A Comparative Study of the Pathogenicity of Fusarium circinatum and other Fusarium Species in Polish Provenances of P. sylvestris L.


 ,
,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Fungal Isolates
2.2. In Vitro Virulence Test
2.3. Greenhouse Pathogenicity Tests
2.4. DNA Extraction, Purification and PCR Amplification
2.5. Statistical Analysis
3. Results
3.1. Effect of Fusarium Species on Pine Seedling
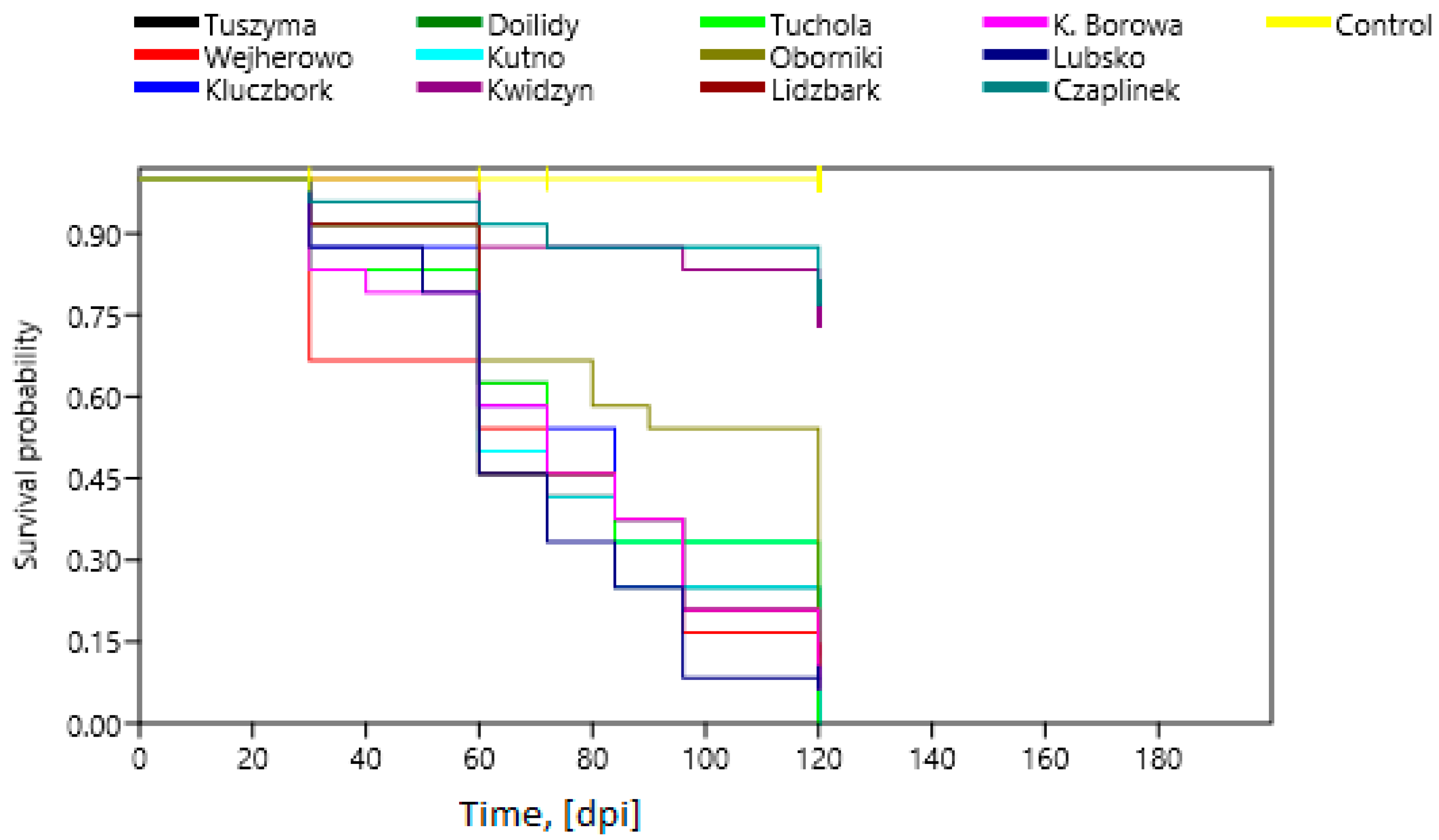
3.2. Greenhouse Pathogenicity Tests
4. Discussion
4.1. Preliminary Assessment of Pathogenicity of Fusarium Circinatum to Polish Pine Provenances
4.2. Virulence of Other Fusarium Species
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Santini, A.; Ghelardini, L.; De Pace, C.; Desprez-Loustau, M.L.; Capretti, P.; Chandelier, A.; Cech, T.; Chira, D.; Diamandis, S.; Gaitniekis, T.; et al. Biogeographical Patterns and Determinants of Invasion by Forest Pathogens in Europe. New Phytol. 2013, 197, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Ganley, R.J.; Watt, M.S.; Kriticos, D.J.; Hopkins, A.J.M.; Manning, L.K. Increased Risk of Pitch Canker to Australasia under Climate Change. Australas. Plant Pathol. 2011, 40, 228–237. [Google Scholar] [CrossRef]
- Watt, M.S.; Ganley, R.J.; Kriticos, D.J.; Manning, L.K. Dothistroma Needle Blight and Pitch Canker: The Current and Future Potential Distribution of Two Important Diseases of Pinus Species. Can. J. For. Res. 2011, 41, 412–424. [Google Scholar] [CrossRef]
- Gordon, T.R.; Kirkpatrick, S.C.; Aegerter, B.J.; Fisher, A.J.; Storer, A.J.; Wood, D.L. Evidence for the Occurrence of Induced Resistance to Pitch Canker, Caused by Gibberella circinate (Anamorph Fusarium circinatum), in Populations of Pinus radiata. For. Pathol. 2011, 41, 227–232. [Google Scholar] [CrossRef]
