Comparison of Root Surface Enzyme Activity of Ericaceous Plants and Picea abies Growing at the Tree Line in the Austrian Alps


1
Institute of Forest Ecology, Universität für Bodenkultur (BOKU), Peter Jordan Strasse 82, 1190 Vienna, Austria
2
CAS Key Laboratory of Mountain Ecological Restoration and Bioresource Utilization & Ecological Restoration and Biodiversity Conservation Key Laboratory of Sichuan Province, Chengdu Institute of Biology, Chinese Academy of Sciences, Chengdu 610041, China
3
Department of Ecology, Mongolian University of Life Science, Zaisan 17024, Mailbox 57, Khan-Uul District, Ulaanbaator 17024, Mongolia
4
School of Forestry, Northeast Forestry University, Harbin 150040, China
5
Global Change Research Institute, Academy of Sciences of the Czech Republic, Department of Landscape Carbon Deposition, Na Sádkách 7, 37005 Ceské Budejovice, Czech Republic
*
Author to whom correspondence should be addressed.
Forests 2018, 9(9), 575; https://doi.org/10.3390/f9090575
Submission received: 31 July 2018
/
Revised: 3 September 2018
/
Accepted: 11 September 2018
/
Published: 17 September 2018
(This article belongs to the Special Issue The Ecology of Fine Roots and Mycorrhizas in Forests)
Abstract
:Plants with ericoid mycorrhizal and ectomycorrhizal associations coexist at the tree line and in many boreal forests. Both ericoid mycorrhizal and ectomycorrhizal roots are known to produce extracellular enzymes, but ericoid mycorrhizal fungi have been demonstrated in vitro to have higher enzyme activities. On hair roots of four ericoid mycorrhizal species (Rhododendron ferrugineum, Vaccinium vitis-idaea, Vaccinium myrtillus, Calluna vulgaris) and on ectomycorrhizal and non-mycorrhizal root tips of Picea abies growing at the tree line (1700 m) in the Austrian Alps, potential activities of eight extracellular root enzymes were estimated. Our results show that the activities of all the different extracellular root enzymes were generally similar among the ericaceous plant species. The mean laccase enzyme activity of ectomycorrhizal root tips of Picea abies was significantly higher than that of both the hair roots and fine roots of the ericaceous species. Leucine-amino-peptidase activity on hair roots was significantly higher than on non-mycorrhizal fine roots for the ericaceous vegetation. However, the mean activity of β-glucuronidase of the ericaceous species was significantly higher in fine roots compared to the hair roots. Generally extracellular root enzyme activity is not higher on ericaceous roots compared to ectomycorrhizas of Picea abies.
1. Introduction
Tree lines are climatically-determined ecotones [1,2], and in the Alps form a transition zone from tree dominated vegetation to ericaceous plants and grass dominated vegetation forms [2]. A number of ericaceous species co-exist at and above the tree line [2]. The tree line zone is also a transition between areas dominated by different mycorrhizal types [3]. The tree species, mostly Picea abies, Pinus mugo, and Larix decidua form ectomycorrhizas while the ericaceous plants form ericoid mycorrhizas. Species such as Rhododendron ferrugineum, Vaccinium myrtillus, and Vaccinium vitis-idaea are the dominant ericaceous plant species at the tree line [4], and often grow in discrete dense patches. Studies conducted at high latitude tree lines in Alaska showed that nitrogen (N) and phosphorus (P) availability is generally low in Alaskan spruce forest as release of N and P from dead organic matter is slow due to temperature limited decomposition [5]. Nitrogen limitation has been suggested to be a major factor restricting plant growth at the alpine tree line [6,7]. A characteristic of ericaceous plants is that they have low litter amounts of N and P, and a function attributed to ericoid mycorrhizas is the ability to mobilize N and P from these low-quality litter substrates using extracellular enzymes [8]. Many studies have demonstrated that ectomycorrhizas of trees also play a major role in the mineral nutrition of the trees through a variety of mechanisms, one of them also being the exudation of extracellular enzymes to degrade organic substrates to release, particularly, nitrogen and phosphorus [9]. Thus, under low N and P conditions at the tree line, the relative extracellular enzyme activities may be a factor effecting N and P acquisition of both trees and ericaceous plants and may limit the spread of trees into the dense ericaceous patches. However, to the best of our knowledge, the surface extracellular enzyme activity of ericoid mycorrhizas has never been determined, presumably because of the thinness of the hair roots and associated problems of obtaining an accurate measurement of root surface area.
In contrast to the lack of investigations on extracellular enzyme activity of ericoid roots mycorrhizas, many investigations of enzyme activity have been conducted on pure cultures under in vitro conditions [10,11]. Haselwandter et al. [12] provided evidence that selected ectomycorrhizal and ericoid fungi have the potential to degrade lignin in axenic culture. The early work of Read and co-workers demonstrated the presence of proteases and phosphatases, whereas later work has shown that these fungi also produce extracellular enzymes capable of degrading components of the plant cell wall such as β-l,4-endoxylanase [13], laccase, and phenol oxidase [14]. In all cases, ericoid fungi have been shown to have high to very high levels of activity of these enzymes [13,15,16,17].
Measuring potential enzymatic activities on root tips using the multi-enzyme assays developed by Pritsch et al. [18] and Courty et al. [9] makes a contribution to estimating the ecological function of ectomycorrhizas [19]. These enzymes play a role in carbon (C), N, or P degradation or mobilization (Table S1). The enzymes β-glucosidase, cellobiohydrolase, β-xylosidase, and β-glucuronidase are all involved in the degradation of cellulose [20,21]. Other commonly assayed enzymes are involved in the degradation of more recalcitrant C-containing substances or release of N. For example, laccase is involved in degradation of lignin [22]. N-acetyl-β-d-glucosaminidase is involved in degradation of chitin, and leucine aminopeptidase in cleavage of polypeptides [23]. Acid phosphatases are able to free phosphate groups from complex organic compounds and are widespread in living organisms.
In the present study, our aims were to compare the activity profiles of ectomycorrhizal root tips and ericoid mycorrhizal hair roots, to detect differences in root surface enzyme activities among different ericaceous species, and to detect differences in root surface enzyme activity of ericaceous species between hair roots and fine roots. The ericoid mycorrhizal roots of ericaceous plants are often referred to as ‘hair roots’, usually having a distal diameter of <100 µm [15]. Several studies showed that the mycorrhizal colonization levels of hair roots of ericaceous plants are up to 99% [24,25]. The hair roots branch from thicker fine roots that contain few or no fungal tissues [26]. We tested the two hypothesis: (i) ericoid mycorrhizal hair roots of ericaceous species have higher root surface enzyme activity than ectomycorrhizal root tips of co-occurring Picea abies trees, and (ii) the activities of all the different extracellular root enzymes were generally similar among the ericaceous plant species. To this end, we measured root surface enzyme activity of four different ericaceous species (Rhododendron ferrugineum, Vaccinium vitis-idaea, Vaccinium myrtillus, and Calluna vulgaris) and the tree species Picea abies that co-exist at the alpine tree line. The eight enzyme assays were determined using photometric and fluorometric techniques, and a method was developed to measure the projected surface area of the fine and hair roots using ImageJ software (National Institute of Mental Health, Bethesda, MA, USA) in order to compare the enzyme activity of tree roots and the hair roots.
