Context Matters: Contrasting Ladybird Beetle Responses to Urban Environments across Two US Regions


1
Environmental Studies Department, University of California, Santa Cruz, Santa Cruz, CA 95064, USA
2
Department of Plant Sciences, University of Göttingen, Göttingen NI 37077, Germany
3
Department of Ecology and Evolutionary Biology, University of Michigan, Ann Arbor, MI 48109, USA
4
Department of Ecology and Evolutionary Biology, Princeton University, Princeton, NJ 08540, USA
*
Author to whom correspondence should be addressed.
Sustainability 2018, 10(6), 1829; https://doi.org/10.3390/su10061829
Submission received: 8 April 2018
/
Revised: 16 May 2018
/
Accepted: 30 May 2018
/
Published: 1 June 2018
(This article belongs to the Special Issue Agroecology in the City: Applying Ecological Principles to Sustainable Urban Agriculture)
Abstract
:Urban agroecosystems offer an opportunity to investigate the diversity and distribution of organisms that are conserved in city landscapes. This information is not only important for conservation efforts, but also has important implications for sustainable agricultural practices. Associated biodiversity can provide ecosystem services like pollination and pest control, but because organisms may respond differently to the unique environmental filters of specific urban landscapes, it is valuable to compare regions that have different abiotic conditions and urbanization histories. In this study, we compared the abundance and diversity of ladybird beetles within urban gardens in California and Michigan, USA. We asked what species are shared, and what species are unique to urban regions. Moreover, we asked how beetle diversity is influenced by the amount and rate of urbanization surrounding sampled urban gardens. We found that the abundance and diversity of beetles, particularly of unique species, respond in opposite directions to urbanization: ladybirds increased with urbanization in California, but decreased with urbanization in Michigan. We propose that in California water availability in gardens and the urbanization history of the landscape could explain the divergent pattern. Thus, urban context is likely a key contributor to biodiversity within habitats and an important consideration for sustainable agricultural practices in urban agroecosystems.
1. Introduction
Urbanization is changing biodiversity patterns and population distributions in cities across the world [1,2]. Urban environments are characterized by changes in abiotic [3] as well as biotic conditions [4]. For example, greater amounts of impervious surface in cities causes urban heat island effects, which increases the temperatures of cities [5] and within urban green spaces [6]. Light pollution from buildings and car traffic extends the duration and intensity of light availability, affecting the circadian rhythms of biotic elements [7]. Irrigation of lawns, parks and gardens adds water resources and maintains the presence of vegetation for organisms to exploit [8,9]. Moreover, the abundance and distribution of species and resources (habitat/food/shelter) in urban areas are often supplemented or altered across time and space [1,10].
Changes in environmental conditions and resource availability have different effects on different taxa and species [1,11]. Some species are able to persist and thrive in altered urban environments, while environmental filters and competition can cause other species to decline [12]. The species that thrive, what some consider “urban exploiters”, are often habitat generalists that are able to live, exploit resources and reproduce in diverse, resource poor environmental conditions [2]. On the other hand, specialist species with particular habitat (food, shelter) requirements may be more sensitive to—and decline with—increasing urbanization because cities do not have the vegetation or resources to support these species [13]. The negative effect(s) of urbanization on species life history and functional traits may lead to biotic homogenization and declines in species richness within urban habitats [14].
The rate at which urbanization occurs (i.e., the speed at which land is converted to impervious surface) could further affect the diversity and distribution of species abundance and richness within urban habitats, and their ability to adapt to certain urban conditions. The percent impervious surface is forecasted to increase by 1.5 million km2 by 2030 [15,16]. Moreover, because cities have distinct development histories, socio-cultural and demographic trends [17], it is important to understand whether and how biodiversity will respond to increasing urbanization (and associated qualitative and quantitative aspects) across multiple urban environmental contexts [16,18]. Elucidating whether certain organisms respond differently or not between unique regions can inform conservation agendas and urban sustainability broadly for various cities across the world [1,19].
Urban agroecosystems such as community and home gardens are high-quality habitats that conserve considerable amounts of biodiversity in cities [20]. These systems are heavily managed by people predominantly for the purpose of cultivating fresh vegetables, fruits, flowers and herbs for self-consumption [21,22,23]. Because urban agroecosystems are usually vegetated and irrigated [24], they provide food and shelter for many arthropods. Certain arthropod groups, for example pollinators and natural enemies, are in turn important for providing ecosystem services like crop pollination and pest control. Previous studies have shown that these arthropod groups are less abundant in gardens where surrounding levels of urbanization are high [25,26]. However, groups respond differentially to urbanization and at different spatial scales [27,28]. Some arthropod groups and species in urban environments including urban agroecosystems may actually increase with urbanization [29,30]. For example, insect pollinator species diversity is greater in some urban regions compared to surrounding suburban and agricultural areas [30,31,32]. Currently, it is unclear whether these patterns are regional phenomena or if these trends are generalizable to other urban regional contexts. We argue that this question warrants further investigation, requiring research that draws comparisons of arthropod biodiversity across spatially distinct regions. Yet studies in urban agroecosystems that compare and synthesize findings across regions with different environmental conditions are rare [33].
Here, we combine data on ladybird beetle abundance and species richness collected from comparable urban agroecosystems in two distinct geographical regions to test whether the response of ladybird beetles to urbanization differs by the environmental context and urbanization history. Ladybird beetles are charismatic arthropods in agroecosystems that provide key natural pest control services, particularly of herbivorous aphids, mites and scale insects [34,35,36]. Because urban agroecosystems are situated among dense human populations, they tend towards organic, environmentally friendly, and human-health-conscious forms of management [37]. Thus, natural pest control is particularly important for these agroecosystems. In this study, we asked: (1) Does the relationship between urbanization (percent impervious surface, rate of development) and predator (ladybird beetle) systems in urban agroecosystems change with environmental context (region)? (2) Which species are shared by, and which are unique to urban agroecosystems of different regions? (3) Do shared species respond differently to urbanization measures in the region than unique species?
2. Methods
2.1. Study Regions
We worked in two regions in the USA—California and Michigan—to collect ladybird beetle data in urban community gardens in these regions (Figure 1). In California, we collected ladybird beetle data from 18 urban gardens in the California central coast in Santa Clara, Santa Cruz, and Monterey counties, which have estimated population densities of 274, 232, and 50 people/sq. km, respectively (2010–2014 U.S. census period) [38]. In Michigan, we collected ladybird beetle data from 13 urban gardens in Washtenaw county, which has an estimated population density of 192 people/sq. km [38]. The gardens in both regions are surrounded by different amounts of impervious surface. Moreover, the gardens differ in vegetation and groundcover composition and structure, but because they are all community gardens, differences in composition and structure are assumed to be relatively similar between regions. The gardens range in size from 444 to 15,525 m2 in California and from 54 to 8778 m2 in Michigan, and are separated by approximately >2 km in California and >0.5 km in Michigan. All of the gardens have been cultivated for 1 to 47 years and do not use chemical pesticides and insecticides.
