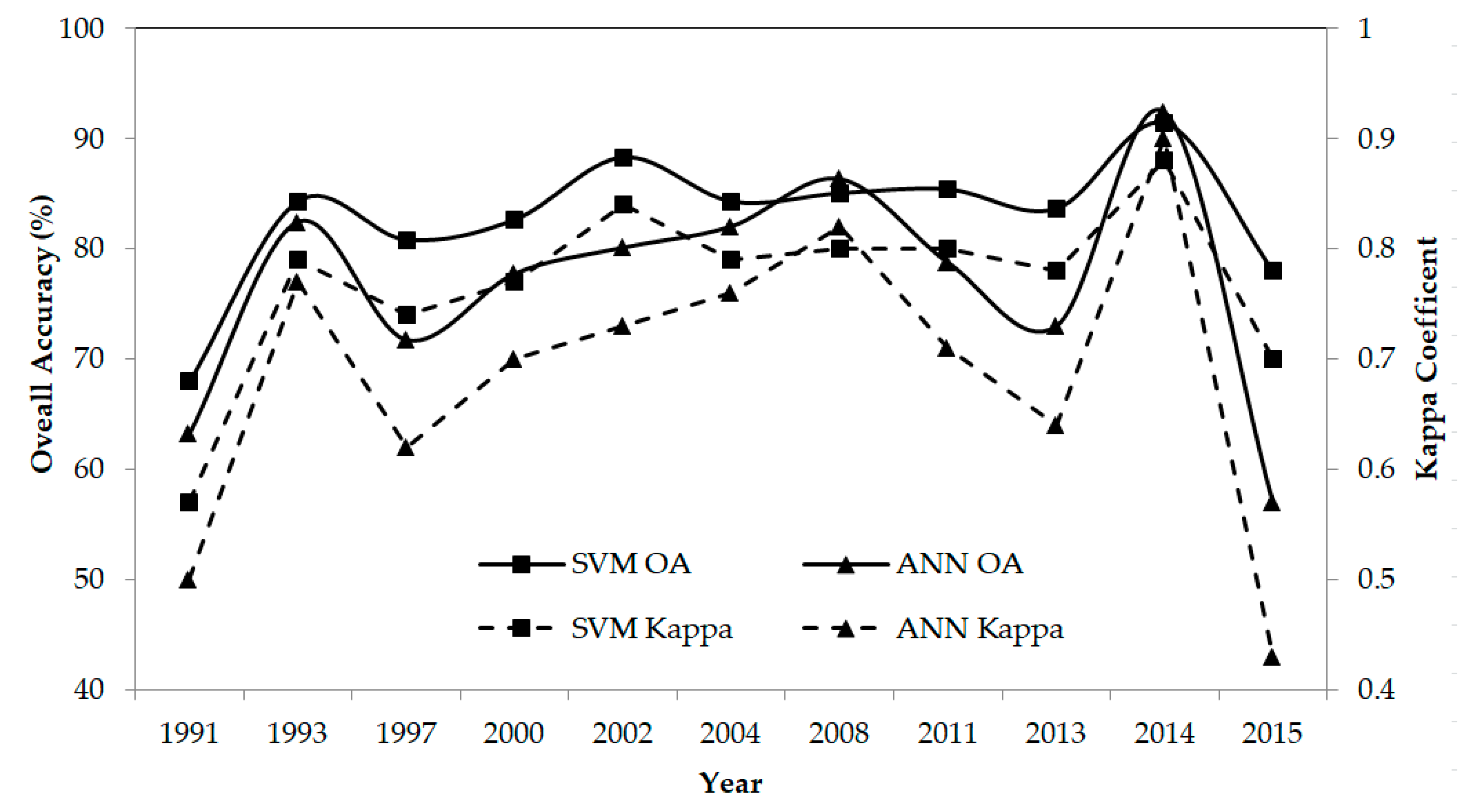
4.1. Accuracy Assessment of the Mangrove Species Classification
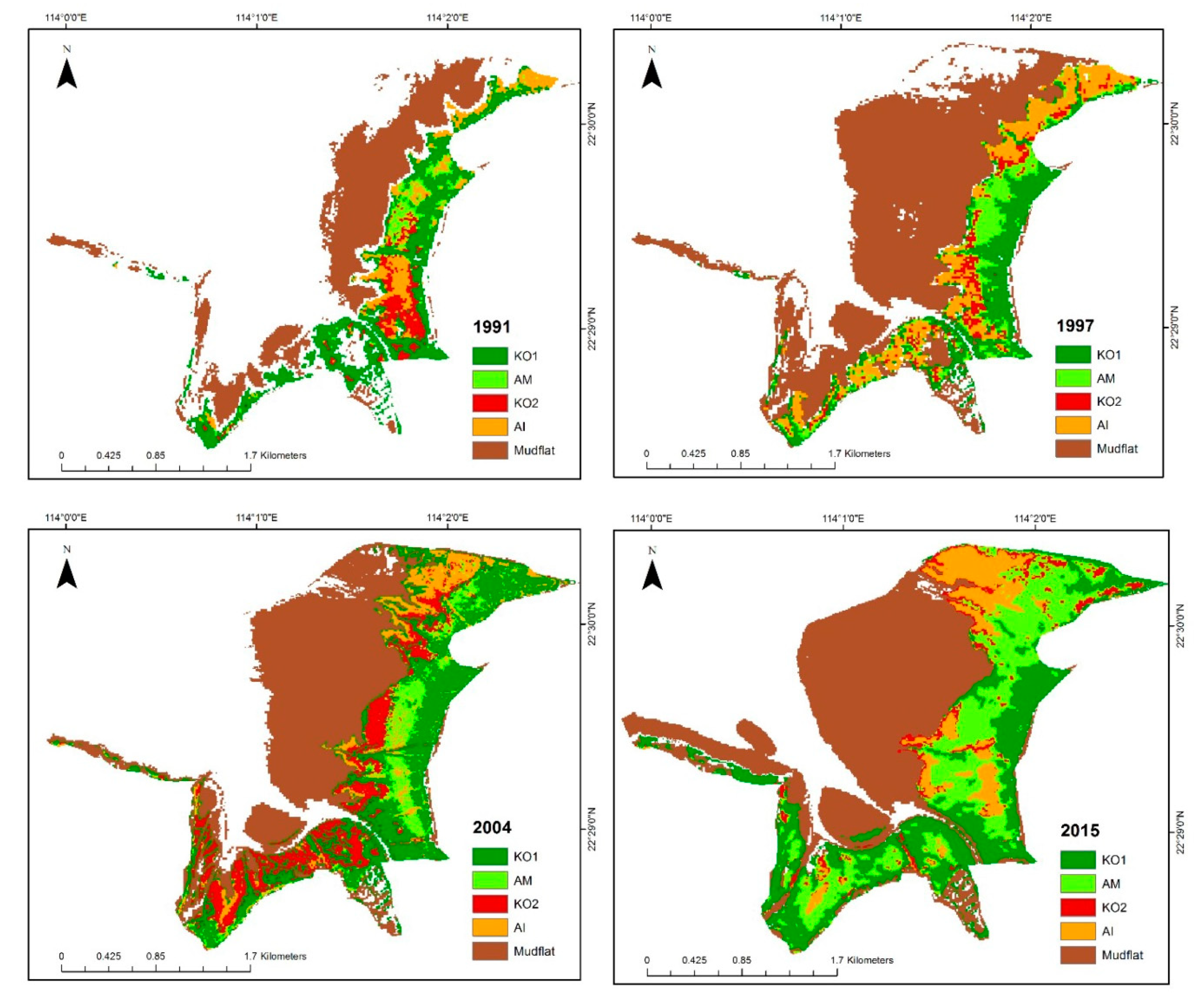
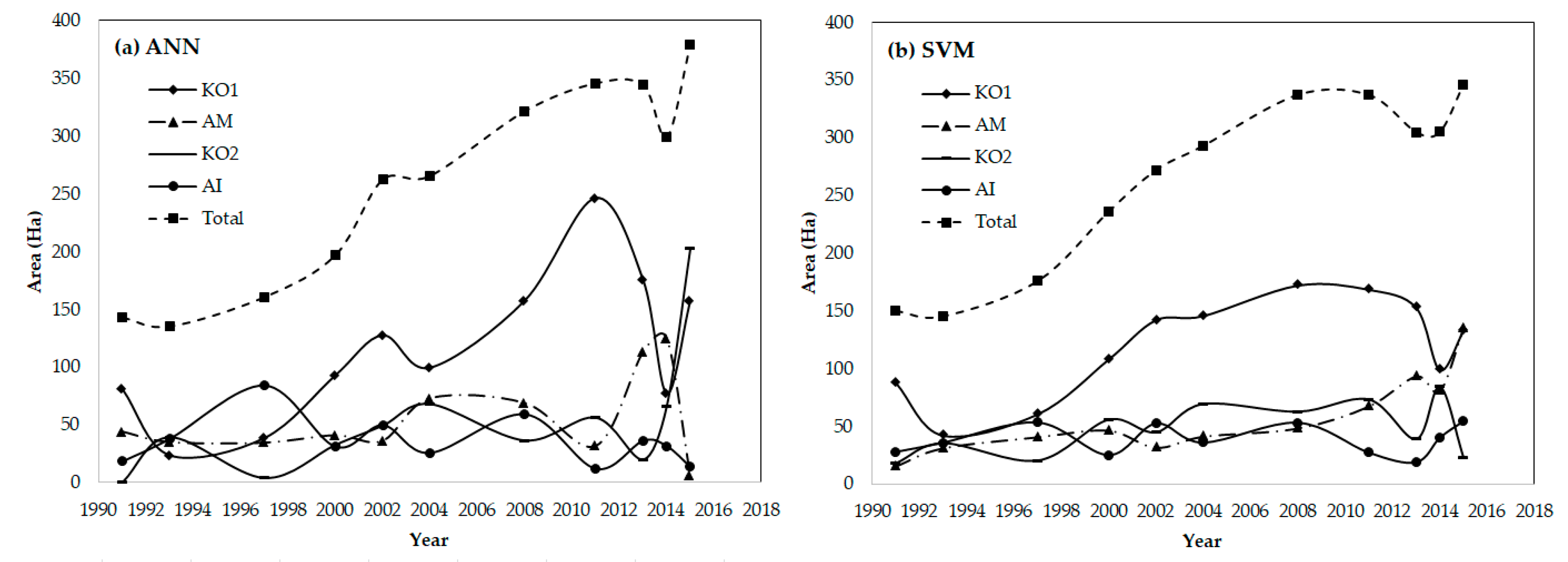
4.2. Mapping Mangrove Zonation in the Last 25 Years
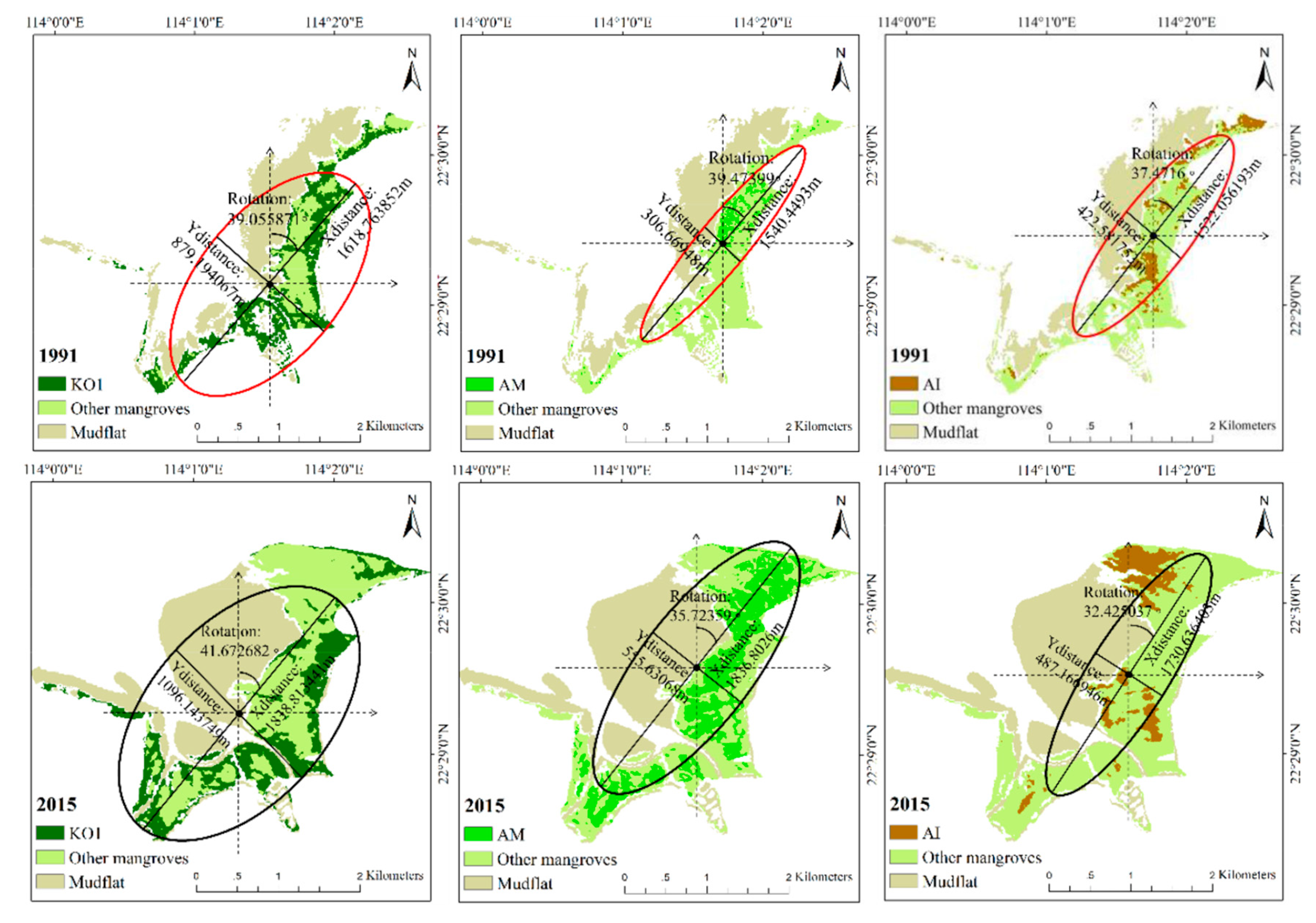
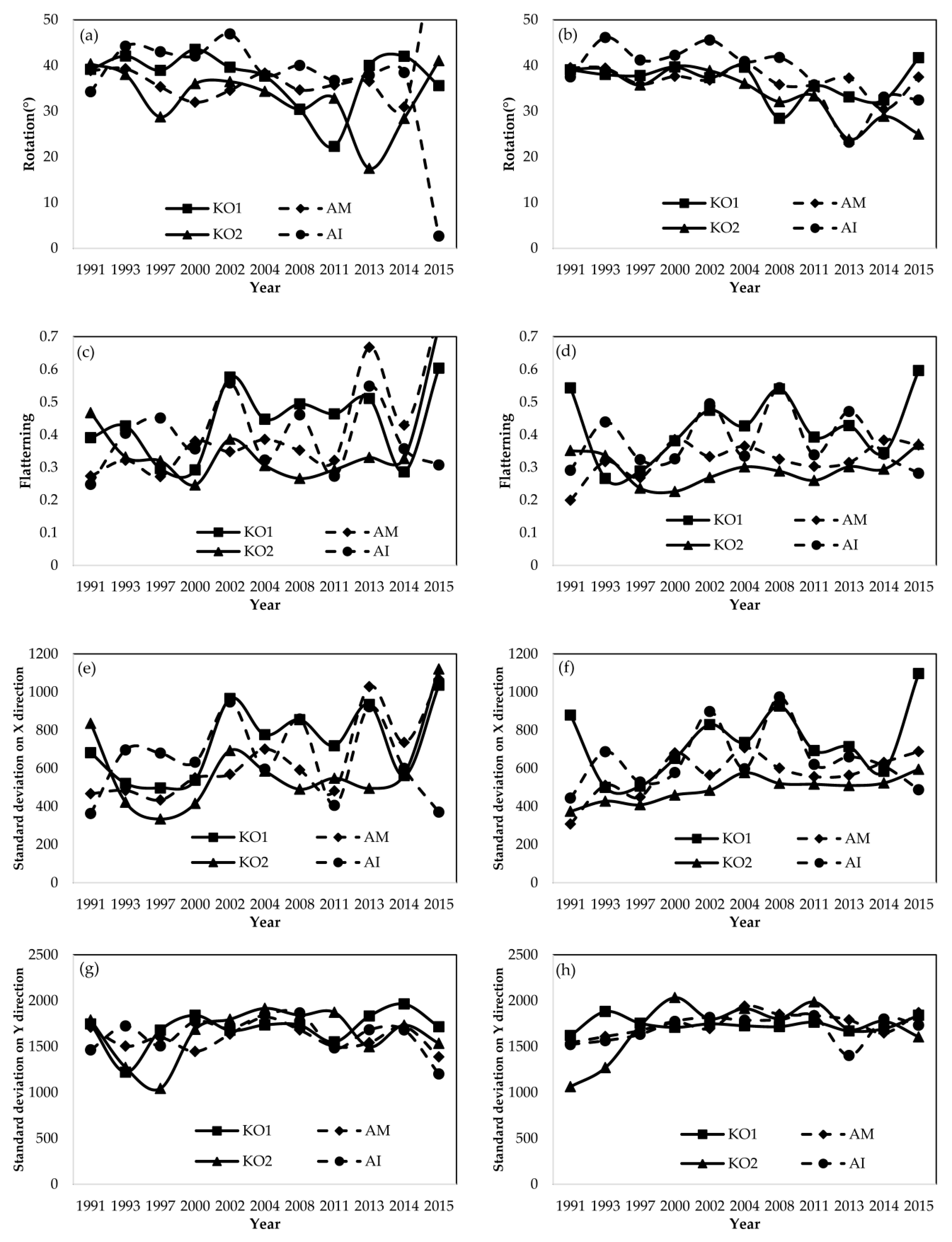
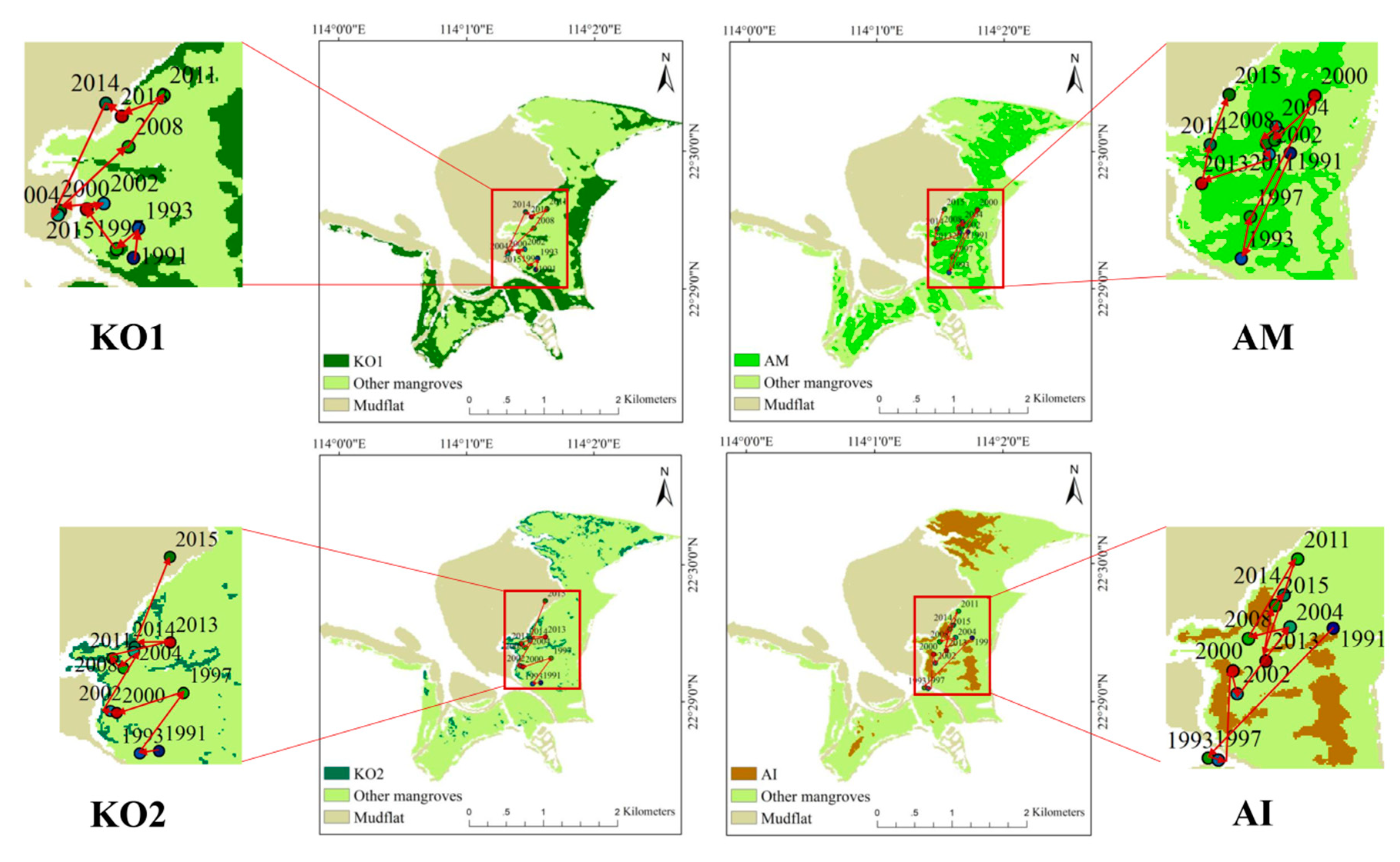
4.3. Temporal Dynamics on Directional Development at Species Level
5. Discussion
5.1. Discussion on the Driving Factors of Mangrove Changes
5.2. Comments on the Sustainable Development of Mai Po Reserve
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Alongi, D.M. Present state and future of the world’s mangrove forests. Environ. Conserv. 2002, 29, 331–349. [Google Scholar] [CrossRef]
- Duke, N.C. Phenologies and litter fall of two mangrove trees, Sonneratia alba Sm. and S. caseolaris (L.) Engl., and their putative hybrid, S.× gulngai NC Duke. Aust. J. Bot. 1988, 36, 473–482. [Google Scholar] [CrossRef]
- Kathiresan, K.; Bingham, B.L. Biology of mangroves and mangrove ecosystems. Adv. Mar. Biol. 2001, 40, 81–251. [Google Scholar]
