Multiple Factors Drive Variation of Forest Root Biomass in Southwestern China

 ,
,
Abstract
:1. Introduction
2. Material and Methods
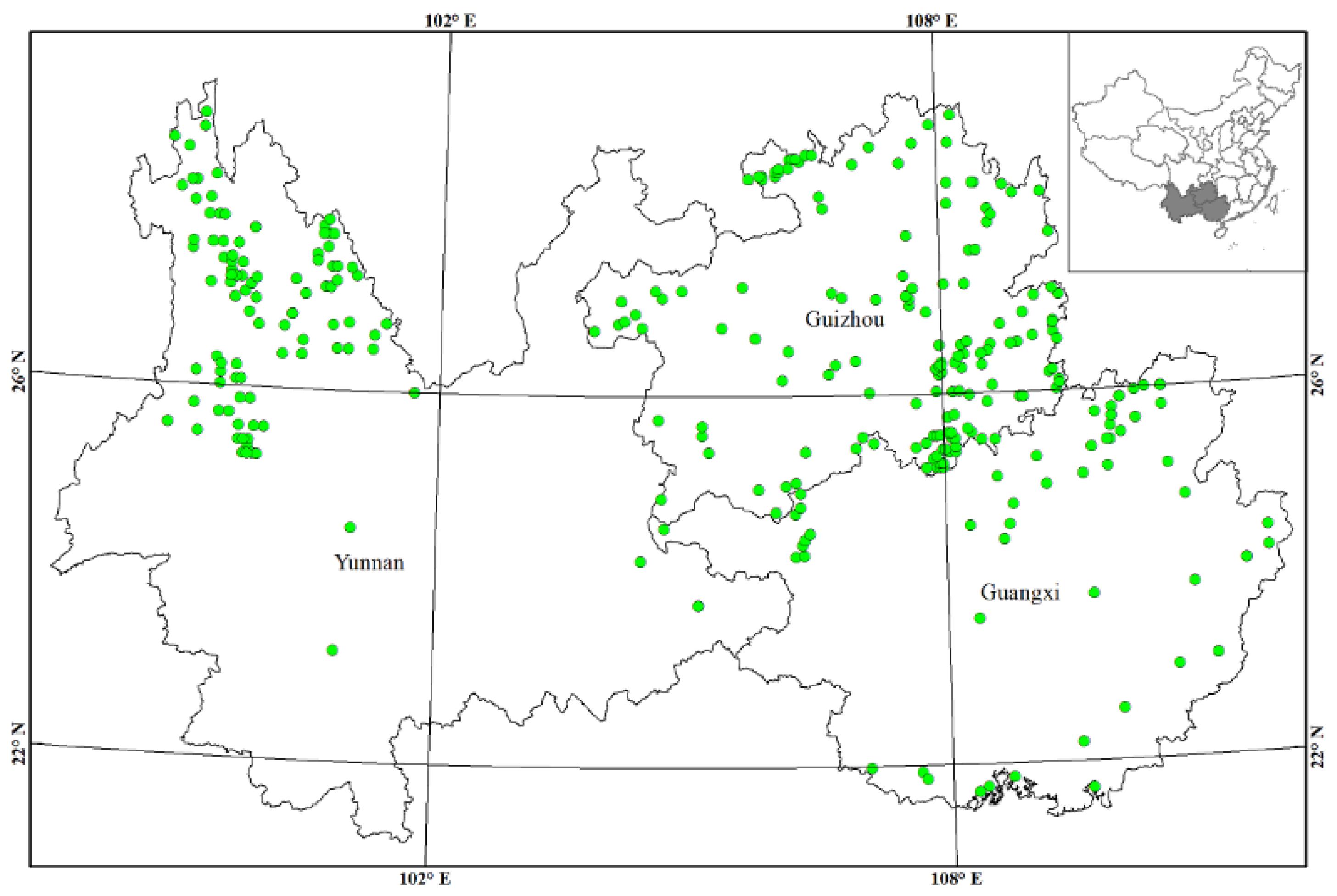
2.1. Study Site
2.2. Forest Biomass Data
2.3. Climatic Variables and Soil Data
2.4. Statistical Analysis
3. Results
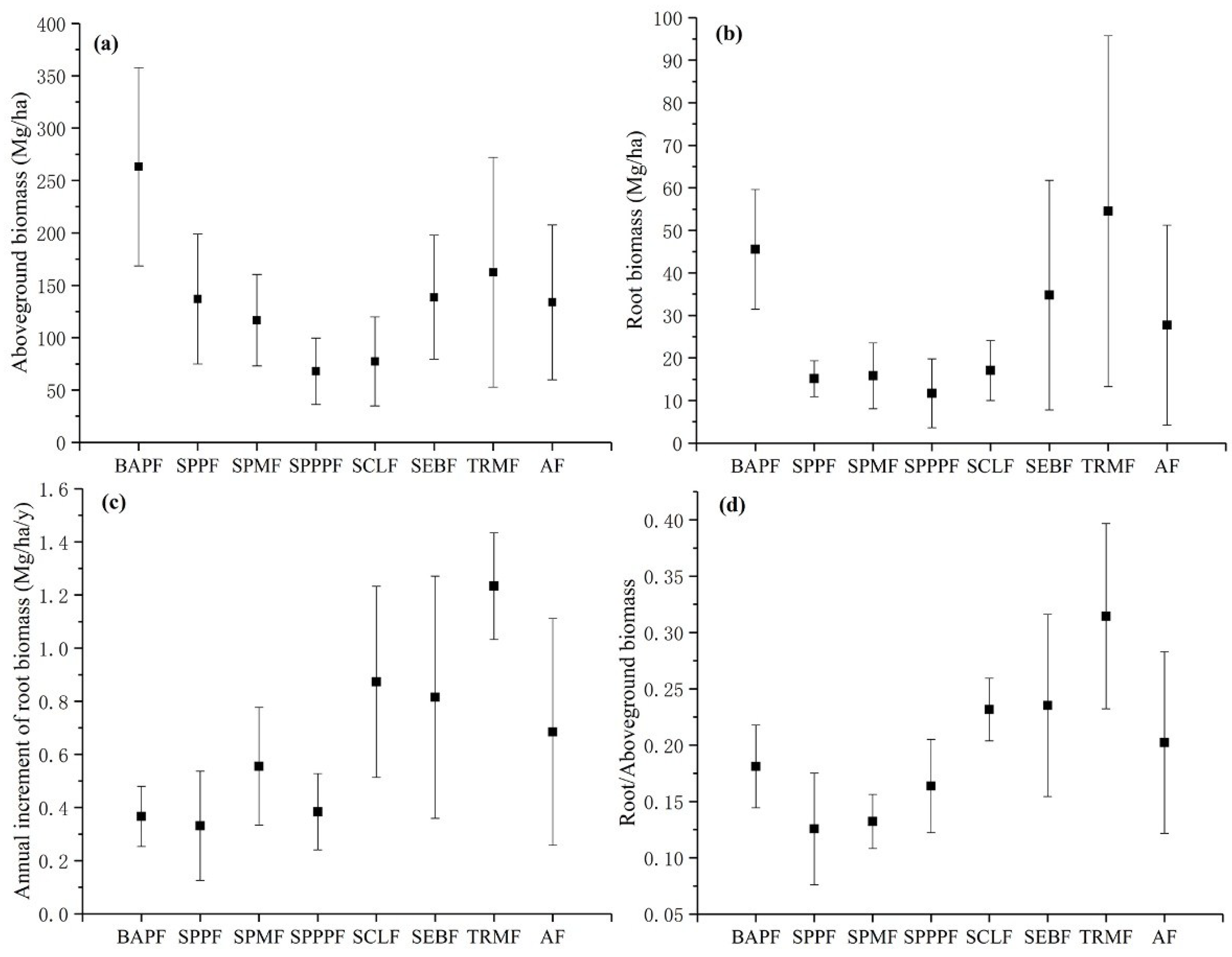
3.1. Statistical and Spatial Patterns of Forest Root Biomass
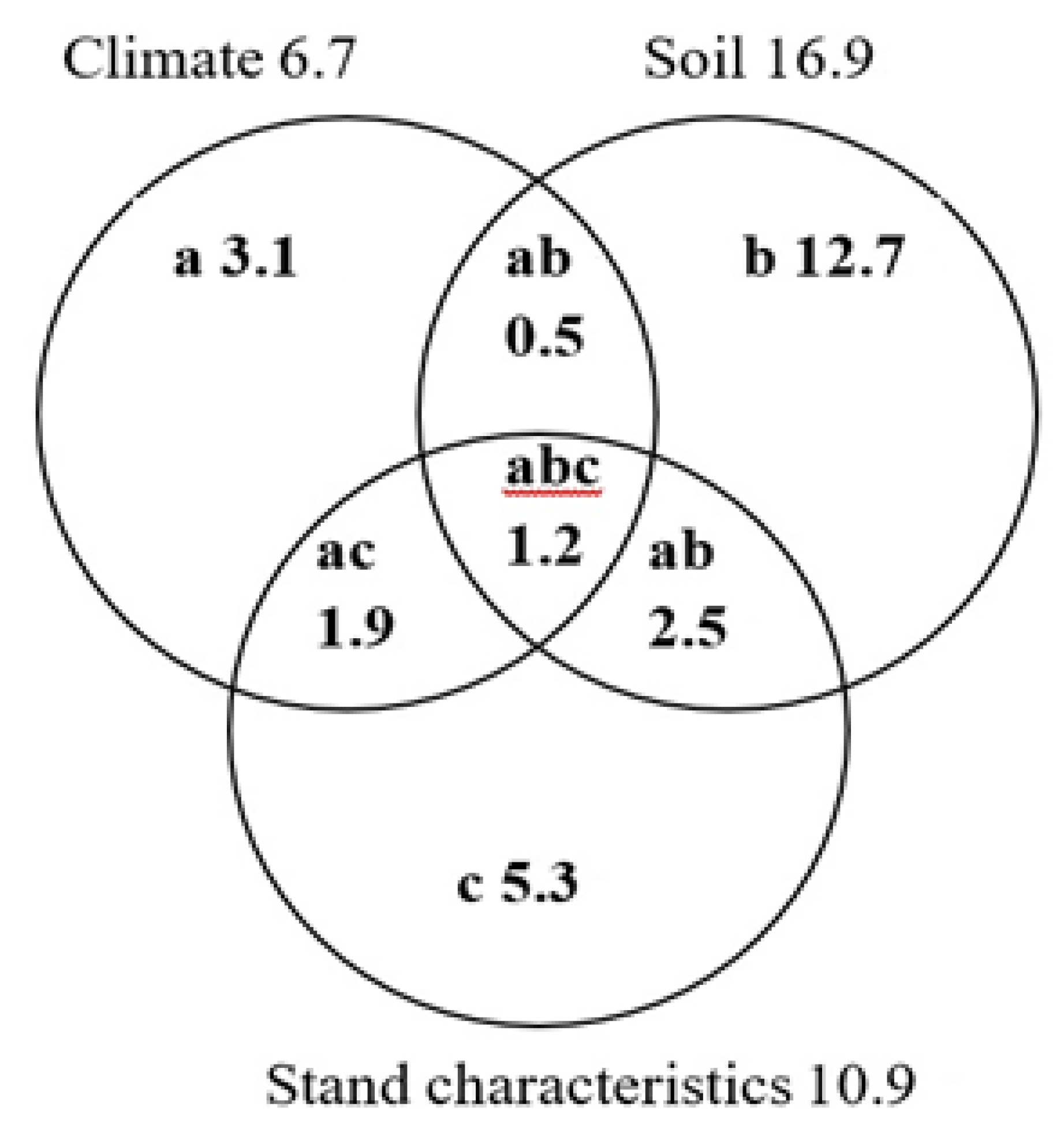
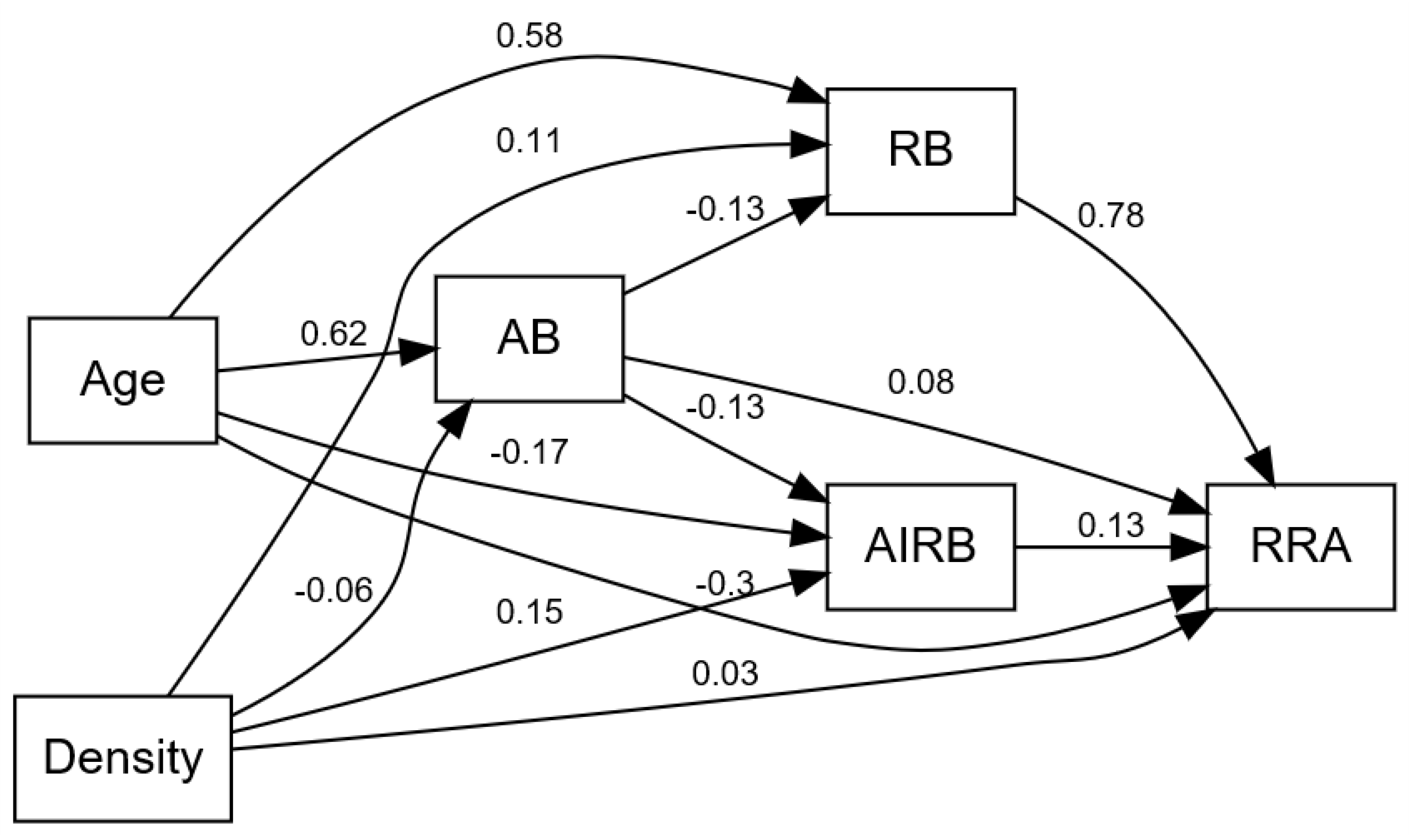
3.2. Factors Influencing Variation in Forest Root Biomass
3.3. Factors Influencing Variation in Forest Root Biomass in Different Forest Types
4. Discussion
4.1. Variation in Forest Root Biomass in Southwestern China
4.2. Factors Influencing Variations in Forest Root Biomass
4.3. Potential of Forest Root Biomass on C Sequestration in Southwestern China
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Houghton, R.A.; Hackler, J.L.; Lawrence, K.T. The U.S. Carbon budget: Contributions from land-use change. Science 1999, 285, 574–578. [Google Scholar] [CrossRef] [PubMed]
- Ni, J. Carbon storage in terrestrial ecosystems of China: Estimates at different spatial resolutions and their responses to climate change. Clim. Chang. 2001, 49, 339–358. [Google Scholar] [CrossRef]
- Fang, J.Y.; Guo, Z.D.; Piao, S.L.; Chen, A.P. Terrestrial vegetation carbon sinks in China, 1981–2000. Sci. China Ser. D Earth Sci. 2007, 50, 1341–1350. [Google Scholar] [CrossRef]
- Cairns, M.; Brown, S.; Helmer, E.; Baumgardner, G. Root biomass allocation in the world’s upland forests. Oecologia 1997, 111, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.Y.; Chen, A.P.; Peng, C.H.; Zhao, S.Q.; Ci, L.J. Changes in forest biomass carbon storage in China between 1949 and 1998. Science 2001, 292, 2320–2322. [Google Scholar] [CrossRef] [PubMed]