2.2. Ladybird Beetle Sampling
To assess ladybird beetle communities in the gardens, we used visual and trapping methods in both regions. In California, we sampled for adult beetles with visual surveys and sticky traps within 20 m2 plots at the center of each of the 18 gardens six times during summer 2014 (17–20 June, 7–10 July, 27–30 July, 19–21 August, 8–10 September, 29 September–1 October). Within the 20 m2, we visually surveyed vegetation and ground cover for adult beetles in eight randomly placed 0.5 × 0.5 m sub-plots. We collected all individuals observed and stored them in vials with ethyl alcohol. At four random locations within the plots, we also placed a 3″ × 5″ yellow sticky trap card (BioQuip Products Inc., Compton, CA, USA) on a galvanized wire stake for 24 h. In Michigan, we sampled for adult beetles by visually surveying five sentinel potted pea plants (Pisum sativum var. Dwarf grey) placed at each of the 13 gardens in Washtenaw County. Any ladybird beetle adults on plants were counted and identified to species. In addition, we swept surrounding vegetation in gardens for adult ladybird beetles using 10 full sweeps of a 0.2 m diameter net. All Michigan gardens were surveyed twice a week from 14 May to 20 July 2012. Sampling effort was consistent in all sites in each region throughout the sampling periods: in Michigan, the same two researchers conducted the sampling within the respective area for 30 min; in California, the same researcher conducted the sampling within the respective area for 60 min. The slight differences in sampling methods and years sampled between regions introduces some limitations discussed later in our conclusions.
We identified all ladybird beetles on traps and in vials to species using identification guides [39] and online resources [40,41]. Total abundance for each site for each species, total species richness, and total species diversity (Shannon’s Diversity Index H) was tabulated across the months. Species diversity includes the relative distribution of species’ abundances and was calculated using the vegan package in R [42]. For the analysis, we categorized species present in both California and Michigan as “shared species”, and categorized species that were not both present in California and Michigan as “unique species”.
2.3. Urban Landscape Analysis
To measure current levels of urbanization and to assess urbanization history, we summarized (1) the mean percent impervious surface surrounding gardens, and (2) the rate at which percent impervious surface has increased over time. For both regions (California, Michigan), we used the package “raster” in R (v 3.4.1) [43,44] to calculate the mean percent impervious surface within buffers of 10, 100, 500, 1000, 2000, 3000 m spatial scales surrounding each garden site based on land cover data from the US Geological Survey’s National Land Cover Database (NLCD) 2011 Percent Developed Imperviousness dataset [45]. Here, a high total percent impervious surface indicates higher degrees of urbanization, and a low percent impervious surface indicates low degrees of urbanization. To calculate the rate of percent impervious surface change over time (henceforth “urbanization rate”), we collected this data at three time periods, as provided by the NLCD: 2001, 2006, 2011. Urbanization rate was calculated as the slope of the regression across these three time periods.
2.4. Statistical Analysis
We ran species accumulation curves to test whether species richness had been sufficiently sampled in both California and Michigan. The expected number of species in each geographic region was calculated using a sample-based rarefaction method known as the Mao Tau estimator [46]. Both regions showed evidence that richness was sufficiently sampled, exhibiting saturation in their species accumulation curves (Figure S1).
We first modeled abundance and richness for each region at multiple spatial scales to determine the best scale at which ladybird beetles respond to urbanization. We built seven generalized linear models (GLM) at 0, 10, 100, 500, 1000, 2000, 3000 m spatial scales assuming Poisson error distributions for count data. The model with the lowest Akaike Information Criterion (AIC) was selected as the best spatial scale for each region [47].
Urbanization rate was calculated by taking the slopes of linear regressions between time and impervious surface (NLCD: 2001, 2006, 2011) for each garden at a scale of 500 m. This was the buffer scale determined earlier to be significant for Michigan. California beetles best responded to impervious cover at 100 m, but at this scale urbanization rate did not vary by garden. Thus, we only analyzed effects of urbanization rate on ladybird beetle abundance, species richness and species diversity at 500 m for both regions. We also ran Pearson’s r tests between values of urbanization rate and impervious surface at both 100 and 500 m to test for correlations between explanatory variables. Urbanization rate and impervious surface were not significantly correlated (Table S1).
To determine whether ladybird beetles significantly responded to percent impervious surface or urbanization rates, we constructed GLMs at the spatial scale appropriate for the region and predictor variable as described above. Abundance and species richness GLMs assumed Poisson error distributions, and diversity GLMs assumed Gaussian. All GLMs were then fit by Laplace approximation and goodness of fit determined by Wald Z tests [47]. This is what we refer to in the text as Poisson and linear regressions.
3. Results
We found 16 ladybird beetle species in California and eight species in Michigan over the sampling periods across the regions (Table 1). Species diversity index values were higher in California (ranging from 1.3 to 2.5 for all species) than in Michigan (ranging from 0 to 1.5). Only four species were shared by California and Michigan, including: C. septempunctata, C. Sanguinea, H. axyridis and O. v-nigrum. Thus, 12 species in California and four species in Michigan were unique to that region.
Total ladybird beetle species abundance, richness and diversity (shared and unique species) were best explained by percent impervious surface at a 100 m spatial scale in California, but were best explained at a 500 m spatial scale in Michigan (Table S2). Total ladybird beetle species abundance and species diversity significantly increased with percent impervious surface in California, but significantly declined with percent impervious surface in Michigan (Table 2; Figure 2). Total species richness also generally increased in California and decreased in Michigan with percent impervious surface (Table 2). The divergent trend between regions was similar for shared species: shared species abundance, richness and diversity significantly increased in California with greater impervious surface, but were not significant for response measures in Michigan (Table 2). Unique species abundance also significantly increased with impervious cover in California, while unique species abundance significantly decreased in Michigan (Table 2; Figure 2). Of note, impervious surface cover gradients were comparable between California and Michigan.
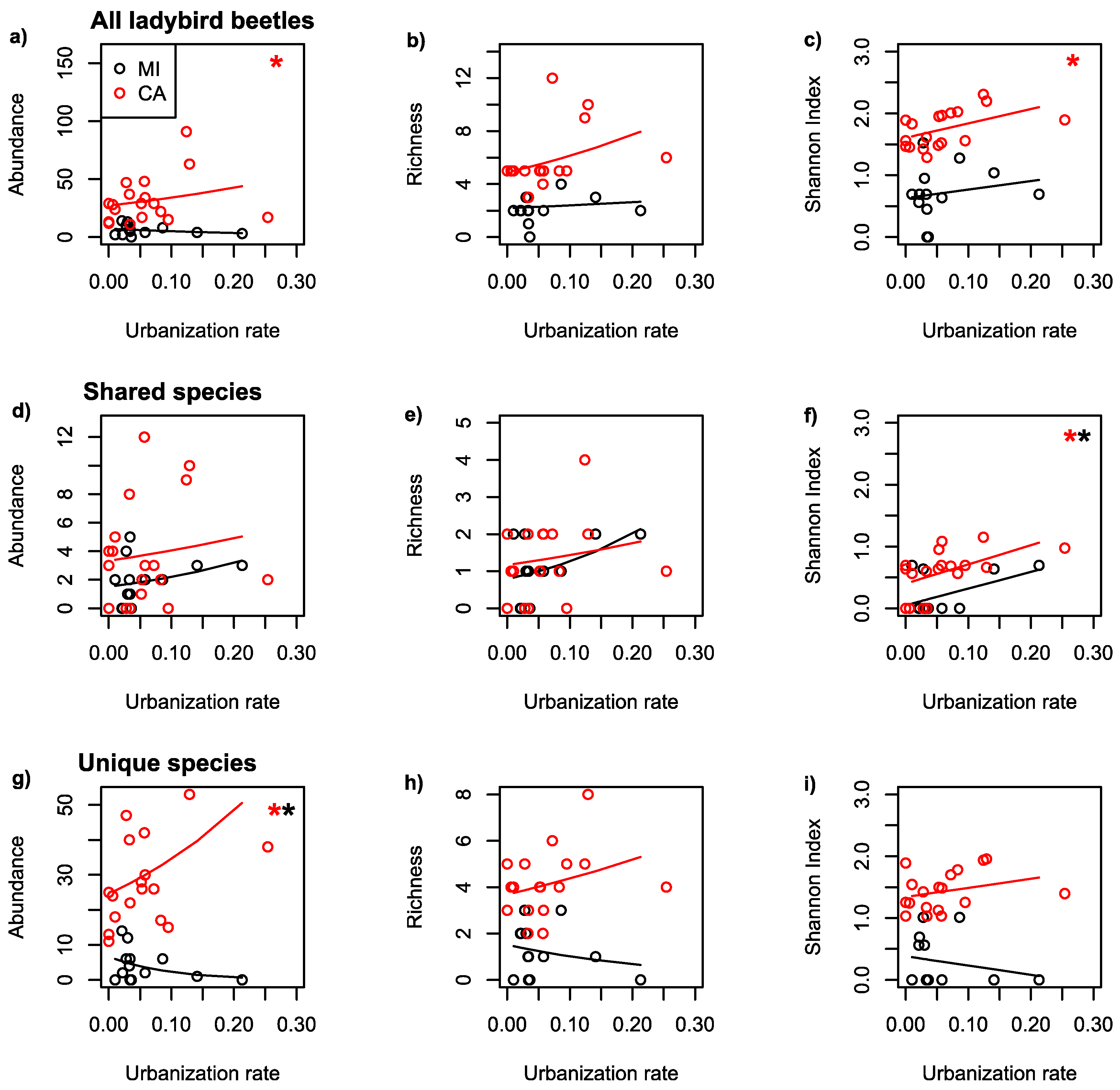
In response to the rate of urbanization surrounding gardens, ladybird beetle abundance and species diversity increased with faster urbanization rates in California (Table 2; Figure 3). Whereas, in Michigan, it was not significant for all response measures (Table 2; Figure 3). Species diversity of shared ladybird species significantly increased with faster urbanization rates for both regions (Table 2; Figure 3). The abundance of unique species significantly increased in California and decreased in Michigan with faster urbanization rates (Table 2; Figure 3).