2. Materials and Methods
2.1. Site Description
The study site is located in the Wasserberg area of the Stift Heiligenkreuz forest estate near Gaal in the central Alps in Austria. At the tree line, five replicate plots were established between 1668 and 1791 m above sea level on a southeast facing slope (47°19′ N, 14°43′ E) [27]. Each plot contained discrete areas dominated by Rhododendron ferrugineum, Vaccinium vitis-idaea, Vaccinium myrtillus, or Calluna vulgaris. The plots were ca. 120 m apart. In addition, five replicated plots were established in a diffuse tree line of Picea abies at an elevation of 1800 m, ca. 300 to 500 m from the plots of ericaceous plants. The trees were estimated to be 90–120 years old.
Soils at the sites are developed from gneiss and are Dystric Cambisols with an H (Layers dominated by organic material, formed from accumulations of undecomposed or partially decomposed organic material at the soil surface which may be underwater) layer of ca. 6 cm and an A (Layers were made of highly decomposed organic matter) layer of ca. 10 cm.
2.2. Root Sampling
On 8 June 2016, one soil core was taken to a depth of 20 cm from the center of the bushes of each of the ericaceous species Rhododendron ferrugineum, Vaccinium vitis-idaea, Vaccinium myrtillus, and Calluna vulgaris on each of the five plots. Five soil cores were taken from each vegetation species in total. From the closed Picea abies stand at the diffuse tree line site, two soil cores per plot were taken using a 7 cm diameter stainless steel corer. In total, 30 soil cores were collected at the tree line sites (Figure S1). Soil samples were directly returned to the laboratory and stored at 4 °C until further analysis within two days. Enzyme activity does not change within two days in intact soil cores [28]. Root segments were taken from each sample of the ericaceous species and carefully cleaned to remove adhering soil particles using a brush and tap water. A 5–7 cm segment of the fine roots from each species was placed under a ZEISS (Stemi 2000-CS, Carl Zeiss, Oberkochen, Germany) dissecting microscope that was connected with an AxioCam ERc5s camera (Carl Zeiss, Oberkochen, Germany). From this segment, a 1–3 cm long root section with either (1) many hair roots or (2) no hair roots (Figure S2) was removed. For each root type, hair roots and fine roots (without hair roots), replicate samples were removed for enzyme analysis. Three root sections of each root type from each ericaceous species at each plot were taken (6 root segments for each of 5 plots for hair roots and 3 root segments for each of 5 plots for fine roots from each ericaceous species). After estimation of enzyme activity, each sample was photographed under the microscope for estimation of projected surface area.
For Picea abies, root segments were removed from each soil core to give a sample with approximately 150–300 root tips per core. For the ectomycorrhizal root tips, all clearly definable ectomycorrhizal root tips from each sample were sorted into morphotypes based on the method described by Agerer [29], using a ZEISS (Stemi 2000-CS) dissecting microscope that was connected with an AxioCam ERc5s camera. In total, 10 clearly defined morphotypes were determined, and identification for ectomycorrhizal taxa was carried out by DNA extraction and PCR analysis (see below and Table S2).
2.3. DNA Extraction and PCR Amplification
The 1.5 mL micro-centrifuge tubes containing the ectomycorrhizal root tips of each morphotype were placed in liquid N for 5–10 min, and the tips were ground with a sterilized glass bar. DNA from the crushed ectomycorrhizal root tips was extracted by using DNeasy Plant Mini kits (QIAGEN, Hilden, Germany), and the extracted DNA was stored at −20 °C until the PCR reactions were run. For the PCR reactions, 1 µL DNA template was mixed with 12.5 µL MyTaq mix (BIOLINE, London, UK), 0.5 µL ITS1F (20 µM) primer (CTTGGTCATTTAGAGGAAGTAA forward), and 0.5 µL ITS4 (20 µM) primer (TCCTCCGCTTATTGATATGC reverse) and then diluted with 10.5 µL distilled deionized H2O. For the PCR, the Thermocyler (TProfessional Basic, Biometra, Germany) cycling parameters were an initial denaturation at 95 °C for 1 min, a second denaturation at 94 °C for 30 s, annealing at 50 °C for 40 s, and extension at 72 °C for 30 s, followed by a final auto-extension step at 72 °C for 4.5 min. The step from the second denaturation to extension was run for 35 cycles. To check the success of the PCR amplification, electrophoresis was carried out using 1% regular agarose gel stained with SERVA DNA Stain G in a 1% Tris-EDTA buffer solution. The gel was then visualized under UV light. If a clear single band was visible on the gel, the PCR products were sent for sequencing. Sequencing was done by Macrogen Inc., Seoul, Korea. Sequencing reactions were performed in a MJ Research PTC-225 Peltier Thermal Cycler using an ABI PRISM® BigDyeTM Terminator Cycle Sequencing Kit with AmpliTaq® DNA polymerase (FS enzyme) (Applied Biosystems, Foster City, CA, USA), following the protocols supplied by the manufacturer. Single-pass sequencing was performed on each template using an ITS4 primer. The fluorescent-labelled fragments were purified from the unincorporated terminators using the BigDye® XTerminator™ purification protocol. The samples were re-suspended in distilled water and subjected to electrophoresis in an ABI PRISM® 3730XL sequencer (Applied Biosystems, Foster City, CA, USA). The sequences obtained were manually checked and edited using Finch TV_1_4_0. Query sequences were compared with sequences on the UNITE and NBCI databases to identify the species of ectomycorrhiza; all but one of the sequences had a similarity of over 97% (Table S2). The sequences were deposited in GenBank with Accession No. MF784606-MF784613 and MF893218. Morphotypes could not be identified using the DNA analysis and were labelled as unknown.
2.4. Root Staining
Roots were sampled and cleaned with tap water. For checking the presence of ericoid mycorrhizas, roots were cut into 1 cm segments, soaked for 15 min in 10% KOH solution at 90 °C in an incubator, cleared in 3% H2O2 for 3 min, rinsed with distilled water 3 times, and then acidified for 5 min in 1% HCl at room temperature. The root segments were then stained with 0.05% trypan blue solution [30] (Figure S3). The stained roots were then mounted in acid glycerol on to glass microscope slides and examined under a ZEISS Axio Lab.A1 microscope (Carl Zeiss, Oberkochen, Germany).
2.5. Estimation of Hair Root and Fine Root Surface Area.
Digital images of the roots were made under a ZEISS (Stemi 2000-CS) microscope using an AxioCam ERc5s camera and were used to calculate the projected surface area of both hair roots and fine roots of ericaceous plants and ectomycorrhizas root tips of Picea abies. For each root section, a digital image was made of a calibration graticule. From the digital images, the projected surface area was determined using ImageJ software (http://imagej.nih.gov/ij/). The color threshold was set to give a high contrast black and white image. Once after the threshold was applied to the images, root area was quantified with the “Analyze particles” command. The root projected area was measured as the number of pixels belonging to the threshold region. It was based on the formula: area = (1/dpi)2, where dpi is the number of dots per inch used in scanning in mm. The ImageJ software offers direct calculation of area, with the correct scale setting. Using this method, the projected surface area of the roots less than 200 µm in diameter, and shown to contain mycorrhizal structures, could be calculated.
