Abstract
When working with rare plant species, applying morphometric techniques is one of the main ways to obtain a representative data set on plant individuals and the state of particular populations growing in different ecological conditions and experiencing different degrees of anthropogenic load. Zygophyllum pinnatum, an understudied species growing on the territory of the Republic of Bashkortostan, the Orenburg region, and the Republic of Kazakhstan, is referred as a rare species. The purpose of the work is to study morphometric parameters and reveal the vitality structure of the Z. pinnatum coenopopulations in two regions of the Russian Federation (the Republic of Bashkortostan and Orenburg region) and in the north-west of the Republic of Kazakhstan (the Aktobe Region). Applying standard morphometric techniques, we analyzed the state of 16 coenopopulations of the species. Judging by most morphometric parameters, the leadership belongs to the individuals from the “Troitsk Cretaceous Mountains” coenopopulation located in the Sol-Iletsky District of Orenburg oblast, where, apparently, the most optimal plant conditions are formed (the absence of human impact and low projective cover of the grass stand to have a favorable effect). The minimum values for most parameters are recorded in the coenopopulations located on the territory of the Republic of Kazakhstan, resulting from arid growing conditions. The variability of most characters is within the normal range of the species response (Cv—5.0–44.6%). The discriminant function analysis revealed the morphostructural similarity of individuals from most coenopopulations. Regarding vitality, eight coenopopulations are prosperous, and another eight are depressed. The condition of the species coenopopulations is stable; however, the species is not provided with proper protection measures. Therefore, further monitoring of its habitats and improvement of environmental measures are necessary.
1. Introduction
Mountain territories are special ecosystems characterized by high biological diversity and a concentration of rare and endemic plant species [1]. Rare montane plants are most susceptible to the threat of loss of floristic diversity, as small range sizes, low population numbers, and narrow ecological niches make them particularly sensitive to changes in environmental factors and anthropogenic stress [2,3,4,5,6]. Therefore, rare mountain plants are important objects of study and protection at the regional and global levels [1,7,8].
The range boundaries of many Asian plant species, which are often represented by isolated and fragmented localities, run through the territory of the Southern Urals. In these disjunctive fragments of the ranges, populations of rare species are in borderline, often stressful, conditions and have peculiar internal organization, structure, morphology, and other biological features. Any human impacts (from environmental pollution to grazing, recreation, and deforestation) have an extremely negative effect on such borderline populations, as they lead to a further reduction of the ranges, up to their complete disappearance. Therefore, identification research and the study of rare species biological diversity in borderline conditions are urgent and top priority environmental tasks. Knowing the species biology and the structure of coenopopulations (CP), it is possible to predict the course of their development and their response to adverse environmental influences. In this regard, an urgent task is to conduct the habitat inventory, assess the state of coenopopulations in specific habitats, and study the biology of rare species, which makes it possible to understand the reasons for their rarity [9,10].
One of the main ways to obtain a representative data set on plant individuals and the state of specific populations growing in different parts of the ecocoenotic range and experiencing various degrees of anthropogenic load is through the use of morphometric techniques [11]. Relying on them, one can obtain valuable information about the state of rare plant species and assess the bioecological characteristics of individuals and populations in a comparative aspect on an objective basis [12,13,14,15,16,17,18,19].
The object of the study is Zygophyllum pinnatum Cham., a representative of the Zygophyllaceae family. The Latin name of the species is given according to the international nomenclature of the World Checklist of Vascular Plants [20]. This is an understudied Eastern European, Western, and Central Asian mountain-steppe species. Its range covers the Trans-Volga region, the south of Western Siberia, Central Asia (the Aral-Caspian region, the Balkhash region, the Karakum Desert, the Tian Shan, and the Pamir-Alay), and the northeast of Iran. It is a xerophyte growing on gypsum outcrops and loamy slopes, less often on alkali soils [21,22,23]. It is on the Red Book lists of the Republic of Bashkortostan [24], Orenburg [25], and Chelyabinsk [26] regions, as well as other regions of Russia. The Southern Urals are the northmost range border of the species; therefore, the studies on the biology of this rare plant are topical in comparison with the coenopopulations from the main species range. The purpose of this study is to identify the features of the morphological variability of Z. pinnatum in the CP of two regions of the Russian Federation (the Republic of Bashkortostan and Orenburg region) and in the northwest of the Republic of Kazakhstan (the Aktobe region).
2. Materials and Methods
Zygophyllum pinnatum is a perennial dwarf subshrub, 10–20 cm tall, with numerous, partly upright stems forming a sod. The caudex is multicipital and ligneous. The leaves are paripinnate, petiolate, with 8–12 small oval leaflets. The flowers are found in leaf axils and 2–3 pcs. per pedicel, upright while flowering, drooping while bearing fruit. Petals are 1.5 times longer than the calyx, obovate, obtuse at the apex, and orange in color. The fruit is a large, round-oval capsule with wide membranous wings. It blooms in May–June, bears fruit in July–August, and reproduces with seeds [21].