- Niklas, K.J. Modeling below- and above-ground biomass for non-woody and woody plants. Ann. Bot. 2005, 95, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Piao, S.; Fang, J.; Zhu, B.; Tan, K. Forest biomass carbon stocks in China over the past 2 decades: Estimation based on integrated inventory and satellite data. J. Geophys. Res. 2005, 110. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Zhao, X.; Bai, Y.; Tang, Z.; Wang, W.; Zhao, Y.; Wan, H.; Xie, Z.; Shi, X.; Wu, B. Carbon pools in China’s terrestrial ecosystems: New estimates based on an intensive field survey. Proc. Natl. Acad. Sci. USA 2018, 115, 4021–4026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poorter, H.; Jagodzinski, A.M.; Ruiz-Peinado, R.; Kuyah, S.; Luo, Y.; Oleksyn, J.; Usoltsev, V.A.; Buckley, T.N.; Reich, P.B.; Sack, L. How does biomass distribution change with size and differ among species? An analysis for 1200 plant species from five continents. New Phytol. 2015, 208, 736–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enquist, B.J.; Niklas, K.J. Global allocation rules for patterns of biomass partitioning across seed plants. Science 2002, 295, 1517–1520. [Google Scholar] [CrossRef] [PubMed]
- Hui, D.F.; Jackson, R.B. Geographical and interannual variability in biomass partitioning in grassland ecosystems: A synthesis of field data. New Phytol. 2006, 169, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Jagodzinski, A.M.; Ziółkowski, J.; Warnkowska, A.; Prais, H. Tree age effects on fine root biomass and morphology over chronosequences of Fagus sylvatica, Quercus robur and Alnus glutinosa stands. PLoS ONE 2016, 11, e0148668. [Google Scholar] [CrossRef] [PubMed]
- Hui, D.; Wang, J.; Le, X.; Shen, W.; Ren, H. Influences of biotic and abiotic factors on the relationship between tree productivity and biomass in China. For. Ecol. Manag. 2012, 264, 72–80. [Google Scholar] [CrossRef]
