
Neuroprotective Effect of Polyvalent Immunoglobulins on Mouse Models of Chemotherapy-Induced Peripheral Neuropathy

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. In Vivo Studies
2.2.1. Animals
2.2.2. Timeline of Treatments

2.2.3. Functional Assessments
von Frey Filament Test
Cold Plate Test
2.2.4. Quantification of Intra-Epidermal Nerve Fiber (IENF) and DRG Neuron Densities
2.2.5. Sciatic Nerve Ultrastructural Analysis
2.3. In Vitro Studies
2.3.1. Cell Lines
- Two diffuse large B-cell lymphoma cell lines, SU-DHL-4 (ATCC® CRL-2957™, ATCC, Manassas, VA, USA) and U-2932 (DSMZ® ACC633), sensitive to VCR (suspension cell culture).
- Two colorectal cancer cell lines, HT29 (ATCC® HTB-38™) and HCT116 (ATCC® CCL-247™), sensitive to OXP (adherent cell culture),
2.3.2. Treatments
2.3.3. Cell Viability Assay
2.4. Data Analysis
3. Results
3.1. Effect of Preventive hIg Treatment on the Onset of VCR-Induced Peripheral Neuropathy in Mice
3.1.1. Immunoglobulins and VCR Did Not Alter Weight Gain of Mice
3.1.2. hIg Alleviated Tactile Hypersensitivity Induced by VCR
3.1.3. hIg Alleviated Sensory Nerve Injuries Induced by VCR
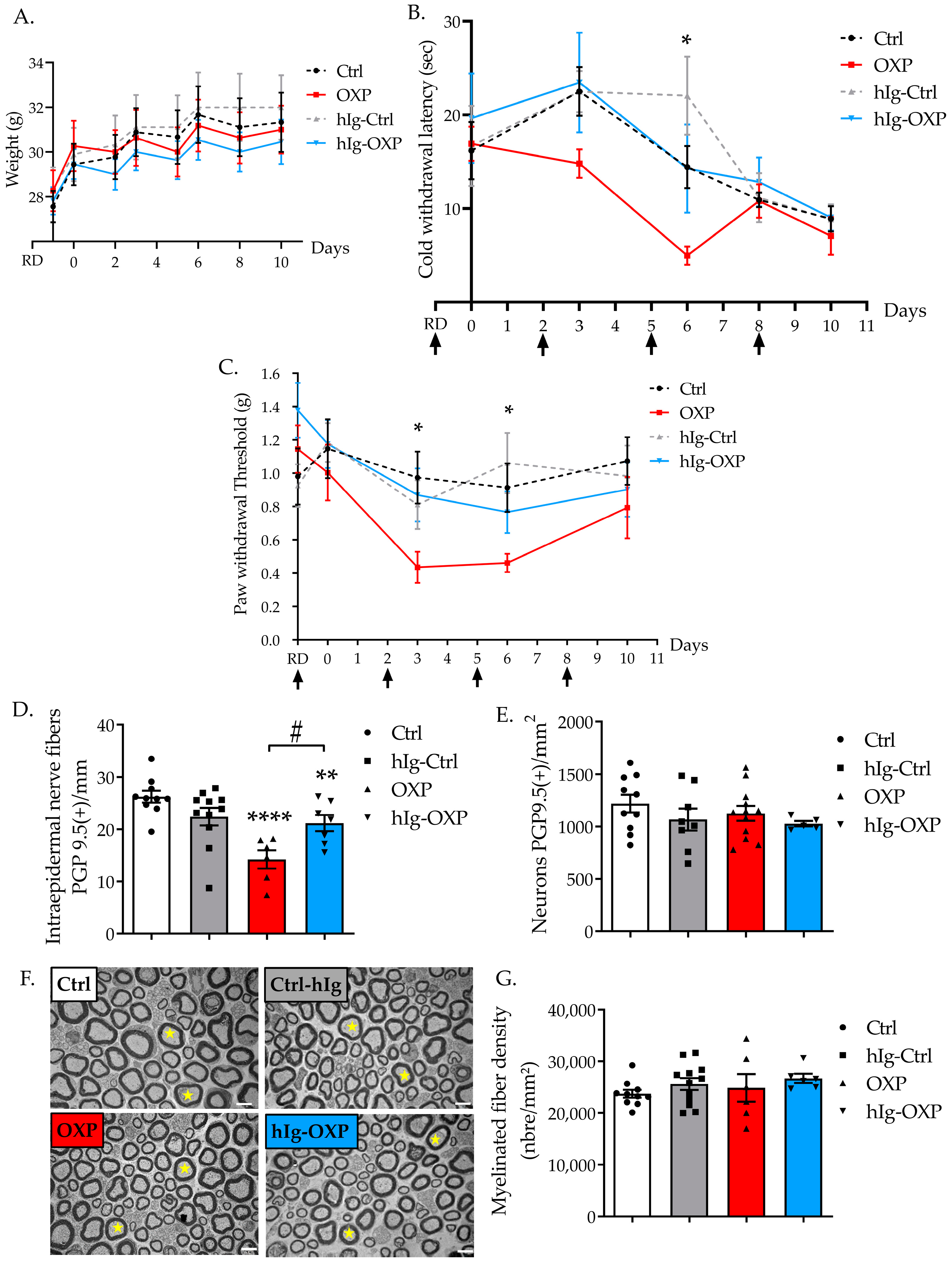
3.2. Effect of Preventive hIg Treatment on the Onset of OXP-Induced Peripheral Neuropathy in Mice
3.2.1. Immunoglobulins and OXP Did Not Alter Weight Gain of Mice
3.2.2. hIg Prevented Cold Hyperalgesia Induced by OXP
3.2.3. hIg Prevented Tactile Allodynia Induced by OXP
3.2.4. hIg Prevented Sensory Nerve Injury Induced by OXP
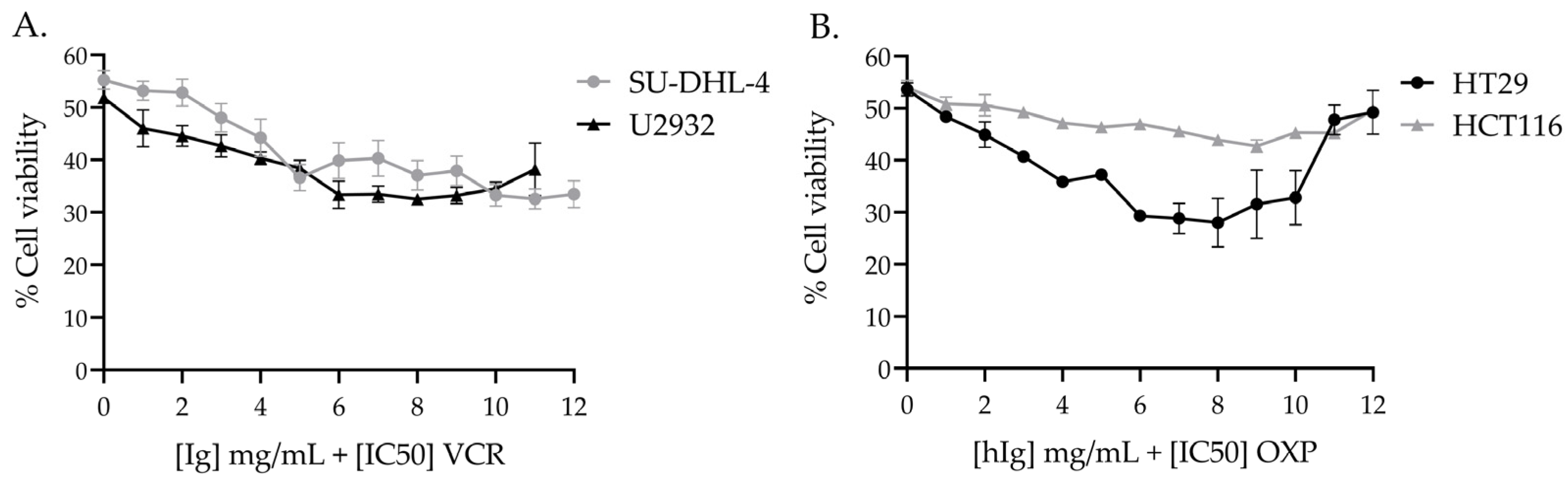
3.3. Effect of hIg on the Chemotherapy-Induced Cytotoxicity of Human Cancer Cell Lines
3.3.1. hIg Did Not Alter VCR-Induced Cytotoxicity in Human Lymphoma Cell Lines and Potentiated VCR Cytotoxicity
3.3.2. hIg Did Not Alter OXP-Induced Cytotoxicity in Colorectal Cancer Human Cell Lines
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seretny, M.; Currie, G.L.; Sena, E.S.; Ramnarine, S.; Grant, R.; MacLeod, M.R.; Colvin, L.A.; Fallon, M. Incidence, Prevalence, and Predictors of Chemotherapy-Induced Peripheral Neuropathy: A Systematic Review and Meta-Analysis. Pain 2014, 155, 2461–2470. [Google Scholar] [CrossRef]