- Gordon, T.R. Pitch Canker Disease of Pines. Phytopathology 2006, 96, 657–659. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA). Risk Assessment of Gibberella circinata for the EU Territory and Identification and Evaluation of Risk Management Options. EFSA J. 2010, 8, 1620. [Google Scholar] [CrossRef]
- Bezos, D.; Martinez-Alvarez, P.; Fernández, M.; Diez, J.J. Epidemiology and Management of Pine Pitch Canker Disease in Europe—A Review. Balt. For. 2017, 23, 279–293. [Google Scholar]
- Landeras, E.; García, P.; Fernández, Y.; Braña, M.; Fernández-Alonso, O.; Méndez-Lodcs, S.; Pérez-Sierra, A.; León, M.; Abad-Campos, P.; Berbegal, M.; et al. Outbreak of Pitch Canker Caused by Fusarium circinatum on Pinus spp. in Northern Spain. Plant Dis. 2005, 89, 1015. [Google Scholar] [CrossRef]
- Bragança, H.; Diogo, E.; Moniz, F.; Amaro, P. First Report of Pitch Canker on Pines Caused by Fusarium circinatum in Portugal. Plant Dis. 2009, 93, 1079. [Google Scholar] [CrossRef]
- Wingfield, M.J.; Hammerbacher, A.; Ganley, R.J.; Steenkamp, E.T.; Gordon, T.R.; Wingfield, B.D.; Coutinho, T.A. Pitch Canker Caused by Fusarium circinatum—A Growing Threat to Pine Plantations and Forests Worldwide. Australas. Plant Pathol. 2008, 37, 319–334. [Google Scholar] [CrossRef]
- European and Mediterranean Plant Protection Organization (EPPO). PQR Database. Paris, France, 2014. Available online: http://www.eppo.int/DATABASES/pqr/pqr.htm (accessed on December 2016).
- Carlucci, A.; Colatruglio, L.; Frisullo, S. First Report of Pitch Canker Caused by Fusarium circinatum on Pinus halepensis and P. pinea in Apulia (Southern Italy). Plant Dis. 2007, 91, 1683. [Google Scholar]
- EPPO. Pm 7/91(1): Gibberella circinata. EPPO Bull, EPPO: Paris, France, 2009; 298–309.