- Guo, Z.; Hu, H.; Li, P.; Li, N.; Fang, J. Spatio-temporal changes in biomass carbon sinks in China’s forests from 1977 to 2008. Sci. China Life Sci. 2013, 56, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Wang, X.; Zhang, X.; Booth, T.H.; Lu, F. Root: Shoot ratios across China’s forests: Forest type and climatic effects. For. Ecol. Manag. 2012, 269, 19–25. [Google Scholar] [CrossRef]
- Hararuk, O.; Smith, M.J.; Luo, Y. Microbial models with data-driven parameters predict stronger soil carbon responses to climate change. Glob. Chang. Biol. 2015, 21, 2439–2453. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.J.; Wang, X.K.; Zhang, X.Q.; Ren, Y.; Poorter, H. Variation in biomass expansion factors for China’s forests in relation to forest type, climate, and stand development. Ann. For. Sci. 2013, 70, 589–599. [Google Scholar] [CrossRef]
- Jackson, R.B.; Canadell, J.; Ehleringer, J.R.; Mooney, H.A.; Sala, O.E.; Schulze, E.D. A global analysis of root distributions for terrestrial biomes. Oecologia 1996, 108, 389–411. [Google Scholar] [CrossRef] [PubMed]
- Poorter, H.; Nagel, O. The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: A quantitative review. Aust. J. Plant Physiol. 2000, 27, 595–607. [Google Scholar] [CrossRef]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, L.; Chen, X.; Tian, X.; Wang, X.; Ren, Y. Biomass allocation patterns across China’s terrestrial biomes. PLoS ONE 2014, 9, e93566. [Google Scholar] [CrossRef] [PubMed]
- Niklas, K.J.; Enquist, B.J. Invariant scaling relationships for interspecific plant biomass production rates and body size. Proc. Natl. Acad. Sci. USA 2001, 98, 2922–2927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, D.; Niklas, K. Above- and below-ground biomass relationships across 1543 forested communities. Ann. Bot. 2007, 99, 95–102. [Google Scholar] [CrossRef] [PubMed]
- McConnaughay, K.D.M.; Coleman, J.S. Biomass allocation in plants: Ontogeny or optimality? A test along three resource gradients. Ecology 1999, 80, 2581–2593. [Google Scholar] [CrossRef]