- Furukawa, K.; Wolanski, E.; Mueller, H. Currents and sediment transport in mangrove forests. Estuar. Coast. Shelf Sci. 1997, 44, 301–310. [Google Scholar] [CrossRef]
- Nagelkerken, I.; Blaber, S.; Bouillon, S.; Green, P.; Haywood, M.; Kirton, L.; Meynecke, J.-O.; Pawlik, J.; Penrose, H.; Sasekumar, A. The habitat function of mangroves for terrestrial and marine fauna: A review. Aquat. Bot. 2008, 89, 155–185. [Google Scholar] [CrossRef]
- Duke, N.C.; Meynecke, J.O.; Dittmann, S.; Ellison, A.M.; Anger, K.; Berger, U.; Cannicci, S.; Diele, K.; Ewel, K.C.; Field, C.D.; et al. A world without mangroves? Science 2007, 317, 41–42. [Google Scholar] [CrossRef] [PubMed]
- Ellison, A.M.; Mukherjee, B.B.; Karim, A. Testing patterns of zonation in mangroves: Scale dependence and environmental correlates in the Sundarbans of Bangladesh. J. Ecol. 2000, 88, 813–824. [Google Scholar] [CrossRef]
- Youssef, T.; Saenger, P. Mangrove zonation in Mobbs Bay—Australia. Estuar. Coast. Shelf Sci. 1999, 49, 43–50. [Google Scholar] [CrossRef]
