
Salmonella as a Promising Curative Tool against Cancer



1
Department of Veterinary Public Health, College of Veterinary Medicine, Jeonbuk National University, Iksan 54596, Korea
2
Department of Preventive Medicine, College of Veterinary Medicine, Chungnam National University, Daejeon 34134, Korea
*
Author to whom correspondence should be addressed.
Pharmaceutics 2022, 14(10), 2100; https://doi.org/10.3390/pharmaceutics14102100
Submission received: 3 September 2022
/
Revised: 28 September 2022
/
Accepted: 28 September 2022
/
Published: 1 October 2022
(This article belongs to the Special Issue Cancer Vaccine Technologies: Current Progress and Future Perspectives)
Abstract
:Bacteria-mediated cancer therapy has become a topic of interest under the broad umbrella of oncotherapy. Among many bacterial species, Salmonella remains at the forefront due to its ability to localize and proliferate inside tumor microenvironments and often suppress tumor growth. Salmonella Typhimurium is one of the most promising mediators, with engineering plasticity and cancer specificity. It can be used to deliver toxins that induce cell death in cancer cells specifically, and also as a cancer-specific instrument for immunotherapy by delivering tumor antigens and exposing the tumor environment to the host immune system. Salmonella can be used to deliver prodrug converting enzymes unambiguously against cancer. Though positive responses in Salmonella-mediated cancer treatments are still at a preliminary level, they have paved the way for developing combinatorial therapy with conventional chemotherapy, radiotherapy, and surgery, and can be used synergistically to combat multi-drug resistant and higher-stage cancers. With this background, Salmonella-mediated cancer therapy was approved for clinical trials by U.S. Food and Drug Administration, but the results were not satisfactory and more pre-clinical investigation is needed. This review summarizes the recent advancements in Salmonella-mediated oncotherapy in the fight against cancer. The present article emphasizes the demand for Salmonella mutants with high stringency toward cancer and with amenable elements of safety by virulence deletions.
1. Introduction
Cancer is a leading cause of death worldwide and a burgeoning health burden with a limited number of successful therapeutics. On average, 10 million people worldwide lose their lives annually due to various cancers [1]. Every cancer requires an accurate diagnosis and prompt treatment at the earliest possibility. Even though most conventional treatment strategies such as surgery, chemotherapy, and radiotherapy remain major life savers, they have serious limitations that can damage healthy tissues [2,3]. Surgical removal of cancers can be effective in certain types and developmental stages of cancers; however, cancer relapse and the possibility of further spread due to metastasis are some of the inherent weaknesses of this method [4]. On the contrary, radiotherapy and chemotherapy provide varying degrees of success and inflict unprecedented failures in cancer treatment, especially distant tumor recurrences and undesirable effects [5,6]. Hence, to fill the gap, novel treatment concepts and strategies are essential as an ideal treatment for cancers. Cancer tumors consist of hypoxic core regions and necrotic centers, which make most cancer treatments incompetent due to lack of oxygen and abnormal vasculature. Studies have demonstrated that such regions are the key features of tumors that lead to treatment failure [7,8,9]. In addition, due to the abnormal vascular architecture, it is a huge challenge to deliver therapeutic agents to the tumor region. Hence, it is evident that a single treatment strategy may not be effective against cancer malignancies, but holistic approaches might bring suboptimal outcomes.
In recent decades, bacteria-mediated cancer treatments (BMCT) have garnered attention as an alternative strategy to treat cancer tumors due to the intrinsic challenges of conventional cancer treatment strategies. Advancements in genetic engineering and recombinant DNA technology had paved the path for developing numerous bacterial strains as model systems to be used in cancer immunotherapy. William Coley’s controversial study, more than a century ago, revealed that some bacterial species may hold the key to creating targeted treatments for cancers that are challenging to cure. According to Coley, the complex cocktail he produced had the potential to shrink cancer tumors; however, the lack of progressive techniques and poor understanding of the mode of action made it difficult to reproduce consistent results. Revitalizing these early findings, scientists around the world have attempted to use novel bacterial species, such as Bifidobacterium, Clostridium, Salmonella, Streptococcus, and Listeria monocytogenes for tumor regression and have brought deep insight into their mode of action.
Accumulating evidence suggests that the cytotoxic effect of several bacteria can be hijacked and used against cancer cells. Some bacteria are naturally capable of homing to tumors, while other invasive species have been exploited to deliver heterologous genes intracellularly. The high rate of replication and invasive characteristics of some species, including Salmonella and Clostridium, are better vector candidates to express the target gene in tumor cells. Bacteria that have higher penetration and dispersion throughout tumors hold a greater ability to regress tumors [10]. Tumors have hypoxic regions where facultative anaerobes can survive and function to produce anti-tumor effects. Efficient delivery of the therapeutic gene to the target tissue or cell is the most significant hurdle for successful gene therapy. Thus, DNA is normally combined with a gene delivery vehicle in order to protect and mediate the effective tissue or cell entry of the payload gene of interest (GOI). The account for the payload GOI is handled by the bacterial invasive character that stands as a model in animal and human infection.
Among known bacterial species suitable for BMCT, Salmonella enterica serovar Typhimurium (ST) is one of the most versatile, as it can grow under both aerobic and anaerobic culture conditions. Therefore, it has no problem spreading systemically in animals under highly aerobic conditions, and eventually localizes in hypoxic tumor regions, which are their preferred sites of colonization. Salmonella has exhibited an immense capability to colonize hypoxic, necrotic, and metastatic tumors; thus, it can complement conventional treatment strategies [11]. The preferential accumulation ratio of ST in tumor regions is between 103 to 104 times more than in normal body tissues, resolving a huge challenge in the target specificity of cancer treatment [12]. Experimental data have confirmed that this localization occurs in a few days in most cases [13]. Thus, Salmonella can be used as a carrier to deliver therapeutics directly into the tumor regions, thereby protecting them from degradation and potential damage from the host immune system. Moreover, Salmonella can be easily tailored in a myriad of ways such as bacterial ghost systems, protein secretion systems, target-oriented and lysis systems, quorum sensing systems, etc. Thus, it has versatility for cancer treatment. Additionally, the simplicity of manufacturing it, its cost-effectiveness, and its rapid mass production place it as a new option for cancer therapy. In the present article, we elaborate on the mechanisms, strategies, and potential engineering methods of Salmonella as a key microbial agent suitable for the next generation of cancer therapy.
2. Bacterial Application for Cancer Therapy
Numerous studies have demonstrated the anti-tumor effects of several bacteria, either by directly killing or modulating immune components of the tumor microenvironment. The natural cytotoxic features of bacteria can result in substantial tumor regression. Therefore, many researchers have exploited non-pathogenic obligate anaerobes and facultative anaerobes which selectively infiltrate and replicate within solid tumors when administered systemically. The ability of bacteria to regress tumors came into the limelight in the early 1800s. BMCT started gaining momentum when Coley’s toxin, a mixture of killed Streptococcus pyogenes and Serratia marcescens developed by Dr. William B. Coley, achieved clinical responses for many malignant tumors [14]. Additionally, the anti-tumor characteristics of various bacteria have been documented, such as Salmonella [15], Escherichia coli, Vibrio cholerae and Listeria monocytogenes [16], Clostridium welchii [17], Clostridium tetani [18], Bifidobacterium infantis [19], Streptococcus pyogenes [20], and Proteus mirabilis [21]. Salmonella holds natural cytotoxicity that regresses tumors when injected in its native form [15]. Similarly, Clostridium, an obligate anaerobe, can regress tumors in mice [17,22,23].
The bacterial implication in cancer can function as a two-edged sword, since the association of certain pathogenic species has been linked to colon cancer. E. coli possesses a genomic island polyketide synthetase codes for the synthesis of colibactin that has been implicated in colorectal cancer [24]. In another scenario, Clostridium sps., especially Clostridium perfringens and Clostridium septicum, has been associated with colorectal cancer [25,26]. Salmonella typhi/paratyphi produces a potent carcinogen N-nitroso compound and has been documented for hepatobiliary carcinoma [27].
3. Attenuated Bacteria for Cancer Therapy
The unique tumor-homing characteristics of bacteria have been exploited for anti-tumor vaccine therapies, and such bacteria can be genetically attenuated to carry and deliver heterogeneous antigens which elicit host immunity. Generally, attenuation by modification on lipopolysaccharides, flagella, or other structural proteins is widely practiced. Moreover, with advancements in molecular biology, genetic modification for the delivery of different agents, such as tumor-associated antigens, immunostimulatory molecules, anti-tumor drugs, and nucleotides (DNA or RNA), are extensively accepted approaches [28]. A bacterial platform targeting cancer therapy is the choice of many researchers, as it possesses many benefits. Genetic manipulation, precise tuning, and limitless functional combinations of desired characteristics using bacteria have brought cancer treatment to a new horizon. Recently, several studies have focused on cancer treatment using the direct delivery of heterogeneous antigens or genes encoding anti-tumor molecules. Bacteria, including Salmonella species, have been significantly attenuated in their virulence in order to rule out a major threat of systemic infection or collateral damage [13,29].
Bacteria can specifically target tumors and actively penetrate tissue, which can induce cytotoxicity and destroy malignant cells [30]. Obligate anaerobes and facultative anaerobes can have an intrinsic tumor-targeting ability, because they can survive intra-tumor hypoxia. The hypoxic area in the tumor facilitates anaerobes and facultative anaerobes, as well as additional nutrients, including purines, and the immunosuppressive environment inhibits the clearance of Salmonella. Transforming growth factor-beta within a tumor inhibits the activation of neutrophils that prevent bacterial clearance from solid tumors [31]. Attenuated Salmonella strains which have been developed as gene delivery vectors carrying herpes simplex virus thymidine kinase (HSV TK) possess anti-tumor activity in mice and are capable of both selective amplification within tumors and expression of effector genes encoding therapeutic proteins [15].
Several strategies have been exploited for BMCT research, such as bacterial tumor-targeting properties, intra-tumoral penetration, bacterial cytotoxicity, expression of anti-cancer agents, host gene-triggering strategies, and immunotherapy. Although efforts are ongoing, the success of BMCT in animal models could not be replicated in humans using ST VNP20009. The attenuated strain with chromosomal deletion of the purI and msbB genes was able to target the tumor and inhibit tumor growth in mice [32], but failed to replicate anti-tumor effects in humans [33].
4. Why Is Salmonella the Best Option?
Salmonella-mediated cancer therapy (SMCT) is gaining popularity over other therapies because of several advantages. For instance, it has characteristic features that are peculiar to the species, such as self-targeting tumor localization and proliferation, and intrinsic anti-tumor nature. Compared with other bacterial species, it has several advantages, such as the relative ease of attenuation and gene manipulation, and it can grow in a hypoxic environment. It has a wide range of hosts, including humans and farm animals. In addition, it can be administered orally and stimulate local and systemic immune responses, highlighting its use as a model vector for cancer vaccine therapy.
4.1. High Tumor Colonization
The inherent facultative anaerobic and intracellular pathogenic characteristics of Salmonella facilitate strong preferential colonization in tumor tissue (Figure 1). Clairmont et al. have documented 1000-fold accumulation of the ST VNP20009 strain in tumors compared to the liver [32]. These attenuated strains were cleared rapidly from systemic circulation, liver, and spleen, whereas proliferation in tumor tissue continued for a longer time [32]. Thus, lower toxicity is observed in the host. Such selective tumor colonization and proliferation are owing to the hypoxic and vascularized environment of the tumor [34]. The mobility of Salmonella toward tumors and their accumulation is influenced by the tumor microenvironment, host reticuloendothelial system, and bacterial metabolism [35].
4.2. Non-Specific Tumor Target
Salmonella has been reported as having invasiveness toward a broad spectrum of murine tumor models. Several documents support its effectiveness in the treatment of melanoma, colon cancer, lung cancer, prostate cancer, cervical cancer, and metastatic T-cell lymphoma. The quiescent cells in the tumor cylindroid model secrete bacterial chemoattractant, and the presence of necrotic and quiescent cells attracts and enables ST to replicate in the central regions of the quiescent tumor cell mass [36]. Other mechanisms revealed that Salmonella uses aspartate receptors to initiate chemotaxis toward tumor cylindroids in vitro. It uses the serine receptor to initiate penetration and the ribose/galactose receptor to enhance affinity toward necrotic tissue [37].
4.3. Inherent Anti-Tumor Character
Salmonella is believed to have an intrinsic oncolytic activity that is mediated through the induction of tumor cell apoptosis. Bacterial invasion releases toxins, deprives the tumor microenvironment of nutrients, and promotes apoptosis [30]. Alternatively, the anti-angiogenic ability of Salmonella could delay tumor progression by inhibiting tumor angiogenesis [38,39]. Findings on the ability of Salmonella to downregulate the expression of hypoxia-inducible factor-1 alpha (HIF-1α) and vascular endothelial growth factor (VEGF) through the AKT/mTOR pathway support the anti-angiogenic ability of Salmonella [38]. Furthermore, bacteria can activate innate immune cells by cytokine stimulation and recruit and activate other immune cells at the tumor site, promoting anti-tumor immunity [40].
4.4. Engineering Plasticity
Salmonella is an excellent system for acquiring attenuation by targeting different approaches. It has flexibility in gene modification that helps to improve the safety of bacterial therapy through the deletion of major virulence factors or the generation of auxotrophic mutants that are incapable of replicating efficiently in an environment deficient in specific nutrients [41]. This increased specificity allows it to function as a “bacterial robot” to deliver anti-tumor therapeutic agents to the tumor site. It can deliver therapeutic payloads in the form of DNA, RNA, or protein. For this, the Salmonella delivery system needs to be transformed with extrinsic gene expression plasmids. The engineering plasticity of Salmonella enhances its efficacy in cancer therapy through improving safety, specificity, and delivery of anti-tumor therapeutic agents. It has been exploited for the delivery of cytotoxic agents such as cytolysin A, PE38, and diphtheria toxin, apoptosis-inducing proteins such as Fas ligand and TNF-related, apoptosis-inducing ligand (TRAIL), as well as apoptin and immunomodulatory cytokines, including IL-2, IL-12, IL-18, and IFN-γ [34,42,43,44,45,46]. Engineering plasticity enables the application of Salmonella in the expression of anti-cancer agents, “prodrugs,” or tumor-specific antigens at tumor sites. At present, the expression of oncogene-silencing RNA and a strategy of incorporating tumor-killing nanoparticles are being developed [30,47].