- Mokany, K.; Raison, R.J.; Prokushkin, A.S. Critical analysis of root: Shoot ratios in terrestrial biomes. Glob. Chang. Biol. 2006, 12, 84–96. [Google Scholar] [CrossRef]
- Cheng, D.; Zhong, Q.; Niklas, K.J.; Ma, Y.; Yang, Y.; Zhang, J. Isometric scaling of above- and below-ground biomass at the individual and community levels in the understorey of a sub-tropical forest. Ann. Bot. 2015, 115, 303–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Fang, J.; Zhu, B. Forest biomass and root-shoot allocation in northeast China. For. Ecol. Manag. 2008, 255, 4007–4020. [Google Scholar] [CrossRef]
- Yang, Y.H.; Fang, J.Y.; Ji, C.J.; Han, W.X. Above- and belowground biomass allocation in Tibetan grasslands. J. Veg. Sci. 2009, 20, 177–184. [Google Scholar] [CrossRef]
- Bruun, H.H.; Moen, J.; Virtanen, R.; Grytnes, J.A.; Oksanen, L.; Angerbjörn, A. Effects of altitude and topography on species richness of vascular plants, bryophytes and lichens in alpine communities. J. Veg. Sci. 2006, 17, 37–46. [Google Scholar] [CrossRef]
- Burkes, E.C.; Will, R.E.; Barron-Gafford, G.A.; Teskey, R.O.; Shiver, B. Biomass partitioning and growth efficiency of intensively managed Pinus taeda and Pinus elliottii stands of different planting densities. For. Sci. 2003, 49, 224–234. [Google Scholar]
- Ruiz-Robleto, J.; Villar, R. Relative growth rate and biomass allocation in ten woody species with different leaf longevity using phylogenetic independent contrasts (PICs). Plant Biol. 2005, 7, 484–494. [Google Scholar] [CrossRef] [PubMed]
- Majdi, H.; Ohrvik, J. Interactive effects of soil warming and fertilization on root production, mortality, and longevity in a Norway spruce stand in Northern Sweden. Glob. Chang. Biol. 2004, 10, 182–188. [Google Scholar] [CrossRef]
- Zhang, H.; Song, T.Q.; Wang, K.L.; Yang, H.; Yue, Y.M.; Zeng, Z.X.; Peng, W.X.; Zeng, F.P. Influences of stand characteristics and environmental factors on forest biomass and root-shoot allocation in southwest China. Ecol. Eng. 2016, 91, 7–15. [Google Scholar] [CrossRef]