The studies of the species populations were carried out in 2015–2019 on the territory of the Russian Federation in the Cis-Urals of the Republic of Bashkortostan (RB, 3 administrative districts), Orenburg region (OR, 5 districts); on the Sub-Ural plateau of the Republic of Kazakhstan (RK) in the Aktobe region (AR, 2 districts). Within the study areas from north to south, we laid a transect stretching for 570 km from the central Cis-Urals of the Republic of Bashkortostan (Karmaskalinsky District) to the chalk mountains of the Terektitau area, Oiyl District, the Aktobe region, and the Republic of Kazakhstan. A total of 16 CPs of the species were studied. The names of coenopopulations were given according to the nearest settlement or geographical object.
An individual in a middle-aged generative state was taken as a counting unit, as is customary in methodological works within the population-ontogenetic approach [10,27,28]. The study of morphometry in natural conditions was carried out according to the method designed by V.N. Golubev [29] and was based on 25 middle-aged generative individuals from each of all sixteen Z. pinnatum populations. A total of 400 individuals were used in the analysis. The entire perennial axis of the plant from the substrate surface was taken for a generative shoot. The measurements were taken in the phases of flowering and the beginning of fructification since the species flowering period is long and the plants feature both flowers and fruits at the same time. We considered the following parameters: number of generative shoots per 1 plant, pcs.–Ngs; generative shoot height, cm–h; shoot diameter, cm–d; number of leaves per one generative shoot, pcs.–Nlv; leaf length, cm–Llv; leaf width, cm–Slv; number of leaflets, pcs.–Nlf; leaflet length, cm–Llf; leaflet width, cm–Slf; shrub diameter, cm–db; number of fruits per one shoot, pcs.–Nfr; fruit length, cm–Lfr; fruit width, cm–Sfr.
The multivariate analysis was performed using the Statistica 6.0 program (StatSoft Inc., Round Rock, Texas, USA) [30,31]. The phenotypic distance (the Mahalanobis distance) was calculated within the linear discriminant analysis. In the cluster analysis, the Euclidean distance was used as a distance between the samples in the morphological features complex; a dendrogram was constructed using single-linkage clustering [32].
The technique for assessing the vitality composition was based on the differentiation of plants of a similar ontogenetic state into vitality classes. The factor analysis, which allowed identifying a determining set of characters, was preliminarily carried out. The vitality spectra were compiled, and the coenopopulation quality index and vitality types were determined [10].
The static analysis was carried out in MS Excel 2010 using the Statistica 6.0 statistical software package; the coefficient of variation and variability range were assessed [33].
3. Results
The localization of the studied CPs is presented in Figure 1, their brief descriptions are given in Table 1. Is found in various types of petrophytic and calciphytic steppes of the Festuco-Brometea Br.-Bl. et Tx. ex Soó 1947 class, up to calciphytic, suffruticulose communities within the desert communities of the Anabasietea cretaceae Golovanov in Golovanov et al., 2021 class. In the northern part of the gradient, the predominant vegetation type is the Agropyron pectinatum community of the order Helictotricho-Stipetalia Toman 1969, the Festuco-Brometea class. Such coenoses encompass stony steppes characterized by a high frequency of petrophytes: Allium rubens Schrad. ex Willd., Hedysarum grandiflorum Pall., Koeleria sclerophylla P.A. Smirn., Onosma simplicissima L., in combination with true steppe species: Agropyron pectinatum (M. Bieb.) P. Beauv., Artemisia marschalliana Spreng., Stipa capillata L., Stipa lessingiana Trin. and Rupr., etc.
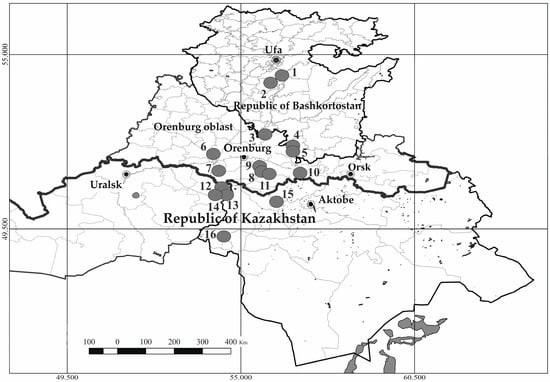
Figure 1.
Localities of the studied cenopopulations of Zygophyllum pinnatum Cham. (figures specify serial numbers of coenopopulations).