4. Discussion
The influence of urbanization on biodiversity can change with environmental (regional) context. This comparative study between two urban regions in the US—California and Michigan—shows that organisms respond differently to urbanization depending on region. Ladybird beetles have a contrasting response to the intensity of urbanization as well as the rate at which urbanization occurs in different regional contexts, and we found only one unidirectional relationship between species diversity and urbanization rate between regions. The contrasting response is most apparent in the abundance of all species and unique species. We hypothesize that the effect of urbanization on unique species is driving this divergent pattern.
Our first question was whether the relationship between urbanization and ladybird beetles in urban agroecosystems changes with environmental context. We found that urban gardens are supporting more abundant and diverse ladybird beetle populations in more urban areas in California, while in Michigan, ladybird beetles in urban gardens decline in abundance, species richness and diversity with increasing amounts of impervious cover and faster urbanization rates in most instances. Urbanization is clearly driving the abundance, species richness and behavior of ladybird beetles in California, as we have found in previous studies [29,50]. However, we show that this is not the case in another environmental context (Michigan). Though not specifically sampled in urban gardens, a majority of taxa decline in abundance and species richness with urbanization [2]. This is particularly apparent for vertebrates [18], but is also often the case for invertebrates [51,52,53]. In contrast, plant species generally increase with urbanization presumably because non-native species introductions outweigh extinctions in this group and because plants have smaller geographical ranges than mobile organisms with high dispersal abilities [54]. Organisms with larger ranges may be more sensitive to urbanization because urbanization can fragment migratory corridors and impede dispersal [55,56]. Given that we observed divergent geographical responses to urbanization most strongly for unique ladybird species and one similar response to urbanization by shared species, differences in dispersal ranges could possibly explain our results. If unique beetles to California have larger geographical distributions—often related to species dispersal ability and range size [57]—than beetles unique to Michigan, our results would be consistent with the dispersal hypothesis. However, we did not find strong evidence for this hypothesis in our results, because the reported geographic distribution for these species is relatively narrow for California beetles versus Michigan beetles (Table 1).
It is important to note, however, that though general trends in taxonomic responses to urbanization exist, all taxa that have been examined at multiple spatial scales or contexts exhibit some degree of divergence in responses to urbanization (6.9 to 33.3% of studies in a given taxon report different responses to urbanization depending on context) [2,28,30,58,59]. At larger spatial scales, urbanization is correlated with dense human populations that also coincide historically with nutrient-rich and biodiverse regions [60,61]. McKinney suggests that this can produce an apparent positive effect of urbanization on species abundance and richness [2]. Moreover, the longer periods of warm temperatures due to urbanization (i.e., urban heat island effects) may increase insect population abundance because of increased reproductive capacity [62], a common physiological response for arthropods [63]. At smaller spatial scales, local effects including management intensity and the destruction of habitat and pollution may impose negative effects of urbanization on species abundance and richness [64]. However, our results do not support this hypothesis given that ladybird beetles responded negatively to urbanization at larger spatial scales in Michigan, and positively at smaller spatial scales in California (Figure 2, Table 1).
The rate of urbanization, not only the amount of impervious surface, was important for explaining beetle abundance and diversity but exhibited different patterns depending on the group. Interestingly, while the abundance and diversity of unique species similarly diverged in regional responses to urbanization rate as to amount of impervious surface, shared species all showed positive unidirectional responses to urbanization rate. The predictor variables are not significantly correlated, and thus could theoretically have divergent effects (Table S1). Urbanization rates were actually relatively similar in California and in Michigan (at 500 m), which could explain why shared species had similar responses in each region while unique species had opposite responses. The differences in ladybird biodiversity between regions is therefore likely best explained by a species-level response: specifically, the response of unique species to California versus Michigan. Indeed, the strongest pattern from our study is the response of unique species to both the percent impervious surface and to the rate of urbanization, with abundance of unique beetles significantly increasing in California but decreasing in Michigan. This suggests that there are environmental filters at regional as well as local scales for species’ traits that allow them to thrive in more urban areas and habitats [12,65] in California, and that in Michigan, those species are not present. Only species with traits that allow them to persist in urban environments should similarly increase with increasing rates of urbanization across regions. Indeed, traits including habitat, diet breadth and foraging efficiency have explained the global expansion of the ladybird species Harmonia axyridis into urban areas [66,67]. The similar unidirectional response of shared species to urbanization rate in our study supports this hypothesis.
Thus, our results may be explained by the legacy of land use change in each region and species’ life histories/traits. California gardens have more species and more unique species that are not found in Michigan. Michigan gardens have fewer species, and 50% of those species were also found in California gardens. Most of the shared species across regions are aphidophagous (eat aphids), while in California the unique species to the region largely eat mites, scales and fungus (Table 1). For example, the fungus feeder Psyllobora vigintimaculata is very abundant in California and has a different life history than other species (Figure S2). The presence of species with these feeding preferences could be because of the industrial agricultural crops grown within and near our urban garden sites in California, historically and currently. Some of the region was once an orchard landscape, known as the “Valley of Heart’s Delight”, that has historically grown diverse fruit and nut trees [68]. Fruit trees and landscaped shrubs are often prone to scale, mite and mealybug pest damage along with crops like strawberries and tomatoes [69]. As discussed earlier, human populations preferentially settle in biodiverse areas (or “biodiversity hotspots”) [61,70], and cities can coincide with threatened species distribution [71,72], possibly explaining positive relationships between ladybirds and urbanity. The legacy of agriculture in turn has permanent effects on ecosystems, and the influx of nutrients and irrigation can also create biodiversity hotspots [60]. Natural enemies were historically introduced from e.g., New Zealand and Australia for biological control of pests in the orchards. For example, the Dusky ladybird beetle, Nephus binaevatus, was released into California from New Zealand in 1922 to help control mealybugs in orchards [73]. This non-native species is unique to the California garden sites, particularly in very urban sites in Santa Clara county that were once orchards. Our sampled garden sites in California contain fruit trees such as citrus and stone fruit. Thus, some of the unique ladybird beetles like N. binaevatus and also Coccinella septempuctata are legacies of agricultural industrialization and urbanization unique to California, and/or may be present in gardens due to the availability of their prey/host.