- Zhang, H.; Song, T.Q.; Wang, K.L.; Du, H.; Yue, Y.M.; Wang, G.X.; Zeng, F.P. Biomass and carbon storage in an age-sequence of Cyclobalanopsis glauca plantations in southwest China. Ecol. Eng. 2014, 73, 184–191. [Google Scholar] [CrossRef]
- Yuan, D.X. The geology environment and hydro-ecological problem of karst region. Land Res. South China 2003, 1, 21–25. [Google Scholar]
- Du, H.; Wang, K.L.; Peng, W.X.; Zeng, F.P.; Song, T.Q.; Zhang, H.; Lu, S.Y. Spatial heterogeneity of soil mineral oxide components in depression between karst hills, Southwest China. Chin. Geogr. Sci. 2014, 24, 163–179. [Google Scholar] [CrossRef]
- Cheng, J.Z.; Lee, X.Q.; Theng, B.K.G.; Zhang, L.K.; Fang, B.; Li, F.S. Biomass accumulation and carbon sequestration in an age-sequence of Zanthoxylum bungeanum plantations under the Grain for Green Program in karst regions, Guizhou province. Agric. For. Meteorol. 2015, 203, 88–95. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, K.L.; Zeng, Z.X.; Du, H.; Zeng, F.P. Biomass and carbon storage across a chronosequence of Juglans regia plantations in the karst region, southwest China. Forests 2017, 8, 103. [Google Scholar] [CrossRef]
- Peng, W.X.; Wang, K.L.; Song, T.Q.; Zeng, F.P.; Wang, J.R. Controlling and restoration models of complex degradation vulnerable Karst ecosystem. Acta Ecol. Sin. 2008, 28, 811–820. [Google Scholar]
- Zhang, H.; Wang, K.L.; Xu, X.L.; Song, T.Q.; Xu, Y.F.; Zeng, F.P. Biogeographical patterns of biomass allocation in leaves, stems and roots in China’s forests. Sci. Rep. 2015, 5, 15997. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Song, T.Q.; Wang, K.L.; Wang, G.X.; Liao, J.X.; Xu, G.H.; Zeng, F.P. Biogeographical patterns of biomass allocation vary with climate, soil and forest characteristics in China. Environ. Res. Lett. 2015, 10, 044014. [Google Scholar] [CrossRef]