Recently, protein drug delivery to tumors was demonstrated by Raman et al. The group used intracellular delivery of Salmonella to express constitutive two-chain active caspase-3, an engineered form of caspase-3 that causes apoptotic cell death. They developed the strain by engineering genetic circuits to enable autonomous deposition of protein payloads directly into cancer cells that decreased tumor growth and reduced breast metastases [48]. Transforming growth factor alpha (TGFα) is a natural ligand for epidermal growth factor receptor (EGFR), a receptor highly expressed in tumor cells [49]. Researchers employed the ΔppGpp Salmonella mutant to deliver a recombinant drug TGFα-PE38, an immunotoxin comprising a modified Pseudomonas exotoxin A (PE38) conjugated with TGFα, and investigated the process through which it undermined the tumor by inducing the expression of pro-inflammatory cytokines from macrophages and neutrophils, such as IL-1β and TNFα [46,50]. The recombinant protein produced by bacteria effectively regressed the solid tumor growth and induced tumor cell apoptosis [46].
5. Salmonella enterica Serovar Typhimurium
ST, a gram-negative pathogenic bacteria [51], is one of the most extensively studied and promising bacterial mediators of cancer immunotherapy. ST is the choice of many researchers because of its ability to grow in both aerobic and anaerobic environments, resulting in its colonization in both non-hypoxic and hypoxic tumors [52]. It has high tumor specificity and deep tissue penetration potential. It possesses chemoreceptors, such as the aspartate receptor, that initiate chemotaxis toward viable tumor tissue, serine receptors to induce tissue penetration, and a ribose receptor to regulate migration toward necrotic tissue [37]. In addition, it has been engineered and designed in many studies that have explored cancer-targeting therapeutic agents (Table 1). ST has been engineered as a delivery vector for anti-tumor effects by expressing the pro-apoptotic Fas ligand in breast cancer and colon carcinomas in mice models [53]. TRAIL, a natural inducer of apoptosis and tumor cell death, was used in ST-based cancer therapy under the control of a prokaryotic radiation-inducible promoter, recA. This study has shown the inhibition of mammary tumor growth and substantially increased rates of survival [54].
Since the recognition that bacteria could be used for cancer therapy in the 19th century, Salmonella has been widely studied. It has been the best option for researchers as it covers many features, including high tumor-targeting capacity, tumor specificity, deep tissue penetration, and engineering plasticity. ST with auxotrophic mutations has demonstrated prominent anti-cancer potential [13,15,67,68]. Attenuated Salmonella with modification on lipopolysaccharide has been reported to have an anti-tumor effect [69]. ST with a modified lipid A (ST VNP20009) has achieved anti-tumor responses in dogs [70]. Lipid A-modified ST replicated more than 1000 times in tumors compared to normal tissue [68]. Auxotrophs for aromatic compounds, such as tryptophan [13] and purine, revealed Salmonella to be a good candidate for cancer therapeutics.
6. ST VNP20009 Strain
Several ST mutants have been evaluated for cancer therapy. The strains that are studied most frequently are ST VNP20009, A1-R, and other mutants. Among them, the ST VNP20009 strain was generated to obtain stable attenuated virulence by deletion of the purl gene for purine auxotrophy, and endotoxicity was reduced by the deletion of the msbB gene with a lipid-A modification [32]. ST VNP20009 renders selective accumulation at a ratio of >1000:1 over normal organs and specificity toward the tumor tissue, and is highly attenuated with a virulence reduction of about 10,000-fold [12]. This strain has been used in many studies and is the only strain to be evaluated in phase I clinical trials for metastatic melanoma or renal cell carcinoma in humans [33,71]. Mice were implanted with murine melanoma, and tumor growth was inhibited in the human tumor xenografts by 57–95% with a single dose of ST VNP20009 [72]. Aside from solid tumors, ST VNP20009 can induce apoptosis in multiple types of leukemia cells and prolong the survival of the MLL-AF9-induced acute myeloid leukemia-carrying mice [73]. Zheng et al. have demonstrated transgenic expressions of proteins by ST VNP20009 [74], and it has been used for the delivery of cycle-inhibiting factor genes for the treatment of colon cancer [75]. The lack of tumor regression in clinical trials of ST VNP20009 [33] compelled further modification, and the phoP/phoQ system was deleted in order to enhance tumor-targeting ability [76]. Complete genome sequencing of ST VNP2009 revealed nonsynonymous single nucleotide polymorphisms and purM deletion, demanding further study for effective human application [77].
A new method for tumor localization and improved safety has been demonstrated by coating VNP20009 with tumor cell-derived nanoshells [78]. The carcinoma cell-mimetic bacteria (CCMB) were released from carcinoma cells using UV irradiation, with an extra membrane from apoptotic bodies of invaded tumor cells as coating shells. The presence of a tumor cell-derived membrane in CCMB elicits low inflammation and enhances homologous-targeting tumor localization. This strategy exhibited tumor regression and metastasis inhibition and stands as a promising biotherapy for tumor treatment [78].
7. ST A1-R Strain
ST A1-R strain is a tumor-seeking leucine-arginine auxotrophic mutant developed by nitrosoguanidine mutagenesis that grows in viable as well as necrotic regions of tumors, but is restricted to normal tissue. It has been characterized to treat various tumor models in mice such as prostate, breast, pancreatic, and ovarian cancer, as well as sarcoma and glioma [79]. Mice implanted with metastatic PC-3 human prostate tumors were cured by nearly 70% when injected weekly with ST A1-R [80]. Likewise, mouse models with disseminated and metastatic human ovarian cancer cell line SKOV3-FGP demonstrated remarkable tumor size reduction and survival compared to untreated mice [81]. Administration of ST A1-R to nude mice with primary osteosarcoma and lung metastasis was highly effective, especially against metastasis [79]. Bone metastasis is a lethal and morbid late stage of breast cancer. ST A1-R treatment reduced bone growth in highly metastatic human breast cancer in nude mice, and also helped to prevent and inhibit breast cancer bone metastasis [82]. This strain also has the capability of decoying quiescent cancer cells to the S/G2/M phase and sensitizing them to cytotoxic chemotherapy [83].
8. Other Auxotrophic Mutant Strains
Auxotrophic mutant strains are an attractive alternative modulation of Salmonella for killing tumor cells. We engineered ST auxotrophic for tryptophan as a candidate for cancer therapy. This engineered mutant has improved the ability to target and colonize the tumors, reduced the fitness in healthy tissues, and inhibited primary tumor growth and lung metastases [13]. Liang et al. modified the auxotrophic Salmonella vector harboring ∆aroA and ∆purM mutations to deliver antitumor molecules, including the angiogenesis inhibitor endostatin and apoptosis inducer TRAIL, then evaluated anti-tumor efficacy. This strain significantly suppressed tumor growth and prolonged the survival of colon carcinoma and melanoma-bearing mice [84]. Attenuated ST aroA strains that secrete prostate-specific antigens in combination with cholera toxin subunit B induced cytotoxic CD8+ T cells, which efficiently prevented tumor growth in mice [85]. Another application of aroA deleted ST (Δdam, ΔaroA) demonstrated the suppression of tumor angiogenesis, tumor growth, and metastasis while delivering a murine MHC class I antigen epitopes of Legumain, a protein highly expressed in tumor-associated macrophage (TAM) [86].
9. Anti-Tumor Mechanism of Salmonella
The anti-cancer activities of Salmonella have been well documented, but the details of killing tumor cells are still under debate, and accumulated evidence supports the different mechanisms proposed. Various intrinsic features of Salmonella and the tumor microenvironment play a crucial role during the proliferation, progression, and metastasis of tumor cells (Figure 2).
9.1. Hypoxic Environment and Tumor Vasculature
Chronic and acute hypoxia occurs in the tumor due to unevenly distributed and chaotic tumor vasculature, which is associated with decreased oxygen supply and disturbed cell proliferation. A hypoxic environment favors facultative anaerobic bacteria such as Salmonella. Tumor angiogenesis can cause an abnormal vasculature that is more vascularized than the corresponding normal tissue [102]. Such a high and unruly vascularizing tumor microenvironment enhances the chance of Salmonella invasion. Higher localization of such facultative anaerobic bacteria starts killing tumor cells for their nutrients and survival, which is further supported by chaotic vasculature. A positive correlation between higher vascularity and destruction of tumor blood vessels by the tumor-targeting double-auxotrophic mutant ST A1-R has been demonstrated [39].
9.2. Abundant Nutrients and Competitive Nature of Salmonella
The tumor microenvironment hosts diverse micronutrients. A study in murine pancreatic and lung adenocarcinoma models revealed that nutrients available to tumors differ from those present in circulation that influence cancer cell metabolism [87]. Metabolic adaptation in tumors accumulates metabolites that serve as a substrate for the generation of energy and biomass, as well as altering normal gene expression [103]. This creates an immunosuppressive microenvironment inside the tumor that could ease the survival and growth of attenuated auxotrophic bacteria by supplying nutrients and providing protection from the immuno-surveillance of the host [104]. By using this knowledge, auxotrophic Salmonella for leucine, arginine, and tryptophan has been exploited for the treatment of prostate cancer and breast cancer in mice models [13,105].
9.3. Tumor Penetration
The potential for deep tumor penetration has made Salmonella the best candidate for BMCT. Conventional chemotherapeutic drug distribution relies on passive transport, limiting uniform delivery in the inner tumor region. Live Salmonella gathers energy from surrounding abundant nutrients and invades deep tumor tissue. After systemic administration of ST, it is dispersed in all the regions of the solid tumor from the edge to the core center, and apoptosis is induced [99]. Such a spatial distribution of bacteria within tumor tissue is influenced by bacterial motility; bacteria with higher motility penetrate deeper [106]. Other groups believe that ST migration in the tumor is a passive process that is independent of motility and chemotaxis, however, they have used different strains and time points post-infection to examine the tumor-colonization events [35]. Other factors including host immune response can determine tumor penetration. Neutrophils prevent the spread of bacteria from the necrotic region into tumor tissue, and their depletion increases intra-tumor bacterial colonization [107].
9.4. Apoptosis and Autophagy-Inducing Intrinsic Anti-Tumor Action
Numerous investigations have shown that Salmonella has direct killing effects on cancer cells. Accumulation of Salmonella in tumors induces apoptosis [99]. High-resolution multiphoton tomographic images have revealed that genetically engineered ST A1-R infected cancer cells expanded, burst, and finally lost viability [101]. Salmonella has an intrinsic ability to kill cancer cells by inducing both cellular apoptosis and autophagy [100,108]. Although the exact mechanism underlying the induction of apoptosis by Salmonella is less defined, competition for nutrients with cancer cells and the release of bacterial toxins may induce apoptosis. It could also induce autophagy, a scavenger process that is present at a lower level in tumor cells than in their normal counterparts. Salmonella induces the autophagic signaling pathway in a dose- and time-dependent manner via downregulation of the protein kinase B (AKT)/mammalian target of the rapamycin (mTOR) pathway [109]. The AKT/mTOR pathway plays a significant role in cellular physiology and homeostasis. Downregulation of this pathway reduces the expression of matrix metalloproteinase 9 (MMP-9), an oncoprotein involved in metastasis [110].
9.5. Inhibition of Angiogenesis
Angiogenesis plays a vital role in the development and progression of tumors. HIF-1α and VEGF play a significant role in tumor angiogenesis. Salmonella invasion in a tumor can downregulate the expression of HIF-1α and VEGF, and inhibits tumor angiogenesis via the AKT/mTOR pathway [38]. Another mechanism for the suppression of angiogenesis involves a recently identified tumor angiogenesis inhibitor protein, connexin 43 (Cx43), which functions by downregulating VEGF via HIF-1α [111]. Furthermore, higher tumor vasculature has been correlated with the destruction of tumor blood vessels with the tumor-targeting double-auxotrophic mutant ST A1-R [39,112].
9.6. Immunomodulation in Tumor Tissue
Salmonella can manipulate immune components of the tumor niche in favor of tumor inhibition by shifting the tumor microenvironment from immunosuppressive to immunogenic (Figure 1). This process involves alterations to both cellular and soluble components of the immune system which, in turn, affects the phenotypic and functional properties of immune cells. Salmonella infection has been reported to increase the infiltration of macrophages [88], natural killer (NK) cells, CD4+ helper T cells, CD8+ cytotoxic T cells [89], and B cells [90]. Tumor cells release chemo-attractants such as colony-stimulating factor 1 (CSF-1) and the chemokine C-C motif chemokine ligand 2 (CCL2) to recruit monocytes that differentiate into M2 macrophages [113]. M2 macrophage polarization supports the growth and malignancy of tumors by suppressing the anti-tumor immune responses of the host by secreting immunosuppressive molecules, such as arginase 1 (Arg1) [96] and the cytokine IL-10 [114]. Upon Salmonella invasion, TAMs increase the expression of M1 macrophage activation markers, such as the IFN-γ-dependent Sca-1 and MHC class II proteins [88]. This is a paradigm for shifting M2 to M1. These M1 macrophages orchestrate protective anti-tumor immune responses through the expression of nitric oxide synthase (NOS2) and TNF-α [98].
Regulatory T (Treg) cells mitigate anti-tumor immunity by inhibiting tumor antigen-specific cytotoxic T-lymphocytes (CTL) [115]. Intra-tumoral injection of attenuated Salmonella has immunotherapeutic potential, as it reduces Treg cells in a colon cancer model [93]. Salmonella treatment downregulates the cell surface molecule, CD44 present in both Treg and cancer cells [94]. Cx43 is a ubiquitous protein for gap junction formation but is normally lost during melanoma progression. It has been reported to suppress the growth of melanoma and is involved in peptide transfer to activate T cells and induce anti-tumor immunity [116]. Salmonella infection can induce the upregulation of Cx43 and facilitate gap junction formation. Dendritic cells can collect peptide antigens via gap junctions and present antigenic peptides from the tumor cells. Antigenic presentation activates cytotoxic T cells against the tumor antigen, which can ultimately control the growth of distant uninfected tumors [92].
9.7. Orchestration of TAM Function and Polarization
TAMs are a major component of the leukocytic infiltrate of tumors, and have served as a paradigm for cancer-related inflammation, promotion tumor growth, invasion, metastasis, and drug resistance [117]. TAM has an M2-like phenotype, although there is a state of constant transition between the two forms (M1 and M2) [118]. M1 macrophages are well-known for anti-tumor functions, whereas the M2 macrophages promote the occurrence and metastasis of tumor cells, tumor angiogenesis, and tumor progression [119]. TAM functions as a double-edged sword, with the ability to express pro- and anti-tumor activity, and has excellent reprogramming potential toward immunological stimuli such as IFN-γ or IFN-α. Upon bacterial invasion, TAM is associated with an innate response compelled to produce inflammatory cytokines. This stimulation in TAM re-programs immunosuppressive M2 macrophages into immunostimulatory M1 macrophages [120,121]. Salmonella invasion increases high mobility group box 1 (HMGB1) secretion in tumors to coax the polarization of macrophages in favor of an M1-like phenotype, which is reflected by increased inducible nitric oxide synthase (iNOS) and interleukin 1 Beta (IL-1β) production [122]. In a separate investigation, the M2 macrophage phenotype was reversed into the M1 macrophage in a co-culture of M2 macrophage with a genetically engineered ST YB1 strain. This suggests that ST redirects M2:M1 macrophage transition in TAM by switching the macrophage from the CD206high/HLA-DRlow phenotype to the CD206low/HLA-DRhigh phenotype in order to undermine breast tumor growth [123]. Thus, generated M1 macrophages mediate cytotoxicity by releasing tumor-killing molecules ROS and NO [124], as well as by antibody-dependent, cell-mediated cytotoxicity [125]. This event orchestrates the tumor microenvironment toward protective anti-tumor immune responses.