- Jones, D.; Zhao, F.; Brell, J.; Lewis, M.A.; Loprinzi, C.L.; Weiss, M.; Fisch, M.J. Neuropathic Symptoms, Quality of Life, and Clinician Perception of Patient Care in Medical Oncology Outpatients with Colorectal, Breast, Lung, and Prostate Cancer. J. Cancer Surviv. Res. Pract. 2015, 9, 1–10. [Google Scholar] [CrossRef]
- Maletic, V.; Raison, C.L. Neurobiology of Depression, Fibromyalgia and Neuropathic Pain. Front. Biosci. Landmark 2009, 14, 5291–5338. [Google Scholar] [CrossRef]
- McWilliams, L.A.; Cox, B.J.; Enns, M.W. Mood and Anxiety Disorders Associated with Chronic Pain: An Examination in a Nationally Representative Sample. Pain 2003, 106, 127–133. [Google Scholar] [CrossRef]
- Lees, J.G.; Makker, P.G.S.; Tonkin, R.S.; Abdulla, M.; Park, S.B.; Goldstein, D.; Moalem-Taylor, G. Immune-Mediated Processes Implicated in Chemotherapy-Induced Peripheral Neuropathy. Eur. J. Cancer 2017, 73, 22–29. [Google Scholar] [CrossRef]
- Kiguchi, N.; Maeda, T.; Kobayashi, Y.; Kondo, T.; Ozaki, M.; Kishioka, S. The Critical Role of Invading Peripheral Macrophage-Derived Interleukin-6 in Vincristine-Induced Mechanical Allodynia in Mice. Eur. J. Pharmacol. 2008, 592, 87–92. [Google Scholar] [CrossRef]
- Liu, X.; Tonello, R.; Ling, Y.; Gao, Y.-J.; Berta, T. Paclitaxel-Activated Astrocytes Produce Mechanical Allodynia in Mice by Releasing Tumor Necrosis Factor-α and Stromal-Derived Cell Factor 1. J. Neuroinflamm. 2019, 16, 209. [Google Scholar] [CrossRef]
- Wahlman, C.; Doyle, T.M.; Little, J.W.; Luongo, L.; Janes, K.; Chen, Z.; Esposito, E.; Tosh, D.K.; Cuzzocrea, S.; Jacobson, K.A.; et al. Chemotherapy-Induced Pain Is Promoted by Enhanced Spinal Adenosine Kinase Levels through Astrocyte-Dependent Mechanisms. Pain 2018, 159, 1025–1034. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-S.; Li, Y.-Y.; Cui, W.; Li, L.-B.; Zhang, Z.-C.; Tian, B.-P.; Zhang, G.-S. Melatonin Attenuates Pain Hypersensitivity and Decreases Astrocyte-Mediated Spinal Neuroinflammation in a Rat Model of Oxaliplatin-Induced Pain. Inflammation 2017, 40, 2052–2061. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, Y.; de Carvalho-Barbosa, M.; Kavelaars, A.; Heijnen, C.J.; Albrecht, P.J.; Dougherty, P.M. Dorsal Root Ganglion Infiltration by Macrophages Contributes to Paclitaxel Chemotherapy-Induced Peripheral Neuropathy. J. Pain 2016, 17, 775–786. [Google Scholar] [CrossRef] [PubMed]