Table 1.
Characteristics of the investigated habitats of Zygophyllum pinnatum Cham.
In the south of RB and in OR, Z. pinnatum grows as part of the syntaxa within the classes Festuco-Brometea and Anabasietea cretaceae. Such associations as Athemido trotzkianae–Artemisietum salsoloidis Golovanov et al., 2021 (class Anabasietea cretaceae) refer to the most common types of communities, which are the communities of cretaceous massifs with exposure to the south and the west on rubbly, less erodible substrates with the presence of fine soil, widespread within the Sub-Ural Plateau [33]. Onosmato simplicissimae–Anthemietum trotzkianae Golovanov et al., 2021 (class Anabasietea cretaceae) are the communities of cretaceous outcrops of the Sub-Ural Plateau’s northern part and the Obshchy Syrt, transitional on a latitudinal gradient from the communities of the Anabasietea cretaceae class to petrophytic steppes of the Festuco-Brometea class, characteristic of the southern Cis-Urals [34]. Such community coenoses are characterized by the presence of species typical for the cretaceous outcrops of the region (Alyssum tortuosum Waldst. and Kit. ex Willd., Anthemis trotzkiana Claus, Atraphaxis decipiens Jaub. and Spach, Hedysarum tscherkassovae Knjasev, Matthiola fragrans Bunge, Psephellus marschallianus (Spreng.) K. Koch, etc.). Common petrophytes (Hedysarum grandiflorum Pall., H. razoumowianum Fisch. and Helm ex DC., Onosma simplicissima L., Scabiosa isetensis L., etc.) become more frequent in the associations at the northern boundary of the class communities.
Zygophyllum pinnatum is less common in the coenoses of the Elytrigion pruiniferae Korolyuk et al., 2022 alliance, comprising xeropetrophytic vegetation of the Southern Urals steppe zone [35]. These are such associations as: Elytrigietum pruiniferae Lebedeva in Korolyuk et al., 2022 (the central association of the alliance, encompassing xeropetrophytic coenoses of the hillocky and low-mountain massifs of OR and the south of RB) and Elytrigio pruiniferae–Stipetum sareptanae Golovanov in Korolyuk et al., 2022 (coenoses associated with hillocky massifs located to the south of the latitudinal segment of the Ural River, as well as the Southern Urals foothills). The above-mentioned communities are characterized by the presence of xerophytes: Allium tulipifolium Ledeb., Alyssum turkestanicum Regel & Schmalh., Ferula tatarica Fisch. ex Spreng, Linaria odora (M. Bieb.) Fisch., Poa bulbosa L., Pseudoroegneria geniculata (Trin.) Á. Löve, Thymus guberlinensis Iljin, Tulipa sylvestris L., etc.
In the Republic of Kazakhstan, the species was recorded only in the communities of the Anabasietea cretaceae class (association Athemido trotzkianae–Artemisietum salsoloidis).
Table 2 presents the results of studying the plant’s morphometric parameters in sixteen Z. pinnatum coenopopulations. The study revealed that the individuals from CP 13, which is located on the southern slope of the “Troitsk Cretaceous Mountains” natural monument in the Sol-Iletsky District of OR, surpass the individuals from other CPs in most indicators. Apparently, the most favorable conditions for plant growth are formed there (ecological, thermal, and humidity optimums). Additionally, the absence of human impact and low projective cover of the grass stand have a favorable effect. Also, CPs 2 and 3, located at gypsum outcrops in the northern part of the study area, feature high parameter values. The minimum values for most parameters are recorded for CPs 15 and 16, located on the territory of the RK, as well as for CP 14 (OR). These CPs occupy rubbly slopes of chalk hills with exposure to the south and west, featuring sparse desertificated plant communities. In this case, extreme arid growing conditions adversely affect the habitus of plants. The above-mentioned factors are associated with the largest number of generative shoots, which is also observed in CPs 2, 3, and 13, while the smallest number is observed in CPs 15 and 16. The generative shoot height parameter varies from 6.9 cm (CP 15) to 18.3 (CP 13). The largest number of leaves is recorded in CPs 2 and 13, the smallest in CPs 7, 16, and 14. The number of leaflets parameter varies from 6.8 in CP 13 to 11.7 in CP 5. According to such characters as the length and width of leaf and leaflet, no significant differences among coenopopulations were revealed; only in CPs 2 and 13, these characters are slightly higher, while in the southernmost CPs 14–16, they are lower. The maximum values for the number of fruits were recorded in CP 16, which is probably due to the high reproductive response in arid climate conditions. According to the coefficient of variation variability range, most characters have a normal degree of variability (Cv—5.0–44.6%). Wide and significant variation was revealed for the number of generative shoots (Cv—48.1–78.4%) and the number of fruits (Cv—55.2% and 66.0%). Increased character variability in plants in a CP is observed in cases of significant differentiation of individuals in terms of morphological structure. It is interpreted as a manifestation of phenotypic plasticity and the ability of a plant to adapt to environmental conditions.