In a different approach, suppression of TAMs can be achieved through designing cancer vaccines against proteins which is overexpress by TAMs, such as Legumain and others. Legumain encodes an asparaginyl endopeptidase that is highly upregulated in murine and human tumor tissues [86]. Administration of Legumain or its MHC class I antigen epitopes as a minigene vaccine through Salmonella induces specific CD8+ T-cell response and suppresses tumor angiogenesis, tumor growth, and metastasis [86,126].
9.8. Release of Cytotoxic Chemicals
The release of cytotoxic compounds such as perforin and granzyme is a defense mechanism during Salmonella treatment which helps to kill cancer cells. The expression of immunomodulatory molecules, such as cytokines and chemokines, stimulates the host immune system to clear tumors. As a consequence, Salmonella has been explored as a vector for the expression of cytotoxic agents such as cytolysin A, Fas ligand (FASL), TNF-α, TRAIL, IL-2, and IL-18 [53,54,66,127,128]. ST engineered to express the proapoptotic cytokine FASL inhibited the growth of primary tumors by an average of 59% for breast tumors and 82% for colon tumors [53]. Likewise, cytotoxic protein (HlyE) was expressed in the hypoxic regions of murine mammary tumors with the aid of a highly hypoxia-inducible promoter that increased tumor necrosis and reduced tumor growth [65]. ST engineered for the secretion of murine TRAIL under the control of the prokaryotic radiation inducible RecA promoter, activated apoptosis, delayed mammary tumor growth, and reduced mortality by 76% [54]. Chen et al. demonstrated similar results for suppressing melanoma by TRAIL under the control of the hypoxia-induced nirB promoter [63].
9.9. Role of the Type III Secretion System
Salmonella harbors a well known virulence factor, the type III secretion system (T3SS) encoded by “Salmonella pathogenicity island 1” (SPI1) to facilitate direct injection of effector proteins into host cells, interfere with intracellular signaling pathways, manipulate host cell cytoskeleton network, and enable colonization [129]. They are adapted to produce and store most of the effector proteins to secrete through the T3SS into the host cells [130]. This powerful system can be manipulated in order to translocate heterogeneous antigens by fusion with effector proteins. Cellular invasion promoted by SPI1 is supported by injecting bacterial effector molecules encoded by Salmonella pathogenicity island 2 (SPI2), which attempt to restore themselves inside the host by restructuring endosomes into Salmonella-containing vacuoles (SCVs) to evade host defense mechanisms and enable intracellular survival [131,132]. Thus, the invasion of the native form or bacterial delivery system is facilitated by the combined function of SPI1 and SPI2. Thus, the established Salmonella can induce apoptosis, and it has been supported by the apoptotic tumor cell death through the expression of Salmonella effectors such as a virulence factor SpvB (an ADP-ribosyl transferase enzyme) through the activation of caspases 3 and 7 [133]. Salmonella effector SipA has been reported to downregulate the expression of P-glycoprotein which is highly expressed in several cancers such as colon, breast, kidney, and lymphoma [134,135].
Regions of T3SS proteins such as needles, inner rod proteins, and flagellins are perceived by nucleotide-binding oligomerization domain (NOD)-like receptor (NLR) family apoptosis inhibitory protein (NAIP) to activate the NAIP-NLR family caspase-associated recruitment domain-containing protein 4 (NLRC4). This results in NLRC4 inflammasome assembly and triggers the cleavage and activation of caspase-1 and caspase-8. Activation of caspase-1 mediates pyroptosis and stimulation of caspase-8 triggers caspase-3/7 promoting apoptosis that ultimately results in inflammatory death of host cells termed as PANoptosis [136,137,138]. Internalization of effector proteins modulates the host immune response and secretes proinflammatory cytokines or rearranges the host cell cytoskeleton, facilitating bacterial invasion [91,129]. In order to strengthen the antigen delivery for cancer vaccine development, Salmonella T3SS is a promising strategy. It was demonstrated that the delivery of the NY-ESO-1 tumor antigen through T3SS elicited the regression of established NY-ESO-1-expressing tumors in mice [139]. T3SS from Salmonella has been exploited in order to translocate heterologous antigens MHC class I-peptide p60 (217–225) from Listeria monocytogenes into the cytosol of host cells, which induced antigen-specific CD8 T cells, for significant tumor regression [140]. SPI2 effector protein SseF was used to deliver tumor-associated antigens (TAAs) survivin into the cytosol of antigen-presenting cells for optimal immunogenicity [141]. Likewise, another SPI2 effector protein, SspH2, was used to produce a chimeric construct with the p60 protein of Listeria monocytogenes, which induces a p60-specific CD4 and CD8 T-cell response in vaccinated mice [142]. The assisted delivery of TAA via SPI-2-regulated T3SS produced anti-tumor activity in a mouse model.
10. Current Approach with Combination Therapy
Salmonella has been used with other therapeutic agents to enhance the efficacy of anti-cancer activities. It has been used in combination with chemotherapy, radiotherapy, immune checkpoint inhibitors, and immunomodulatory cytokines. The combined administration of Salmonella with chemotherapy reduces toxicity compared with individual therapy with bacteria or chemotherapeutics. For this, a murine melanoma model was treated with VNP20009 and cyclophosphamide, which induced a significant decrease in microvessel density and serum VEGF levels compared with either treatment alone. [143]. Similarly, the combination of anti-angiogenic agent HM-3 (a polypeptide inhibiting angiogenesis) and VNP20009 harboring expression plasmids for siRNA targeting Sox2 demonstrated efficient treatment for lung cancer [144]. Another well known ST A1-R strain was implemented in combination with the chemotherapeutic drugs temozolomide, doxorubicin, and anti-angiogenic agents, which significantly suppressed the growth of tumors in patient-derived orthotopic xenograft models [59,60,145]. The co-administration of radiotherapy and BMCT produced prominent anti-tumor effects compared to either of the treatments alone. The combination of X-rays either with VNP20009 or ΔppGpp ST expressing cytolysin A (ClyA) or γ-radiation with Salmonella BRD509 induced a significant suppression of the tumor or delayed tumor growth [146,147,148]. In addition, the treatment with A1-R post-surgical excision of tumors significantly inhibited surgery-induced breast cancer metastasis [149]. In combination therapy, the use of prodrug strategy along with Salmonella-expressing, prodrug-activating enzymes such as HSV TK, carboxypeptidase G2 (CPG2), and cytosine deaminase have more promising tumor retardation capabilities compared to the use of the therapeutic strain alone [15,56,150].
11. Cancer Vaccines Delivered by Salmonella
Being an intracellular pathogen, with vast survival in different organs of the host, attenuated Salmonella has been widely used as a vaccine delivery system against various diseases [151]. As mentioned earlier, Salmonella is a multifaceted antagonist for cancer [152,153], apart from that, using auxotrophic Salmonella as a therapeutic and prophylactic vaccine delivery system is also an ideal strategy. Medina et al. have demonstrated the anti-cancer effect of auxotrophic ST (∆aroA) as a vaccine delivery system that expresses β-gal as a model TAA against aggressive fibrosarcoma [154]. In another study, SPI-2 and T3SS of Salmonella were used to deliver survivin as a TAA into antigen-presenting cells, and the PsifB::sseJ promoter/effector combination was found to have an excellent anti-cancer immune response of CD8 infiltration in the tumor environment [155]. Similarly, elevated effector-memory CTL responses against CT26 colon cancer and orthotopic delayed brain tumor glioblastoma in mice were found after immunization with survivin, which was fused to the SseF effector protein and kept under the regulation of SsrB, the key regulator of SPI2 [141]. Heat shock protein 70, as an immuno-chaperone fused with SopE of Salmonella T3SS, has elicited a considerable CTL response against murine melanoma [156]. A multi-antigen DNA vaccine encoding fusion antigenic domains of tyrosine hydroxylase, survivin, and PHOX2B, delivered by auxotrophic ST (∆aroA, ∆guaAB), has been demonstrated to exhibit significant elicitation of the CTL response, INFγ production, and excellent suppression of neuroblastoma in a mouse model [157]. In addition to the CTL responses, the elicitation of humoral response by a Salmonella-based oral DNA vaccine with a MG7-Ag mimotope against gastric cancer was confirmed [158]. An H2O2-inactivated S. Typhimurium RE88 (∆aroA, ∆dam) has been established to induce anti-cancer immunity by using ovalbumin as a model antigen [159]. Salmonella has also been studied for the expression of oncogenic virus antigens. A recombinant ST that produced Human Papillomavirus Type 16 (HPV16) L1 Virus-like Particles (VLPs) induced the anti-tumor immune response in prophylactic as well as therapeutic contexts [160]. The same group has constructed a Salmonella that has expressed major capsid protein L1 of HPV16 virus via plasmid, and has shown the induction of anti-HPV16 neutralizing and humoral immune responses [161]. They have also demonstrated the intravaginal immunization of the HPV16-L1 Salmonella construct and its innate, adaptive, Th1, and Th2 mucosal immune responses [162]. Thus, Salmonella can be used to deliver oncogenic viral antigens for prophylactic vaccine development. In addition, Salmonella infection triggers the formation of gap junctions in melanoma that are typically lacking in tumor cells. The transfer of tumor antigens to dendritic cells and the resultant induction of immune responses depend on these gap junctions [92]. Moreover, Salmonella has the ability to induce MHC class I and II immune responses by delivering cancer-related antigens via bacterial surface and translocating the antigen or its gene to the antigen-presenting cells, respectively. Salmonella has the virtue of being used as a delivery vehicle for extrinsic cancer antigens, oncogenic viral antigens, and to display the intrinsic antigens of the active tumor to achieve anti-cancer immunity based on these aspects.
12. Application of Salmonella in Tumor Targeting and Detection
Tumor targeting and accumulation phenotypes have made Salmonella the best player in the creation of genetically engineered strains for detecting tumors. Strains expressing fluorescent proteins are well studied for the purpose of visualizing and locating the tumor region in vivo [16]. Another approach for positioning tumor-specific Salmonella used positron emission tomography which locates the engineered ST VNP20009 strain in tumors by expressing the HSV1-TK reporter gene that can selectively phosphorylate radiolabeled 2′-Fluro-1-β-D-arabinofuranosyl-5-iodo-uracil [163].
Salmonella was engineered to express the fluorescent protein ZsGreen. It has a high sensitivity that can detect tumors 2600 times smaller than the current limit of tomographic techniques [164]. Since Salmonella preferentially accumulates in tumors and microscopic metastases, this approach would provide a method to detect a tumor, monitor treatment efficacy, and identify metastatic onset.
13. Clinical Trials
The success of attenuated ST strain VNP20009 with chromosomal deletion of the purI and msbB genes in order to target tumor and inhibit its growth in mice takes the lead for a Phase I human clinical trial in metastatic cancer patients through intravenous infusion [33]. In this study, none of the patients experienced tumor regression, although VNP20009 could be safely administered to patients. Tumor localization was noted at the highest tolerated dose in only 3 patients [33]. Further improvement on the VNP20009 was incorporated by inserting a gene encoding E. coli cytosine deaminase (CD) and designated the strain as TAPET-CD [165]. Three patients received an intratumoral injection of TAPET-CD; two patients had intratumor evidence of bacterial colonization that persisted for at least 15 days after the initial injection. The conversion of 5-fluorocytosine to 5-fluorouracil as a result of CD expression was demonstrated in these two patients with a tumor-to-plasma ratio of 3:1. This signifies the localization of TAPET-CD in tumors with minimal systemic spread. In another study, oral vaccination with live attenuated Salmonella typhi, carrying an expression plasmid encoding VEGFR2, increased vaccine-specific T cell responses in patients with advanced pancreatic cancer [166]. Aside from human clinical trials, VNP20009 has been administered in a Phase I trial on dogs. The study revealed bacterial tumor localization in 42% of cases and major anti-tumor response in 15% of cases [70]. The exciting anti-tumor activity in tumor-bearing dogs or mice models was not reflected in human clinical trials. This discrepancy in anti-tumor effects of engineered ST in animal and human models demand an in-depth study on the anti-tumor mechanism, especially in humans. Human clinical trials for anti-cancer treatment using Salmonella are listed in Table 2.
14. Limitations of Bacteria-Mediated Cancer Therapy
The race for bacteria-mediated tumor therapy is at its peak, as it holds potential for the treatment of different tumors. The main concern while using bacterial vaccines is their ubiquitous nature. Since many bacteria, including Salmonella and Listeria, used in BMCT are prevalent in the environment, and their pre-exposure commonly induces immunity against these pathogens. Therefore, the preexisting or vaccine-induced vector-specific immunity could potentially block therapeutic genes and vaccine delivery. In a survey, it was found that there was a preexisting cellular immunity to L. monocytogenes in 60% of human donors tested [168]. The current practice of genetic modification by deletion of genes responsible for the biosynthesis of lipopolysaccharide, amino acids, or purines affects bacterial virulence that modulates bacterial invasion, tumor localization, and immune stimulation. Excessive attenuation downregulates invasive potential, while less attenuation is pathogenic. Thus, balanced attenuation is a major concern for the development of effective anti-tumor ST strains, since insufficient attenuation might result in proliferation at non-designated organs, and may even cause severe septic shock while excessive attenuation affects its invasion potential and immunogenic presentation. While administering ST for treatment, termination of bacteria from the treated patient is another issue that needs to be resolved. Although antibiotics are the remedy, there is a high chance of acquiring antibiotic resistance. So far, bacterial application for cancer therapy needs to address safety issues despite having a promising success rate. SMCT is still facing many problems, and as a result, most phase I clinical trials have not been satisfactory. Furthermore, intrinsic factors such as tumor architecture, growth rate, and blood supply, as well as extrinsic factors, especially the entry of bacteria into tumors, their growth within tumors, and clearance from the peripheral circulation and tumors, might have a pivotal role in the success of SMCT in clinical application. Therefore, the complicated interactive mechanism between Salmonella, tumor cells, host immune cells, and inflammatory reactions needs maximum exploration to understand the process so that SMCT could be applied to human cancer therapy.