- Tan, Q.J.; Wang, W.L.; Chen, H.S.; Qin, Z.S.; Zheng, S.F.; Zhang, H.; Du, H.; Song, T.Q. Coupling relationships between plant community and soil characteristics in canyon karst region in south-west China. Bangladesh J. Bot. 2017, 46, 1117–1127. [Google Scholar]
- Condit, R. Research in large, long-term tropical forest plots. Trends Ecol. Evol. 1995, 10, 18–22. [Google Scholar] [CrossRef] [Green Version]
- Han, W.X.; Fang, J.Y.; Reich, P.B.; Ian Woodward, F.; Wang, Z.H. Biogeography and variability of eleven mineral elements in plant leaves across gradients of climate, soil and plant functional type in China. Ecol. Lett. 2011, 14, 788–796. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, M.; Wardle, D.A. Structural equation modelling reveals plant-community drivers of carbon storage in boreal forest ecosystems. Biol. Lett. 2010, 6, 116–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosseel, Y. Lavaan: An R package for structural equation modelling. J. Stat. Softw. 2012, 48, 1–36. [Google Scholar] [CrossRef]
- Keeling, H.C.; Phillips, O.L. The global relationship between forest productivity and biomass. Glob. Ecol. Biogeogr. 2007, 16, 618–631. [Google Scholar] [CrossRef]
- Jagodziński, A.M.; Oleksyn, J. Ecological consequences of silviculture at variable stand densities. I. Stand growth and development. Sylwan 2009, 153, 75–85. [Google Scholar]
- Jagodziński, A.M.; Oleksyn, J. Ecological consequences of silviculture at variable stand densities. II. Biomass production and allocation, nutrient retention. Sylwan 2009, 153, 147–157. [Google Scholar]
- Castaneda-Moya, E.; Twilley, R.R.; Rivera-Monroy, V.H. Allocation of biomass and net primary productivity of mangrove forests along environmental gradients in the Florida coastal everglades, USA. For. Ecol. Manag. 2013, 307, 226–241. [Google Scholar] [CrossRef]
- Austin, J.M.; Mackey, B.G.; Van Niel, K.P. Estimating forest biomass using satellite radar: An exploratory study in a temperate Australian Eucalyptus forest. For. Ecol. Manag. 2003, 176, 575–583. [Google Scholar] [CrossRef]