Table 2.
Intrapopulation variability of morphometric parameters Zygophyllum pinnatum Cham.
To assess the intra- and interpopulation variability of Z. pinnatum under the influence of external factors, a discriminant function analysis was carried out, which showed that the Wilks λ values are very low (at p < 0.000), which indicates an overall high statistical significance of the obtained results. Turning to discriminant function analysis, one can not only assess the significance of interpopulation polymorphism and estimate the “distances” between populations but also designate those characters from among the considered that primarily determine interpopulation differences. The maximum contribution to the differentiation of groups is made by the diameter of the shrub (F = 27.93913), and the minimum contribution is made by the leaflet width (3.58747). The average distances of each individual from the center of the population to which it belongs are calculated. It was revealed that CP 5 has the maximum morphostructural diversity (30.49 ± 1.754), and CP 14 has the minimum (7.97 ± 0.710). Low phenotypic similarity was revealed between CP 13 and CPs 4, 5, 11, and 16 (80.559–87.533), and high phenotypic similarity was revealed between CPs 1–5, 1–6, 1–10, 4–5, 7–8, 7–15, and 12–15 (7.331–10.838).
The values of squared Mahalanobis distances between coenopopulations are calculated. The discriminant function analysis makes it possible to reveal both cases of almost complete phenotypic overlap among individuals from locally different populations as well as the differences among individuals in the structure of vegetative and generative organs. The maximum distance was revealed between CP 13 and CPs 4, 5, 11, 16 (80.559–87.533), and the smallest—between CP 1 and CPs 5, 6, 10, CPs 4–5, 7–8, 12–15 (7.331–10.838). The higher the phenotypic similarity of individuals between coenopopulations, the smaller the distance between them. It is possible that these CPs were once fragmented from a single population into isolated loci.
Figure 2 presents the visualization of the discriminant model, where the individuals of all 16 studied Z. pinnatum coenopopulations are displayed in the space of the first and second canonical roots. It can be seen that in most coenopopulations, plant individuals are homotypic in morphological structure, which is not entirely expected. The species grows in different thermal and humidity conditions, while individuals are phenotypically similar to each other. It is possible that the specific soil conditions in which the species grows (screes and outcrops of carbonate and gypsum rocks characterized by dry, mobile, and meager substrate) largely neutralize the influence of the climatic factor. High variations are observed only in the number of fruits and generative shoots. CP 12 and some individuals from CPs 3, 7, 8, 10, and 16 stay away from the general massif and occupy their own area; the overlap among them is insignificant. These individuals have minimum or maximum indicators in some morphometric parameters, which detach them from the CP to which they belong. In general, based on the discriminant function analysis, we established that the studied Z. pinnatum coenopopulations significantly differ from each other at a Wilks λ of 0.010 and p = 0.000.
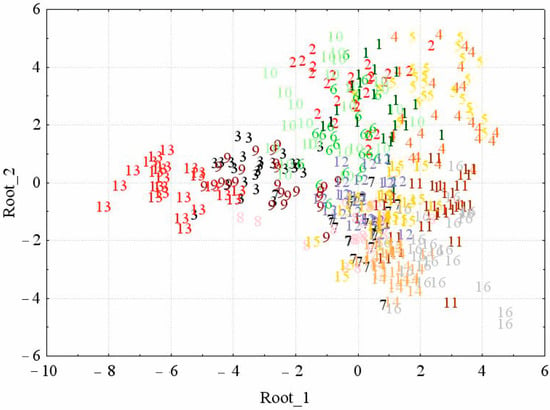
Figure 2.
The results of a discriminant analysis of coenopopulations of Zygophyllum pinnatum Cham. by the totality of morphometric characters in the space of the first and second canonical roots (1–16 population numbers).
In order to establish the way the character groups of the vegetative and generative spheres of Z. pinnatum plants correlate with each other, we carried out a canonical correlation analysis (Figure 3). The results of the analysis revealed that the canonical correlation coefficient R between vegetative and generative structures equals 0.723 and is 100% statistically significant (p = 0.000). It demonstrates that the development of generative organs largely depends on the development of vegetative structures, which is quite explicable since the maximum development of vegetative organs allows the generative sphere of plants to develop successfully. Strong positive correlations are observed between the leaf width and length, the leaflet width and length, the fruit width and length, as well as between the leaf length and the generative shoot height. An extensive number of strong positive correlations were found in CPs 2, 7, and 10. These coenopopulations are located in gypsified areas of petrophytic steppes, where the species are exposed to quite favorable growing conditions. Most CPs feature mean correlations between the characters. CPs 11 and 12 are characterized by very few correlations. The lowest correlation coefficients are typical for populations growing in dry cretaceous habitats in OR. With the deterioration of habitat conditions, the mean correlation degree may decrease, which may be indicative of an adaptive reaction of plants.