15. Future Perspectives and Conclusions
In this review, we elaborated on the anti-cancer therapeutic potential of Salmonella. Special features such as tumor-targeting capacity, tumor tissue penetration, immunomodulatory effects, genomic engineering plasticity, and delivery efficacy highlighted Salmonella as a genuine candidate for cancer therapy. Current knowledge was elaborated on in order to facilitate the development of safer and more effective Salmonella bacterial strains for cancer treatment. Although the anti-tumor potential of Salmonella has been elucidated, the precise mechanism of immune activation against cancer by Salmonella is still not clear. Even though the anti-tumor potential of Salmonella has been established in murine models and dog phase trials, clinical trials in human patients have not been effective. The success rate of tumor suppression by Salmonella in humans is still inadequate. Thus, Salmonella-based human oncotherapy still demands immense immunological and molecular mechanism studies on human prospects. Cancer patients and animals are highly immuno-compromised; thus, even mild systemic bacterial infection may lead to serious adverse effects. Hence, avirulent, non-endotoxic Salmonella strains are highly envisaged. Moreover, Salmonella-based oncotherapy requires consecutive inoculation. Upon subsequent inoculation, Salmonella should not be nullified by host immunity. Therefore, strains which are less immunogenic but highly invasive to tumors have a good scope. Genetic manipulation of Salmonella by simple genetic engineering or synthetic bioengineering is well known to induce the production of anti-cancer drugs. Furthermore, BMCT is the proof of concept that can either be used as a monotherapy or in combination with other anti-cancer therapies in order to achieve better clinical outcomes. The ongoing continuous development and innovation of Salmonella-based cancer therapy possesses great promise for a new paradigm of cancer therapy that could pave the way for its succeeding emergence as one of the mainstream therapeutic interventions to tackle various types of cancer.
Author Contributions
Conceptualization, R.P.A. and J.H.L.; methodology, R.P.A., C.S. and A.S.; formal analysis, R.P.A., C.S. and A.S.; investigation, R.P.A., C.S. and A.S.; resources, R.P.A., C.S. and A.S.; data curation, R.P.A.; writing—original draft preparation, R.P.A.; writing—review and editing, R.P.A., C.S., and A.S.; visualization, R.P.A. and J.H.L.; supervision, J.H.L.; project administration, J.H.L.; funding acquisition, J.H.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National University Development Project in 2021 and by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (2019R1A6A1A03033084).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Authors are grateful to all who assisted in the preparation of this manuscript. All individuals included in this section have consented to the acknowledgement.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Minchinton, A.I.; Tannock, I.F. Drug penetration in solid tumours. Nat. Rev. Cancer 2006, 6, 583–592. [Google Scholar] [CrossRef] [PubMed]
- St Jean, A.T.; Zhang, M.; Forbes, N.S. Bacterial therapies: Completing the cancer treatment toolbox. Curr. Opin. Biotechnol. 2008, 19, 511–517. [Google Scholar] [CrossRef] [Green Version]
- Keung, E.Z.; Fairweather, M.; Raut, C.P. Surgical Management of Metastatic Disease. Surg. Clin. North Am. 2016, 96, 1175–1192. [Google Scholar] [CrossRef] [PubMed]
- Dutt, S.; Ahmed, M.M.; Loo, B.W., Jr.; Strober, S. Novel Radiation Therapy Paradigms and Immunomodulation: Heresies and Hope. Semin. Radiat. Oncol. 2020, 30, 194–200. [Google Scholar] [CrossRef]
- Kocakavuk, E.; Anderson, K.J.; Varn, F.S.; Johnson, K.C.; Amin, S.B.; Sulman, E.P.; Lolkema, M.P.; Barthel, F.P.; Verhaak, R.G.W. Radiotherapy is associated with a deletion signature that contributes to poor outcomes in patients with cancer. Nat. Genet. 2021, 53, 1088–1096. [Google Scholar] [CrossRef]
- Jing, X.; Yang, F.; Shao, C.; Wei, K.; Xie, M.; Shen, H.; Shu, Y. Role of hypoxia in cancer therapy by regulating the tumor microenvironment. Mol. Cancer 2019, 18, 157. [Google Scholar] [CrossRef] [Green Version]
- Rohwer, N.; Cramer, T. Hypoxia-mediated drug resistance: Novel insights on the functional interaction of HIFs and cell death pathways. Drug Resist. Updat. 2011, 14, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Gray, L.H.; Conger, A.D.; Ebert, M.; Hornsey, S.; Scott, O.C. The concentration of oxygen dissolved in tissues at the time of irradiation as a factor in radiotherapy. Br. J. Radiol. 1953, 26, 638–648. [Google Scholar] [CrossRef]
- Dang, L.H.; Bettegowda, C.; Huso, D.L.; Kinzler, K.W.; Vogelstein, B. Combination bacteriolytic therapy for the treatment of experimental tumors. Proc. Natl. Acad. Sci. USA 2001, 98, 15155–15160. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, V.H.; Min, J.J. Salmonella-Mediated Cancer Therapy: Roles and Potential. Nucl. Med. Mol. Imaging 2017, 51, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Tjuvajev, J.; Blasberg, R.; Luo, X.; Zheng, L.M.; King, I.; Bermudes, D. Salmonella-based tumor-targeted cancer therapy: Tumor amplified protein expression therapy (TAPET) for diagnostic imaging. J. Control. Release 2001, 74, 313–315. [Google Scholar] [CrossRef]
- Jawalagatti, V.; Kirthika, P.; Lee, J.H. Targeting primary and metastatic tumor growth in an aggressive breast cancer by engineered tryptophan auxotrophic Salmonella Typhimurium. Mol. Ther. Oncolytics 2022, 25, 350–363. [Google Scholar] [CrossRef]
- McCarthy, E.F.; The Toxins of William, B. Coley and the treatment of bone and soft-tissue sarcomas. Iowa Orthop. J. 2006, 26, 154–158. [Google Scholar]
- Pawelek, J.M.; Low, K.B.; Bermudes, D. Tumor-targeted Salmonella as a novel anticancer vector. Cancer Res. 1997, 57, 4537–4544. [Google Scholar] [PubMed]
- Yu, Y.A.; Shabahang, S.; Timiryasova, T.M.; Zhang, Q.; Beltz, R.; Gentschev, I.; Goebel, W.; Szalay, A.A. Visualization of tumors and metastases in live animals with bacteria and vaccinia virus encoding light-emitting proteins. Nat. Biotechnol. 2004, 22, 313–320. [Google Scholar] [CrossRef]
- Parker, R.C.; Plummer, H.C.; Siebenmann, C.O.; Chapman, M.G. Effect of histolyticus infection and toxin on transplantable mouse tumors. Proc. Soc. Exp. Biol. Med. 1947, 66, 461–467. [Google Scholar] [CrossRef]
- Malmgren, R.A.; Flanigan, C.C. Localization of the vegetative form of Clostridium tetani in mouse tumors following intravenous spore administration. Cancer Res. 1955, 15, 473–478. [Google Scholar]
- Kohwi, Y.; Imai, K.; Tamura, Z.; Hashimoto, Y. Antitumor effect of Bifidobacterium infantis in mice. Gan 1978, 69, 613–618. [Google Scholar]
- Maletzki, C.; Linnebacher, M.; Kreikemeyer, B.; Emmrich, J. Pancreatic cancer regression by intratumoural injection of live Streptococcus pyogenes in a syngeneic mouse model. Gut 2008, 57, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Arakawa, M.; Sugiura, K.; Reilly, H.C.; Stock, C.C. Oncolytic effect of Proteus mirabilis upon tumor-bearing animals. II. Effect on transplantable mouse and rat tumors. Gan 1968, 59, 117–122. [Google Scholar] [PubMed]
- Moese, J.R.; Moese, G. Oncolysis by Clostridia. I. Activity of Clostridium Butyricum (M-55) and Other Nonpathogenic Clostridia against the Ehrlich Carcinoma. Cancer Res. 1964, 24, 212–216. [Google Scholar] [PubMed]
- Mohr, U.; Hondius Boldingh, W.; Emminger, A.; Behagel, H.A. Oncolysis by a new strain of Clostridium. Cancer Res. 1972, 32, 1122–1128. [Google Scholar] [PubMed]
- Iftekhar, A.; Berger, H.; Bouznad, N.; Heuberger, J.; Boccellato, F.; Dobrindt, U.; Hermeking, H.; Sigal, M.; Meyer, T.F. Genomic aberrations after short-term exposure to colibactin-producing E. coli transform primary colon epithelial cells. Nat. Commun. 2021, 12, 1003. [Google Scholar] [CrossRef] [PubMed]
- Kwong, T.N.Y.; Wang, X.; Nakatsu, G.; Chow, T.C.; Tipoe, T.; Dai, R.Z.W.; Tsoi, K.K.K.; Wong, M.C.S.; Tse, G.; Chan, M.T.V.; et al. Association Between Bacteremia From Specific Microbes and Subsequent Diagnosis of Colorectal Cancer. Gastroenterology 2018, 155, 383–390.e8. [Google Scholar] [CrossRef] [PubMed]
- Ohara, T.; Yoshino, K.; Kitajima, M. Possibility of preventing colorectal carcinogenesis with probiotics. Hepatogastroenterology 2010, 57, 1411–1415. [Google Scholar]
- Caygill, C.P.; Braddick, M.; Hill, M.J.; Knowles, R.L.; Sharp, J.C. The association between typhoid carriage, typhoid infection and subsequent cancer at a number of sites. Eur. J. Cancer Prev. 1995, 4, 187–193. [Google Scholar] [CrossRef]
- Pandey, M.; Choudhury, H.; Vijayagomaran, P.A.; Lian, P.N.P.; Ning, T.J.; Wai, N.Z.; Xian-Zhuang, N.; Le Er, C.; Rahmah, N.S.N.; Kamaruzzaman, N.D.B.; et al. Recent Update on Bacteria as a Delivery Carrier in Cancer Therapy: From Evil to Allies. Pharm. Res. 2022, 39, 1115–1134. [Google Scholar] [CrossRef]
- Jawalagatti, V.; Kirthika, P.; Lee, J.H. Oral mRNA Vaccines against Infectious Diseases—A Bacterial Perspective [Invited]. Front. Immunol. 2022, 13, 884862. [Google Scholar] [CrossRef]
- Forbes, N.S. Engineering the perfect (bacterial) cancer therapy. Nat. Rev. Cancer 2010, 10, 785–794. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.J.; Sun, Y.; Nabel, G.J. Regulation of the proinflammatory effects of Fas ligand (CD95L). Science 1998, 282, 1714–1717. [Google Scholar] [CrossRef] [PubMed]
- Clairmont, C.; Lee, K.C.; Pike, J.; Ittensohn, M.; Low, K.B.; Pawelek, J.; Bermudes, D.; Brecher, S.M.; Margitich, D.; Turnier, J.; et al. Biodistribution and genetic stability of the novel antitumor agent VNP20009, a genetically modified strain of Salmonella typhimurium. J. Infect. Dis. 2000, 181, 1996–2002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toso, J.F.; Gill, V.J.; Hwu, P.; Marincola, F.M.; Restifo, N.P.; Schwartzentruber, D.J.; Sherry, R.M.; Topalian, S.L.; Yang, J.C.; Stock, F.; et al. Phase I study of the intravenous administration of attenuated Salmonella typhimurium to patients with metastatic melanoma. J. Clin. Oncol. 2002, 20, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Liang, K.; Liu, Q.; Li, P.; Luo, H.; Wang, H.; Kong, Q. Genetically engineered Salmonella Typhimurium: Recent advances in cancer therapy. Cancer Lett. 2019, 448, 168–181. [Google Scholar] [CrossRef] [PubMed]
- Stritzker, J.; Weibel, S.; Seubert, C.; Gotz, A.; Tresch, A.; van Rooijen, N.; Oelschlaeger, T.A.; Hill, P.J.; Gentschev, I.; Szalay, A.A. Enterobacterial tumor colonization in mice depends on bacterial metabolism and macrophages but is independent of chemotaxis and motility. Int. J. Med. Microbiol. 2010, 300, 449–456. [Google Scholar] [CrossRef]
- Kasinskas, R.W.; Forbes, N.S. Salmonella typhimurium specifically chemotax and proliferate in heterogeneous tumor tissue in vitro. Biotechnol. Bioeng. 2006, 94, 710–721. [Google Scholar] [CrossRef]
- Kasinskas, R.W.; Forbes, N.S. Salmonella typhimurium lacking ribose chemoreceptors localize in tumor quiescence and induce apoptosis. Cancer Res. 2007, 67, 3201–3209. [Google Scholar] [CrossRef] [Green Version]
- Tu, D.G.; Chang, W.W.; Lin, S.T.; Kuo, C.Y.; Tsao, Y.T.; Lee, C.H. Salmonella inhibits tumor angiogenesis by downregulation of vascular endothelial growth factor. Oncotarget 2016, 7, 37513–37523. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Zhang, L.; Hoffman, R.M.; Zhao, M. Vessel destruction by tumor-targeting Salmonella typhimurium A1-R is enhanced by high tumor vascularity. Cell Cycle 2010, 9, 4518–4524. [Google Scholar] [CrossRef] [Green Version]
- Henderson, B.; Poole, S.; Wilson, M. Microbial/host interactions in health and disease: Who controls the cytokine network? Immunopharmacology 1996, 35, 1–21. [Google Scholar] [CrossRef]
- Zhou, S.; Gravekamp, C.; Bermudes, D.; Liu, K. Tumour-targeting bacteria engineered to fight cancer. Nat. Rev. Cancer 2018, 18, 727–743. [Google Scholar] [CrossRef] [PubMed]