The ladybird beetles in California—particularly those that are unique—may be more likely able to withstand environmental disturbance because they have been historically used in human-dominated systems for e.g., biological control. Non-native species to a region often have a greater ability to survive in a variety of habitats—including disturbed habitats—than native species [74]. In disturbed urban landscapes, more abundant species are habitat generalists and/or non-native to a region [2,75]. These species have ecological traits that allow them to exploit resources and persist [76], and environmental filters have been used to explain taxonomic differences between urban habitats [12,77]. Most (though not all) of the ladybird species that we observed in Michigan sites are native species from the greater Northeast and Midwest region (Table 1). This could explain why abundance and richness of beetles declines with urbanization in Michigan: many of the species are native and are less likely to survive and adapt to environmental disturbances like urbanization as non-native and invasive species [78].
Abiotic factors associated with urbanization and different environmental contexts may better explain the contrasts in ladybird diversity patterns in California and Michigan agroecosystems. Ladybirds must avidly consume water (e.g., dew, rain) for their survival [79], and water availability often drives their movement ecology and life cycle [80]. Thus, climate patterns (temperature, precipitation) can affect ladybird distribution [39,79], and significant climatic contrasts between regions could explain divergent patterns in ladybird abundance and species richness in gardens. In comparison to Michigan, which has a temperate climate with four defined seasons, California has on average warmer temperatures throughout the year and two seasons, one dry (summer; April to September) and one wet (winter; October to March). In California, urban gardens in the summer dry months (with <1 cm precipitation per month much in the form of fog) may provide an important irrigated habitat in comparison to the surrounding urban matrix. Indeed, supplemental irrigation in cities maintains and supports biodiversity during drought events [9,81,82], as California experienced during the time of this study. Michigan has an opposite precipitation pattern with relatively wet summers, receiving on average 8 to 9 cm/month in the summer [83]. Thus, limited water availability in the urban matrix may not affect arthropod distributions as strongly in Michigan. On the other hand, urban gardens in California may act as critical sources of water, food, and shelter and promote greater ladybird beetle foraging and fecundity rates [79], ultimately increasing their populations. If this is the case, then urban gardens in California may function as sources of populations rather than sinks, as has previously been suggested for biodiversity in urban habitats [1,25,84,85]. To test this hypothesis, we suggest examining habitats outside of gardens in these respective regions and their local conditions (in water availability, prey) to see whether ladybird abundance and diversity conservation is greater within gardens versus outside of gardens in natural habitats [86] and if it is explained by local conditions.
The slight differences in sampling methods and years sampled between regions are caveats in our study. Specific sampling methods as well as annual variation within regions may influence differences between regions. Moreover, although we only focused on landscape factors to better understand the ecology of urban agroecosystems across regions, local garden attributes including vegetation characteristics could also be important for species distribution [87,88]. Indeed, these caveats are generally the cruces of comparative studies and synthesis research. Although such studies are critical, comparing biodiversity assessments from multiple regions and research groups are inherently challenging to conduct and to analyze due to inconsistencies among methodological approaches in ecology, and research funding and timing. Given these limitations, this work shows strong relationships between landscape factors and local biodiversity with available comparable data across two regions. This is a first step towards a greater understanding of regional effects on urban biodiversity.
5. Conclusions
Ladybird beetles provide important pest control services and understanding their responses to urbanization can inform sustainable agricultural management in urban gardens. In this comparative study using urban agroecosystems as a field system, we show that organisms like ladybirds exhibit opposite responses to urbanization in different regional environmental contexts. Thus, we cannot expect universal responses of organism groups to urbanization. Based on previous research, region-specific responses to urbanization appear ubiquitous. Spatial scale is currently presumed to have a positive correlation with species abundance and diversity in increasingly urban environments. Here, we show that at least for ladybird beetles that is not the case. We suggest that region-specific responses are more likely due to the environmental factors (abiotic, biotic), urbanization history of the region, and the natural histories of species unique to each region. The natural history of species that are unique to urban agroecosystems could impart insight into how urban areas may be designed to conserve species that are more sensitive to urban environments. Future urban agroecology research should seek to draw more comparisons across distinct regions rather than generalizing responses of biodiversity to urbanization.
Supplementary Materials
The following are available online at https://www.mdpi.com/2071-1050/10/6/1829/s1, Figure S1: Species accumulation curves, Figure S2: Histograms of ladybird species sampled, Table S1: Correlations between explanatory variables, Table S2: Ladybird beetle sensitivity to percent impervious surface at various spatial scales.
Author Contributions
All authors conceived of and designed the study and performed the data collection; T.W.O. and K.L. analyzed the data; M.H.E. drafted the manuscript. All authors provided feedback on and contributed significantly to the manuscript.
Acknowledgments
We thank the field and lab work provided by P. Bichier, Y. Bravo, J. Burks, H. Cohen, V. De la Fuente, K. Ennis, A. García, P. Gillette, M. Hash, R. Hruska, B. Ferguson, M. MacDonald, L. Marin, H. Morales, M.E. Narvaez Cuellar, M. Otoshi, M. Plascencia, R. Quistberg, G. Santiz Ruiz, R. Schreiber-Brown, S-S. Thomas, L. Hawkes, B. Madden, A. Ho and D. Kowalsky. We thank the two anonymous reviewers for their helpful comments on the manuscript. Thank you to S. Philpott, J. Vandermeer, and I. Perfecto for considerable research support. We thank the community gardens and gardeners for hosting our research in California: Aptos Community Garden, Beach Flats Community Garden, Berryessa Community Garden, the Center for Agroecology and Sustainable Food Systems, Chinatown Community Garden, Coyote Creek Community Garden, El Jardín at Emma Prusch Park, The Forge at Santa Clara University, Giving Garden at Faith Lutheran Church, Homeless Garden Project, La Colina Community Garden, Laguna Seca Community Garden, Live Oak Green Grange Community Garden, MEarth at Carmel Valley Middle School, Mi Jardín Verde Community Garden at All Saints’ Episcopal Church, Our Green Thumb Garden at Middlebury Institute of International Studies, and Salinas Community Garden at St. George’s Episcopal Church. We thank Project Grow Community Gardens for hosting our research in Michigan. This research was financially supported by the Center for Agroecology and Sustainable Food Systems and the Environmental Studies Department at the University of California, Santa Cruz. M.H.E. was supported by a National Science Foundation Graduate Research Fellowship Grant No. 174835. T.W.O was funded by the Department of Ecology and Evolutionary Biology and the Rackham Graduate School at the University of Michigan. This material is based upon work supported by the National Science Foundation under Grant No. 1711167 to T.W.O.
Conflicts of Interest
The authors declare no conflict of interest. The founding sponsors had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, and in the decision to publish the results. The data was collected with site approval.