- Carreiras, J.M.B.; Pereira, J.M.C.; Campagnolo, M.L.; Shimabukuro, Y.E. Assessing the extent of agriculture/pasture and secondary succession forest in the Brazilian Legal Amazon using SPOT VEGETATION data. Remote Sens. Environ. 2006, 101, 283–298. [Google Scholar] [CrossRef]
- Guo, Z.D.; Hu, H.F.; Pan, Y.D.; Birdsey, R.A.; Fang, J.Y. Increasing biomass carbon stocks in trees outside forests in China over the last three decades. Biogeoscienes 2014, 11, 4115–4122. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site Conditions | RB (Mg/ha) | AIRB (Mg/ha/year) | RRA |
|---|---|---|---|
| Longitude (E, °C) | −0.227 ** | 0.277 ** | −0.011 |
| Latitude (N, °C) | −0.062 | −0.267 ** | −0.093 |
| Altitude (m) | 0.218 ** | −0.287 ** | 0.008 |
| Root Parameters | Models | Equations | p | R2 |
|---|---|---|---|---|
| RB (Mg/ha) | 1 | RB = 0.313AGE + 12.956 | <0.001 | 0.215 |
| 2 | RB = 0.348AGE − 2.993LAT + 89.689 | <0.001 | 0.252 | |
| 3 | RB = 0.378AGE − 3.040LAT + 0.002 DEN + 85.859 | <0.001 | 0.265 | |
| AIRB (Mg/ha/year) | 1 | AIRB =0.031MAT + 0.241 | <0.001 | 0.098 |
| 2 | AIRB = 0.028MAT + 0.00007DEN + 0.183 | <0.001 | 0.137 | |
| 3 | AIRB = 0.020MAT + 0.00067DEN + 0.000245MAP − 0.014 | <0.001 | 0.151 | |
| 4 | AIRB = 0.013MAT + 0.00005DEN + 0.000242MAP − 0.002AGE + 0.188 | <0.001 | 0.163 | |
| 5 | AIRB = 0.017MAT + 0.00006DEN + 0.000284MAP − 0.002AGE + 0.146TP + 0.188 | <0.001 | 0.173 | |
| RRA | 1 | RRA = 0.023PH + 0.071 | <0.004 | 0.062 |
| BAFs | Forest Types | Equations | p | R2 |
|---|---|---|---|---|
| RB (Mg/ha) | BAPF | RB = 0.024DEN + 0.263AGE − 33.613TN + 77.663 | <0.001 | 0.663 |
| SPPF | RB = 0.001DEN − 6.071TP + 20.255 | <0.001 | 0.301 | |
| SPMF | RB = −0.01ALT + 0.232AGE + 14.998 | <0.001 | 0.460 | |
| SPPPF | RB = 0.627AGE + 2.025LON − 216.558 | <0.001 | 0.666 | |