- Rogers, K.; Lymburner, L.; Salum, R.; Brooke, B.P.; Woodroffe, C.D. Mapping of mangrove extent and zonation using high and low tide composites of Landsat data. Hydrobiologia 2017, 803, 49–68. [Google Scholar] [CrossRef]
- Bullock, E.L.; Fagherazzi, S.; Nardin, W.; Vo-Luong, P.; Nguyen, P.; Woodcock, C.E. Temporal patterns in species zonation in a mangrove forest in the Mekong Delta, Vietnam, using a time series of Landsat imagery. Cont. Shelf Res. 2017, 147, 144–154. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, T.; Liu, M.; Jia, M.; Lin, H.; Chu, L.; Devlin, A.T. Potential of Combining Optical and Dual Polarimetric SAR Data for Improving Mangrove Species Discrimination Using Rotation Forest. Remote Sens. (Basel) 2018, 10, 467. [Google Scholar] [CrossRef]
- Zhang, H.; Xu, R. Exploring the optimal integration levels between SAR and optical data for better urban land cover mapping in the Pearl River Delta. Int. J. Appl. Earth Obs. Geoinf. 2018, 64, 87–95. [Google Scholar] [CrossRef]
- Wan, L.; Zhang, H.; Wang, T.; Li, G.; Lin, H. Mangrove species discrimination from very high resolution imagery using gaussian markov random field model. Wetlands 2017. accepted. [Google Scholar]