2.6. Potential Activity of Extracellular Root Enzymes
Using six replicates of hair root and three replicates of fine roots per species per plot, potential ectoenzyme activities were determined using the high-throughput photometric and fluorimetric 96-well black microplate assays described by Pritsch and Garbaye [31] and Courty et al. [9] (the analysis on the hair roots was repeated two times using separate root samples). For ectomycorrhizas and non-mycorrhizal roots tips, 3 replicates (3 single root tips) for each morphotype were used for enzyme analysis. In total, the enzyme activity of 10 different ectomycorrhizal taxa was determined (Table S3). For comparison to the extracellular enzyme activity of ericoid mycorrhizas, we used the mean values of the 10 ectomycorrhizal taxa.
Eight enzyme activities were measured: β-glucosidase (BG), N-acetyl-β-d-glucosaminidase (NAG), acid phosphatase (AP), leucine aminopeptidase (LAP), β-xylosidase (Xyl), cellobiohydrolase (Cel), β-Glucuronidase (Glr), and laccase. The substrates for the BG, NAG, AP, LAP, Xyl, Cel, and Glr assays were 4-methylumbelliferone (MUF)-β-d-glucopyranoside, MUF-N-actyl-β-d-glucosamine, MUF-phosphate, l-leucine-7-amido-4-methylcoumarin hydrochloride, MUF-xyloside, MUF-cellobioside, and MUF-glucuronide, respectively. For the determination of laccase activity, clear flat-bottom 96-well microfiltration plates (Sarstedt, Newton, NC, USA) were used, and the root tips were incubated in 100 µL of 2,2’-azinobis-3-ethylbenzothiazoline-6-sulfonate (ABTS; Sigma, St Quentin Fallavier, France) solution at 21 °C for 1 h. After incubation, 120 μL of incubation solution was added, and the intensity of the green color that had developed was immediately measured at 420 nm using an xMark™ Microplate Absorbance Spectrophotometer (Bio-Rad, Hercules, CA, USA) [27].
Laccase activity was expressed as μmol mm−2 min−1 of projected area of root tips. The rest of the enzymes activities were expressed as pmol mm−2 min−1 of projected area of root tips.
2.7. Soil Properties
The H-layer of the soil of the cores was sieved using a 2 mm mesh size sieve. Soil pH was determined on field moist soil using a 1:2 soil suspension in distilled water. For analysis of total C and total N, the sieved soil was then dried at 80 °C for 24 h. The dried soil was finely ground in a mortar, and C and N were determined in 100–150 mg samples using automated dry combustion (LECO TruSpec CN, LECO, St. Joseph, MI, USA).
2.8. Statistical Analysis
T-tests were used for comparison of the average enzymatic activity among different vegetation species, means and standard errors of soil parameters were calculated from each plot, the Student-Newman-Keuls test was used for Post-Hoc multiple comparison in SPSS. We refer to a p value of ≤0.05 as statistically significant and indicate in some cases a p value of ≤0.1 as marginally significant.
3. Results
3.1. Soil Chemical Properties
Table 1 shows that there were no significant differences in soil pH, percentage C and N, and the CN ratio among the four ericaceous vegetation species. The soil of the Picea abies stand had a lower pH and higher levels of soil C and N, and a higher CN ratio than those of the soil under the ericaceous plants.
3.2. Root Surface Enzyme Activity
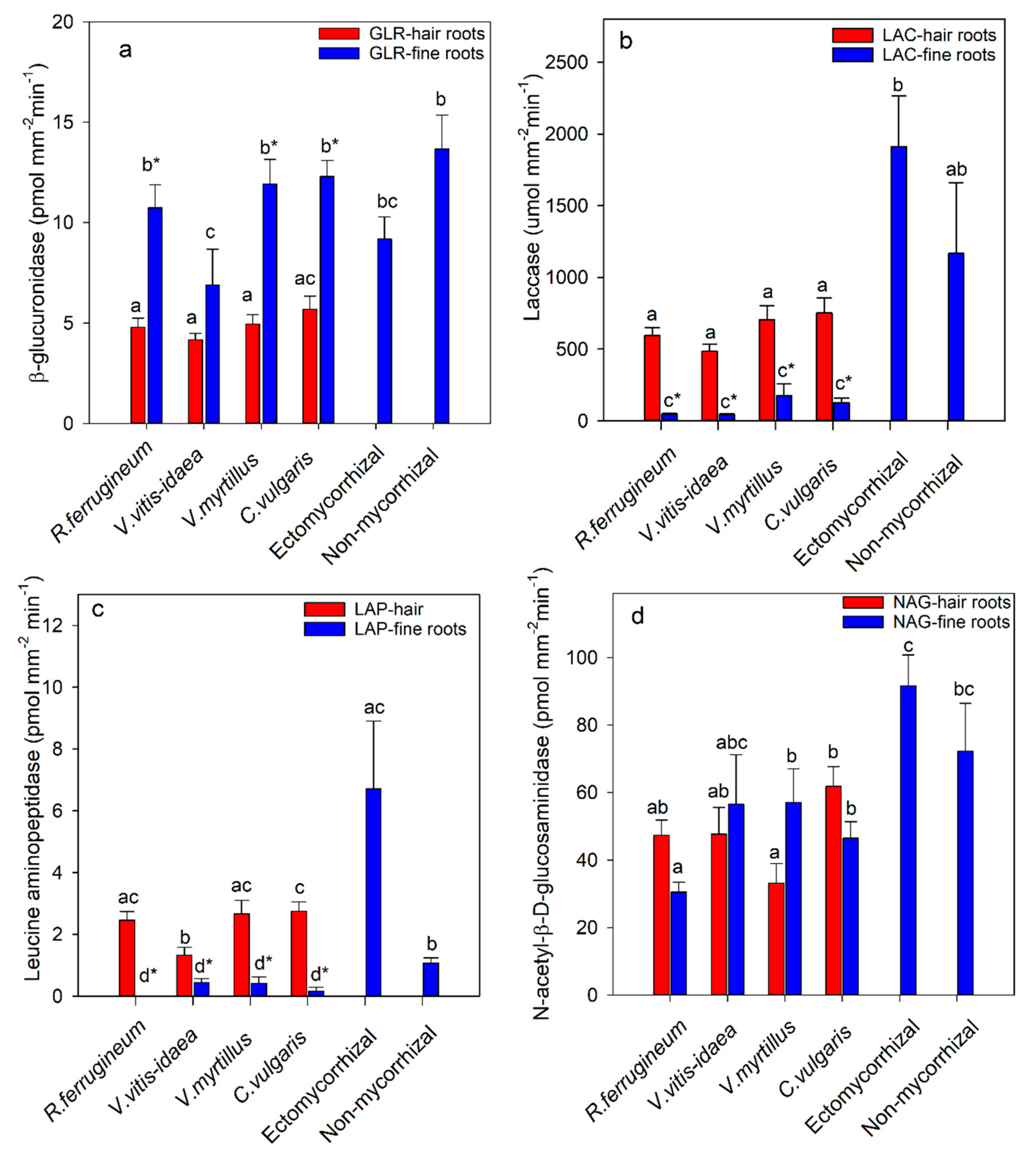