- al-Ramadi, B.K.; Fernandez-Cabezudo, M.J.; El-Hasasna, H.; Al-Salam, S.; Bashir, G.; Chouaib, S. Potent anti-tumor activity of systemically-administered IL2-expressing Salmonella correlates with decreased angiogenesis and enhanced tumor apoptosis. Clin. Immunol. 2009, 130, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.; Duong, M.T.; Zuo, C.; Qin, Y.; Zhang, Y.; Guo, Y.; Hong, Y.; Zheng, J.H.; Min, J.J. Targeting of pancreatic cancer cells and stromal cells using engineered oncolytic Salmonella typhimurium. Mol. Ther. 2022, 30, 662–671. [Google Scholar] [CrossRef] [PubMed]
- de Lima Fragelli, B.D.; Camillo, L.; de Almeida Rodolpho, J.M.; de Godoy, K.F.; de Castro, C.A.; Brassolatti, P.; da Silva, A.J.; Borra, R.C.; de Freitas Anibal, F. Antitumor Effect of IL-2 and TRAIL Proteins Expressed by Recombinant Salmonella in Murine Bladder Cancer Cells. Cell. Physiol. Biochem. 2021, 55, 460–476. [Google Scholar]
- Guan, G.F.; Zhao, M.; Liu, L.M.; Jin, C.S.; Sun, K.; Zhang, D.J.; Yu, D.J.; Cao, H.W.; Lu, Y.Q.; Wen, L.J. Salmonella typhimurium mediated delivery of Apoptin in human laryngeal cancer. Int. J. Med. Sci. 2013, 10, 1639–1648. [Google Scholar] [CrossRef] [Green Version]
- Lim, D.; Kim, K.S.; Kim, H.; Ko, K.C.; Song, J.J.; Choi, J.H.; Shin, M.; Min, J.J.; Jeong, J.H.; Choy, H.E. Anti-tumor activity of an immunotoxin (TGFalpha-PE38) delivered by attenuated Salmonella typhimurium. Oncotarget 2017, 8, 37550–37560. [Google Scholar] [CrossRef] [Green Version]
- Blache, C.A.; Manuel, E.R.; Kaltcheva, T.I.; Wong, A.N.; Ellenhorn, J.D.; Blazar, B.R.; Diamond, D.J. Systemic delivery of Salmonella typhimurium transformed with IDO shRNA enhances intratumoral vector colonization and suppresses tumor growth. Cancer Res. 2012, 72, 6447–6456. [Google Scholar] [CrossRef] [Green Version]
- Raman, V.; Van Dessel, N.; Hall, C.L.; Wetherby, V.E.; Whitney, S.A.; Kolewe, E.L.; Bloom, S.M.K.; Sharma, A.; Hardy, J.A.; Bollen, M.; et al. Intracellular delivery of protein drugs with an autonomously lysing bacterial system reduces tumor growth and metastases. Nat. Commun. 2021, 12, 6116. [Google Scholar] [CrossRef]
- Deng, J.; Guo, Y.; Jiang, Z.; Yang, M.; Li, H.; Wang, J. Enhancement of ovarian cancer chemotherapy by delivery of multidrug-resistance gene small interfering RNA using tumor targeting Salmonella. J. Obs. Gynaecol. Res. 2015, 41, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Phan, T.X.; Nguyen, V.H.; Dinh-Vu, H.V.; Zheng, J.H.; Yun, M.; Park, S.G.; Hong, Y.; Choy, H.E.; Szardenings, M.; et al. Salmonella typhimurium Suppresses Tumor Growth via the Pro-Inflammatory Cytokine Interleukin-1beta. Theranostics 2015, 5, 1328–1342. [Google Scholar] [CrossRef] [Green Version]
- Akoachere, J.F.; Tanih, N.F.; Ndip, L.M.; Ndip, R.N. Phenotypic characterization of Salmonella typhimurium isolates from food-animals and abattoir drains in Buea, Cameroon. J. Health Popul. Nutr. 2009, 27, 612–618. [Google Scholar]
- Semenov, A.V.; van Overbeek, L.; Termorshuizen, A.J.; van Bruggen, A.H. Influence of aerobic and anaerobic conditions on survival of Escherichia coli O157:H7 and Salmonella enterica serovar Typhimurium in Luria-Bertani broth, farm-yard manure and slurry. J. Env. Manag. 2011, 92, 780–787. [Google Scholar] [CrossRef] [PubMed]
- Loeffler, M.; Le’Negrate, G.; Krajewska, M.; Reed, J.C. Inhibition of tumor growth using salmonella expressing Fas ligand. J. Natl. Cancer Inst. 2008, 100, 1113–1116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganai, S.; Arenas, R.B.; Forbes, N.S. Tumour-targeted delivery of TRAIL using Salmonella typhimurium enhances breast cancer survival in mice. Br. J. Cancer 2009, 101, 1683–1691. [Google Scholar] [CrossRef] [PubMed]
- King, I.; Bermudes, D.; Lin, S.; Belcourt, M.; Pike, J.; Troy, K.; Le, T.; Ittensohn, M.; Mao, J.; Lang, W.; et al. Tumor-targeted Salmonella expressing cytosine deaminase as an anticancer agent. Hum. Gene 2002, 13, 1225–1233. [Google Scholar] [CrossRef] [Green Version]
- Friedlos, F.; Lehouritis, P.; Ogilvie, L.; Hedley, D.; Davies, L.; Bermudes, D.; King, I.; Martin, J.; Marais, R.; Springer, C.J. Attenuated Salmonella targets prodrug activating enzyme carboxypeptidase G2 to mouse melanoma and human breast and colon carcinomas for effective suicide gene therapy. Clin. Cancer Res. 2008, 14, 4259–4266. [Google Scholar] [CrossRef] [Green Version]
- Murakami, T.; Hiroshima, Y.; Zhang, Y.; Zhao, M.; Kiyuna, T.; Hwang, H.K.; Miyake, K.; Homma, Y.; Mori, R.; Matsuyama, R.; et al. Tumor-Targeting Salmonella typhimurium A1-R Promotes Tumoricidal CD8(+) T Cell Tumor Infiltration and Arrests Growth and Metastasis in a Syngeneic Pancreatic-Cancer Orthotopic Mouse Model. J. Cell Biochem. 2018, 119, 634–639. [Google Scholar] [CrossRef]
- Igarashi, K.; Kawaguchi, K.; Kiyuna, T.; Miyake, K.; Miyake, M.; Li, S.; Han, Q.; Tan, Y.; Zhao, M.; Li, Y.; et al. Tumor-targeting Salmonella typhimurium A1-R combined with recombinant methioninase and cisplatinum eradicates an osteosarcoma cisplatinum-resistant lung metastasis in a patient-derived orthotopic xenograft (PDOX) mouse model: Decoy, trap and kill chemotherapy moves toward the clinic. Cell Cycle 2018, 17, 801–809. [Google Scholar]
- Murakami, T.; DeLong, J.; Eilber, F.C.; Zhao, M.; Zhang, Y.; Zhang, N.; Singh, A.; Russell, T.; Deng, S.; Reynoso, J.; et al. Tumor-targeting Salmonella typhimurium A1-R in combination with doxorubicin eradicate soft tissue sarcoma in a patient-derived orthotopic xenograft (PDOX) model. Oncotarget 2016, 7, 12783–12790. [Google Scholar] [CrossRef] [Green Version]
- Kawaguchi, K.; Igarashi, K.; Murakami, T.; Chmielowski, B.; Kiyuna, T.; Zhao, M.; Zhang, Y.; Singh, A.; Unno, M.; Nelson, S.D.; et al. Tumor-targeting Salmonella typhimurium A1-R combined with temozolomide regresses malignant melanoma with a BRAF-V600E mutation in a patient-derived orthotopic xenograft (PDOX) model. Oncotarget 2016, 7, 85929–85936. [Google Scholar] [CrossRef] [Green Version]
- Kawaguchi, K.; Igarashi, K.; Murakami, T.; Kiyuna, T.; Zhao, M.; Zhang, Y.; Nelson, S.D.; Russell, T.A.; Dry, S.M.; Singh, A.S.; et al. Salmonella typhimurium A1-R targeting of a chemotherapy-resistant BRAF-V600E melanoma in a patient-derived orthotopic xenograft (PDOX) model is enhanced in combination with either vemurafenib or temozolomide. Cell Cycle 2017, 16, 1288–1294. [Google Scholar] [CrossRef] [PubMed]
- Zuo, S.G.; Chen, Y.; Wu, Z.P.; Liu, X.; Liu, C.; Zhou, Y.C.; Wu, C.L.; Jin, C.G.; Gu, Y.L.; Li, J.; et al. Orally administered DNA vaccine delivery by attenuated Salmonella typhimurium targeting fetal liver kinase 1 inhibits murine Lewis lung carcinoma growth and metastasis. Biol. Pharm. Bull. 2010, 33, 174–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Yang, B.; Cheng, X.; Qiao, Y.; Tang, B.; Chen, G.; Wei, J.; Liu, X.; Cheng, W.; Du, P.; et al. Salmonella-mediated tumor-targeting TRAIL gene therapy significantly suppresses melanoma growth in mouse model. Cancer Sci. 2012, 103, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Zhu, X.; Chen, L.; Li, S.; Ren, D. Oral administration of attenuated S. typhimurium carrying shRNA-expressing vectors as a cancer therapeutic. Cancer Biol. 2008, 7, 145–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, R.M.; Green, J.; Williams, P.J.; Tazzyman, S.; Hunt, S.; Harmey, J.H.; Kehoe, S.C.; Lewis, C.E. Bacterial delivery of a novel cytolysin to hypoxic areas of solid tumors. Gene 2009, 16, 329–339. [Google Scholar] [CrossRef] [Green Version]
- Loeffler, M.; Le’Negrate, G.; Krajewska, M.; Reed, J.C. IL-18-producing Salmonella inhibit tumor growth. Cancer Gene 2008, 15, 787–794. [Google Scholar] [CrossRef] [Green Version]
- Hoiseth, S.K.; Stocker, B.A. Aromatic-dependent Salmonella typhimurium are non-virulent and effective as live vaccines. Nature 1981, 291, 238–239. [Google Scholar] [CrossRef]
- Low, K.B.; Ittensohn, M.; Le, T.; Platt, J.; Sodi, S.; Amoss, M.; Ash, O.; Carmichael, E.; Chakraborty, A.; Fischer, J.; et al. Lipid A mutant Salmonella with suppressed virulence and TNFalpha induction retain tumor-targeting in vivo. Nat. Biotechnol. 1999, 17, 37–41. [Google Scholar] [CrossRef]
- Felgner, S.; Kocijancic, D.; Frahm, M.; Curtiss, R., 3rd; Erhardt, M.; Weiss, S. Optimizing Salmonella enterica serovar Typhimurium for bacteria-mediated tumor therapy. Gut Microbes 2016, 7, 171–177. [Google Scholar] [CrossRef] [Green Version]
- Thamm, D.H.; Kurzman, I.D.; King, I.; Li, Z.; Sznol, M.; Dubielzig, R.R.; Vail, D.M.; MacEwen, E.G. Systemic administration of an attenuated, tumor-targeting Salmonella typhimurium to dogs with spontaneous neoplasia: Phase I evaluation. Clin. Cancer Res. 2005, 11, 4827–4834. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, C.; Nemunaitis, J. A phase I trial of genetically modified Salmonella typhimurium expressing cytosine deaminase (TAPET-CD, VNP20029) administered by intratumoral injection in combination with 5-fluorocytosine for patients with advanced or metastatic cancer. Protocol no: CL-017. Version: April 9, 2001. Hum. Gene 2001, 12, 1594–1596. [Google Scholar]
- Luo, X.; Li, Z.; Lin, S.; Le, T.; Ittensohn, M.; Bermudes, D.; Runyab, J.D.; Shen, S.Y.; Chen, J.; King, I.C.; et al. Antitumor effect of VNP20009, an attenuated Salmonella, in murine tumor models. Oncol. Res. 2001, 12, 501–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Lu, M.; Lai, Y.; Zhang, X.; Li, Y.; Mao, P.; Liang, Z.; Mu, Y.; Lin, Y.; Zhao, A.Z.; et al. Inhibition of acute leukemia with attenuated Salmonella typhimurium strain VNP20009. Biomed. Pharm. 2020, 129, 110425. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.M.; Luo, X.; Feng, M.; Li, Z.; Le, T.; Ittensohn, M.; Trailsmith, M.; Bermudes, D.; Lin, S.L.; King, I.C. Tumor amplified protein expression therapy: Salmonella as a tumor-selective protein delivery vector. Oncol. Res. 2000, 12, 127–135. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, J.; Gu, M.; Li, G.; Ni, J.; Fan, M. Antitumor Effect of Cycle Inhibiting Factor Expression in Colon Cancer via Salmonella VNP20009. Anticancer Agents Med. Chem. 2020, 20, 1722–1727. [Google Scholar] [CrossRef]
- Cheng, X.; Zhang, X.; Zhou, Y.; Zhang, C.; Hua, Z.C. A Salmonella Typhimurium mutant strain capable of RNAi delivery: Higher tumor-targeting and lower toxicity. Cancer Biol. 2014, 15, 1068–1076. [Google Scholar] [CrossRef] [Green Version]
- Broadway, K.M.; Modise, T.; Jensen, R.V.; Scharf, B.E. Complete genome sequence of Salmonella enterica serovar Typhimurium VNP20009, a strain engineered for tumor targeting. J. Biotechnol. 2014, 192 Pt A, 177–178. [Google Scholar] [CrossRef]
- Liu, R.; Cao, Z.P.; Wang, L.; Wang, X.Y.; Lin, S.S.; Wu, F.; Pang, Y.; Liu, J.Y. Multimodal oncolytic bacteria by coating with tumor cell derived nanoshells. Nano Today 2022, 45, 101537. [Google Scholar] [CrossRef]
- Hoffman, R.M. Tumor-targeting amino acid auxotrophic Salmonella typhimurium. Amino Acids 2009, 37, 509–521. [Google Scholar] [CrossRef]
- Zhao, M.; Geller, J.; Ma, H.; Yang, M.; Penman, S.; Hoffman, R.M. Monotherapy with a tumor-targeting mutant of Salmonella typhimurium cures orthotopic metastatic mouse models of human prostate cancer. Proc. Natl. Acad. Sci. USA 2007, 104, 10170–10174. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, Y.; Miwa, S.; Zhang, Y.; Hiroshima, Y.; Yano, S.; Uehara, F.; Yamamoto, M.; Toneri, M.; Bouvet, M.; Matsubara, H.; et al. Efficacy of tumor-targeting Salmonella typhimurium A1-R on nude mouse models of metastatic and disseminated human ovarian cancer. J. Cell. Biochem. 2014, 115, 1996–2003. [Google Scholar] [PubMed]