- Wang, T.; Zhang, H.; Lin, H.; Fang, C. Textural–spectral feature-based species classification of mangroves in Mai Po Nature Reserve from Worldview-3 imagery. Remote Sens. (Basel) 2016, 8, 24. [Google Scholar] [CrossRef]
- Jia, M.M.; Liu, M.Y.; Wang, Z.M.; Mao, D.H.; Ren, C.Y.; Cui, H.S. Evaluating the Effectiveness of Conservation on Mangroves: A Remote Sensing-Based Comparison for Two Adjacent Protected Areas in Shenzhen and Hong Kong, China. Remote Sens. (Basel) 2016, 8, 627. [Google Scholar] [CrossRef]
- Viennois, G.; Proisy, C.; Feret, J.B.; Prosperi, J.; Sidik, F.; Suhardjono; Rahmania, R.; Longepe, N.; Germain, O.; Gaspar, P. Multitemporal Analysis of High-Spatial-Resolution Optical Satellite Imagery for Mangrove Species Mapping in Bali, Indonesia. IEEE J. Stars 2016, 9, 3680–3686. [Google Scholar] [CrossRef]
- Young, L.; Melville, D. Conservation of the Deep Bay environment. In The Marine Biology of the South China Sea, Proceedings of the First International Conference on the Marine Biology of Hong Kong and the South China Sea, Hong Kong, China, 28 October–3 November 1990, 1st ed.; Brain, M., Ed.; Hong Kong University Press: Hong Kong, China, 1993; Volume 1, pp. 211–232. [Google Scholar]