- Chelkowski, J. (Ed.) Fusarium: Mycotoxins, Taxonomy, Pathogenicity; Elsevier: Amsterdam, The Netherlands, 1989; Volume 2, p. 492. [Google Scholar]
- Martín-Pinto, P.; Pajares, J.; Díez, J. Pathogenicity of Fusarium verticillioides and Fusarium oxysporum on Pinus nigra Seedlings in Northwest Spain. For. Pathol. 2008, 38, 78–82. [Google Scholar] [CrossRef]
- Anderson, I.C.; Campbell, C.D.; Prosser, J.I. Diversity of Fungi in Organic Soils under a Moorland–Scots Pine (Pinus sylvestris L.) Gradient. Environ. Microbiol. 2003, 5, 1121–1132. [Google Scholar] [CrossRef] [PubMed]
- Machón, P.; Pajares, J.A.; Diez, J.J.; Alves-Santos, F.M. Influence of the Ectomycorrhizal Fungus Laccaria laccata on Pre-Emergence, Post-Emergence and Late Damping-Off by Fusarium oxysporum and F. verticillioides on Stone Pine Seedlings. Symbiosis 2009, 49, 101–109. [Google Scholar] [CrossRef]
- Duda, B.; Sierota, Z.H. Survival of Scots Pine Seedlings after Biological and Chemical Control of Damping-off in Plastic Greenhouses. For. Pathol. 1987, 17, 110–117. [Google Scholar] [CrossRef]
- Bełka, M.; Mańka, M. Damping-off of Scots pine (Pinus sylvestris) Seedlings in 2004 and 2005 in Nowy Dwór Forest Nursery versus Soil Fungi Communities. Phytopathologia 2010, 58, 63–67. [Google Scholar]
- Martínez-Alvarez, P.; Pando, V.; Diez, J.J. Alternative Species to Replace Monterey Pine Plantations Affected by Pitch Canker Caused by Fusarium circinatum in Northern Spain. Plant Pathol. 2014, 63, 1086–1094. [Google Scholar] [CrossRef]
- James, R.L.; Dumroese, R.K.; Wenny, D.L. Fusarium proliferaturm is a Common, Aggressive Pathogen of Container-Grown Conifer Seedling. Phytopathology 1995, 85, 1129. [Google Scholar]
- Stewart, J.E.; Abdo, Z.; Dumroese, R.K.; Klopfenstein, N.B.; Kim, M.S. Virulence of Fusarium oxysporum and F. Commune to Douglas-Fir (Pseudotsuga menziesii) Seedlings. For. Pathol. 2012, 42, 220–228. [Google Scholar] [CrossRef]
- Schweigkofler, W.; O’Donnell, K.; Garbelotto, M. Detection and Quantification of Airborne Conidia of Fusarium circinatum, the Causal Agent of Pine Pitch Canker, from Two California Sites by Using a Real-Time PCR approach Combined with a Simple Spore Trapping Method. Appl. Environ. Microbiol. 2004, 70, 3512–3520. [Google Scholar] [CrossRef] [PubMed]
- Glass, N.L.; Donaldson, G.C. Development of Primer Sets Designed for Use with the PCR to Amplify Conserved Genes from Filamentous Ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [PubMed]
- Kristensen, R.; Torp, M.; Kosiak, B.; Holst-Jensen, A. Phylogeny and Toxigenic Potential is Correlated in Fusarium Species as Revealed by Partial Translation Elongation Factor 1 Alpha Gene Sequences. Mycol. Res. 2005, 109, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Gardes, M.; Bruns, T.D. Its Primers with Enhanced Specificity for Basidiomycetes—Application to the Identification of Mycorrhizae and Rusts. Mol. Ecol. 1995, 2, 113–118. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Method and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miiller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1997, 215, 403–410. [Google Scholar] [CrossRef]
- Therneau, T. A Package for Survival Analysis in S. R Package Version 2.41-3. Available online: https://CRAN.R-project.org/package=survival (accessed on 18 December 2017).