Of the eight enzymes measured, only β-glucuronidase, laccase, leucine aminopeptidase, and N-acetyl-β-d-glucosaminidase showed consistent significant differences. The mean activity of β-glucuronidase of the ericaceous species was significantly higher in fine roots compared to that of the hair roots (Figure 1a). The activity in fine roots of Vaccinium vitis-idaea was significantly lower than that of the other three species. The β-glucuronidase enzyme activity of ectomycorrhizal root tips of Picea abies was significantly higher than that of hair roots of the ericaceous species (Figure 1a). For all ericaceous species, the hair roots had significant higher laccase enzyme activity than that of fine roots. However, no significant differences were found in laccase enzyme activity among the ericaceous species (Figure 1b). The mean laccase enzyme activity of ectomycorrhizal root tips of Picea abies was significantly higher than that of both the hair roots and fine roots of the ericaceous species (Figure 1b). The activities of three ectomycorrhizal taxa were over five times that of the hair roots of the ericaceous species (Table 2). The potential activities of leucine aminopeptidase in hair roots of all ericaceous species were higher than those in fine roots (Figure 1c), and the activity in hair roots of Vaccinium vitis-idaea was significantly lower than those of the other three species. In the fine roots, no significant differences were shown among species, and the activities were very low in all species. The mean leucine aminopeptidase activity in ectomycorrhizal roots of Picea abies was significantly higher than that in both non-mycorrhizal roots tips and fine roots of the ericaceous species, but not the hair roots except those of Vaccinium vitis-idaea (Figure 1c). The high mean activity of the ectomycorrhizal root tips is due primarily to the very high activity of one of the taxa, Russula xerampelina (Table 2). This taxon had significantly higher leucine aminopeptidase activity than those of the other ectomycorrhizal taxa (Table 2). Hair roots of ericaceous species had similar activity of N-acetyl-β-d-glucosaminidase with exception of Calluna vulgaris which had significantly higher activity than that of Vaccinium myrtillus (Figure 1d). Mean enzyme activities of ectomycorrhizal root tips of Picea abies were significantly higher than that of the hair roots of all ericaceous species (Figure 1d). Although some individual taxa had similar activity to that of hair roots, the majority of the taxa had higher activity (Table 2). However, no significant differences were found in N-acetyl-β-d-glucosaminidase activity between hair roots and fine roots for all ericaceous species with exception of Vaccinium myrtillus (Figure 1d).
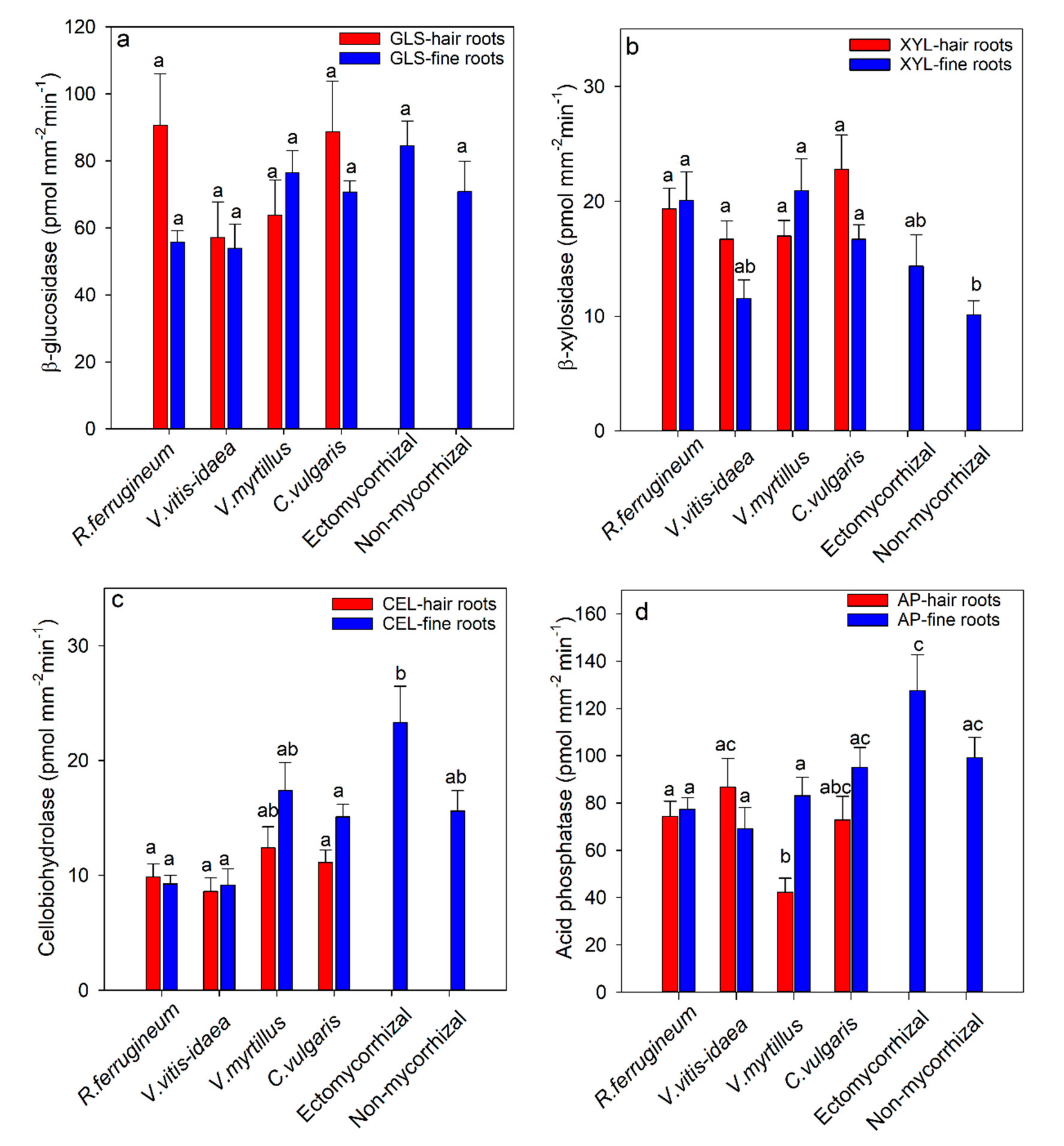
The enzyme activities of β-glucosidase, β-xylosidase, cellobiohydrolase, and acid phosphatase in the roots of the ericaceous species and ectomycorrhizas of Picea abies are shown in Figure 2. For β-glucosidase (Figure 2a), β-xylosidase (Figure 2b), and cellobiohydrolase (Figure 2c), there were no differences among ericaceous species both for hair roots and fine roots, and no significant differences between hair and fine roots (Figure 2). For cellobiohydrolase, the activity of ectomycorrhizal root tips was significantly higher than that of hair roots of all ericaceous species except Vaccinium myrtillus (Figure 2c). The majority of the ectmycorrhizal taxa had higher cellobiohydrolase activity compared to that of fine roots and hair roots of all ericaceous species (Table S3). For β-xylosidase, significantly lower activity was found in non-mycorrhizal root tips of Picea abies compared to that of hair roots of ericaceous species (Figure 2b). Figure 2d shows that acid phosphatase activity of Rhododendron ferrugineum in hair roots was significantly higher than that of the hair roots of Vaccinium myrtillus, and that acid phosphatase activity in ectomycorrhizal root tips of Picea abies was significant higher than that of the hair roots of Rhododendron ferrugineum and Vaccinium myrtillus. The high mean activity of the ectomycorrhizal root tips is due primarily to the very high activity of one of the taxa, Russula xerampelina (Table S3). There were no significant differences in acid phosphatase activity between fine roots and hair roots for all ericaceous species with the exception of Vaccinium myrtillus (Figure 2d).