- Miwa, S.; Yano, S.; Zhang, Y.; Matsumoto, Y.; Uehara, F.; Yamamoto, M.; Hiroshima, Y.; Kimura, H.; Hayashi, K.; Yamamoto, N.; et al. Tumor-targeting Salmonella typhimurium A1-R prevents experimental human breast cancer bone metastasis in nude mice. Oncotarget 2014, 5, 7119–7125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yano, S.; Zhang, Y.; Zhao, M.; Hiroshima, Y.; Miwa, S.; Uehara, F.; Kishimoto, H.; Tazawa, H.; Bouvet, M.; Fujiwara, T.; et al. Tumor-targeting Salmonella typhimurium A1-R decoys quiescent cancer cells to cycle as visualized by FUCCI imaging and become sensitive to chemotherapy. Cell Cycle 2014, 13, 3958–3963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, K.; Zhang, R.; Luo, H.; Zhang, J.; Tian, Z.; Zhang, X.; Zhang, Y.; Ali, M.K.; Kong, Q. Optimized Attenuated Salmonella Typhimurium Suppressed Tumor Growth and Improved Survival in Mice. Front. Microbiol. 2021, 12, 774490. [Google Scholar] [CrossRef]
- Fensterle, J.; Bergmann, B.; Yone, C.L.; Hotz, C.; Meyer, S.R.; Spreng, S.; Goebel, W.; Rapp, U.R.; Gentschev, I. Cancer immunotherapy based on recombinant Salmonella enterica serovar Typhimurium aroA strains secreting prostate-specific antigen and cholera toxin subunit B. Cancer Gene 2008, 15, 85–93. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Zhou, H.; Krueger, J.; Kaplan, C.; Lee, S.H.; Dolman, C.; Markowitz, D.; Wu, W.; Liu, C.; Reisfeld, R.A.; et al. Targeting tumor-associated macrophages as a novel strategy against breast cancer. J. Clin. Investig. 2006, 116, 2132–2141. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, M.R.; Danai, L.V.; Lewis, C.A.; Chan, S.H.; Gui, D.Y.; Kunchok, T.; Dennstedt, E.A.; Vander Heiden, M.G.; Muir, A. Quantification of microenvironmental metabolites in murine cancers reveals determinants of tumor nutrient availability. Elife 2019, 8, e44235. [Google Scholar] [CrossRef]
- Kaimala, S.; Al-Sbiei, A.; Cabral-Marques, O.; Fernandez-Cabezudo, M.J.; Al-Ramadi, B.K. Attenuated Bacteria as Immunotherapeutic Tools for Cancer Treatment. Front. Oncol. 2018, 8, 136. [Google Scholar] [CrossRef] [Green Version]
- Grille, S.; Moreno, M.; Bascuas, T.; Marques, J.M.; Munoz, N.; Lens, D.; Chabalgoity, J.A. Salmonella enterica serovar Typhimurium immunotherapy for B-cell lymphoma induces broad anti-tumour immunity with therapeutic effect. Immunology 2014, 143, 428–437. [Google Scholar] [CrossRef]
- Lee, C.H.; Hsieh, J.L.; Wu, C.L.; Hsu, H.C.; Shiau, A.L. B cells are required for tumor-targeting Salmonella in host. Appl. Microbiol. Biotechnol. 2011, 92, 1251–1260. [Google Scholar] [CrossRef]
- Raymond, B.; Young, J.C.; Pallett, M.; Endres, R.G.; Clements, A.; Frankel, G. Subversion of trafficking, apoptosis, and innate immunity by type III secretion system effectors. Trends Microbiol. 2013, 21, 430–441. [Google Scholar] [CrossRef] [PubMed]
- Saccheri, F.; Pozzi, C.; Avogadri, F.; Barozzi, S.; Faretta, M.; Fusi, P.; Rescigno, M. Bacteria-induced gap junctions in tumors favor antigen cross-presentation and antitumor immunity. Sci. Transl. Med. 2010, 2, 44ra57. [Google Scholar] [CrossRef] [PubMed]
- Hong, E.H.; Chang, S.Y.; Lee, B.R.; Pyun, A.R.; Kim, J.W.; Kweon, M.N.; Ko, H.J. Intratumoral injection of attenuated Salmonella vaccine can induce tumor microenvironmental shift from immune suppressive to immunogenic. Vaccine 2013, 31, 1377–1384. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Chopra, A.K. An enteric pathogen Salmonella enterica serovar Typhimurium suppresses tumor growth by downregulating CD44high and CD4T regulatory (Treg) cell expression in mice: The critical role of lipopolysaccharide and Braun lipoprotein in modulating tumor growth. Cancer Gene 2010, 17, 97–108. [Google Scholar] [CrossRef]
- Kuan, Y.D.; Lee, C.H. Salmonella overcomes tumor immune tolerance by inhibition of tumor indoleamine 2, 3-dioxygenase 1 expression. Oncotarget 2016, 7, 374–385. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, P.C.; Quiceno, D.G.; Zabaleta, J.; Ortiz, B.; Zea, A.H.; Piazuelo, M.B.; Delgado, A.; Correa, P.; Brayer, J.; Sotomayor, E.M.; et al. Arginase I production in the tumor microenvironment by mature myeloid cells inhibits T-cell receptor expression and antigen-specific T-cell responses. Cancer Res. 2004, 64, 5839–5849. [Google Scholar] [CrossRef] [Green Version]
- Renner, C.; Held, G.; Ohnesorge, S.; Bauer, S.; Gerlach, K.; Pfitzenmeier, J.P.; Pfreundschuh, M. Role of perforin, granzymes and the proliferative state of the target cells in apoptosis and necrosis mediated by bispecific-antibody-activated cytotoxic T cells. Cancer Immunol. Immunother. 1997, 44, 70–76. [Google Scholar] [CrossRef]
- Martinez, F.O.; Sica, A.; Mantovani, A.; Locati, M. Macrophage activation and polarization. Front. Biosci. 2008, 13, 453–461. [Google Scholar] [CrossRef] [Green Version]
- Ganai, S.; Arenas, R.B.; Sauer, J.P.; Bentley, B.; Forbes, N.S. In tumors Salmonella migrate away from vasculature toward the transition zone and induce apoptosis. Cancer Gene 2011, 18, 457–466. [Google Scholar] [CrossRef] [Green Version]
- Chang, W.W.; Lee, C.H. Salmonella as an innovative therapeutic antitumor agent. Int. J. Mol. Sci. 2014, 15, 14546–14554. [Google Scholar] [CrossRef] [Green Version]
- Uchugonova, A.; Zhao, M.; Zhang, Y.; Weinigel, M.; Konig, K.; Hoffman, R.M. Cancer-cell killing by engineered Salmonella imaged by multiphoton tomography in live mice. Anticancer Res. 2012, 32, 4331–4337. [Google Scholar] [PubMed]
- Viallard, C.; Larrivee, B. Tumor angiogenesis and vascular normalization: Alternative therapeutic targets. Angiogenesis 2017, 20, 409–426. [Google Scholar] [CrossRef] [PubMed]
- Pavlova, N.N.; Zhu, J.; Thompson, C.B. The hallmarks of cancer metabolism: Still emerging. Cell Metab. 2022, 34, 355–377. [Google Scholar] [CrossRef] [PubMed]
- Forbes, N.S.; Munn, L.L.; Fukumura, D.; Jain, R.K. Sparse initial entrapment of systemically injected Salmonella typhimurium leads to heterogeneous accumulation within tumors. Cancer Res. 2003, 63, 5188–5193. [Google Scholar]
- Zhao, M.; Yang, M.; Li, X.M.; Jiang, P.; Baranov, E.; Li, S.; Xu, M.; Penman, S.; Hoffman, R.M. Tumor-targeting bacterial therapy with amino acid auxotrophs of GFP-expressing Salmonella typhimurium. Proc. Natl. Acad. Sci. USA 2005, 102, 755–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toley, B.J.; Forbes, N.S. Motility is critical for effective distribution and accumulation of bacteria in tumor tissue. Integr. Biol. 2012, 4, 165–176. [Google Scholar] [CrossRef] [Green Version]
- Westphal, K.; Leschner, S.; Jablonska, J.; Loessner, H.; Weiss, S. Containment of tumor-colonizing bacteria by host neutrophils. Cancer Res. 2008, 68, 2952–2960. [Google Scholar] [CrossRef] [Green Version]
- Mi, Z.; Feng, Z.C.; Li, C.; Yang, X.; Ma, M.T.; Rong, P.F. Salmonella-Mediated Cancer Therapy: An Innovative Therapeutic Strategy. J. Cancer 2019, 10, 4765–4776. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.H.; Lin, S.T.; Liu, J.J.; Chang, W.W.; Hsieh, J.L.; Wang, W.K. Salmonella induce autophagy in melanoma by the downregulation of AKT/mTOR pathway. Gene 2014, 21, 309–316. [Google Scholar] [CrossRef]
- Tsao, Y.T.; Kuo, C.Y.; Cheng, S.P.; Lee, C.H. Downregulations of AKT/mTOR Signaling Pathway for Salmonella-Mediated Suppression of Matrix Metalloproteinases-9 Expression in Mouse Tumor Models. Int. J. Mol. Sci. 2018, 19, 1630. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.K.; Chen, M.C.; Leong, H.F.; Kuo, Y.L.; Kuo, C.Y.; Lee, C.H. Connexin 43 suppresses tumor angiogenesis by down-regulation of vascular endothelial growth factor via hypoxic-induced factor-1alpha. Int. J. Mol. Sci. 2014, 16, 439–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiyuna, T.; Tome, Y.; Uehara, F.; Murakami, T.; Zhang, Y.; Zhao, M.; Kanaya, F.; Hoffman, R.M. Tumor-targeting Salmonella typhimurium A1-R Inhibits Osteosarcoma Angiogenesis in the In Vivo Gelfoam(R) Assay Visualized by Color-coded Imaging. Anticancer. Res. 2018, 38, 159–164. [Google Scholar] [PubMed]
- Galdiero, M.R.; Bonavita, E.; Barajon, I.; Garlanda, C.; Mantovani, A.; Jaillon, S. Tumor associated macrophages and neutrophils in cancer. Immunobiology 2013, 218, 1402–1410. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol. 2004, 25, 677–686. [Google Scholar] [CrossRef]
- Josefowicz, S.Z.; Lu, L.F.; Rudensky, A.Y. Regulatory T cells: Mechanisms of differentiation and function. Annu. Rev. Immunol. 2012, 30, 531–564. [Google Scholar] [CrossRef]
- Varela-Vazquez, A.; Guitian-Caamano, A.; Carpintero-Fernandez, P.; Fonseca, E.; Sayedyahossein, S.; Aasen, T.; Penuela, S.; Mayan, M.D. Emerging functions and clinical prospects of connexins and pannexins in melanoma. Biochim. Biophys. Acta Rev. Cancer 2020, 1874, 188380. [Google Scholar] [CrossRef]
- DeNardo, D.G.; Ruffell, B. Macrophages as regulators of tumour immunity and immunotherapy. Nat. Rev. Immunol. 2019, 19, 369–382. [Google Scholar] [CrossRef]
- Biswas, S.K.; Gangi, L.; Paul, S.; Schioppa, T.; Saccani, A.; Sironi, M.; Bottazzi, B.; Doni, A.; Vincenzo, B.; Pasqualini, F.; et al. A distinct and unique transcriptional program expressed by tumor-associated macrophages (defective NF-kappaB and enhanced IRF-3/STAT1 activation). Blood 2006, 107, 2112–2122. [Google Scholar] [CrossRef] [Green Version]
- Boutilier, A.J.; Elsawa, S.F. Macrophage Polarization States in the Tumor Microenvironment. Int. J. Mol. Sci. 2021, 22, 6995. [Google Scholar] [CrossRef]
- De Palma, M.; Mazzieri, R.; Politi, L.S.; Pucci, F.; Zonari, E.; Sitia, G.; Mazzoleni, S.; Moi, D.; Venneri, M.A.; Indraccolo, S.; et al. Tumor-targeted interferon-alpha delivery by Tie2-expressing monocytes inhibits tumor growth and metastasis. Cancer Cell 2008, 14, 299–311. [Google Scholar] [CrossRef] [Green Version]
- Duluc, D.; Corvaisier, M.; Blanchard, S.; Catala, L.; Descamps, P.; Gamelin, E.; Ponsoda, S.; Delneste, Y.; Hebbar, M.; Jeannin, P. Interferon-gamma reverses the immunosuppressive and protumoral properties and prevents the generation of human tumor-associated macrophages. Int. J. Cancer 2009, 125, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Pangilinan, C.R.; Wu, L.H.; Lee, C.H. Salmonella Impacts Tumor-Induced Macrophage Polarization, and Inhibits SNAI1-Mediated Metastasis in Melanoma. Cancers 2021, 13, 2894. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Xu, J.; Wang, Q.; Zhang, A.Q.; Wang, K. An obligatory anaerobic Salmonella typhimurium strain redirects M2 macrophages to the M1 phenotype. Oncol. Lett. 2018, 15, 3918–3922. [Google Scholar] [PubMed] [Green Version]
- Bernsmeier, C.; van der Merwe, S.; Perianin, A. Innate immune cells in cirrhosis. J. Hepatol. 2020, 73, 186–201. [Google Scholar] [CrossRef] [PubMed]
- Bruns, H.; Buttner, M.; Fabri, M.; Mougiakakos, D.; Bittenbring, J.T.; Hoffmann, M.H.; Beier, F.; Pasemann, S.; Jitschin, R.; Hofmann, A.D.; et al. Vitamin D-dependent induction of cathelicidin in human macrophages results in cytotoxicity against high-grade B cell lymphoma. Sci. Transl. Med. 2015, 7, 282ra47. [Google Scholar] [CrossRef]
- Lewen, S.; Zhou, H.; Hu, H.D.; Cheng, T.; Markowitz, D.; Reisfeld, R.A.; Xiang, R.; Luo, Y. A Legumain-based minigene vaccine targets the tumor stroma and suppresses breast cancer growth and angiogenesis. Cancer Immunol. Immunother. 2008, 57, 507–515. [Google Scholar] [CrossRef]
- Nguyen, V.H.; Kim, H.S.; Ha, J.M.; Hong, Y.; Choy, H.E.; Min, J.J. Genetically engineered Salmonella typhimurium as an imageable therapeutic probe for cancer. Cancer Res. 2010, 70, 18–23. [Google Scholar] [CrossRef] [Green Version]
- Sorenson, B.S.; Banton, K.L.; Frykman, N.L.; Leonard, A.S.; Saltzman, D.A. Attenuated Salmonella typhimurium with IL-2 gene reduces pulmonary metastases in murine osteosarcoma. Clin. Orthop. Relat. Res. 2008, 466, 1285–1291. [Google Scholar] [CrossRef] [Green Version]
- Deng, W.; Marshall, N.C.; Rowland, J.L.; McCoy, J.M.; Worrall, L.J.; Santos, A.S.; Strynadka, N.C.J.; Finlay, B.B. Assembly, structure, function and regulation of type III secretion systems. Nat. Rev. Microbiol. 2017, 15, 323–337. [Google Scholar] [CrossRef]
- Galan, J.E.; Wolf-Watz, H. Protein delivery into eukaryotic cells by type III secretion machines. Nature 2006, 444, 567–573. [Google Scholar] [CrossRef]
- Rajashekar, R.; Liebl, D.; Seitz, A.; Hensel, M. Dynamic remodeling of the endosomal system during formation of Salmonella-induced filaments by intracellular Salmonella enterica. Traffic 2008, 9, 2100–2116. [Google Scholar] [CrossRef] [PubMed]