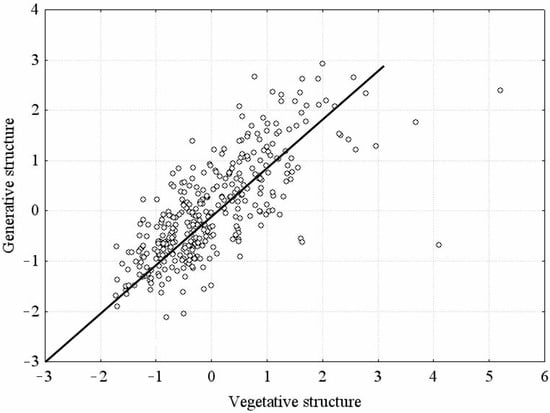
Figure 3.
Correlation of vegetative and generative structures in individuals of Zygophyllum pinnatum Cham.
The cluster analysis based on the mean values of the plants morphometric parameters resulted in the construction of a dendrogram reflecting the differences between the Z. pinnatum samples (Figure 4). When using the single linkage method, the studied populations, at a distance of 12.7, were divided into two clusters, one of which is clearly represented by CP 4, located on a slight slope with lime-sandstone outcrops exposed to a limited anthropogenic load. The second cluster, at a distance of 10.8, encompasses the rest of the populations, including a singled-out CP 12, which is characterized by the minimum morphometric values for a number of parameters. The last group, at a distance of 10.4, is split into two subclusters: the first one, at a distance of 10.1, comprises the southernmost CPs growing on the territory of the RK; the second one, at a distance of 9.8, includes CP 8. The subcluster, at a distance of 9.7, mainly encompasses thriving populations and is split, in turn, into 3 groups: the first one comprises CPs 3 and 7, which are characterized by the smallest fruits; CP 1 stands apart with the biggest fruits; and CPs 13 and 9, characterized by the tallest plants, coalesce in a separate subcluster at a distance of 5.8.
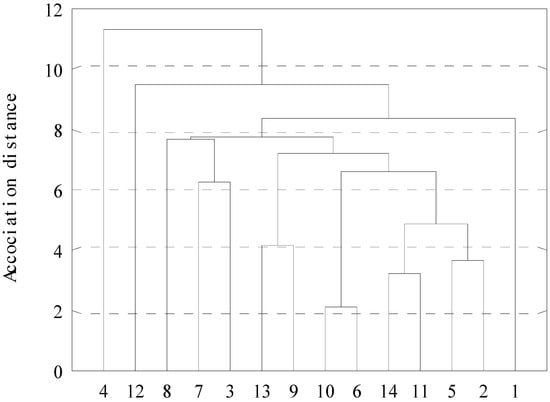
Figure 4.
Dendrogram of differences in samples of Zygophyllum pinnatum Cham. by average sample values of morphometric parameters of plants (1–16 coenopopulation numbers).
Table 3 presents the results of the Z. pinnatum individuals distribution by vitality classes in coenopopulations. Eight of the studied CPs are prosperous, and the proportion of individuals with high vitality (Q = 0.38–0.50) is seen as dominant. These CPs grow on high slopes (20–45°), predominantly on gypsum outcrops in the northern and central parts of the studied gradient. The remaining half of the CPs are characterized as depressed, with low vitality (Q = 0–0.28). They are confined mainly to the cretaceous substrate in the central and southern parts of the gradient. Six CPs lack specimens of the highest class completely; they are located mainly on cretaceous outcrops in the south of OR and on the territory of the RK.
Table 3.
Distribution of individuals in Zygophyllum pinnatum Cham. on the vitality classes.
4. Discussion
Thus, the conducted studies show that on the territory of the Republic of Bashkortostan, the Orenburg region, and the Republic of Kazakhstan, the rare Z. pinnatum species grows in various types of petrophytic and calciphytic steppes of the Festuco-Brometea class, up to calciphytic suffruticulose desertificated communities of the Anabasietea cretaceae class. The main syntaxon characteristic of the species in the northern part of the gradient is the Agropyron pectinatum community; the Athemido trotzkianae–Artemisietum salsoloidis and Onosmato simplicissimae–Anthemietum trotzkianae associations are to the south. The greatest diversity of phytocoenoses with Z. pinnatum participation is recorded in Orenburg oblast.