4. Discussion
Roots tips of trees and their fungal symbionts have been shown to produce extracellular enzymes that degrade organic substrates [9]. There were clear differences in extracellular enzyme activity between the hair roots and fine roots of the ericaceous species, or the ectomycorrhizal roots and non-mycorrhizal of the Picea abies roots, suggesting differences in the production of enzymes between different tissues. This was particularly evident for laccase and leucine aminopeptidase. In fine roots of the ericaceous species we found no evidence of mycorrhizal colonization. Burke and Cairney [14] showed that laccase is produced by a number of ectomycorrhizal fungi and by the ericoid mycorrhizal fungus Rhizoscyphus ericae. Kanunfre and Zancan [32] showed that Thelephora terrestris, a hymenomycete that forms ectomycorrhizas with conifers, also produces laccase in pure culture. In contrast to laccase and leucine aminopeptidase, the similar activities of N-acetyl-β-d-glucosaminidase, acid phosphatases, β-glucosidase, cellobiohydrolase, and β-xylosidase between hair roots and fine roots, and also between ectomycorrhizas and non-mycorrhizal root tips of Picea abies, suggest that these are produced both by the plant and the ericoid mycorrhizal fungus. In in vitro culture, Burke and Cairney [13] showed that ericoid mycorrhizal fungus produced a complete cellulase enzyme complex (cellulase, cellobiohydrolase, and β-d-glucosidase) along with components of the hemicellulolytic complex (endoxylanase, β-d-xylosidase, α-d- and β-d-arabinosidase, and glucoronidase) and mannanolytic complex (mannanase, β-d-mannosidase, and α-d- and β-d-galactosidase). The ericoid mycorrhizal fungus Hymenoscyphus ericae produces high levels of acid phosphatase [33,34]. Higher activity of β-glucuronidase was found on the fine roots of the ericaceous species and non-mycorrhizal root tips of Picea abies. This result suggests that the β-glucuronidase enzyme is primarily produced by the plant. This conclusion confirmed results from Tedersoo et al. [35], which showed that most ectomycorrhizal fungi do not produce β-glucuronidase in significant amounts compared with that of non-ectomycorrhizal roots in a tropical rainforest ecosystem. We could not find any study that reported β-glucuronidase enzyme in ericoid mycorrhizas.
Among the four ericaceous plant species, there were few differences in activity of the root extracellular enzymes. One exception was the lower activity of leucine aminopeptidase on the hair roots of Vaccinium vitis-idaea, which was not reflected in the soil N status or the C/N ratio. The similarity of the root extracellular enzymes among the species and the similarity of the soil chemical parameters suggests that there are no major differences in nutrient mobilization from organic substrates among these co-existing species.
Using digital image analysis, we were able to obtain an estimate of the projected surface area of hair roots, which allowed comparison of the activity of extracellular enzymes between ericoid mycorrhiza and ectomycorrhiza in symbiosis. To the best of our knowledge, such comparisons until now have been restricted to in vitro studies in pure cultures [16,36]. The activities of both laccase and N-acetyl-β-d-glucosaminidase were significantly higher in ectomycorrhizal root tips than that of hair roots of ericoid species. This result is consistent with the conclusion of Adamczyk et al. [37], who found the highest N-acetyl-β-d-glucosaminidase enzyme activities in soil planted Pinus sylvestris, compared to that planted with ericoid mycorrhizal plants. However, this is increasing evidence to show that ectomycorrhizas are inhabited by a number of secondary colonizers such as ascomycete fungi [38] and bacteria [39], all of which can influence extracellular enzyme activity. Vohník et al. [40] also showed that senescing ectomycorrhizas of Picea abies have co-associated ascomycetes, some of which formed ericoid mycorrhizas on Vaccinium myrtillus. One of the identified ectomycorrhizal taxa on Picea abies at the tree line, an ascomycete Helotiales, had the highest laccase activity. Different species of Helotiales form ectomycorrhizas but are also common secondary colonizers of other ectomycorrhizas [38].
Generally, there were few differences in activity of the different enzymes among the species of ericaceous plants, and also between the ericaceous plants and Picea abies. As discussed above, the activities of laccase and N-acetyl-β-d-glucosaminidase were higher in ectomycorrhizal root tips than in ericoid mycorrhizal hair roots, and the activities of all other enzymes were similar between ectomycorrhizal root tips and ericoid mycorrhizal hair roots. Studies using in vitro pure cultures have generally shown that ericoid mycorrhizal fungi have high enzyme activity [16,41,42]. This higher activity has led to the suggestion that ericoid mycorrhizal fungi have a high capacity to mobilize N from recalcitrant compounds [4,16]. In our study, the higher enzyme activity of ectomycorrhizal root tips may be a function of the amount of fungal biomass directly in contact with the assay solution. For ectomycorrhizas, the assay solution is directly in contact with the high fungal surface area of the hyphal mantle, whereas in ericoid mycorrhizas, the majority of the fungal biomass is enclosed by the root tissue, but clearly active. This is in contrast to in vitro studies where similar amounts of fungal biomass are exposed to the assay solution. In contrast to the enzyme activity at the root surface in tree line soils, the activity of N-acetyl-β-d-glucosaminidase was similar in soil from under Rhododendron ferrugineum compared to Picea abies or Pinus mugo [27]. Thus, in the field, the high proliferation of ericoid roots [43] or dense root mats [4] compared to the more dispersed ectomycorrhizal roots may compensate for the lower surface area enzyme activity in the ericaceous roots.
5. Conclusions
Among the ericaceous plant species, the activities of all the different extracellular root enzymes were generally similar. Generally, extracellular root enzyme activity is not higher on ericaceous roots compared to the ectomycorrhizas of Picea abies.