- Bethea, J.R.; Fischer, R. Role of Peripheral Immune Cells for Development and Recovery of Chronic Pain. Front. Immunol. 2021, 12, 641588. [Google Scholar] [CrossRef] [PubMed]
- Zis, P.; Liampas, A.; Pozotou, T.; Parperis, K.; Artemiadis, A.; Hadjigeorgiou, G. Immunoglobulin Use for the Management of Painful Peripheral Neuropathy: A Systematic Review and Meta-Analysis. Pain Ther. 2022, 11, 1219–1227. [Google Scholar] [CrossRef] [PubMed]
- Sapir, T.; Shoenfeld, Y. Facing the Enigma of Immunomodulatory Effects of Intravenous Immunoglobulin. Clin. Rev. Allergy Immunol. 2005, 29, 185–199. [Google Scholar] [CrossRef] [PubMed]
- Perez, E.E.; Orange, J.S.; Bonilla, F.; Chinen, J.; Chinn, I.K.; Dorsey, M.; El-Gamal, Y.; Harville, T.O.; Hossny, E.; Mazer, B.; et al. Update on the Use of Immunoglobulin in Human Disease: A Review of Evidence. J. Allergy Clin. Immunol. 2017, 139, S1–S46. [Google Scholar] [CrossRef] [PubMed]
- Meregalli, C.; Monza, L.; Chiorazzi, A.; Scali, C.; Guarnieri, C.; Fumagalli, G.; Alberti, P.; Pozzi, E.; Canta, A.; Ballarini, E.; et al. Human Intravenous Immunoglobulin Alleviates Neuropathic Symptoms in a Rat Model of Paclitaxel-Induced Peripheral Neurotoxicity. Int. J. Mol. Sci. 2021, 22, 1058. [Google Scholar] [CrossRef] [PubMed]
- Meregalli, C.; Marjanovic, I.; Scali, C.; Monza, L.; Spinoni, N.; Galliani, C.; Brivio, R.; Chiorazzi, A.; Ballarini, E.; Rodriguez-Menendez, V.; et al. High-Dose Intravenous Immunoglobulins Reduce Nerve Macrophage Infiltration and the Severity of Bortezomib-Induced Peripheral Neurotoxicity in Rats. J. Neuroinflamm. 2018, 15, 232. [Google Scholar] [CrossRef] [PubMed]
- Bessaguet, F.; Danigo, A.; Bouchenaki, H.; Duchesne, M.; Magy, L.; Richard, L.; Sturtz, F.; Desmoulière, A.; Demiot, C. Neuroprotective Effect of Angiotensin II Type 2 Receptor Stimulation in Vincristine-Induced Mechanical Allodynia. Pain 2018, 159, 2538–2546. [Google Scholar] [CrossRef]
- Bouchenaki, H.; Danigo, A.; Bernard, A.; Bessaguet, F.; Richard, L.; Sturtz, F.; Balayssac, D.; Magy, L.; Demiot, C. Ramipril Alleviates Oxaliplatin-Induced Acute Pain Syndrome in Mice. Front. Pharmacol. 2021, 12, 712442. [Google Scholar] [CrossRef]
- Chaplan, S.R.; Bach, F.W.; Pogrel, J.W.; Chung, J.M.; Yaksh, T.L. Quantitative Assessment of Tactile Allodynia in the Rat Paw. J. Neurosci. Methods 1994, 53, 55–63. [Google Scholar] [CrossRef]