- Wingfield, M.J.; Jacobs, A.; Coutinho, T.A.; Ahumada, R.; Wingfield, B.D. First Report of the Pitch Canker Fungus, Fusarium circinatum, on Pines in Chile. Plant Pathol. 2002, 51, 397. [Google Scholar] [CrossRef]
- Hodge, G.R.; Dvorak, W.S. Variation in Pitch Canker Resistance Among Provenances of Pinus patula and Pinus tecunumanii from Mexico and Central America. New For. 2007, 33, 193–206. [Google Scholar] [CrossRef]
- Enebak, S.A.; Stanosz, G.R. Responses of Conifer Species of the Great Lakes Region of North America to Inoculation with the Pitch Canker Pathogen Fusarium circinatum. For. Pathol. 2003, 33, 333–338. [Google Scholar] [CrossRef]
- Iturritxa, E.; Ganley, R.J.; Raposo, R.; García-Serna, I.; Mesanza, N.; Kirkpatrick, S.C.; Gordon, T.R. Resistance Levels of Spanish Conifers Against Fusarium circinatum and Diplodia pinea. For. Pathol. 2013, 43, 488–495. [Google Scholar] [CrossRef]
- Pérez-Sierra, A.; Landeras, E.; León, M.; Berbegal, M.; García-Jiménez, J.; Armengol, J. Characterization of Fusarium circinatum from Pinus spp. in Northern Spain. Mycol. Res. 2007, 111, 832–839. [Google Scholar] [CrossRef] [PubMed]
- Martín-García, J.; Lukačevičová, A.; Flores-Pacheco, J.A.; Diez, J.J.; Dvořák, M. Evaluation of the Susceptibility of Several Czech Conifer Provenances to Fusarium circinatum. Forests 2018, 9, 72. [Google Scholar]
- Storer, A.J.; Gordon, T.R.; Clark, S.L. Association of the Pitch Canker Fungus, Fusarium Subglutinans f. sp. pini with Monterey Pine Seeds and Seedlings in California. Plant Pathol. 1998, 47, 649–656. [Google Scholar] [CrossRef]
- Schmale, D.G.; Gordon, T.R. Variation in Susceptibility to Pitch Canker Disease, caused by Fusarium circinatum, in Native Stands of Pinus muricata. Plant Pathol. 2003, 52, 720–725. [Google Scholar] [CrossRef]
- Swett, C.L.; Kirkpatrick, S.C.; Gordon, T.R. Evidence for a Hemibiotrophic Association of the Pitch Canker Pathogen Fusarium circinatum with Pinus radiata. Plant Dis. 2016, 100, 79–84. [Google Scholar] [CrossRef]
- Martín-García, J.; Paraschiv, M.; Flores-Pacheco, J.A.; Chira, D.; Diez, J.J.; Fernández, M. Susceptibility of Several Northeastern Conifers to Fusarium circinatum and Strategies for Biocontrol. Forests 2017, 8, 318. [Google Scholar]
- Nowakowska, J.A.; Zachara, T.; Konecka, A. Genetic Variability of Scots pine (Pinus sylvestris L.) and Norway Spruce (Picea abies L. Karst.) Natural Regeneration Compared with Their Maternal Stands. For. Res. Pap. 2014, 75, 47–54. [Google Scholar] [CrossRef]
- Bobowicz, M.A.; Korczyk, A.F. Variability of Eight Polish Populations of Pinus sylvestris L. Expressed in Traits of Cones. Acta Soc. Bot. Pol. 1994, 63, 191–198. [Google Scholar] [CrossRef]
- Vivas, M.; Zas, R.; Solla, A. Screening of Maritime Pine (Pinus pinaster) for Resistance to Fusarium circinatum, the Causal Agent of Pitch Canker Disease. Forestry 2011, 85, 185–192. [Google Scholar] [CrossRef]
- Swett, C.L.; Gordon, T.R. Latent Infection by Fusarium circinatum Influences Susceptibility of Monterey Pine Seedlings to Pitch Canker. In Proceedings of the Fourth International Workshop on the Genetics of Host-Parasite Interactions in Forestry: Disease and Insect Resistance in Forest Trees, Eugene, OR, USA, 31 July–5 August 2011; pp. 159–161. [Google Scholar]