Supplementary Materials
The following are available online at https://www.mdpi.com/1999-4907/9/9/575/s1, Table S1: Extracellular enzymes, their activities in soil, and the two fluorometrically labelled assays (4-methylumbelliferone-MUF and 7-amino-4-methyl coumarin-AMC) containing substrates used in the present analysis; Table S2: Identification of ectomycorrhizal taxa associated with Picea abies at tree line (1668 to 1791 m). Table S3: β-glucosidase, Acid phosphatase, Cellobiohydrolase, and β-xylosidase activity on root tips of 10 different ectomycorrhizal taxa of Picea abies at tree line (1668 to 1791 m). Figure S1: Images of the tree line site (1668 to 1791 m) at Wasserberg in the central Alps, Austria; Figure S2: Profile of ericoid mycorrhizal roots collected from the tree line (1668 to 1791 m) at Wasserberg in the central Alps, Austria in June 2016; Figure S3: Ericoid mycorrhizal roots structure.
Author Contributions
D.G. conceived and designed the experiments; L.W. and B.O. performed the experiments; L.W. analyzed the data; L.W. and D.G. wrote the paper. W.D. gave advice about micro climate.
Funding
L.W. was supported by the China scholarship council, and D.G. was supported by a Marie Curie grant GPF333996 LINKTOFUN. B.O. was supported by a Eurasia Pacific Uninet Fellowship and Erasmus-Mundus Post-Doc mobility grant. This work was also supported by the Ministry of Education, Youth and Sports of CR within the National Sustainability Program NPU I, grant No. LO1415.
Acknowledgments
We thank the Stift Heiligenkreuz for giving us access to the sites. We thank Frauke Neumann and Marcel Hirsch for technical support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Christian, K.; Jens, P. A world-wide study of high altitude treeline temperatures. J. Biogeogr. 2004, 31, 713–732. [Google Scholar] [CrossRef]
- Körner, C. A re-assessment of high elevation treeline positions and their explanation. Oecologia 1998, 115, 445–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Read, D.J. Mycorrhizas in ecosystems. Experientia 1991, 47, 376–391. [Google Scholar] [CrossRef]
- Read, D.J.; Leake, J.R.; Perez-Moreno, J. Mycorrhizal fungi as drivers of ecosystem processes in heathland and boreal forest biomes. Can. J. Bot. 2004, 82, 1243–1263. [Google Scholar] [CrossRef]
- Schulze, E.; Kelliher, F.M.; Korner, C.; Lloyd, J.; Leuning, R. Relationships among Maximum Stomatal Conductance, Ecosystem Surface Conductance, Carbon Assimilation Rate, and Plant Nitrogen Nutrition: A Global Ecology Scaling Exercise. Ann. Rev. Ecol. Syst. 1994, 25, 629–662. [Google Scholar] [CrossRef]
- Dawes, M.A.; Schleppi, P.; Hättenschwiler, S.; Rixen, C.; Hagedorn, F. Soil warming opens the nitrogen cycle at the alpine treeline. Glob. Chang. Biol. 2017, 23, 421–434. [Google Scholar] [CrossRef] [PubMed]
- Thébault, A.; Clément, J.-C.; Ibanez, S.; Roy, J.; Geremia, R.A.; Pérez, C.A.; Buttler, A.; Estienne, Y.; Lavorel, S. Nitrogen limitation and microbial diversity at the treeline. Oikos 2014, 123, 729–740. [Google Scholar] [CrossRef] [Green Version]
- Cornelissen, J.; Aerts, R.; Cerabolini, B.; Werger, M.; van der Heijden, M. Carbon cycling traits of plant species are linked with mycorrhizal strategy. Oecologia 2001, 129, 611–619. [Google Scholar] [CrossRef] [PubMed]
- Courty, P.-E.; Karin, P.; Michael, S.; Anton, H.; Jean, G. Activity profiling of ectomycorrhiza communities in two forest soils using multiple enzymatic tests. New Phytol. 2005, 167, 309–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, R.M.; Cairney, J.W.G. Carbohydrate oxidases in ericoid and ectomycorrhizal fungi: A possible source of Fenton radicals during the degradation of lignocellulose. New Phytol. 1998, 139, 637–645. [Google Scholar] [CrossRef]
- Leake, J.R.; Read, D.J. Proteinase activity in mycorrhizal fungi. New Phytol. 1990, 116, 123–128. [Google Scholar] [CrossRef]
- Haselwandter, K.; Bobleter, O.; Read, D.J. Degradation of 14C-labelled lignin and dehydropolymer of coniferyl alcohol by ericoid and ectomycorrhizal fungi. Arch. Microbiol. 1990, 153, 352–354. [Google Scholar] [CrossRef]
- Burke, R.M.; Cairney, J.W.G. Carbohydrolase production by the ericoid mycorrhizal fungus Hymenoscyphus ericae under solid-state fermentation conditions. Mycol. Res. 1997, 101, 1135–1139. [Google Scholar] [CrossRef]
- Burke, R.; Cairney, J. Laccases and other polyphenol oxidases in ecto- and ericoid mycorrhizal fungi. Mycorrhiza 2002, 12, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Read, D.J. The Structure and Function of the Ericoid Mycorrhizal Root. Ann. Bot. 1996, 77, 365–374. [Google Scholar] [CrossRef] [Green Version]
- Bending, G.D.; Read, D.J. Nitrogen mobilization from protein-polyphenol complex by ericoid and ectomycorrhizal fungi. Soil Biol. Biochem. 1996, 28, 1603–1612. [Google Scholar] [CrossRef]
- Kerley, J.S.; Read, J.D. The biology of mycorrhiza in the Ericaceae. New Phytol. 1995, 131, 369–375. [Google Scholar] [CrossRef]
- Pritsch, K.; Raidl, S.; Marksteiner, E.; Blaschke, H.; Agerer, R.; Schloter, M.; Hartmann, A. A rapid and highly sensitive method for measuring enzyme activities in single mycorrhizal tips using 4-methylumbelliferone-labelled fluorogenic substrates in a microplate system. J. Microbiol. Methods 2004, 58, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Courty, P.-E.; Pouysegur, R.; Buée, M.; Garbaye, J. Laccase and phosphatase activities of the dominant ectomycorrhizal types in a lowland oak forest. Soil Biol. Biochem. 2006, 38, 1219–1222. [Google Scholar] [CrossRef]
- Henrissat, B.; Davies, G. Structural and sequence-based classification of glycoside hydrolases. Curr. Opin. Struct. Biol. 1997, 7, 637–644. [Google Scholar] [CrossRef]
- Kuroyama, H.; Tsutsui, N.; Hashimoto, Y.; Tsumuraya, Y. Purification and characterization of a β-glucuronidase from Aspergillus niger. Carbohydr. Res. 2001, 333, 27–39. [Google Scholar] [CrossRef]