- Allchorne, A.J.; Broom, D.C.; Woolf, C.J. Detection of Cold Pain, Cold Allodynia and Cold Hyperalgesia in Freely Behaving Rats. Mol. Pain 2005, 1, 1744–8069. [Google Scholar] [CrossRef]
- Lauria, G.; Cornblath, D.R.; Johansson, O.; McArthur, J.C.; Mellgren, S.I.; Nolano, M.; Rosenberg, N.; Sommer, C. EFNS Guidelines on the Use of Skin Biopsy in the Diagnosis of Peripheral Neuropathy. Eur. J. Neurol. Off. J. Eur. Fed. Neurol. Soc. 2005, 12, 747–758. [Google Scholar] [CrossRef] [PubMed]
- Pasetto, L.M.; D’Andrea, M.R.; Rossi, E.; Monfardini, S. Oxaliplatin-Related Neurotoxicity: How and Why? Crit. Rev. Oncol. Hematol. 2006, 59, 159–168. [Google Scholar] [CrossRef]
- Jordan, B.; Margulies, A.; Cardoso, F.; Cavaletti, G.; Haugnes, H.S.; Jahn, P.; Le Rhun, E.; Preusser, M.; Scotté, F.; Taphoorn, M.J.B.; et al. Systemic Anticancer Therapy-Induced Peripheral and Central Neurotoxicity: ESMO–EONS–EANO Clinical Practice Guidelines for Diagnosis, Prevention, Treatment and Follow-Up. Ann. Oncol. 2020, 31, 1306–1319. [Google Scholar] [CrossRef]
- Loprinzi, C.L.; Lacchetti, C.; Bleeker, J.; Cavaletti, G.; Chauhan, C.; Hertz, D.L.; Kelley, M.R.; Lavino, A.; Lustberg, M.B.; Paice, J.A.; et al. Prevention and Management of Chemotherapy-Induced Peripheral Neuropathy in Survivors of Adult Cancers: ASCO Guideline Update. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2020, 38, 3325–3348. [Google Scholar] [CrossRef]
- Burgess, J.; Ferdousi, M.; Gosal, D.; Boon, C.; Matsumoto, K.; Marshall, A.; Mak, T.; Marshall, A.; Frank, B.; Malik, R.A.; et al. Chemotherapy-Induced Peripheral Neuropathy: Epidemiology, Pathomechanisms and Treatment. Oncol. Ther. 2021, 9, 385–450. [Google Scholar] [CrossRef]
- Staff, N.P.; Cavaletti, G.; Islam, B.; Lustberg, M.; Psimaras, D.; Tamburin, S. Platinum-Induced Peripheral Neurotoxicity: From Pathogenesis to Treatment. J. Peripher. Nerv. Syst. JPNS 2019, 24, S26–S39. [Google Scholar] [CrossRef]
- Tanaka, J.; Kajii, M. Intravenous Immunoglobulin Suppresses Chemotherapy-Induced Peripheral Neurotoxicity Via Macrophage Modulation in Rats and Mice. Med. Res. Arch. 2022, 10, 1–10. [Google Scholar] [CrossRef]