References
- McKinney, M.L. Urbanization, biodiversity, and conservation. Bioscience 2002, 52, 883–890. [Google Scholar] [CrossRef]
- McKinney, M.L. Effects of urbanization on species richness: A review of plants and animals. Urban Ecosyst. 2008, 11, 161–176. [Google Scholar] [CrossRef]
- Kaye, J.P.; Groffman, P.M.; Grimm, N.B.; Baker, L.A.; Pouyat, R.V. A distinct urban biogeochemistry? Trends Ecol. Evol. 2006, 21, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Grimm, N.B.; Faeth, S.H.; Golubiewski, N.E.; Redman, C.L.; Wu, J.; Bai, X.; Briggs, J.M. Global change and the ecology of cities. Science 2008, 319, 756–760. [Google Scholar] [CrossRef] [PubMed]
- Oke, T.R. City size and the urban heat island. Atmos. Environ. 1973, 7, 769–779. [Google Scholar] [CrossRef]
- Lin, B.B.; Egerer, M.H.; Liere, H.; Jha, S.; Bichier, P.; Philpott, S.M. Local- and landscape-scale land cover affects microclimate and water use in urban gardens. Sci. Total Environ. 2018, 610–611, 570–575. [Google Scholar] [CrossRef] [PubMed]
- Longcore, T.; Rich, C. Ecological light pollution. Front. Ecol. Environ. 2004, 2, 191–198. [Google Scholar] [CrossRef]
- Hope, D.; Gries, C.; Zhu, W.; Fagan, W.F.; Redman, C.L.; Grimm, N.B.; Nelson, A.L.; Martin, C.; Kinzig, A. Socioeconomics drive urban plant diversity. Proc. Natl. Acad. Sci. USA 2003, 100, 8788–8792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faeth, S.H.; Warren, P.S.; Shochat, E.; Marussich, W.A. Trophic dynamics in urban communities. Bioscience 2005, 55, 399–407. [Google Scholar] [CrossRef]
- Pickett, S.T.A.; Cadenasso, M.L. Altered resources, disturbance, and heterogeneity: A framework for comparing urban and non-urban soils. Urban Ecosyst. 2009, 12, 23–44. [Google Scholar] [CrossRef]
- Faeth, S.H.; Saari, S.; Bang, C. Urban Biodiversity: Patterns, Processes and Implications for Conservation, eLS 2012; John Wiley Sons, Ltd.: Hoboken, NJ, USA, 2012. [Google Scholar] [CrossRef]
- Aronson, M.F.J.; Nilon, C.H.; Lepczyk, C.A.; Parker, T.S.; Warren, P.S.; Cilliers, S.S.; Goddard, M.A.; Hahs, A.K.; Herzog, C.; Katti, M.; et al. Hierarchical filters determine community assembly of urban species pools. Ecology 2016, 97, 2952–2963. [Google Scholar] [CrossRef] [PubMed]
- Mackin-Rogalska, R.; Pinowski, J.; Solon, J.; Wojcik, Z. Changes in vegetation, avifauna, and small mammals in a suburban habitat. Pol. Ecol. Stud. 1988, 14, 293–330. [Google Scholar]
- McKinney, M.L.; Lockwood, J.L. Biotic homogenization: A few winners replacing many losers in the next mass extinction. Trends Ecol. Evol. 1999, 5347, 450–453. [Google Scholar] [CrossRef]
- Seto, K.C.; Fragkias, M.; Gu, B. A Meta-Analysis of Global Urban Land Expansion. PLoS ONE 2011, 6, e23777. [Google Scholar] [CrossRef] [PubMed]
- Seto, K.C.; Guneralp, B.; Hutyra, L.R. Global forecasts of urban expansion to 2030 and direct impacts on biodiversity and carbon pools. Proc. Natl. Acad. Sci. USA 2012, 109, 16083–16088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, S.S. Incorporating critical elements of city distinctiveness into urban biodiversity conservation. Biodivers. Conserv. 2015, 24, 683–700. [Google Scholar] [CrossRef]
- Aronson, M.F.; La Sorte, F.A.; Nilon, C.H.; Katti, M.; Goddard, M.A.; Lepczyk, C.A.; Warren, P.S.; Williams, N.S.; Cilliers, S.; Clarkson, B.; et al. A global analysis of the impacts of urbanization on bird and plant diversity reveals key anthropogenic drivers. Proc. R. Soc. B 2014, 281, 20133330. [Google Scholar] [CrossRef] [PubMed]
- McKinney, M.L. Urbanization as a major cause of biotic homogenization. Biol. Conserv. 2006, 127, 247–260. [Google Scholar] [CrossRef]
- Goddard, M.A.; Dougill, A.J.; Benton, T.G. Scaling up from gardens: Biodiversity conservation in urban environments. Trends Ecol. Evol. 2010, 25, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Mougeot, L.J.A. Urban agriculture: Definition, presence, potentials and risks, and policy challenges. In Growing Cities, Growing Food: Urban Agriculture on the Policy Agenda. A Reader on Urban Agriculture; Bakker, N., Dubbeling, M., Gündel, S., Sabel-Koschella, U., De Zeeuw, H., Eds.; DSE/ETC: Feldafing, Germany, 2000; pp. 99–117. [Google Scholar]
- Lovell, S.T. Multifunctional urban agriculture for sustainable land use planning in the United States. Sustainability 2010, 2, 2499–2522. [Google Scholar] [CrossRef]
- Zezza, A.; Tasciotti, L. Urban agriculture, poverty, and food security: Empirical evidence from a sample of developing countries. Food Policy 2010, 35, 265–273. [Google Scholar] [CrossRef]
- Faeth, S.H.; Bang, C.; Saari, S. Urban biodiversity: Patterns and mechanisms. Ann. N. Y. Acad. Sci. 2011, 1223, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Connor, E.F.; Hafernik, J.; Levy, J.; Moore, V.L.; Rickman, J.K. Insect conservation in an urban biodiversity hotspot: The San Francisco Bay Area. J. Insect Conserv. 2002, 6, 247–259. [Google Scholar] [CrossRef]
- Quistberg, R.D.; Bichier, P.; Philpott, S.M. Landscape and local correlates of bee abundance and species richness in urban gardens. Environ. Entomol. 2016, 45, 592–601. [Google Scholar] [CrossRef] [PubMed]
- Bennett, A.B.; Gratton, C. Measuring natural pest suppression at different spatial scales affects the importance of local variables. Environ. Entomol. 2012, 41, 1077–1085. [Google Scholar] [CrossRef] [PubMed]
- Egerer, M.H.; Arel, C.; Otoshi, M.D.; Quistberg, R.D.; Bichier, P.; Philpott, S.M. Urban arthropods respond variably to changes in landscape context and spatial scale. J. Urban Ecol. 2017, 3. [Google Scholar] [CrossRef] [Green Version]
- Egerer, M.H.; Bichier, P.; Philpott, S.M. Landscape and local habitat correlates of lady beetle abundance and species richness in urban agriculture. Ann. Entomol. Soc. Am. 2016, 110, 97–103. [Google Scholar] [CrossRef]
- Hall, D.M.; Camilo, G.R.; Tonietto, R.K.; Ollerton, J.; Ahrné, K.; Arduser, M.; Ascher, J.S.; Baldock, K.C.R.; Fowler, R.; Frankie, G.; et al. The city as a refuge for insect pollinators. Conserv. Biol. 2017, 31, 24–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldock, K.C.R.; Goddard, M.A.; Hicks, D.M.; Kunin, E.; Mitschunas, N.; Osgathorpe, L.M.; Potts, S.G.; Robertson, K.M.; Scott, A.V.; Stone, G.N.; et al. Where is the UK’s pollinator biodiversity? The importance of urban areas for flower-visiting insects. Proc. R. Soc. B 2015, 282, 20142849. [Google Scholar] [CrossRef] [PubMed]
- Cane, J.H.; Minckley, R.L.; Kervin, L.J.; Roulston, T.H.; Williams, N.M. Complex responses within a desert bee guild (Hymenoptera: Apiformes) to urban habitat fragmentation. Ecol. Appl. 2006, 16, 632–644. [Google Scholar] [CrossRef]
- Lin, B.B.; Philpott, S.M.; Jha, S. The future of urban agriculture and biodiversity-ecosystem services: Challenges and next steps. Basic Appl. Ecol. 2015, 16, 189–201. [Google Scholar] [CrossRef]
- Cardinale, B.J.; Harvey, C.T.; Gross, K.; Ives, A.R. Biodiversity and biocontrol: Emergent impacts of a multi-enemy assemblage on pest suppression and crop yield in an agroecosystem. Ecol. Lett. 2003, 6, 857–865. [Google Scholar] [CrossRef]
- Obrycki, J.J.; Harwood, J.D.; Kring, T.J.; O’Neil, R.J. Aphidophagy by Coccinellidae: Application of biological control in agroecosystems. Biol. Control 2009, 51, 244–254. [Google Scholar] [CrossRef]
- Evans, E.W. Lady beetles as predators of insects other than Hemiptera. Biol. Control 2009, 51, 255–267. [Google Scholar] [CrossRef]
- Oberholtzer, L.; Dimitri, C.; Pressman, A. Organic Agriculture in U.S. Urban Areas: Building Bridges between Organic Farms and Education. In Proceedings of the IFOAM Organic World Congress 2014, Istanbul, Turkey, 13–15 October 2014. [Google Scholar]
- US Census Bureau. American Community Survey 5-Year Estimates, County; US Census Bureau: Suitland, MD, USA, 2014.