A certain confinement of Z. pinnatum to the substrate type is traced as well. Within the forest-steppe zone of the Bashkir Cis-Urals (in the northernmost part of its range), the species occurs exclusively on gypsum outcrops. To the south, it can be found both on gypsum outcrops and other rocks (lime-sandstones, chalks, etc.). Within the territory of the Republic of Kazakhstan, the species was most often recorded at the outcrops of chalk and marl.
The study of morphometric parameters revealed that the individuals from CP 13, which is located on the southern slope of the “Troitsk Cretaceous Mountains” natural monument in the Sol-Iletsky District of OR where, apparently, the most favorable growing conditions are formed, lead by most indicators. Also, according to the main indicators, the optimal conditions for Z. pinnatum growth develop closer to the northern border of the range, in more favorable, in terms of temperature and water regime, climatic conditions, where the species is found mainly on gypsum outcrops. Southward morphometric indicators decrease, and the state of populations worsens. The minimum values for most parameters were recorded in individuals from the coenopopulations located on the territory of the Republic of Kazakhstan, as well as in the “Troitsk Cretaceous Mountains 2” CP (OR), where insufficient moisture adversely affects the plant habitus. The variability of most characters lies within the normal range of the species response (Cv—5.0–44.6%). The variability of parameters is indicative of phenotypic differences in characters and may signify the high adaptability of individuals to different habitat conditions.
The vitality analysis revealed that eight coenopopulations are prosperous, while another eight are depressed. Populations with high vitality were found in conditions more favorable for moisture on gypsum outcrops in the northern and central parts of the studied gradient, while individuals with low vitality were found in arid conditions on cretaceous substrates in the central and southern parts of the gradient.
The analysis of Z. pinnatum phenotypic structure showed that the individuals in most coenopopulations are phenotypically similar, which is confirmed by a low variability of parameters. Higher variations are observed only in the number of fruits and generative shoots. Individuals with the highest morphostructural diversity are confined to lime-sandstones, such as in the “Verblyudka” CP (OR), with the minimum diversity—to cretaceous substrates, such as the “Troitsk Cretaceous Mountains 2” CP (OR). Low morphostructural diversity may indicate the fragmentation of isolated loci from a once-single population.
The most comfortable conditions for the growth of the species in the investigated territories are in the southern part of the Orenburg region, with an average annual rainfall of 300–350 mm and dark-chestnut soils. Also prospering are CPs located to the north, on the territory of the Republic of Bashkortostan (average annual rainfall is 400–450 mm), with a predominance of chernozem soils. To the south, in the Republic of Kazakhstan, in more arid conditions (average annual rainfall is 150–200 mm, soils are light chestnut), the vital indicators of the species are significantly reduced.
5. Conclusions
The analysis of morphometric parameters of plants and their variability and vitality in 16 cenopopulations of perennial dwarf subshrub Z. pinnatum, growing in petrophytic and calcifytic steppes of the Festuco–Brometea class and semi-shrub desert communities of the Anabasietea cretaceae class, revealed morphostructural diversity and vital condition of the species in different habitat conditions on the gradient: the Republic of Bashkortostan–Orenburg region–the Republic of Kazakhstan. Prosperous cenopopulations are located mainly in the northern and central parts, while depressive populations are located in the central and southern parts of the gradient.
More than half of the studied Z. pinnatum coenopopulations are not provided with proper protection measures. At the northernmost border of the range in the Republic of Bashkortostan, the species is protected only within one natural monument. Within Orenburg oblast, Z. pinnatum is protected much better. In the Republic of Kazakhstan, the studied coenopopulations are not protected. Despite the satisfactory state of the species coenopopulations, it is necessary to monitor its habitats further and improve environmental protection measures, which include the creation of new conservation areas, especially since other rare species, such as Hedysarum grandiflorum, Anthemis trotzkiana, Matthiola fragrans, Lepidium meyeri, etc., grow along with it. In particular, this applies to the coenopopulations at the northernmost border of the range, encountered on the territory of the Republic of Bashkortostan.
Author Contributions
Conceptualization, A.M., L.A., Y.G. and O.K.; Methodology, A.M. and Y.G.; Software, A.M. and Y.G.; Validation, A.M., L.A., Y.G. and O.K.; Formal Analysis, A.M., L.A. and Y.G.; Investigation, A.M., L.A., Y.G. and O.K.; Resources, A.M., L.A., Y.G. and O.K.; Data Curation, A.M. and L.A.; Writing—Original Draft Preparation, A.M. and L.A.; Writing—Review and Editing, A.M., L.A. and Y.G.; Visualization, A.M.; Supervision, L.A.; Project Administration, A.M.; Funding Acquisition, A.M., L.A., Y.G. and O.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
This study does not require ethical approval.