- Liss, V.; Swart, A.L.; Kehl, A.; Hermanns, N.; Zhang, Y.; Chikkaballi, D.; Bohles, N.; Deiwick, J.; Hensel, M. Salmonella enterica Remodels the Host Cell Endosomal System for Efficient Intravacuolar Nutrition. Cell Host Microbe 2017, 21, 390–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mesa-Pereira, B.; Medina, C.; Camacho, E.M.; Flores, A.; Santero, E. Novel tools to analyze the function of Salmonella effectors show that SvpB ectopic expression induces cell cycle arrest in tumor cells. PLoS ONE 2013, 8, e78458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mercado-Lubo, R.; Zhang, Y.; Zhao, L.; Rossi, K.; Wu, X.; Zou, Y.; Castillo, A.; Leonard, J.; Bortell, R.; Greiner, D.L.; et al. A Salmonella nanoparticle mimic overcomes multidrug resistance in tumours. Nat. Commun. 2016, 7, 12225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siccardi, D.; Mumy, K.L.; Wall, D.M.; Bien, J.D.; McCormick, B.A. Salmonella enterica serovar Typhimurium modulates P-glycoprotein in the intestinal epithelium. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G1392–G1400. [Google Scholar] [CrossRef] [Green Version]
- Sundaram, B.; Kanneganti, T.D. Advances in Understanding Activation and Function of the NLRC4 Inflammasome. Int. J. Mol. Sci. 2021, 22, 1048. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Z.; Ruan, J.; Pan, Y.; Magupalli, V.G.; Wu, H.; Lieberman, J. Inflammasome-activated gasdermin D causes pyroptosis by forming membrane pores. Nature 2016, 535, 153–158. [Google Scholar] [CrossRef] [Green Version]
- Christgen, S.; Zheng, M.; Kesavardhana, S.; Karki, R.; Malireddi, R.K.S.; Banoth, B.; Place, D.E.; Briard, B.; Sharma, B.R.; Tuladhar, S.; et al. Identification of the PANoptosome: A Molecular Platform Triggering Pyroptosis, Apoptosis, and Necroptosis (PANoptosis). Front. Cell Infect. Microbiol. 2020, 10, 237. [Google Scholar] [CrossRef]
- Nishikawa, H.; Sato, E.; Briones, G.; Chen, L.M.; Matsuo, M.; Nagata, Y.; Ritter, G.; Jager, E.; Nomura, H.; Kondo, S.; et al. In vivo antigen delivery by a Salmonella typhimurium type III secretion system for therapeutic cancer vaccines. J. Clin. Investig. 2006, 116, 1946–1954. [Google Scholar] [CrossRef] [Green Version]
- Roider, E.; Jellbauer, S.; Kohn, B.; Berchtold, C.; Partilla, M.; Busch, D.H.; Russmann, H.; Panthel, K. Invasion and destruction of a murine fibrosarcoma by Salmonella-induced effector CD8 T cells as a therapeutic intervention against cancer. Cancer Immunol. Immunother. 2011, 60, 371–380. [Google Scholar] [CrossRef]
- Xiong, G.; Husseiny, M.I.; Song, L.; Erdreich-Epstein, A.; Shackleford, G.M.; Seeger, R.C.; Jackel, D.; Hensel, M.; Metelitsa, L.S. Novel cancer vaccine based on genes of Salmonella pathogenicity island 2. Int. J. Cancer 2010, 126, 2622–2634. [Google Scholar]
- Panthel, K.; Meinel, K.M.; Sevil Domenech, V.E.; Trulzsch, K.; Russmann, H. Salmonella type III-mediated heterologous antigen delivery: A versatile oral vaccination strategy to induce cellular immunity against infectious agents and tumors. Int. J. Med. Microbiol. 2008, 298, 99–103. [Google Scholar] [CrossRef]
- Jia, L.J.; Wei, D.P.; Sun, Q.M.; Jin, G.H.; Li, S.F.; Huang, Y.; Hua, Z.C. Tumor-targeting Salmonella typhimurium improves cyclophosphamide chemotherapy at maximum tolerated dose and low-dose metronomic regimens in a murine melanoma model. Int. J. Cancer 2007, 121, 666–674. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; He, J.; Cheng, H.; Zhu, Z.; Xu, H. Enhanced therapeutic effect of an antiangiogenesis peptide on lung cancer in vivo combined with salmonella VNP20009 carrying a Sox2 shRNA construct. J. Exp. Clin. Cancer Res. 2016, 35, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiroshima, Y.; Zhang, Y.; Murakami, T.; Maawy, A.; Miwa, S.; Yamamoto, M.; Yano, S.; Sato, S.; Momiyama, M.; Mori, R.; et al. Efficacy of tumor-targeting Salmonella typhimurium A1-R in combination with anti-angiogenesis therapy on a pancreatic cancer patient-derived orthotopic xenograft (PDOX) and cell line mouse models. Oncotarget 2014, 5, 12346–12357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Platt, J.; Sodi, S.; Kelley, M.; Rockwell, S.; Bermudes, D.; Low, K.B.; Pawelek, J. Antitumour effects of genetically engineered Salmonella in combination with radiation. Eur. J. Cancer 2000, 36, 2397–2402. [Google Scholar] [CrossRef]
- Liu, X.; Jiang, S.; Piao, L.; Yuan, F. Radiotherapy combined with an engineered Salmonella typhimurium inhibits tumor growth in a mouse model of colon cancer. Exp. Anim. 2016, 65, 413–418. [Google Scholar] [CrossRef] [Green Version]
- Yoon, W.S.; Kim, S.; Seo, S.; Park, Y. Salmonella typhimurium with gamma-radiation induced H2AX phosphorylation and apoptosis in melanoma. Biosci. Biotechnol. Biochem. 2014, 78, 1082–1085. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Miwa, S.; Zhang, N.; Hoffman, R.M.; Zhao, M. Tumor-targeting Salmonella typhimurium A1-R arrests growth of breast-cancer brain metastasis. Oncotarget 2015, 6, 2615–2622. [Google Scholar] [CrossRef] [Green Version]
- Dresselaers, T.; Theys, J.; Nuyts, S.; Wouters, B.; de Bruijn, E.; Anne, J.; Lambin, P.; Van Hecke, P.; Landuyt, W. Non-invasive 19F MR spectroscopy of 5-fluorocytosine to 5-fluorouracil conversion by recombinant Salmonella in tumours. Br. J. Cancer 2003, 89, 1796–1801. [Google Scholar] [CrossRef] [Green Version]
- Roland, K.L.; Brenneman, K.E. Salmonella as a vaccine delivery vehicle. Expert Rev. Vaccines 2013, 12, 1033–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chorobik, P.; Czaplicki, D.; Ossysek, K.; Bereta, J. Salmonella and cancer: From pathogens to therapeutics. Acta Biochim. Pol. 2013, 60, 285–297. [Google Scholar] [CrossRef] [Green Version]
- Bolhassani, A.; Zahedifard, F. Therapeutic live vaccines as a potential anticancer strategy. Int. J. Cancer 2012, 131, 1733–1743. [Google Scholar] [CrossRef] [PubMed]
- Medina, E.; Guzman, C.A.; Staendner, L.H.; Colombo, M.P.; Paglia, P. Salmonella vaccine carrier strains: Effective delivery system to trigger anti-tumor immunity by oral route. Eur. J. Immunol. 1999, 29, 693–699. [Google Scholar] [CrossRef]
- Xu, X.; Hegazy, W.A.; Guo, L.; Gao, X.; Courtney, A.N.; Kurbanov, S.; Liu, D.; Tian, G.; Manuel, E.R.; Diamond, D.J.; et al. Effective cancer vaccine platform based on attenuated salmonella and a type III secretion system. Cancer Res. 2014, 74, 6260–6270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Zhou, P.; Cai, J.; Yang, G.; Liang, S.; Ren, D. Tumor antigen delivered by Salmonella III secretion protein fused with heat shock protein 70 induces protection and eradication against murine melanoma. Cancer Sci. 2010, 101, 2621–2628. [Google Scholar] [CrossRef]
- Stegantseva, M.V.; Shinkevich, V.A.; Tumar, E.M.; Meleshko, A.N. Multi-antigen DNA vaccine delivered by polyethylenimine and Salmonella enterica in neuroblastoma mouse model. Cancer Immunol. Immunother. 2020, 69, 2613–2622. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.C.; Ding, J.; Pan, B.R.; Yu, Z.C.; Han, Q.L.; Meng, F.P.; Liu, N.; Fan, D.M. Development of an oral DNA vaccine against MG7-Ag of gastric cancer using attenuated salmonella typhimurium as carrier. World J. Gastroenterol. 2003, 9, 1191–1195. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Bai, T.; Tian, Y.; Zhou, B.; Wang, Y.; Yang, L. H2O2-Inactivated Salmonella typhimurium RE88 Strain as a New Cancer Vaccine Carrier: Evaluation in a Mouse Model of Cancer. Drug Des. Devel. 2021, 15, 209–222. [Google Scholar] [CrossRef]
- Revaz, V.; Benyacoub, J.; Kast, W.M.; Schiller, J.T.; De Grandi, P.; Nardelli-Haefliger, D. Mucosal vaccination with a recombinant Salmonella typhimurium expressing human papillomavirus type 16 (HPV16) L1 virus-like particles (VLPs) or HPV16 VLPs purified from insect cells inhibits the growth of HPV16-expressing tumor cells in mice. Virology 2001, 279, 354–360. [Google Scholar] [CrossRef] [Green Version]
- Baud, D.; Ponci, F.; Bobst, M.; De Grandi, P.; Nardelli-Haefliger, D. Improved efficiency of a Salmonella-based vaccine against human papillomavirus type 16 virus-like particles achieved by using a codon-optimized version of L1. J. Virol. 2004, 78, 12901–12909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Echchannaoui, H.; Bianchi, M.; Baud, D.; Bobst, M.; Stehle, J.C.; Nardelli-Haefliger, D. Intravaginal immunization of mice with recombinant Salmonella enterica serovar Typhimurium expressing human papillomavirus type 16 antigens as a potential route of vaccination against cervical cancer. Infect. Immun. 2008, 76, 1940–1951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soghomonyan, S.A.; Doubrovin, M.; Pike, J.; Luo, X.; Ittensohn, M.; Runyan, J.D.; Balatoni, J.; Finn, R.; Tjuvajev, J.G.; Blasberg, R.; et al. Positron emission tomography (PET) imaging of tumor-localized Salmonella expressing HSV1-TK. Cancer Gene 2005, 12, 101–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panteli, J.T.; Forkus, B.A.; Van Dessel, N.; Forbes, N.S. Genetically modified bacteria as a tool to detect microscopic solid tumor masses with triggered release of a recombinant biomarker. Integr. Biol. (Camb) 2015, 7, 423–434. [Google Scholar] [CrossRef] [Green Version]
- Nemunaitis, J.; Cunningham, C.; Senzer, N.; Kuhn, J.; Cramm, J.; Litz, C.; Cavagnolo, R.; Cahill, A.; Clairmont, C.; Sznol, M. Pilot trial of genetically modified, attenuated Salmonella expressing the E. coli cytosine deaminase gene in refractory cancer patients. Cancer Gene 2003, 10, 737–744. [Google Scholar] [CrossRef]
- Schmitz-Winnenthal, F.H.; Hohmann, N.; Schmidt, T.; Podola, L.; Friedrich, T.; Lubenau, H.; Springer, M.; Wieckowski, S.; Breiner, K.M.; Mikus, G.; et al. A phase 1 trial extension to assess immunologic efficacy and safety of prime-boost vaccination with VXM01, an oral T cell vaccine against VEGFR2, in patients with advanced pancreatic cancer. Oncoimmunology 2018, 7, e1303584. [Google Scholar] [CrossRef] [Green Version]
- Heimann, D.M.; Rosenberg, S.A. Continuous intravenous administration of live genetically modified salmonella typhimurium in patients with metastatic melanoma. J. Immunother. 2003, 26, 179–180. [Google Scholar] [CrossRef] [Green Version]
- Leong, M.L.; Hampl, J.; Liu, W.; Mathur, S.; Bahjat, K.S.; Luckett, W.; Dubensky, T.W., Jr.; Brockstedt, D.G. Impact of preexisting vector-specific immunity on vaccine potency: Characterization of listeria monocytogenes-specific humoral and cellular immunity in humans and modeling studies using recombinant vaccines in mice. Infect. Immun. 2009, 77, 3958–3968. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Therapeutic strategy for Salmonella-mediated cancer therapy. Salmonella has been exploited to localize and regress tumors via the delivery of anti-cancer molecules to induce cytotoxicity. The intrinsic tumor-targeting feature of Salmonella has been hijacked to deliver the gene of interest or secondary metabolites to suppress the tumor. Recently, combination therapy with conventional treatment strategies is more effective in the treatment of various cancers. Another promising approach is the bio-imaging of the tumor by using a fluorescence marker that enhances the localization and helps in targeting the metastatic tumor.
Figure 1.
Therapeutic strategy for Salmonella-mediated cancer therapy. Salmonella has been exploited to localize and regress tumors via the delivery of anti-cancer molecules to induce cytotoxicity. The intrinsic tumor-targeting feature of Salmonella has been hijacked to deliver the gene of interest or secondary metabolites to suppress the tumor. Recently, combination therapy with conventional treatment strategies is more effective in the treatment of various cancers. Another promising approach is the bio-imaging of the tumor by using a fluorescence marker that enhances the localization and helps in targeting the metastatic tumor.