- Gordon, R.D. The Coccinellidae (Coleoptera) of America north of Mexico. J. N. Y. Entomol. Soc. 1985, 93. Available online: https://www.zin.ru/Animalia/Coleoptera/addpages/Andrey_Ukrainsky_Library/References_files/Gordon85a.htm (accessed on 1 March 2018).
- Discover Life Discover Life. Available online: www.discoverlife.org (accessed on 10 September 2015).
- Iowa State University Department of Entomology BugGuide. Available online: www.bugguide.net (accessed on 10 October 2014).
- Oksanen, J. Community Ecology Package. Available online: https://github.com/vegandevs/vegan (accessed on 15 April 2018).
- Hijmans, R.J. Geographic Data Analysis and Modeling; R Package Raster Version 2.6-7; R Foundation: Vienna, Austria, 2015; Available online: https://cran.r-project.org/web/packages/raster/ (accessed on 1 October 2017).
- R Development Core Team. R development core team. In R: A Language and Environment for Statistical Computing; R Foundation: Vienna, Austria, 2016; Volume 55, pp. 275–286. [Google Scholar]
- Xian, G.Z.; Homer, C.G.; Dewitz, J.; Fry, J.; Hossain, N.; Wickham, J. Change of impervious surface area between 2001 and 2006 in the conterminous United States. Photogramm. Eng. Remote Sens. 2011, 77, 758–762. [Google Scholar]
- Colwell, R.K.; Chao, A.; Gotelli, N.J.; Lin, S.; Mao, C.X.; Chazdon, R.L.; Longino, J.T. Models and estimators linking rarefaction, extrapolation and comparison of assemblages. J. Plant Ecol. 2012, 5, 3–21. [Google Scholar] [CrossRef]
- Bolker, B.M.; Brooks, M.E.; Clark, C.J.; Geange, S.W.; Poulsen, J.R.; Stevens, M.H.H.; White, J.-S.S. Generalized linear mixed models: A practical guide for ecology and evolution. Trends Ecol. Evol. 2009, 24, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Casey, T.L. A revision of the American Coccinellidae. J. N. Y. Entomol. Soc. 1899, 7, 71–169. [Google Scholar]
- Hodek, I.; Honěk, A. Ecology of Coccinellidae; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1996; Volume 54. [Google Scholar]
- Egerer, M.H.; Liere, H.; Bichier, P.; Philpott, S.M. Cityscape quality and resource manipulation affect natural enemy biodiversity in and fidelity to urban agroecosystems. Landsc. Ecol. 2018, 1–14. [Google Scholar] [CrossRef]
- McIntyre, N.E.; Rango, J.; Fagan, W.F.; Faeth, S.H. Ground arthropod community structure in a heterogeneous urban environment. Landsc. Urban Plan. 2001, 52, 257–274. [Google Scholar] [CrossRef] [Green Version]
- Morse, C.C.; Huryn, A.D.; Cronan, C. Impervious surface area as a predictor of the effects of urbanization on stream insect communities in Maine, USA. Environ. Monit. Assess. 2003, 89, 95–127. [Google Scholar] [CrossRef] [PubMed]
- Fattorini, S. Insect extinction by urbanization: A long term study in Rome. Biol. Conserv. 2011, 144, 370–375. [Google Scholar] [CrossRef]
- Gaston, K.J.; Quinn, R.M.; Blackburn, T.M.; Eversham, B.C. Species-range size distributions in Britain. Ecography 1998, 21, 361–370. [Google Scholar] [CrossRef]
- Gibb, H.; Hochuli, D.F. Habitat fragmentation in an urban environment: Large and small fragments support different arthropod assemblages. Biol. Conserv. 2002, 106, 91–100. [Google Scholar] [CrossRef]
- Parris, K.M. Urban amphibian assemblages as metacommunities. J. Anim. Ecol. 2006, 75, 757–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lester, S.E.; Ruttenberg, B.I.; Gaines, S.D.; Kinlan, B.P. The relationship between dispersal ability and geographic range size. Ecol. Lett. 2007, 10, 745–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lizée, M.H.; Manel, S.; Mauffrey, J.F.; Tatoni, T.; Deschamps-Cottin, M. Matrix configuration and patch isolation influences override the species-area relationship for urban butterfly communities. Landsc. Ecol. 2012, 27, 159–169. [Google Scholar] [CrossRef]
- Penone, C.; Kerbiriou, C.; Julien, J.F.; Julliard, R.; Machon, N.; Le Viol, I. Urbanisation effect on Orthoptera: Which scale matters? Insect Conserv. Divers. 2013, 6, 319–327. [Google Scholar] [CrossRef]
- Dupouey, J.L.; Dambrine, E.; Laffite, J.D.; Moares, C. Irreversible impact of past land use on forest soils and biodiversity. Ecology 2002, 83, 2978–2984. [Google Scholar] [CrossRef]
- Pautasso, M. Scale dependence of the correlation between human population presence and vertebrate and plant species richness. Ecol. Lett. 2007, 10, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Meineke, E.K.; Dunn, R.R.; Sexton, J.O.; Frank, S.D. Urban warming drives insect pest abundance on street trees. PLoS ONE 2013, 8, e59687. [Google Scholar] [CrossRef] [PubMed]
- Ernsting, G.; Isaaks, A. Ectotherms, temperature, and trade-offs: Size and number of eggs in a carabid beetle. Am. Nat. 2000, 155, 804–813. [Google Scholar] [PubMed]
- Helden, A.J.; Leather, S.R. Biodiversity on urban roundabouts-Hemiptera, management and the species-area relationship. Basic Appl. Ecol. 2004, 5, 367–377. [Google Scholar] [CrossRef]
- Williams, N.S.G.; Schwartz, M.W.; Vesk, P.A.; Mccarthy, M.A.; Hahs, A.K.; Clemants, S.E.; Corlett, R.T.; Richard, P.; Norton, B.A.; Thompson, K.; et al. A conceptual framework for predicting the effects of urban environments on floras. J. Ecol. 2008, 97. [Google Scholar] [CrossRef]
- Roy, H.E.; Brown, P.M.J.; Adriaens, T.; Berkvens, N.; Borges, I.; Clusella-Trullas, S.; Comont, R.F.; De Clercq, P.; Eschen, R.; Estoup, A.; et al. The harlequin ladybird, Harmonia axyridis: Global perspectives on invasion history and ecology. Biol. Invasions 2016, 18, 997–1044. [Google Scholar] [CrossRef]
- Camacho-Cervantes, M.; Ortega-Iturriaga, A.; Del-Val, E. From effective biocontrol agent to successful invader: The harlequin ladybird (Harmonia axyridis) as an example of good ideas that could go wrong. PeerJ 2017, 5, e3296. [Google Scholar] [CrossRef] [PubMed]
- Ingebritsen, S.E.; Jones, D.R. Santa Clara Valley, California; U.S. Geological Survey: Menlo Park, CA, USA, 1999; Volume 1182, pp. 15–22.