| SCLF | RB = 0.003DEN + 10.801 | <0.050 | 0.097 | |
| SEBF | RB = 0.545AGE − 4.369LAT − 1.780LON + 314.346 | <0.001 | 0.416 | |
| TRMF | RB = 1.378AGE + 3.552MAT + 0.923LON − 187.721 | <0.001 | 0.961 | |
| AIRB (Mg/ha/year) | BAPF | AIRB = −0.343TN + 0.000184 + 1.071 | <0.001 | 0.575 |
| SPPF | AIRB = 0.000073DEN − 0.003AGE − 0.012MAT + 0.580 | <0.001 | 0.831 | |
| SPMF | AIRB = 0.039MAT − 0.145 | <0.001 | 0.308 | |
| SPPPF | ― | ― | ― | |
| SCLF | AIRB = 0.000208DEN + 0.470 | <0.012 | 0.154 | |
| SEBF | AIRB = 0.540TP − 0.06AGE + 0.001MAP + 0.101 | <0.001 | 0.173 | |
| TRMF | AIRB = − 0.001AB + 1.362 | <0.001 | 0.964 | |
| RRA | BAPF | RRA = 0.000082DEN − 0.001AGE + 0.54TN | <0.001 | 0.838 |
| SPPF | RRA = 0.000013DEN − 0.001AGE + 0.015LAT − 0.259 | <0.001 | 0.515 | |
| SPMF | RRA = 0.004MAT + 0.000016DEN + 0.046 | <0.001 | 0.493 | |
| SPPPF | ― | ― | ― | |
| SCLF | RRA = −0.006MAT + 0.329 | <0.001 | 0.367 | |
| SEBF | RRA = −0.010LON + 0.001AGE − 0.12LAT − 0.000012DEN + 1.607 | <0.001 | 0.487 | |
| TRMF | RRA = 0.000026DEN − 0.000179AB + 0.001AGE + 0.254 | <0.001 | 0.994 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Wang, K.; Zeng, Z.; Zou, Z.; Xu, Y.; Zeng, F. Multiple Factors Drive Variation of Forest Root Biomass in Southwestern China. Forests 2018, 9, 456. https://doi.org/10.3390/f9080456
Zhang H, Wang K, Zeng Z, Zou Z, Xu Y, Zeng F. Multiple Factors Drive Variation of Forest Root Biomass in Southwestern China. Forests. 2018; 9(8):456. https://doi.org/10.3390/f9080456
Chicago/Turabian StyleZhang, Hao, Kelin Wang, Zhaoxia Zeng, Zhigang Zou, Yanfang Xu, and Fuping Zeng. 2018. "Multiple Factors Drive Variation of Forest Root Biomass in Southwestern China" Forests 9, no. 8: 456. https://doi.org/10.3390/f9080456
APA StyleZhang, H., Wang, K., Zeng, Z., Zou, Z., Xu, Y., & Zeng, F. (2018). Multiple Factors Drive Variation of Forest Root Biomass in Southwestern China. Forests, 9(8), 456. https://doi.org/10.3390/f9080456