Informed Consent Statement
There were no human participants in this study.
Data Availability Statement
Data available on request due to restrictions eg privacy or ethical. The data presented in this study are available on request from the corresponding author. The data are not publicly available.
Acknowledgments
This work is carried out on the topic of the SUBGI UFRC RAS “Biodiversity of Natural Systems and Plant Resources of Russia: State Assessment and Monitoring of Dynamics, Problems of Conservation, Reproduction, Increase and Rational Use” within the framework of the State assignment for the year of 2022, UFRC RAS No. 075-03-2022-001 from 14 January 2022.
Conflicts of Interest
The authors declare no conflict of interest. The sponsors had no role in the design, execution, interpretation, or writing of the study.
References
- Gorchakovskiy, P.L.; Zolotareva, N.V. Phytodiversity of relict steppe enclaves in the Urals: Experience in comparative assessment. Russ. J. Ecol. 2006, 37, 378–386. [Google Scholar] [CrossRef]
- Dyke, F.V. Conservation Biology. Foundations, Concepts, Applications; Springer: Dordrecht, The Netherlands, 2008; 459p. [Google Scholar]
- Karimova, O.A.; Abramova, L.M.; Golovanov, Y.M. Analysis of the Current Status of Populations of Rare Plant Species of Nature Monument of Troicki Chalk Mountains (Orenburg Region). Arid. Ecosyst. 2017, 7, 41–48. [Google Scholar] [CrossRef]
- Mazangi, A.; Ejtehadi, H.; Mirshamsi, O.; Ghassemzadeh, F.; Hosseinianyousefkhani, S.S. Effects of climate change on the distribution of endemic Ferula xylorhachis Rech. f. (Apiaceae: Scandiceae) in Iran: Predictions from ecological niche models. Russ. J. Ecol. 2016, 47, 349–354. [Google Scholar] [CrossRef]
- Pauli, H.; Gottfried, M.; Dirnböck, T.; Dullinger, S.; Grabherr, G. Assessing the long-term dynamics of endemic plants at summit habitats. In Alpine Biodiversity in Europe; Ecological Studies (Analysis and Synthesis); Springer: Berlin/Heidelberg, Germany, 2003; Volume 167, pp. 195–207. [Google Scholar] [CrossRef]
- Pshegusov, R.H.; Chadaeva, V.A.; Tanija, I.V.; Abramova, L.M.; Mustafina, A.N. Life strategy and long-term climatogenic dynamics of the endemic Caucasian species Fritillaria latifolia Willd. Sci. Notes Kazan Univ. Ser. Nat. Sci. 2019, 161, 571–589. [Google Scholar] [CrossRef]
- Darbaeva, T.E. Partial Floras of the Cretaceous Uplands of Northwestern Kazakhstan; Western Kazakhstan University: Uralsk, Kazakhstan, 2006; 265p. [Google Scholar]
- Golovanov, Y.M.; Abramova, L.M. Chalky highlands in Orenburg oblast, a unique habitat for rare plant species and plant communities. Arid. Ecosyst. 2019, 9, 89–96. [Google Scholar] [CrossRef]
- Beissinger, S.R. Population viability analysis: Past, present, future. In Population Viability Analysis; University of Chicago: Chicago, IL, USA, 2002; pp. 5–15. [Google Scholar]
- Zlobin, Y.A.; Sklyar, V.G.; Klimenko, A.A. Populations of Rare Plant Species: Theoretical Bases and Methods of Study; University Book: Sumy, Ukraine, 2013; 439p. [Google Scholar]
- Laface, V.L.A.; Musarella, C.M.; Sorgonà, A.; Spampinato, G. Analysis of the Population Structure and Dynamic of Endemic Salvia ceratophylloides Ard. (Lamiaceae). Sustainability 2022, 14, 10295. [Google Scholar] [CrossRef]
- Brigham, C.А.; Schwartz, M.W. Population Viability in Plants: Conservation, Management and Modeling of Rare Plants; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2003; 366p. [Google Scholar]
- Callaway, R.M.; Pennings, S.C.; Richards, C.L. Phenotypic plasticity and interactions among plants. Ecology 2003, 84, 1115–1128. [Google Scholar] [CrossRef]