- Thurston, C.F. The structure and function of fungal laccases. Microbiology 1994, 140, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Sinsabaugh, R.L.; Lauber, C.L.; Weintraub, M.N.; Ahmed, B.; Allison, S.D.; Crenshaw, C.; Contosta, A.R.; Cusack, D.; Frey, S.; Gallo, M.E. Stoichiometry of soil enzyme activity at global scale. Ecol. Lett. 2008, 11, 1252–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baba, T.; Hirose, D.; Sasaki, N.; Watanabe, N.; Kobayashi, N.; Kurashige, Y.; Karimi, F.; Ban, T. Mycorrhizal Formation and Diversity of Endophytic Fungi in Hair Roots of Vaccinium oldhamii Miq. in Japan. Microbes Environ. 2016, 31, 186–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grunewaldt-Stocker, G.; von den Berg, C.; Knopp, J.; von Alten, H. Interactions of ericoid mycorrhizal fungi and root pathogens in Rhododendron: In vitro tests with plantlets in sterile liquid culture. Plant Root 2013, 7, 33–48. [Google Scholar] [CrossRef] [Green Version]
- Perotto, S.; Martino, E.; Abbà, S.; Vallino, M. 14 Genetic Diversity and Functional Aspects of Ericoid Mycorrhizal Fungi. In Fungal Associations; Hock, B., Ed.; Springer: Berlin, Germany, 2012; pp. 255–285. [Google Scholar]
- Wang, L.X.; Otgonsuren, B.; Godbold, D.L. Mycorrhizas and soil ecosystem function of co-existing woody vegetation islands at the alpine tree line. Plant Soil 2017, 411, 467–481. [Google Scholar] [CrossRef] [PubMed]
- Mosca, E.; Montecchio, L.; Scattolin, L.; Garbaye, J. Enzymatic activities of three ectomycorrhizal types of Quercus robur L. in relation to tree decline and thinning. Soil Biol. Biochem. 2007, 39, 2897–2904. [Google Scholar] [CrossRef]
- Agerer, R. Colour Atlas of Ectomycorrhizae; Einhorn-Verlag: Schwabisch Gmund, Germany, 1997. [Google Scholar]
- Brundrett, M.; Bougher, N.; Dell, B.; Grove, T. Working Ylith Mycorrhizas in Forestry and Agriculture; Australian Centre for International Agricultural Research: Canberra, Australia, 1996. [Google Scholar]
- Pritsch, K.; Garbaye, J. Enzyme secretion by ECM fungi and exploitation of mineral nutrients from soil organic matter. Ann. For. Sci. 2011, 68, 25–32. [Google Scholar] [CrossRef] [Green Version]
- Kanunfre, C.C.; Zancan, G.T. Physiology of exolaccase production by Thelephora terrestris. FEMS Microbiol. Lett. 1998, 161, 151–156. [Google Scholar] [CrossRef] [Green Version]
- Pearson, V.; Read, D.J. The physiology of the mycorrhizal endophyte of Calluna vulgaris. Trans. Br. Mycol. Soc. 1975, 64, 1–7. [Google Scholar] [CrossRef]
- Straker, C.J.; Mitchell, D.T. The activity and characterization of acid phosphatase in endomycorrhizal fungi of the ericaceae. New Phytol. 1986, 104, 243–256. [Google Scholar] [CrossRef]
- Tedersoo, L.; Naadel, T.; Bahram, M.; Pritsch, K.; Buegger, F.; Leal, M.; Kõljalg, U.; Põldmaa, K. Enzymatic activities and stable isotope patterns of ectomycorrhizal fungi in relation to phylogeny and exploration types in an afrotropical rain forest. New Phytol. 2012, 195, 832–843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leake, J.; Read, D. Proteinase activity in mycorrhizal fungi III. New Phytol. 1991, 117, 309–317. [Google Scholar] [CrossRef]
- Adamczyk, B.; Ahvenainen, A.; Sietiö, O.-M.; Kanerva, S.; Kieloaho, A.-J.; Smolander, A.; Kitunen, V.; Saranpää, P.; Laakso, T.; Straková, P.; et al. The contribution of ericoid plants to soil nitrogen chemistry and organic matter decomposition in boreal forest soil. Soil Biol. Biochem. 2016, 103, 394–404. [Google Scholar] [CrossRef]
- Tedersoo, L.; Kadri, P.; Teele, J.; Genevieve, G.; Kadri, P.; Heidi, T. Ascomycetes associated with ectomycorrhizas: Molecular diversity and ecology with particular reference to the Helotiales. Environ. Microbiol. 2009, 11, 3166–3178. [Google Scholar] [CrossRef] [PubMed]
- Burke, D.J.; Dunham, S.M.; Kretzer, A.M. Molecular analysis of bacterial communities associated with the roots of Douglas fir (Pseudotsuga menziesii) colonized by different ectomycorrhizal fungi. FEMS Microbiol. Ecol. 2008, 65, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Vohník, M.; Mrnka, L.; Lukešová, T.; Bruzone, M.C.; Kohout, P.; Fehrer, J. The cultivable endophytic community of Norway spruce ectomycorrhizas from microhabitats lacking ericaceous hosts is dominated by ericoid mycorrhizal Meliniomyces variabilis. Fungal Ecol. 2013, 6, 281–292. [Google Scholar] [CrossRef]
- Bending, G.D.; Read, D.J. Lignin and soluble phenolic degradation by ectomycorrhizal and ericoid mycorrhizal fungi. Mycol. Res. 1997, 101, 1348–1354. [Google Scholar] [CrossRef]
- Bending, G.D.; Read, D.J. Effects of the soluble polyphenol tannic acid on the activities of ericoid and ectomycorrhizal fungi. Soil Biol. Biochem. 1996, 28, 1595–1602. [Google Scholar] [CrossRef]
- Wurzburger, N.; Hendrick, R.L. Rhododendron thickets alter N cycling and soil extracellular enzyme activities in southern Appalachian hardwood forests. Pedobiologia 2007, 50, 563–576. [Google Scholar] [CrossRef]
Figure 1.
β-glucuronidase (a), laccase (b), leucine amino peptidase (c), and N-acetyl-β-d-glucosaminidase (d) activity of fine roots and hair roots of ericaceous species (Rhododendron ferrugineum, Vaccinium vitis-idaea, Vaccinium myrtillus, or Calluna vulgaris) and ectomycorrhizal and non-mycorrhizal root tips of Picea abies at tree line (1668 to 1791 m). The activity is calculated on the basis of projected surface area. The roots were collected at the Wasserberg, Gaal in the central Alps, Austria. Bars show means ± SE. Bars not followed by the same letters are significantly different (p ≤ 0.05) among ericaceous species, ectomycorrhizal species, or non-mycorrhizal roots tips (ab). Asterisks indicate significant differences between hair roots and fine roots for the same ericaceous species (p ≤ 0.05).
Figure 1.
β-glucuronidase (a), laccase (b), leucine amino peptidase (c), and N-acetyl-β-d-glucosaminidase (d) activity of fine roots and hair roots of ericaceous species (Rhododendron ferrugineum, Vaccinium vitis-idaea, Vaccinium myrtillus, or Calluna vulgaris) and ectomycorrhizal and non-mycorrhizal root tips of Picea abies at tree line (1668 to 1791 m). The activity is calculated on the basis of projected surface area. The roots were collected at the Wasserberg, Gaal in the central Alps, Austria. Bars show means ± SE. Bars not followed by the same letters are significantly different (p ≤ 0.05) among ericaceous species, ectomycorrhizal species, or non-mycorrhizal roots tips (ab). Asterisks indicate significant differences between hair roots and fine roots for the same ericaceous species (p ≤ 0.05).