- Kwaśna, H.; Bateman, G.L. Microbial Communities in Roots of Pinus sylvestris Seedlings with Damping-Off Symptoms in Two Forest Nurseries as Determined by ITS1/2 rDNA Sequencing. For. Pathol. 2009, 39, 239–248. [Google Scholar] [CrossRef]
- Lazreg, F.; Belabid, L.; Sanchez, J.; Gallego, E.; Bayaa, B. Pathogenicity of Fusarium spp. Associated with Diseases of Aleppo-Pine Seedlings in Algerian Forest Nurseries. J. For. Sci. 2014, 60, 115–120. [Google Scholar] [CrossRef]
- Romón, P.; Iturrondobeitia, J.C.; Gibson, K.; Lindgren, B.S.; Goldarazena, A. Quantitative Association of Bark Beetles with Pitch Canker Fungus and Effects of Verbenone on their Semiochemical Communication in Monterey Pine Forests in Northern Spain. Environ. Entomol. 2007, 36, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Bezos, D.; Martínez-Álvarez, P.; Diez, J.J. The Pine Shoot Beetle Tomicus piniperda as a Plausible Vector of Fusarium circinatum in Northern Spain. Ann. For. Sci. 2015, 72, 1079. [Google Scholar] [CrossRef]


{kind=link}
| Species | Code | Host | Location, Country | Latitude | Longitude |
|---|---|---|---|---|---|
| Fusarium circinatum [20] | FcCa6 | Pinus radiata | Comillas, Cantabria, Spain | 43°20′16.2″ N | 4°18′17.1″ W |
| Fusarium poae | A | Winter wheat ears | Bohemia NE Poland * | 54°09′45″ N | 9°00′40″ E |
| Fusarium graminearum | A | Winter wheat grain | Ozon NE Poland * | 54°09′45″ N | 9°00′40″ E |
| Fusarium tricinctum | A | Winter wheat grain | Julius NE Poland * | 54°09′45″ N | 9°00′40″ E |
| Fusarium oxysporum | D | Winter wheat | Julius NE Poland * | 54°09′45″ N | 9°00′40″ E |
| Fusarium culmorum | D | Winter wheat | Julius NE Poland * | 54°09′45″ N | 9°00′40″ E |
| Fusarium graminearum | D | Winter wheat | Julius NE Poland * | 54°09′45″ N | 9°00′40″ E |
| Scots Pine Provenance * | Latitude | Longitude |
|---|---|---|
| Tuszyma | 50°11′4.342″ N | 21°29′53.38″ E |
| Wejherowo | 54°36′6.144″ N | 18°13′46.42″ E |
| Kluczbork | 51°58′23.639″ N | 18°12′32.983″ E |
| Dojlidy | 53°9′5″ N | 23°8′48.094″ E |
| Kutno | 52°18′9.413″ N | 19°17′40.454″ E |
| Kwidzyn | 53°44′12.331″ N | 18°55′45.452″ E |
| Tuchola | 53°40′9.624″ N | 17°54′31.928″ E |
| Oborniki Śląskie | 51°17′53.093″ N | 16°54′41.685″ E |
| Lidzbark | 53°15′3.828″ N | 19°46′13.794″ E |
| Karczma Borowa | 51°51′4.417″ N | 16°36′54.071″ E |
| Lubsko | 51°47′22.074″ N | 14°58′30.576″ E |
| Czaplinek | 53°33′44.406″ N | 16°13′37.974″ E |
| Jablonna | 52°23′1.454″ N | 20°56′1.816″ E |
| Klosnowo | 53°47′60″ N | 17°37′60″ E |
| Lasowice | 50°54′16.443″ N | 18°15′19.283″ E |
| Debno | 52°37′14.16″ N | 14°37′30.632″ E |
| Isolate | Klosnowo | Lasowice | Dębno | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Root Inhibition, % | Mortality, % | Points | Virulence Score | Root Inhibition, % | Mortality, % | Points | Virulence Score | Root Inhibition, % | Mortality, % | Points | Virulence Score | |
