Effects of Arbuscular Mycorrhizal Fungi on the Vegetative Vigor of Ailanthus altissima (Mill.) Swingle Seedlings under Sustained Pot Limitation

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Nursery Propagation of Seedlings and Experimental Design
2.2. Biometric Parameters
2.3. Stable Isotopes Analysis
2.4. Statistical Analysis
3. Results
3.1. Growth Parameters
3.2. Leaf Traits
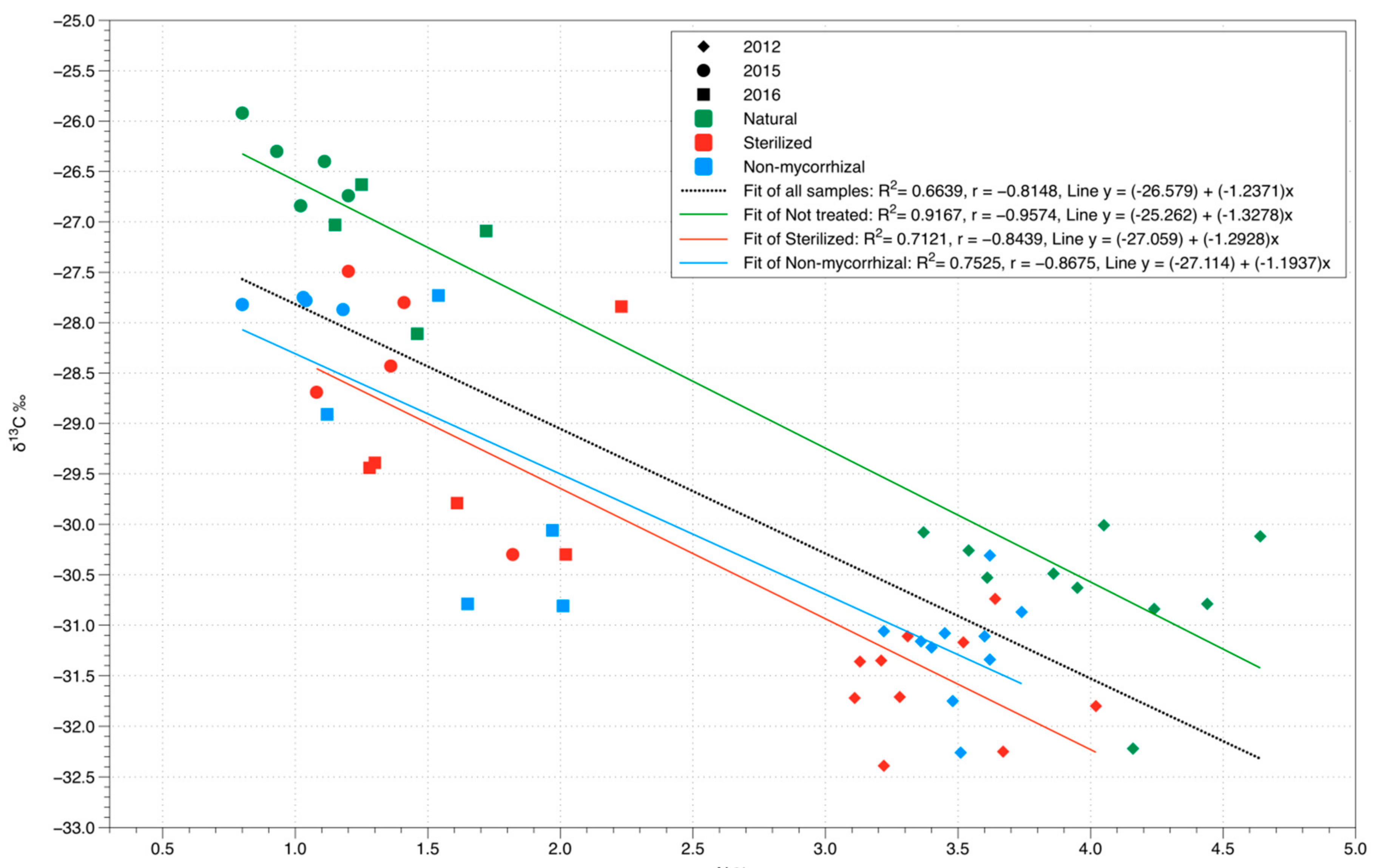
3.3. Stable Isotope Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Alpert, P.; Bone, E.; Holzapfel, C. Invasiveness, invasibility and the role of environmental stress in the spread of non-native plants. Perspect. Plant Ecol. Evol. Syst. 2000, 3, 52–66. [Google Scholar] [CrossRef]
- Blumenthal, D. Interrelated causes of plant invasion. Science 2005, 310, 243–244. [Google Scholar] [CrossRef] [PubMed]
- Lo Verde, G.; La Mantia, T. The role of native flower visitors in pollinating Opuntia ficus-indica (L.) Mill., naturalized in Sicily. Acta Oecol. 2011, 37, 413–417. [Google Scholar] [CrossRef]
- Mitchell, C.E.; Agrawal, A.A.; Bever, J.D.; Gilbert, G.S.; Hufbauer, R.A.; Klironomos, J.N.; Maron, J.L.; Morris, W.F.; Parker, I.M.; Power, A.G.; et al. Biotic interactions and plant invasions. Ecol. Lett. 2006, 9, 726–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pringle, A.; Bever, J.D.; Gardes, M.; Parrent, J.L.; Rillig, M.C.; Klironomos, J.N. Mycorrhizal symbioses and plant invasions. Annu. Rev. Ecol. Evol. 2009, 40, 699–715. [Google Scholar] [CrossRef]
- Rodríguez-Echeverría, S.; Crisóstomo, J.A.; Nabais, C.; Freitas, H. Belowground mutualists and the invasive ability of Acacia longifolia in coastal dunes of Portugal. Biol. Invasions 2009, 11, 651–661. [Google Scholar] [CrossRef]
- Beckstead, J.; Parker, I.M. Invasiveness of Ammophila arenaria: Release from soil-borne pathogens? Ecology 2003, 84, 2824–2831. [Google Scholar] [CrossRef]
- Keane, R.M.; Crawley, M.J. Exotic plant invasions and the enemy release hypothesis. Trends Ecol. Evol. 2002, 17, 164–170. [Google Scholar] [CrossRef]
- Reinhart, K.O.; Callaway, R. Soil biota facilitate Acer invasions in Europe and North America. Ecol. Appl. 2004, 14, 1737–1745. [Google Scholar] [CrossRef]
- Stinson, K.A.; Campbell, S.A.; Powell, J.R.; Wolfe, B.E.; Callaway, R.M.; Thelen, G.C.; Hallett, S.G.; Prati, D.; Klironomos, J.N. Invasive plant suppresses the growth of native tree seedlings by disrupting belowground mutualisms. PLoS ONE 2006, 4, e140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badalamenti, E.; Gristina, L.; Laudicina, V.A.; Novara, A.; Pasta, S.; La Mantia, T. The impact of Carpobrotus cfr. acinaciformis (L.) L. Bolus on soil nutrients, microbial communities structure and native plant communities in Mediterranean ecosystems. Plant Soil 2016, 409, 19–34. [Google Scholar] [CrossRef]
- Badalamenti, E.; La Mantia, T. Stem-injection of herbicide for control of Ailanthus altissima (Mill.) Swingle: A practical source of power for drilling holes in stems. iForest 2013, 6, 123–126. [Google Scholar] [CrossRef]