- UCCE. Master Gardener Program Home, Garden, Turf, and Landscape Pests. Available online: http://ipm.ucanr.edu/ (accessed on 1 March 2018).
- Cincotta, R.P.; Wisnewski, J.; Engelman, R. Human population in the biodiversity hotspots. Nature 2000, 404, 25–27. [Google Scholar] [CrossRef] [PubMed]
- Ives, C.D.; Lentini, P.E.; Threlfall, C.G.; Ikin, K.; Shanahan, D.F.; Garrard, G.E.; Bekessy, S.A.; Fuller, R.A.; Mumaw, L.; Rayner, L.; et al. Cities are hotspots for threatened species. Glob. Ecol. Biogeogr. 2016, 25, 117–126. [Google Scholar] [CrossRef]
- Schwartz, M.W.; Jurjavcic, N.L.; O’Brien, J.M. Conservation’s disenfranchised urban poor. Bioscience 2002, 52, 601–606. [Google Scholar] [CrossRef]
- Martin, N.A. Dusky Lady Beetle—Nephus binaevatus. Available online: http://nzacfactsheets.landcareresearch.co.nz/Index.html (accessed on 1 March 2018).
- Cadotte, M.; Lovett-Doust, J. Ecological and taxonomic differences between rare and common plants of southwestern Ontario. Ecoscience 2002, 9, 397–406. [Google Scholar] [CrossRef]
- Deguines, N.; Julliard, R.; Flores, M.; Fontaine, C. Functional homogenization of flower visitor communities with urbanization. Ecol. Evol. 2016, 6, 1967–1976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cadotte, M.W.; Lovett-doust, J. Ecological patterns and biological invasions: Using regional species inventories in macroecology. Biol. Invasions 2006, 8, 809–821. [Google Scholar] [CrossRef]
- Ossola, A.; Nash, M.A.; Christie, F.J.; Hahs, A.K.; Livesley, S.J. Urban habitat complexity affects species richness but not environmental filtering of morphologically-diverse ants. PeerJ 2015, 3, e1356. [Google Scholar] [CrossRef] [PubMed]
- Finlayson, A.C.J.; Landry, K.M.; Alyokhin, A. V Abundance of native and non-native lady beetles (Coleoptera: Coccinellidae) in different habitats in Maine. Ann. Entomol. Soc. Am. 2008, 101, 1078–1087. [Google Scholar] [CrossRef]
- Roy, H.E.; Brown, P.M.J.; Comont, R.F.; Poland, R.L.; Sloggett, J.J. Ladybirds; Pelagic Publishing Ltd.: Exeter, UK, 2013; Volume 10, ISBN 190780739X. [Google Scholar]
- Hagen, K.S. Biology and ecology of predaceous coccinellidae. Annu. Rev. Entomol. 1962, 7, 289–326. [Google Scholar] [CrossRef]
- Rees, M.; Roe, J.H.; Georges, A. Life in the suburbs: Behavior and survival of a freshwater turtle in response to drought and urbanization. Biol. Conserv. 2009, 142, 3172–3181. [Google Scholar] [CrossRef]
- Parris, K.M.; Hazell, D.L. Biotic effects of climate change in urban environments: The case of the grey-headed flying-fox (Pteropus poliocephalus) in Melbourne, Australia. Biol. Conserv. 2005, 124, 267–276. [Google Scholar] [CrossRef]
- US Climate Data California Climate Data. Available online: http://www.usclimatedata.com/climate.php (accessed on 1 November 2017).
- Taylor, P.D.; Fahrig, L.; Henein, K.; Merriam, G. Connectivity is a vital element of landscape structure. Oikos 1993, 68, 571–573. [Google Scholar] [CrossRef]
- Rudd, H.; Vala, J.; Schaefer, V. Importance of backyard habitat in a comprehensive biodiversity conservation strategy: A connectivity analysis of urban green spaces. Restor. Ecol. 2002, 10, 368–375. [Google Scholar] [CrossRef]
- Levy, J.M.; Connor, E.F. Are gardens effective in butterfly conservation? A case study with the pipevine swallowtail, Battus philenor. J. Insect Conserv. 2004, 8, 323–330. [Google Scholar] [CrossRef]
- Burks, J.M.; Philpott, S.M. Local and landscape drivers of parasitoid abundance, richness, and composition in urban gardens. Environ. Entomol. 2017, 46, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Plascencia, M.; Philpott, S.M. Floral abundance, richness, and spatial distribution drive urban garden bee communities. Bull. Entomol. Res. 2017, 107, 658–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Study regions in Michigan (a) and in California (b) where community gardens (black circles) were sampled. Increasing percent impervious surface (NLCD 2011) shown with increasing shaded color.
Figure 1.
Study regions in Michigan (a) and in California (b) where community gardens (black circles) were sampled. Increasing percent impervious surface (NLCD 2011) shown with increasing shaded color.

Figure 2.
Effect of impervious surface on abundance, richness and diversity of ladybird beetles. Regressions of abundance, richness and diversity (Shannon Index) of Michigan (black lines and points) and California ladybird beetles (red lines and points) as a function of % impervious surface at 500 m for MI and 100 m for CA. All species combined (a–c, top row), species that are shared by both CA and MI (d–f, middle row) and species unique to each region (g–i, bottom row). * indicate significant regressions (p < 0.05). In (b), Poisson regressions for MI and CA are partially significant (p < 0.10).
Figure 2.
Effect of impervious surface on abundance, richness and diversity of ladybird beetles. Regressions of abundance, richness and diversity (Shannon Index) of Michigan (black lines and points) and California ladybird beetles (red lines and points) as a function of % impervious surface at 500 m for MI and 100 m for CA. All species combined (a–c, top row), species that are shared by both CA and MI (d–f, middle row) and species unique to each region (g–i, bottom row). * indicate significant regressions (p < 0.05). In (b), Poisson regressions for MI and CA are partially significant (p < 0.10).

Figure 3.
Effect of urbanization rate on abundance, richness and diversity of ladybird beetles. Regressions of abundance, richness and diversity (Shannon Index) of Michigan (black lines and points) and California ladybird beetles (red lines and points) as a function of urbanization rate at 500 m. All species combined (a–c, top row), species that are shared by both California and Michigan (d–f, middle row) and species unique to each region (g–i, bottom row). * indicate significant regressions (p < 0.05).
Figure 3.
Effect of urbanization rate on abundance, richness and diversity of ladybird beetles. Regressions of abundance, richness and diversity (Shannon Index) of Michigan (black lines and points) and California ladybird beetles (red lines and points) as a function of urbanization rate at 500 m. All species combined (a–c, top row), species that are shared by both California and Michigan (d–f, middle row) and species unique to each region (g–i, bottom row). * indicate significant regressions (p < 0.05).