- Howlett, B.E. A Review of 1995 and a Pictorial Review of the Past Fifty Years; Government Printer: Hong Kong, China, 1996; ISBN 9620202201. [Google Scholar]
- Tam, N.F.; Wong, Y.S.; Lu, C.Y.; Berry, R. Mapping and characterization of mangrove plant communities in Hong Kong. In Asia-Pacific Conference on Science and Management of Coastal Environment; Springer: Dordrecht, The Netherlands, 1997; pp. 25–37. [Google Scholar]
- Agriculture, Fisheries and Conservation Department (AFCD). Mai Po Inner Deep Bay RAMSAR Site Management Plan Executive Summary; Agriculture, Fisheries and Conservation Department: Hong Kong, China, 2011.
- Yipp, M.W.; Hau, C.; Walthew, G. Conservation evaluation of nine Hong Kong mangals. In Proceedings of the Asia-Pacific Symposium on Mangrove Ecosystems, Hong Kong, China, 1–3 September 1993; Springer: Dordrecht, The Netherlands, 1995; pp. 323–333. [Google Scholar]
- Jia, M.; Zhang, Y.; Wang, Z.; Song, K.; Ren, C. Mapping the distribution of mangrove species in the Core Zone of Mai Po Marshes Nature Reserve, Hong Kong, using hyperspectral data and high-resolution data. Int. J. Appl. Earth Obs. Geoinf. 2014, 33, 226–231. [Google Scholar] [CrossRef]
- Wong, F.K.K.; Fung, T. Combining EO-1 Hyperion and Envisat ASAR data for mangrove species classification in Mai Po Ramsar Site, Hong Kong. Int. J. Remote Sens. 2014, 35, 7828–7856. [Google Scholar] [CrossRef]
- Cortes, C.; Vapnik, V. Support-vector networks. Mach. Learn. 1995, 20, 273–297. [Google Scholar] [CrossRef]
- Vapnik, V. The support vector method of function estimation. In Nonlinear Modeling; Springer: Boston, MA, USA, 1998; pp. 55–85. [Google Scholar]
- Song, C.; Woodcock, C.E.; Seto, K.C.; Lenney, M.P.; Macomber, S.A. Classification and change detection using Landsat TM data: When and how to correct atmospheric effects? Remote Sens. Environ. 2001, 75, 230–244. [Google Scholar] [CrossRef]
- Agriculture, Fisheries and Conservation Department (AFCD). Mapping of Distribution and Extent of Mangroves in Hong Kong Using Satellite Image Analysis; Agriculture, Fisheries and Conservation Department: Hong Kong, China, 2008.