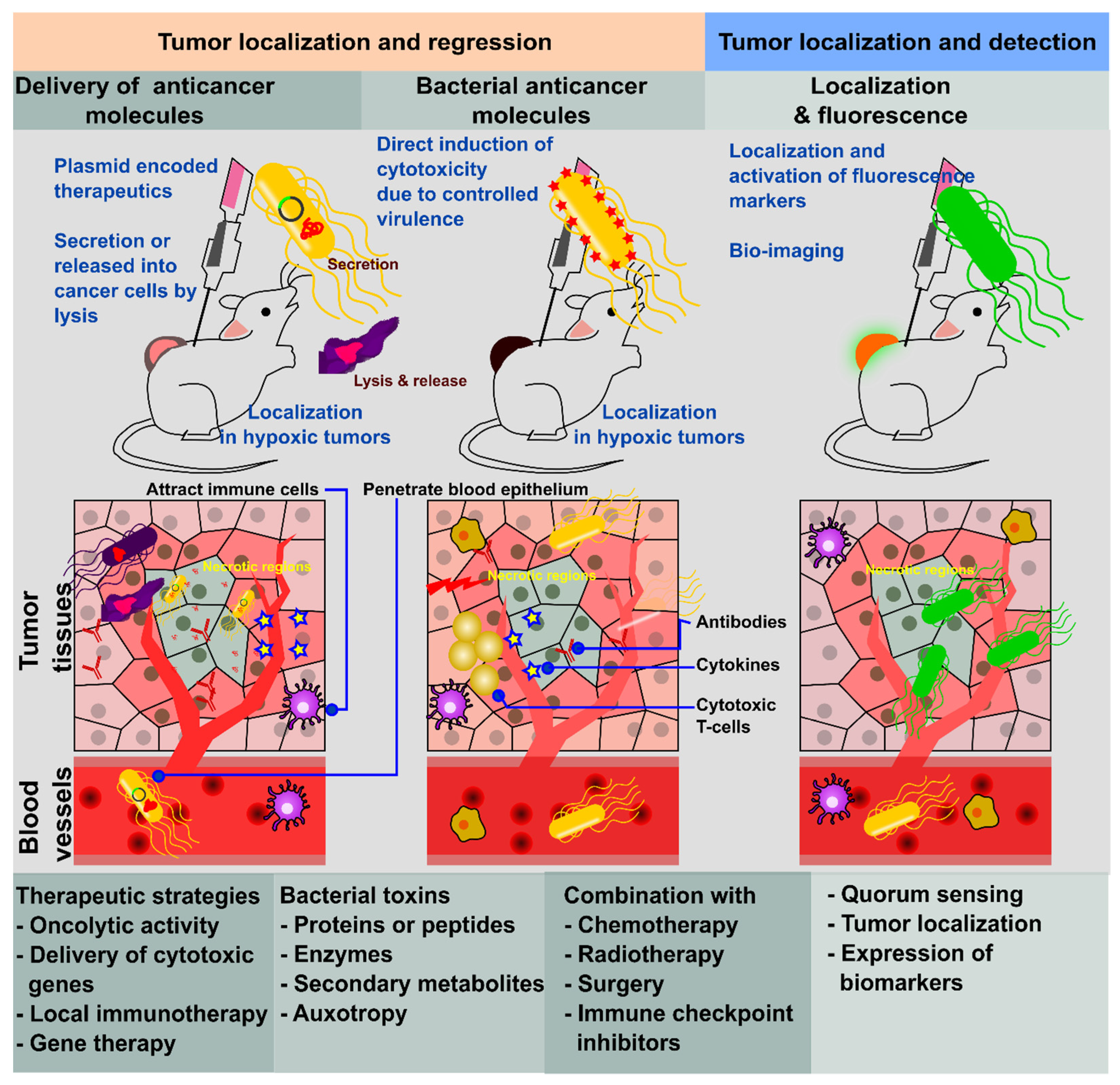
Figure 2.
Schematic diagram illustrating the mechanism of Salmonella to manipulate host immune response for tumor inhibition. Transformation of the tumor microenvironment from immunosuppressive to immunogenic occurs through increased infiltration and reprogramming of anti-tumor immune cells, upregulating the expression of proinflammatory cytokines and inducing a shift in the phenotypic and functional characteristics of immune cells. (1) A niche full of micronutrients supports the aggressive growth of Salmonella [87]. (2) Salmonella invasion increases the infiltration of macrophages [88], CD4+ T cells, CD8+ T cells, NK cells [89], and B cells [90] in the tumor microenvironment. (3) Bacterial invasion upregulates the expression of proinflammatory cytokines [91]. (4) Salmonella infection upregulates the Cx43 protein to enhance an antigenic presentation by dendritic cells [92]. (5) Activation of CD8+ T cells and NK cells downregulates Treg cells [93,94]. (6) Downregulation of immunosuppressive factors favors better immune protective response [95,96]. (7) Cytotoxic T cells release perforin to lyse the tumor cells [38,97]. (8) Transition of M2 macrophage to M1 macrophage orchestrates the protective anti-tumor immune response [88,98]. (9) Salmonella induces apoptosis and autophagy through its intrinsic properties [99,100,101]. (10) Chaotic vasculature is disrupted by inhibition of angiogenesis via downregulation of hypoxia-inducible factor-1 alpha (HIF-1α) and vascular endothelial growth factor (VEGF) [38]. All of the steps are summarized in the text. MΦ: macrophage; IL: interleukin, G-CSF: granulocyte colony-stimulating factor; GM-CSF: granulocyte-macrophage colony-stimulating factor; MIP-1α: macrophage inflammatory protein-1 alpha; ARG-1: arginase-1; TGF-β: transforming growth factor-beta; IDO: Indoleamine 2,3-dioxygenase; TAMs: tumor-associated macrophages; DC: dendritic cell; M1: M1-like macrophage; M2: M2-like macrophage.
Figure 2.
Schematic diagram illustrating the mechanism of Salmonella to manipulate host immune response for tumor inhibition. Transformation of the tumor microenvironment from immunosuppressive to immunogenic occurs through increased infiltration and reprogramming of anti-tumor immune cells, upregulating the expression of proinflammatory cytokines and inducing a shift in the phenotypic and functional characteristics of immune cells. (1) A niche full of micronutrients supports the aggressive growth of Salmonella [87]. (2) Salmonella invasion increases the infiltration of macrophages [88], CD4+ T cells, CD8+ T cells, NK cells [89], and B cells [90] in the tumor microenvironment. (3) Bacterial invasion upregulates the expression of proinflammatory cytokines [91]. (4) Salmonella infection upregulates the Cx43 protein to enhance an antigenic presentation by dendritic cells [92]. (5) Activation of CD8+ T cells and NK cells downregulates Treg cells [93,94]. (6) Downregulation of immunosuppressive factors favors better immune protective response [95,96]. (7) Cytotoxic T cells release perforin to lyse the tumor cells [38,97]. (8) Transition of M2 macrophage to M1 macrophage orchestrates the protective anti-tumor immune response [88,98]. (9) Salmonella induces apoptosis and autophagy through its intrinsic properties [99,100,101]. (10) Chaotic vasculature is disrupted by inhibition of angiogenesis via downregulation of hypoxia-inducible factor-1 alpha (HIF-1α) and vascular endothelial growth factor (VEGF) [38]. All of the steps are summarized in the text. MΦ: macrophage; IL: interleukin, G-CSF: granulocyte colony-stimulating factor; GM-CSF: granulocyte-macrophage colony-stimulating factor; MIP-1α: macrophage inflammatory protein-1 alpha; ARG-1: arginase-1; TGF-β: transforming growth factor-beta; IDO: Indoleamine 2,3-dioxygenase; TAMs: tumor-associated macrophages; DC: dendritic cell; M1: M1-like macrophage; M2: M2-like macrophage.
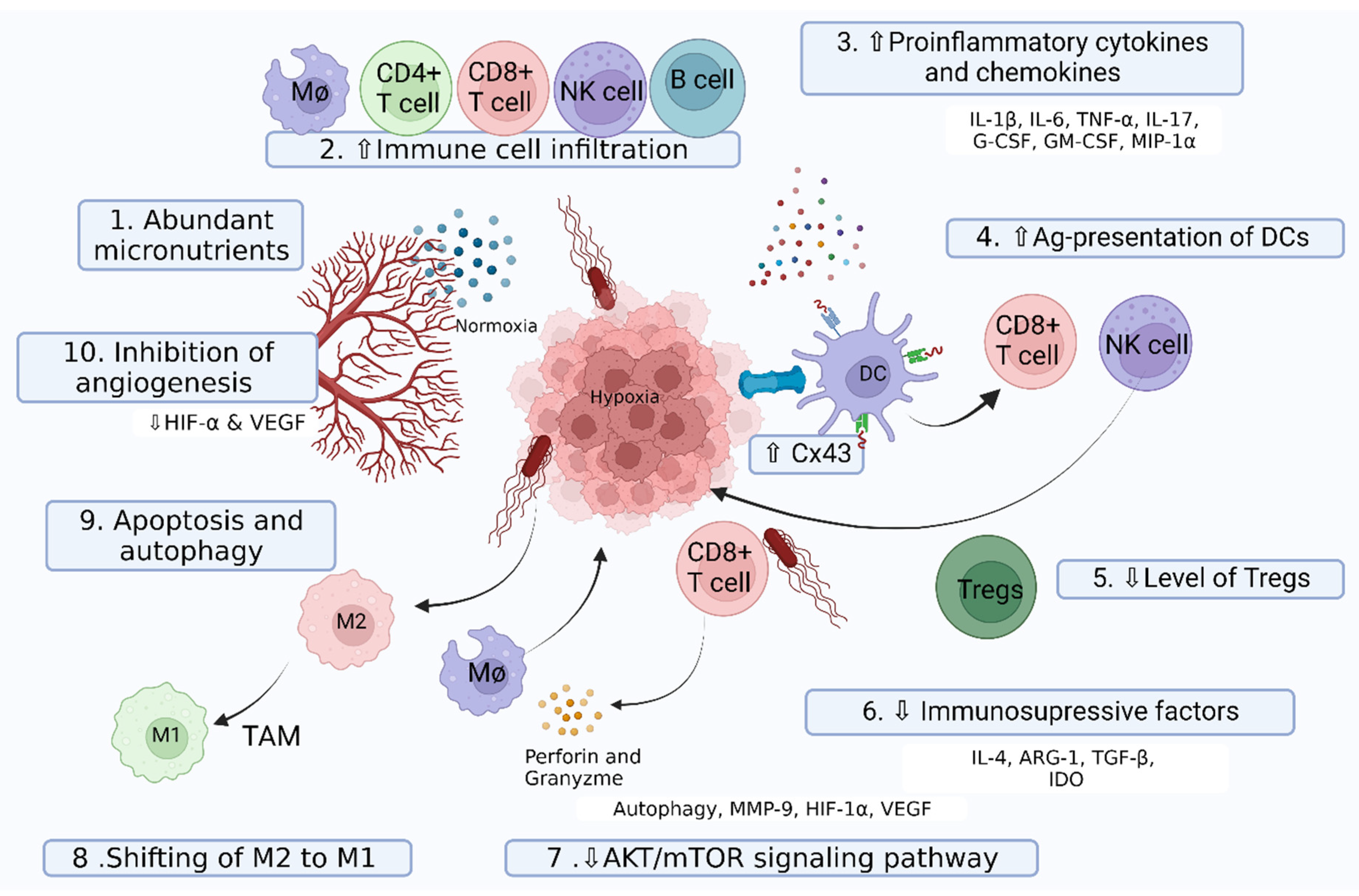

{kind=link}
{kind=link}
Table 1.
Salmonella strains in therapeutic application in cancer.
| Bacteria (Strain) | Strategy/Gene of Interest | Tumor Model | Results/Mode of Action | References |
|---|---|---|---|---|
| S. Typhimurium VNP20009 TAPET-CD | Cytosine deaminase expression from E. coli that converts non-toxic 5-fluorocytosine to the active anti-tumor agent 5-fluorouracil | Mice colon tumors | Mice treated with 5-FC inhibited tumor growth by 88−96% compared to TAPET-CD alone, which inhibited tumor growth by 38−79% | [55] |
| S. Typhimurium VNP20009 | Carboxypeptidase G2 (CPG2) | Murine models of breast and colon cancer | Prodrug-based suicide gene therapy | [56] |
| S. Typhimurium VNP20009 | msbB and purI mutations | melanomas | Attenuated strain preferentially accumulates in tumors and is rapidly cleared from other organs | [32] |
| S. Typhimurium | Deletion of ppGpp | CT26 tumor | Tumor suppression via IL-1β. | [50] |
| S. Typhimurium A1-R | leucine-arginine auxotroph | Pancreatic cancer orthotopic mouse model | Promotes CD8+ T cell infiltration and arrests tumor growth and metastasis. | [57] |
| S. Typhimurium A1-R | Combination with recombinant methioninase or doxorubin or temozolomide | Osteosarcoma, sarcoma, melanoma | Eradicate osteosarcoma and soft tissue sarcoma; regresses malignant melanoma | [58,59,60] |
| S. Typhimurium A1-R | Combination with temozolomide or vemurafenib | Melanoma in patient-derived orthotopic xenograft (PDOX) model | Combinatorial anti-tumor effect and drugs promoted targeting of S. Typhimurium A1-R | [61] |
| S. Typhimurium (ST2514P3) | Tryptophan auxotroph (trpA trpE deletion) | Breast cancer (4T1) | Suppressed the primary tumor growth and pulmonary metastasis | [13] |
| S. Typhimurium | Vascular endothelial growth factor receptor 2 | Lewis lung carcinoma | Tumor suppression and inhibition of pulmonary metastasis | [62] |
| S. Typhimurium VNP20009 | TNF-related apoptosis-inducing ligand (TRAIL) | Mammary tumor, melanoma | Caspase-3-mediated apoptosis in cancer cells | [54,63] |
| S. Typhimurium | Fas ligand (FasL) | breast carcinoma and CT-26 colon carcinoma cells | Tumor growth inhibition by 59% for breast tumors and 82% for colon carcinoma | [53] |
| S. Typhimurium | shRNA- expressing vectors targeting bcl2 | Melanoma | Delayed tumor growth and prolonged survival | [64] |
| S. Typhimurium | Cytotoxic protein (HlyE) | Mammary tumor | Increased tumor necrosis and reduced tumor growth | [65] |
| S. Typhimurium msbB- and purI- | IL-18 | Lewis lung carcinoma | Inhibit the growth of primary subcutaneous tumors and pulmonary metastases | [66] |
| S. Typhimurium | Herpes simplex virus thymidine kinase (HSV TK) | Melanoma | Suppressed tumor growth and a prolonged average survival | [15] |
Table 2.
List of Salmonella strains under human clinical trials for cancer therapy.
| Bacteria | Strain | Cancer Model | Result | Phase | References |
|---|---|---|---|---|---|
| S. Typhimurium | VNP20009 | Metastatic melanoma and renal cell carcinoma | Focal tumor colonization was recorded without tumor regression | I | [33,167] |
| S. Typhimurium | VNP20009 expressing TAPET-CD (Cytosine deaminase) | Squamous cell carcinoma and adenocarcinoma | Intratumoral bacterial colonization in 2 out of 3 patients | I | [165] |
| S. Typhimurium | VNP20009 | Solid tumors | Not provided | I | https://clinicaltrials.gov/ct2/show/NCT00006254 (accessed on 1 September 2022) |
| Salmonella typhi (Express VEGFR2) | Ty21a (VXM01) | Pancreatic cancer | VXM01 vaccination increased vaccine-specific T cell responses | I | [166] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Aganja, R.P.; Sivasankar, C.; Senevirathne, A.; Lee, J.H. Salmonella as a Promising Curative Tool against Cancer. Pharmaceutics 2022, 14, 2100. https://doi.org/10.3390/pharmaceutics14102100
AMA Style
Aganja RP, Sivasankar C, Senevirathne A, Lee JH. Salmonella as a Promising Curative Tool against Cancer. Pharmaceutics. 2022; 14(10):2100. https://doi.org/10.3390/pharmaceutics14102100
Chicago/Turabian StyleAganja, Ram Prasad, Chandran Sivasankar, Amal Senevirathne, and John Hwa Lee. 2022. "Salmonella as a Promising Curative Tool against Cancer" Pharmaceutics 14, no. 10: 2100. https://doi.org/10.3390/pharmaceutics14102100
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.