Figure 2.
β-glucosidase (a), β-xylosidase (b), celliobiohydrolase (c), and acid phosphatase (d) activity of fine roots and hair roots of the ericaceous species (Rhododendron ferrugineum, Vaccinium vitis idaea, Vaccinium myrtillus, Calluna vulgaris) and ectomycorrhizal and non-mycorrhizal root tips of Picea abies at the tree line (1668 to 1791 m). The roots were collected at Wasserberg in the central Alps, Austria. Bars show means ± SE. Bars not followed by the same letters are significantly different (p ≤ 0.05) among ericaceous species, ectomycorrhizal species, or non-mycorrhizal roots tips (ab). There were no significant differences between hair roots and fine roots for the same ericaceous species (p ≤ 0.05).
Figure 2.
β-glucosidase (a), β-xylosidase (b), celliobiohydrolase (c), and acid phosphatase (d) activity of fine roots and hair roots of the ericaceous species (Rhododendron ferrugineum, Vaccinium vitis idaea, Vaccinium myrtillus, Calluna vulgaris) and ectomycorrhizal and non-mycorrhizal root tips of Picea abies at the tree line (1668 to 1791 m). The roots were collected at Wasserberg in the central Alps, Austria. Bars show means ± SE. Bars not followed by the same letters are significantly different (p ≤ 0.05) among ericaceous species, ectomycorrhizal species, or non-mycorrhizal roots tips (ab). There were no significant differences between hair roots and fine roots for the same ericaceous species (p ≤ 0.05).


{kind=link}
{kind=link}
Table 1.
Chemical properties of soils taken from under Rhododendron ferrugineum, Vaccinium vitis-idaea, Vaccinium myrtillus, and Calluna vulgaris at the tree line (1668 to 1791 m) and from a neighboring Picea abies diffuse tree line at Wasserberg in the central Alps, Austria. Mean ± SE. Data points within a row not followed by the same letter are significantly different (p ≤ 0.05) between species.
Table 1.
Chemical properties of soils taken from under Rhododendron ferrugineum, Vaccinium vitis-idaea, Vaccinium myrtillus, and Calluna vulgaris at the tree line (1668 to 1791 m) and from a neighboring Picea abies diffuse tree line at Wasserberg in the central Alps, Austria. Mean ± SE. Data points within a row not followed by the same letter are significantly different (p ≤ 0.05) between species.
| Picea abies | Rhododendron ferrugineum | Vaccinium vitis-idaea | Vaccinium myrtillus | Calluna vulgaris | |
|---|---|---|---|---|---|
| Soil pH (water) | 4.3 ± 0.1 b | 4.8 ± 0.1 a | 4.9 ± 0.04 a | 4.8 ± 0.1 a | 4.7± 0.1 a |
| C% | 31.8 ± 3.1 b | 16.3 ± 1.1 a | 18.4 ± 2.5 a | 21.8 ± 5.6 a | 20.7 ± 3.3 a |
| N% | 1.2 ± 0.1 a | 0.7 ± 0.1 a | 0.9 ± 0.1 a | 0.9 ± 0.2 a | 1.0 ± 0.1 a |
| C/N | 26.4 ± 1.0 b | 22.2 ± 0.9 a | 20.1 ± 0. 8 a | 22.8 ± 1.1 a | 21.5 ± 0.5 a |
Table 2.
β-glucuronidase, laccase, leucine amino peptidase, and N-acetyl-β-d-glucosaminidase activity on root tips of 10 different ectomycorrhizal taxa of Picea abies at the tree line (1668 to 1791 m). The activity was calculated on the basis of projected surface area. The roots were collected at Wasserberg in the central Alps, Austria. Values show means ± SE. Data points within a column not followed by the same letter are significantly different (p ≤ 0.05) between taxa for each enzyme.
Table 2.
β-glucuronidase, laccase, leucine amino peptidase, and N-acetyl-β-d-glucosaminidase activity on root tips of 10 different ectomycorrhizal taxa of Picea abies at the tree line (1668 to 1791 m). The activity was calculated on the basis of projected surface area. The roots were collected at Wasserberg in the central Alps, Austria. Values show means ± SE. Data points within a column not followed by the same letter are significantly different (p ≤ 0.05) between taxa for each enzyme.
| Ectomycorrhizal Taxa | β-glucuronidase (pmol mm−2 min−1) | Laccase (µmol mm−2 min−1) | Leucine Aminopeptidase (pmol mm−2 min−1) | N-acetyl-β-d-glucosaminidase (pmol mm−2 min−1) |
|---|---|---|---|---|
| Thelephoraceae | 11.6 ± 2.1 bc | 4011 ± 891 b | 3.2 ± 1.0 b | 86 ± 12 bcd |
| Russula xerampelina | 2.8 ± 0.4 e | 4053 ± 756 b | 40.6 ± 6.7 a | 103 ± 6 ac |
| Lactarius aurantiacus | 3.8 ± 0.7 de | 253 ± 68 c | 3.9 ± 0.5 b | 107 ± 14 ab |
| Amphinema byssoides | 20.2 ± 3.6 a | 1246 ± 328 c | 4.5 ± 0.6 b | 91 ± 13 abcd |
| Unknown 1 | 4.1 ± 0.5 de | 493 ± 271 c | 3.3 ± 1.8 b | 85 ± 7 bcd |
| Unknown 2 | 10.2 ± 0.5 bcd | 889 ± 241 c | 3.4 ± 0.6 b | 32 ± 1 cd |
| Luellia sp. | 8.0 ± 1.7 bcde | 1407 ± 540 c | 1.9 ± 0.5 b | 106 ± 12 ab |
| Helotiales | 7.5 ± 2.0 cde | 5497 ± 530 a | 2.3 ± 0.2 b | 123 ± 10 ab |
| Leotiomycetes | 9.0 ± 2.0 bcde | 856 ± 211 c | 1. 5 ± 0.4 b | 22 ± 4 d |
| Tylospora sp. | 14.8 ± 3.8 ab | 424 ± 110 c | 2.6 ± 1.2 b | 162 ± 62 a |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, L.; Otgonsuren, B.; Duan, W.; Godbold, D. Comparison of Root Surface Enzyme Activity of Ericaceous Plants and Picea abies Growing at the Tree Line in the Austrian Alps. Forests 2018, 9, 575. https://doi.org/10.3390/f9090575
AMA Style
Wang L, Otgonsuren B, Duan W, Godbold D. Comparison of Root Surface Enzyme Activity of Ericaceous Plants and Picea abies Growing at the Tree Line in the Austrian Alps. Forests. 2018; 9(9):575. https://doi.org/10.3390/f9090575
Chicago/Turabian StyleWang, Lixia, Burenjargal Otgonsuren, Wenbiao Duan, and Douglas Godbold. 2018. "Comparison of Root Surface Enzyme Activity of Ericaceous Plants and Picea abies Growing at the Tree Line in the Austrian Alps" Forests 9, no. 9: 575. https://doi.org/10.3390/f9090575
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.