- Balke, T.; Bouma, T.J.; Horstman, E.M.; Webb, E.L.; Erftemeijer, P.L.; Herman, P.M. Windows of opportunity: Thresholds to mangrove seedling establishment on tidal flats. Mar. Ecol. Prog. Ser. 2011, 440, 1–9. [Google Scholar] [CrossRef]
- Chan, P.; Ching, S.; Erftemeijer, P. Developing a mangrove management strategy in the estuaries of Deep Bay, Shan Pui River and Tin Shui Wai drainage channel. In Proceedings of the CIWEM Hong Kong Water Conference, Hong Kong, China, 11–12 May 2011. [Google Scholar]
- Wong, S.; Li, Y. Hydrographic surveys and sedimentation in Deep Bay, Hong Kong. Environ. Geol. Water Sci. 1990, 15, 111–118. [Google Scholar] [CrossRef]
- Lee, S.-Y. The Mangrove Ecosystem of Deep Bay and the Mai Po Marshes, Hong Kong: Proceedings of the International Workshop on the Mangrove Ecosystem of Deep Bay and the Mai Po Marshes, Hong Kong, China, 3–20 September 1993, 1st ed.; Lee, S.-Y., Ed.; Hong Kong University Press: Hong Kong, China, 1999. [Google Scholar]
- Crase, B.; Liedloff, A.; Vesk, P.A.; Burgman, M.A.; Wintle, B.A. Hydroperiod is the main driver of the spatial pattern of dominance in mangrove communities. Glob. Ecol. Biogeogr. 2013, 22, 806–817. [Google Scholar] [CrossRef]
- Van Loon, A.; Dijksma, R.; Van Mensvoort, M. Hydrological classification in mangrove areas: A case study in Can Gio, Vietnam. Aquat. Bot. 2007, 87, 80–82. [Google Scholar] [CrossRef]
- Leong, R.C.; Friess, D.A.; Crase, B.; Lee, W.K.; Webb, E.L. High-resolution pattern of mangrove species distribution is controlled by surface elevation. Estuar. Coast. Shelf Sci. 2017, 202, 185–192. [Google Scholar] [CrossRef]
- McKee, K.L. Soil physicochemical patterns and mangrove species distribution—Reciprocal effects? J. Ecol. 1993, 477–487. [Google Scholar] [CrossRef]
- McKee, K.L. Growth and physiological responses of neotropical mangrove seedlings to root zone hypoxia. Tree Physiol. 1996, 16, 883–889. [Google Scholar] [CrossRef] [PubMed]
- Youssef, T.; Saenger, P. Photosynthetic gas exchange and accumulation of phytotoxins in mangrove seedlings in response to soil physico-chemical characteristics associated with waterlogging. Tree Physiol. 1998, 18, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Variation of 50-Year Mean Sea Level Trends at Tsim Bei Tsui, Hong Kong. Available online: https://tidesandcurrents.noaa.gov/sltrends/ (accessed on 7 May 2018).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dataset | Acquisition Date | Resolution | |
|---|---|---|---|
| Remote sensing data | SPOT1 HRV | 21 December 1991 | 20 m |
| 26 December 1993 | |||
| 29 January 1997 | |||
| 22 November 2000 | |||
| SPOT4 HRIVR | 8 November 2002 | 10 m | |
| SPOT5 HRG | 11 December 2004 | 10 m | |
| 21 November 2008 | |||
| 10 January 2011 | |||
| GF-1 | 15 November 2013 | 16 m | |
| 20 February 2014 | |||
| 16 December 2015 |
| Date of Satellite Data | Sources of Reference Data |
|---|---|
| 21 December 1991 | Duke and Khan 1999; High Resolution satellite data in 1991 from Google Earth |
| 26 December 1993 | Duke and Khan 1999; High Resolution satellite data in 1991 from Google Earth |
| 29 January 1997 | Duke and Khan 1999; High Resolution satellite data in 2000 from Google Earth |
| 22 November 2000 | High Resolution satellite data in 2000 from Google Earth |
| 8 November 2002 | High Resolution Satellite data in 2002 and 2003 from Google Earth |
| 11 December 2004 | AFCD data; Wong and Fung 2016; High Resolution satellite data in 2003 and 2004 from Google Earth |
| 21 December 2008 | AFCD data; Wong and Fung 2016; High Resolution satellite data in 2008 from Google Earth |
| 10 January 2011 | Wong and Fung 2016; High Resolution satellite data in 2011 |
| 15 November 2013 | Our field data; Google Earth High Resolution images in 2013 |
| 20 February 2014 | Our field data; Google Earth High Resolution images in 2014 |
| 16 December 2015 | Our field data; Google Earth High Resolution images in 2015 |