- Sultan, S.E. Phenotypic plasticity in plants: A case in ecological development. Evol. Dev. 2003, 5, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Klingenberg, C.P.; Duttke, S.; Whelan, S.; Kim, M. Developmental plasticity, morphological variation and evolvability: A multilevel analysis on morphometric integration in the shape of compound leaves. J. Evol. Biol. 2012, 25, 115–129. [Google Scholar] [CrossRef] [PubMed]
- Mustafina, A.N.; Abramova, L.M.; Shigapov, Z.K. Dictamnus Gymnostylis on the South Ural: Biology, Population Structure and Introduction; Gilem: Ufa, Russia, 2014; 184p. [Google Scholar]
- Karimova, O.A.; Mustafina, A.N.; Abramova, L.M. Modern state of natural populations of Medicago cancellata Bieb. rare species in the Bashkortostan Republic. Tomsk. State Univ. J. Biol. 2016, 3, 43–59. [Google Scholar] [CrossRef]
- Karimova, O.A.; Abramova, L.M.; Mustafina, A.N.; Golovanov, Y.M. State of coenopopulations of Anthemis trotzkiana (Asteraceae) in Orenburg Region. Bot. J. 2018, 103, 740–754. [Google Scholar] [CrossRef]
- Abramova, L.M.; Ilyina, V.N.; Karimova, O.A.; Mustafina, A.N. Features of the population organization of the rare species Cephalaria uralensis (Murr.) Schrad. ex Roem. et Schult (Dipsacaceae, Magnoliópsida) in the Trans-Volga and Cis-Urals regions. Povolzhskiy J. Ecol. 2018, 1, 3–15. [Google Scholar] [CrossRef]
- International Nomenclature World Checklist of Vascular Plants. Available online: https://powo.science.kew.org/ (accessed on 11 May 2023).
- Borisova, A.G. Rod. 840. Zygophyllum. In Flora SSSR; Academy of Sciences of USSR: Moscow/Leningrad, Russia, 1949; Volume 14, pp. 182–183. [Google Scholar]
- Ryabinina, Z.N.; Knyazev, M.S. Key of Vascular Plants of Orenburg Region; KMK Scientific Press Ltd.: Moscow, Russia, 2009; 758p. [Google Scholar]
- Kulikov, P.V. Key of Vascular Plants of Chelyabinsk Region; UrО RAN: Ekaterinburg, Russia, 2010; 968p. [Google Scholar]
- Red Book of the Republic of Bashkortostan. In Plants and Mushrooms; MediaPrint: Ufa, Russia, 2011; Volume 1, 384p.
- Red Book of the Orenburg Region: Rare and Endangered Species of Animals, Plants and Fungi; MIR: Voronezh, Russia, 2019; 488p.
- Lagunov, A.V. (Ed.) Red Book of Chelyabinsk Region: Animals, Plants, Mushrooms; Reart: Moscow, Russia, 2017; 504p. [Google Scholar]
- Smirnova, O.V.; Zaugolnova, L.B. (Eds.) Plant Coenopopulations (Basic Concepts and Structure); Nauka: Moscow, Russia, 1976; 217p. [Google Scholar]
- Sharma, S.K.; Pandit, M.K. Morphometric analysis and taxonomic study of Panax bipinnatifidus Seem. (Araliaceae) species complex from Sikkim Himalaya, India. Plant Syst. Evol. 2011, 297, 87–98. [Google Scholar] [CrossRef]
- Golubev, V.N. Basic of Biomorphology of Herbaceous Plants of Central Forest Steppe; Proceedings of the Central Black Earth Reserve named after Alekhin V.V.; Voronezh University: Voronezh, Russia, 1962; Volume 7, 602p. [Google Scholar]
- McLachlant, G.J. Discriminant Analysis and Statistical Pattern Recognition; John Wiley & Sons: Hoboken, NJ, USA, 2005; 544p. [Google Scholar]
- Khalafyan, А.А. STATISTICA 6. Statistical Data Analysis; Binom-Press: Moscow, Russia, 2008; 512p. [Google Scholar]
- Pesenko, Y.A. Principles and Methods of Quantitative Analysis in Faunal Studies; Nauka: Moscow, Russia, 1982; 287p. [Google Scholar]
- Zaytsev, G.N. Mathematics in Experimental Biology; Nauka: Moscow, Russia, 1990; 296p. [Google Scholar]
- Golovanov, Y.M.; Yamalov, S.M.; Lebedeva, M.V.; Korolyuk, A.Y.; Abramova, L.M.; Dulepova, N.A. Vegetation of Cretaceous outcrops of the Poduralsky plateau and adjacent territories. Veg. Russ. 2021, 40, 3–42. [Google Scholar] [CrossRef]
- Korolyuk, A.Y.; Yamalov, S.M.; Lebedeva, M.V.; Golovanov, Y.M.; Dulepova, N.A.; Zolotareva, N.V. Syntaxonomy of xeropetrophytic vegetation of the South Urals: Union Elytrigion pruiniferae all. nov. Veg. Russ. 2022, 43, 88–115. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).