- Fumagalli, G.; Monza, L.; Cavaletti, G.; Rigolio, R.; Meregalli, C. Neuroinflammatory Process Involved in Different Preclinical Models of Chemotherapy-Induced Peripheral Neuropathy. Front. Immunol. 2020, 11, 626687. [Google Scholar] [CrossRef]
- Meng, J.; Qiu, S.; Zhang, L.; You, M.; Xing, H.; Zhu, J. Berberine Alleviate Cisplatin-Induced Peripheral Neuropathy by Modulating Inflammation Signal via TRPV1. Front. Pharmacol. 2021, 12, 774795. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Zhang, Z.; Chen, Z.; Zhang, B.; Zhao, C.; Zhang, Y.; Zhao, C.; Deng, X.; Zhou, Y.; Wu, Y.; et al. Nonspecific Immunoglobulin G Is Effective in Preventing and Treating Cancer in Mice. Cancer Manag. Res. 2019, 11, 2073–2085. [Google Scholar] [CrossRef]
- Krause, I.; Shoenfeld, Y. Intravenous Immunoglobulin Treatment for Fibrosis, Atherosclerosis, and Malignant Conditions. In Adoptive Immunotherapy; Humana Press: Totowa, NJ, USA, 2004; Volume 109, pp. 403–408. ISBN 978-1-59259-862-5. [Google Scholar]
- Tzekova, N.; Heinen, A.; Bunk, S.; Hermann, C.; Hartung, H.-P.; Reipert, B.; Küry, P. Immunoglobulins Stimulate Cultured Schwann Cell Maturation and Promote Their Potential to Induce Axonal Outgrowth. J. Neuroinflamm. 2015, 12, 107. [Google Scholar] [CrossRef]
- Shang, Y.; Zhang, X.; Lu, L.; Jiang, K.; Krohn, M.; Matschos, S.; Mullins, C.S.; Vollmar, B.; Zechner, D.; Gong, P.; et al. Pharmaceutical Immunoglobulin G Impairs Anti-Carcinoma Activity of Oxaliplatin in Colon Cancer Cells. Br. J. Cancer 2021, 124, 1411–1420. [Google Scholar] [CrossRef]
- Hutmacher, D.W.; Loessner, D.; Rizzi, S.; Kaplan, D.L.; Mooney, D.J.; Clements, J.A. Can Tissue Engineering Concepts Advance Tumor Biology Research? Trends Biotechnol. 2010, 28, 125–133. [Google Scholar] [CrossRef]
- Starobova, H.; Monteleone, M.; Adolphe, C.; Batoon, L.; Sandrock, C.J.; Tay, B.; Deuis, J.R.; Smith, A.V.; Mueller, A.; Nadar, E.I.; et al. Vincristine-Induced Peripheral Neuropathy Is Driven by Canonical NLRP3 Activation and IL-1β Release. J. Exp. Med. 2021, 218, e20201452. [Google Scholar] [CrossRef]
- Brigo, F.; Balter, R.; Marradi, P.; Ferlisi, M.; Zaccaron, A.; Fiaschi, A.; Frasson, E.; Bertolasi, L. Vincristine-Related Neuropathy versus Acute Inflammatory Demyelinating Polyradiculoneuropathy in Children with Acute Lymphoblastic Leukemia. J. Child Neurol. 2012, 27, 867–874. [Google Scholar] [CrossRef]