| 1991 | 1993 | ||||||||||||
| KO1 | AM | KO2 | AI | PA | UA | KO1 | AM | KO2 | AI | PA | UA | ||
| KO1 | 165 | 8 | 18 | 27 | 91.16% | 75.69% | KO1 | 174 | 8 | 4 | 3 | 86.14% | 92.06% |
| AM | 14 | 98 | 33 | 3 | 64.90% | 66.22% | AM | 6 | 133 | 31 | 2 | 86.92% | 77.33% |
| KO2 | 0 | 16 | 57 | 25 | 41.91% | 58.16% | KO2 | 7 | 12 | 129 | 10 | 75% | 81.65% |
| AI | 2 | 29 | 28 | 111 | 66.87% | 65.29% | AI | 15 | 0 | 8 | 134 | 89.93% | 85.35% |
| OA | 67.98% | KC | 0.57 | OA | 84.32% | KC | 0.79 | ||||||
| 1997 | 2000 | ||||||||||||
| KO1 | AM | KO2 | AI | PA | UA | KO1 | AM | KO2 | AI | PA | UA | ||
| KO1 | 185 | 3 | 16 | 0 | 93.91% | 90.69% | KO1 | 171 | 0 | 2 | 2 | 100% | 97.71% |
| AM | 12 | 194 | 90 | 0 | 95.10% | 65.54% | AM | 0 | 158 | 3 | 49 | 95.18% | 75.24 |
| KO2 | 0 | 7 | 55 | 4 | 32.93% | 83.33% | KO2 | 0 | 2 | 141 | 31 | 87.04% | 81.03% |
| AI | 0 | 0 | 6 | 148 | 97.37% | 96.10% | AI | 0 | 6 | 16 | 59 | 41.84% | 72.84% |
| OA | 80.83% | KC | 0.74 | OA | 82.66% | KC | 0.77 | ||||||
| 2002 | 2004 | ||||||||||||
| KO1 | AM | KO2 | AI | PA | UA | KO1 | AM | KO2 | AI | PA | UA | ||
| KO1 | 162 | 2 | 8 | 8 | 89.50% | 90% | KO1 | 169 | 11 | 5 | 13 | 89.42% | 83.35% |
| AM | 5 | 174 | 4 | 15 | 93.05% | 87.88% | AM | 6 | 171 | 4 | 42 | 88.14% | 76.68% |
| KO2 | 9 | 1 | 125 | 5 | 87.41% | 89.29% | KO2 | 9 | 2 | 154 | 0 | 92.22% | 93.33% |
| AI | 5 | 10 | 6 | 129 | 82..17% | 86% | AI | 5 | 10 | 4 | 103 | 65.19% | 84.42% |
| OA | 88.32% | KC | 0.84 | OA | 84.32% | KC | 0.79 | ||||||
| 2008 | 2011 | ||||||||||||
| KO1 | AM | KO2 | AI | PA | UA | KO1 | AM | KO2 | AI | PA | UA | ||
| KO1 | 189 | 3 | 2 | 2 | 96.92% | 96.43% | KO1 | 190 | 4 | 2 | 9 | 93.14% | 92.68% |
| AM | 4 | 160 | 1 | 28 | 86.96% | 82.90% | AM | 11 | 147 | 3 | 21 | 86.98% | 80.77% |
| KO2 | 0 | 5 | 143 | 23 | 83.63% | 83.63% | KO2 | 1 | 4 | 155 | 23 | 94.51% | 84.70% |
| AI | 2 | 16 | 25 | 140 | 72.54% | 76.50% | AI | 2 | 14 | 4 | 82 | 60.74% | 80.39% |
| OA | 85.06% | KC | 0.80 | OA | 85.42% | KC | 0.80 | ||||||
| 2013 | 2014 | ||||||||||||
| KO1 | AM | KO2 | AI | PA | UA | KO1 | AM | KO2 | AI | PA | UA | ||
| KO1 | 116 | 6 | 3 | 1 | 89.92% | 92.06% | KO1 | 172 | 12 | 0 | 2 | 86% | 92.47% |
| AM | 13 | 147 | 25 | 6 | 91.30% | 76.96% | AM | 28 | 179 | 4 | 2 | 90.86% | 84.04% |
| KO2 | 0 | 0 | 121 | 27 | 74.69% | 81.76% | KO2 | 0 | 0 | 131 | 3 | 95.62% | 97.76% |
| AI | 0 | 8 | 13 | 138 | 80.23% | 86.79% | AI | 0 | 6 | 2 | 147 | 95.45% | 94.84% |
| OA | 83.65% | KC | 0.78 | OA | 91.42% | KC | 0.88 | ||||||
| 2015 | |||||||||||||
| KO1 | AM | KO2 | AI | PA | UA | ||||||||
| KO1 | 134 | 5 | 0 | 0 | 91.78% | 96.40% | |||||||
| AM | 10 | 178 | 0 | 0 | 96.21% | 67.42% | |||||||
| KO2 | 2 | 2 | 74 | 2 | 40.13% | 70% | |||||||
| AI | 0 | 0 | 63 | 23 | 82.14% | 85.19% | |||||||
| OA | 78.03% | KC | 0.70 | ||||||||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, M.; Zhang, H.; Lin, G.; Lin, H.; Tang, D. Zonation and Directional Dynamics of Mangrove Forests Derived from Time-Series Satellite Imagery in Mai Po, Hong Kong. Sustainability 2018, 10, 1913. https://doi.org/10.3390/su10061913
Liu M, Zhang H, Lin G, Lin H, Tang D. Zonation and Directional Dynamics of Mangrove Forests Derived from Time-Series Satellite Imagery in Mai Po, Hong Kong. Sustainability. 2018; 10(6):1913. https://doi.org/10.3390/su10061913
Chicago/Turabian StyleLiu, Mingfeng, Hongsheng Zhang, Guanghui Lin, Hui Lin, and Danling Tang. 2018. "Zonation and Directional Dynamics of Mangrove Forests Derived from Time-Series Satellite Imagery in Mai Po, Hong Kong" Sustainability 10, no. 6: 1913. https://doi.org/10.3390/su10061913
APA StyleLiu, M., Zhang, H., Lin, G., Lin, H., & Tang, D. (2018). Zonation and Directional Dynamics of Mangrove Forests Derived from Time-Series Satellite Imagery in Mai Po, Hong Kong. Sustainability, 10(6), 1913. https://doi.org/10.3390/su10061913