| Fusarium poae A | 14.4 ± 2.8 a | 4.2 ± 0.07 a | 22.54 ± 0.4 a | 12.5 | 16.2 ± 0.9 a | 0 d | 22.13 ± 0.2 a | 14.5 | 15.7 ± 0.2 a | 4.2 ± 0.1 a | 22.17 ± 1.7 a | 14 |
| Fusarium graminearum A | 18.9 ± 1.1 b | 4.2 ± 0.03 a | 21.75 ± 2.1 a | 16 | 19.8 ± 1.1 b | 4.2 ± 0.1 a | 21.33 ± 0.9 a | 18.5 | 17.8 ± 1.8 b | 4.2 ± 0.1 a | 21.96 ± 1.6 a | 15 |
| Fusarium tricinctum A | 64.5 ± 2.6 c | 16.7 ± 2.7 b | 16.87 ± 0.9 b | 40.5 | 59.8 ± 3.4 c | 12.5 ± 1 b | 16 ± 1.1 b | 44.5 | 58.2 ± 3.5 c | 20.8 ± 1.4 b | 16.79 ± 1.3 b | 41 |
| Fusarium oxysporum D | 88.7 ± 6.8 d | 62.5 ± 12.1 c | 11.7 ± 0.72 c | 66.5 | 80.4 ± 7.8 d | 66.7 ± 9 c | 12.2 ± 1.1 c | 64 | 79.4 ± 6.7 d | 66.7 ± 5.1 c | 12.87 ± 0.9 c | 60.5 |
| Fusarium culmorum D | 18.5 ± 1.3 b | 4.2 ± 0.04 a | 21.62 ± 2.0 a | 17 | 20.5 ± 1.91 b | 0 d | 21.8 ± 1.7 a | 16 | 20.4 ± 1.4 b | 0 d | 21.8 ± 1.4 a | 16 |
| Fusarium graminearum D | 19.6 ± 2.1 b | 0 d | 22.41 ± 2.1 a | 13 | 18.5 ± 1.5 b | 0 d | 22.17 ± 1.8 a | 14 | 20.5 ± 2.3 b | 0 d | 21.79 ± 1.7 a | 16 |
| Fusarium circinaum | 98.8 ± 1.1 e | 100 e | 1.62 ± 0.02 e | 99 | 96.7 ± 2.1 e | 100 e | 1.62 ± 0.02 e | 99 | 98.8 ± 1.1 e | 97.1 ± 2.1 e | 1.62 ± 0.02 e | 98 |
| Control | 1.7 ± 0.1 e | 0 d | 24.8 ± 1.7 a | 1 | 1 ± 0.1 e | 0 d | 24.9 ± 1.7 a | 0.5 | 1.2 ± 0.1 e | 0 d | 24.9 ± 1.7 a | 0.5 |
| F. circinatum | F. poae A | F. graminearum A | F. tricinctum A | F. oxysporum D | F. culmorum D | F. graminearum D | Control | |
|---|---|---|---|---|---|---|---|---|
| Mortality, % | 90 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Weakened plants, % | 5 | 0 | 0 | 0 | 5 | 0 | 0 | 0 |
| Lesion length, mm2 | 10.4 ± 2.1 b | 2.15 ± 0.08 a | 2.27 ± 0.21 a | 2.35 ± 0.64 a | 2.45 ± 0.37 a | 1.9 ± 0.62 a | 2.36 ± 0.5 a | 1.3 ± 0.09 a |
| Range, mm | 3.7–14.2 | 1–2.2 | 1.2–4.2 | 1.4–3.6 | 1.7–3.8 | 1–2.2 | 1.1–2.7 | 1–1.4 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Davydenko, K.; Nowakowska, J.A.; Kaluski, T.; Gawlak, M.; Sadowska, K.; García, J.M.; Diez, J.J.; Okorski, A.; Oszako, T. A Comparative Study of the Pathogenicity of Fusarium circinatum and other Fusarium Species in Polish Provenances of P. sylvestris L. Forests 2018, 9, 560. https://doi.org/10.3390/f9090560
Davydenko K, Nowakowska JA, Kaluski T, Gawlak M, Sadowska K, García JM, Diez JJ, Okorski A, Oszako T. A Comparative Study of the Pathogenicity of Fusarium circinatum and other Fusarium Species in Polish Provenances of P. sylvestris L. Forests. 2018; 9(9):560. https://doi.org/10.3390/f9090560
Chicago/Turabian StyleDavydenko, Kateryna, Justyna Anna Nowakowska, Tomasz Kaluski, Magdalena Gawlak, Katarzyna Sadowska, Jorge Martín García, Julio Javier Diez, Adam Okorski, and Tomasz Oszako. 2018. "A Comparative Study of the Pathogenicity of Fusarium circinatum and other Fusarium Species in Polish Provenances of P. sylvestris L." Forests 9, no. 9: 560. https://doi.org/10.3390/f9090560
APA StyleDavydenko, K., Nowakowska, J. A., Kaluski, T., Gawlak, M., Sadowska, K., García, J. M., Diez, J. J., Okorski, A., & Oszako, T. (2018). A Comparative Study of the Pathogenicity of Fusarium circinatum and other Fusarium Species in Polish Provenances of P. sylvestris L. Forests, 9(9), 560. https://doi.org/10.3390/f9090560