- Young, G.; Angiolillo, A.L. Long-Term Treatment of Refractory Thrombocytopenia in a Patient with Wiskott-Aldrich Syndrome with Vincristine, Immunoglobulin, and Methylprednisolone. Am. J. Hematol. 1999, 62, 183–185. [Google Scholar] [CrossRef]
- Dumas, G.; Rousseau, B.; Rodrigues, M.J.; Tlemsani, C.; Goldwasser, F. Acquired Type II von Willebrand Syndrome in Locally Advanced Bladder Cancer Successfully Treated with Intravenous Immunoglobulin and Chemotherapy. Clin. Genitourin. Cancer 2016, 14, e95–e97. [Google Scholar] [CrossRef]
- Takano, H.; Tanaka, M.; Koike, R.; Nagai, H.; Arakawa, M.; Tsuji, S. Effect of Intravenous Immunoglobulin in Lambert–Eaton Myasthenic Syndrome with Small-cell Lung Cancer: Correlation with the Titer of Anti–Voltage-gateo Calcium Channel Antibody. Muscle Nerve 1994, 17, 1073–1075. [Google Scholar] [CrossRef]
- Phuphanich, S.; Brock, C. Neurologic Improvement after High-Dose Intravenous Immunoglobulin Therapy in Patients with Paraneoplastic Cerebellar Degeneration Associated with Anti-Purkinje Cell Antibody. J. Neurooncol. 2007, 81, 67–69. [Google Scholar] [CrossRef]
- Merimsky, O.; Meller, I.; Inbar, M.; Bar-Yehuda, S.; Shoenfeld, Y.; Fishman, P. A Possible Role for IVIg in the Treatment of Soft Tissue Sarcoma: A Clinical Case and an Experimental Model. Int. J. Oncol. 2002, 20, 839–843. [Google Scholar] [CrossRef]
- Schachter, J.; Katz, U.; Mahrer, A.; Barak, D.; David, L.Z.B.; Nusbacher, J.; Shoenfeld, Y. Efficacy and Safety of Intravenous Immunoglobulin in Patients with Metastatic Melanoma. Ann. N. Y. Acad. Sci. 2007, 1110, 305–314. [Google Scholar] [CrossRef]
- Yasuma, R.; Cicatiello, V.; Mizutani, T.; Tudisco, L.; Kim, Y.; Tarallo, V.; Bogdanovich, S.; Hirano, Y.; Kerur, N.; Li, S.; et al. Intravenous Immune Globulin Suppresses Angiogenesis in Mice and Humans. Signal Transduct. Target. Ther. 2016, 1, 15002. [Google Scholar] [CrossRef]
- Damianovich, M.; Solomon, A.S.; Blank, M.; Shoenfeld, Y. Attenuation of Colon Carcinoma Tumor Spread by Intravenous Immunoglobulin. Ann. N. Y. Acad. Sci. 2007, 1110, 567–577. [Google Scholar] [CrossRef]
- Prasad, N.K.; Papoff, G.; Zeuner, A.; Bonnin, E.; Kazatchkine, M.D.; Ruberti, G.; Kaveri, S.V. Therapeutic Preparations of Normal Polyspecific IgG (IVIg) Induce Apoptosis in Human Lymphocytes and Monocytes: A Novel Mechanism of Action of IVIg Involving the Fas Apoptotic Pathway. J. Immunol. 1998, 161, 3781–3790. [Google Scholar] [CrossRef]