{kind=link}
{kind=link}
{kind=link}
Table 1.
Ladybird beetle species sampled in California and in Michigan. We present: the respective region the species was found in, their feeding habits, the ecological role that they play in agroecosystems, their nativity in their respective region [39,48,49], and their current geographic distribution in the USA [39,40,41]. (CA = California; MI = Michigan; NA = North America).
Table 1.
Ladybird beetle species sampled in California and in Michigan. We present: the respective region the species was found in, their feeding habits, the ecological role that they play in agroecosystems, their nativity in their respective region [39,48,49], and their current geographic distribution in the USA [39,40,41]. (CA = California; MI = Michigan; NA = North America).
| Species | Region Observed | Feeds on | Ecological Function in Agroecosystems | Origin | Distribution in US |
|---|---|---|---|---|---|
| Adalia bipunctata | CA | aphids and mites | predator/pest control | native | West coast, Northeast, few Midwest records (historically most of US and Canada) |
| Coccinella californica | CA | mostly aphids | predator/pest control | native | West coast CA |
| Cycloneda polita | CA | mostly aphids | predator/pest control | native | West coast US and British Columbia |
| Hippodamia convergens | CA | mostly aphids | predator/pest control | native | Throughout US and western Canada |
| Hyperaspis quadrioculata | CA | aphids and scale insects | predator/pest control | native | Central to south CA |
| Nephus binaevatus | CA | aphids and scale insects | predator/pest control | non-native | Central to south CA |
| Psyllobora vigintimaculata | CA | fungus | fungus and mildew control | native | Throughout US and Canada |
| Scymnus cervicalis | CA | mites and scale insects | predator/pest and mite control | native | East US to south CA |
| Scymnus coniferarum | CA | mites and scale insects | predator/pest and mite control | native | CA and scattered west NA records |
| Scymnus marginicollis | CA | mites and scale insects | predator/pest and mite control | native | CA to British Columbia; scattered NA records |
| Scymnus nebulosus | CA | mites and scale insects | predator/pest and mite control | native | South CA to Canada |
| Stethorus punctum | CA | mites and scale insects | predator/pest and mite control | native | West coast US; Northeast, west to north Great Plains |
| Coleomegilla maculata | MI | mostly aphids | predator/pest control | native | East NA to southwest US |
| Cryptolaemus montrouzieri | MI | mites and scale insects | predator/pest and mite control | non-native | Throughout US |
| Hippodamia variegata | MI | mostly aphids | predator/pest control | native | Northeastern to middle US and Canada |
| Propylea quatuordecimpunctata | MI | mostly aphids | predator/pest control | non-native | Throughout NA (native to the Palaearctic) |
| Coccinella septempunctata | MI, CA | mostly aphids | predator/pest control | non-native | Throughout NA (native to the Old World) |
| Cycloneda sanguinea | MI, CA | mostly aphids | predator/pest control | native | West to south CA; NC and FL |
| Harmonia axyridis | MI, CA | mostly aphids | predator/pest control | non-native | Throughout US and southern Canada, except northern Rockies |
| Olla v-nigrum | MI, CA | mostly aphids | predator/pest control | native | Throughout US, except ME and Pacific Northwest |
Table 2.
Results of regressions predicting ladybird beetle abundance (AB), richness (RI) and Shannon’s Diversity Index (SH) as a function of percent impervious surface (IS) and urbanization rate (UR). Scale indicates the spatial scale in meters used to calculate predictor variables IS and UR. Coefficients and p values are derived from Wald Z tests, which assess goodness of fit of generalized linear models to data assuming Poisson error distributions (AB, RI) or Gaussian error distribution (SH).
Table 2.
Results of regressions predicting ladybird beetle abundance (AB), richness (RI) and Shannon’s Diversity Index (SH) as a function of percent impervious surface (IS) and urbanization rate (UR). Scale indicates the spatial scale in meters used to calculate predictor variables IS and UR. Coefficients and p values are derived from Wald Z tests, which assess goodness of fit of generalized linear models to data assuming Poisson error distributions (AB, RI) or Gaussian error distribution (SH).
| Dataset | Region | Scale | Predicted | Predictor | Coefficient | p-Value |
|---|---|---|---|---|---|---|
| All | MI | 500 | AB | IS | −0.015 | 0.01 |
| All | CA | 100 | AB | IS | 0.019 | <0.001 |
| All | MI | 500 | RI | IS | −0.018 | 0.08 |
| All | CA | 100 | RI | IS | 0.009 | 0.06 |
| All | MI | 500 | SH | IS | −0.013 | 0.02 |
| All | CA | 100 | SH | IS | 0.006 | 0.05 |
| Shared | MI | 500 | AB | IS | −0.001 | 0.92 |
| Shared | CA | 100 | AB | IS | 0.031 | <0.001 |
| Shared | MI | 500 | RI | IS | −0.016 | 0.29 |
| Shared | CA | 100 | RI | IS | 0.021 | 0.05 |
| Shared | MI | 500 | SH | IS | −0.003 | 0.54 |
| Shared | CA | 100 | SH | IS | 0.007 | 0.09 |
| Unique | MI | 500 | AB | IS | −0.023 | 0.004 |
| Unique | CA | 100 | AB | IS | 0.018 | <0.001 |
| Unique | MI | 500 | RI | IS | −0.020 | 0.16 |
| Unique | CA | 100 | RI | IS | 0.004 | 0.46 |
| Unique | MI | 500 | SH | IS | −0.005 | 0.44 |
| Unique | CA | 100 | SH | IS | 0.003 | 0.76 |
| All | MI | 500 | AB | UR | −3.524 | 0.15 |
| All | CA | 500 | AB | UR | 2.231 | <0.001 |
| All | MI | 500 | RI | UR | 0.932 | 0.77 |
| All | CA | 500 | RI | UR | 2.292 | 0.11 |
| All | MI | 500 | SH | UR | 1.372 | 0.55 |
| All | CA | 500 | SH | UR | 2.331 | 0.04 |
| Shared | MI | 500 | AB | UR | 3.710 | 0.22 |
| Shared | CA | 500 | AB | UR | 1.925 | 0.29 |
| Shared | MI | 500 | RI | UR | 4.665 | 0.23 |
| Shared | CA | 500 | RI | UR | 2.024 | 0.50 |
| Shared | MI | 500 | SH | UR | 2.698 | 0.09 |
| Shared | CA | 500 | SH | UR | 3.110 | 0.02 |
| Unique | MI | 500 | AB | UR | −10.88 | 0.01 |
| Unique | CA | 500 | AB | UR | 3.376 | <0.001 |
| Unique | MI | 500 | RI | UR | −4.020 | 0.47 |
| Unique | CA | 500 | RI | UR | 1.705 | 0.33 |
| Unique | MI | 500 | SH | UR | −1.506 | 0.48 |
| Unique | CA | 500 | SH | UR | 1.473 | 0.24 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Egerer, M.; Li, K.; Ong, T.W.Y. Context Matters: Contrasting Ladybird Beetle Responses to Urban Environments across Two US Regions. Sustainability 2018, 10, 1829. https://doi.org/10.3390/su10061829
AMA Style
Egerer M, Li K, Ong TWY. Context Matters: Contrasting Ladybird Beetle Responses to Urban Environments across Two US Regions. Sustainability. 2018; 10(6):1829. https://doi.org/10.3390/su10061829
Chicago/Turabian StyleEgerer, Monika, Kevin Li, and Theresa Wei Ying Ong. 2018. "Context Matters: Contrasting Ladybird Beetle Responses to Urban Environments across Two US Regions" Sustainability 10, no. 6: 1829. https://doi.org/10.3390/su10061829
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.