- Lin, K.-L.; Chao, T.-C.; Chen, M.-H. Intravenous Immunoglobulin to Suppress Progression in a Patient with Advanced Breast Cancer. J. Breast Cancer 2022, 25, 253–258. [Google Scholar] [CrossRef]
- Rayburn, E. Anti-Inflammatory Agents for Cancer Therapy. Mol. Cell. Pharmacol. 2009, 1, 29–43. [Google Scholar] [CrossRef]
- Guo, Y.; Tian, X.; Wang, X.; Xiao, Z. Adverse Effects of Immunoglobulin Therapy. Front. Immunol. 2018, 9, 1299. [Google Scholar] [CrossRef]
- Zuercher, A.W.; Spirig, R.; Baz Morelli, A.; Käsermann, F. IVIG in Autoimmune Disease—Potential next Generation Biologics. Autoimmun. Rev. 2016, 15, 781–785. [Google Scholar] [CrossRef]
- Zuercher, A.W.; Spirig, R.; Baz Morelli, A.; Rowe, T.; Käsermann, F. Next-Generation Fc Receptor–Targeting Biologics for Autoimmune Diseases. Autoimmun. Rev. 2019, 18, 102366. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Anticancer Agent | Cell Line | Initial Seeding Density | IC50 Obtained |
|---|---|---|---|
| VCR | U2932 | 1 × 104 cells | 0.95 nM |
| SU-DHL-4 | 2 × 104 cells | 1.06 nM | |
| OXP | HT29 | 0.7 × 104 cells | 10.88 µM |
| HCT116 | 0.4 × 104 cells | 16.85 µM |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mroué, M.; Bessaguet, F.; Nizou, A.; Richard, L.; Sturtz, F.; Magy, L.; Bourthoumieu, S.; Danigo, A.; Demiot, C. Neuroprotective Effect of Polyvalent Immunoglobulins on Mouse Models of Chemotherapy-Induced Peripheral Neuropathy. Pharmaceutics 2024, 16, 139. https://doi.org/10.3390/pharmaceutics16010139
Mroué M, Bessaguet F, Nizou A, Richard L, Sturtz F, Magy L, Bourthoumieu S, Danigo A, Demiot C. Neuroprotective Effect of Polyvalent Immunoglobulins on Mouse Models of Chemotherapy-Induced Peripheral Neuropathy. Pharmaceutics. 2024; 16(1):139. https://doi.org/10.3390/pharmaceutics16010139
Chicago/Turabian StyleMroué, Mohamad, Flavien Bessaguet, Angélique Nizou, Laurence Richard, Franck Sturtz, Laurent Magy, Sylvie Bourthoumieu, Aurore Danigo, and Claire Demiot. 2024. "Neuroprotective Effect of Polyvalent Immunoglobulins on Mouse Models of Chemotherapy-Induced Peripheral Neuropathy" Pharmaceutics 16, no. 1: 139. https://doi.org/10.3390/pharmaceutics16010139