- Vilà, M.; Tessier, M.; Suehs, C.M.; Brundu, G.; Carta, L.; Galanidis, A.; Lambdon, P.; Manca, M.; Médail, F.; Moragues, E.; et al. Local and regional assessment of the impacts of plant invaders on vegetation structure and soil properties of Mediterranean islands. J. Biogeogr. 2006, 33, 853–861. [Google Scholar] [CrossRef]
- Knapp, L.B.; Canham, C.D. Invasion of an old-growth forest in New York by Ailanthus altissima: Sapling growth and recruitment in canopy gaps. J. Torrey Bot. Soc. 2000, 127, 307–315. [Google Scholar] [CrossRef]
- Kowarik, I.; Säumel, I. Biological flora of central Europe: Ailanthus altissima (Mill.) Swingle. Perspect. Plant Ecol. Evol. Syst. 2007, 8, 207–237. [Google Scholar] [CrossRef]
- Maringer, J.; Wohlgemuth, T.; Neff, C.; Pezzatti, G.B.; Conedera, M. Post-fire spread of alien plant species in a mixed broad-leaved forest of the insubric region. Flora 2012, 207, 19–29. [Google Scholar] [CrossRef]
- Trifilò, P.; Raimondo, F.; Nardini, A.; Lo Gullo, M.A.; Salleo, S. Drought resistance of Ailanthus altissima: Root hydraulics and water relations. Tree Physiol. 2004, 24, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Wickert, K.L.; O’Neal, E.S.; Davis, D.D.; Kasson, M.T. Seed production, viability, and reproductive limits of the invasive Ailanthus altissima (Tree-of-Heaven) within invaded environments. Forests 2017, 8, 226. [Google Scholar] [CrossRef]
- Heisey, R.M. Identification of an allelopathic compound from Ailanthus altissima (Simaroubaceae) and characterization of its herbicidal activity. Am. J. Bot. 1996, 83, 192–200. [Google Scholar] [CrossRef]
- Gómez-Aparicio, L.; Canham, C.D. Neighbourhood analyses of the allelopathic effects of the invasive tree Ailanthus altissima in temperate forests. J. Ecol. 2008, 96, 447–458. [Google Scholar] [CrossRef] [Green Version]
- Callaway, R.M.; Bedmar, E.J.; Reinhart, K.O.; Silvan, C.G.; Klironomos, J.N. Effects of soil biota from different ranges on Robinia invasion: Acquiring mutualists and escaping pathogens. Ecology 2011, 92, 1027–1035. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Qiu, Y.-L. Phylogenetic distribution and evolution of mycorrhizas in land plants. Mycorrhiza 2006, 16, 299–363. [Google Scholar] [CrossRef] [PubMed]
- Nijjer, S.; Rogers, W.E.; Siemann, E. The effect of mycorrhizal inoculum on the growth of five native tree species and the invasive Chinese Tallow tree (Sapium sebiferum). Texas J. Sci. 2004, 56, 357–368. [Google Scholar]
- Cardinale, M.; Brusetti, L.; Lanza, A.; Orlando, S.; Daffonchio, D.; Puglia, A.M.; Quatrini, P. Rehabilitation of Mediterranean anthropogenic soils using symbiotic wild legume shrubs: Plant establishment and impact on the soil bacterial community structure. Appl. Soil Ecol. 2010, 46, 1–8. [Google Scholar] [CrossRef]
- Huebner, C.D.; McQuattie, C.; Rebbeck, J. Mycorrhizal associations in Ailanthus altissima (Simaroubaceae) from forested and non-forested sites. J. Torrey Bot. Soc. 2007, 134, 27–33. [Google Scholar] [CrossRef]
- Badalamenti, E.; La Mantia, T.; Quatrini, P. Arbuscular mycorrhizal fungi positively affect growth of Ailanthus altissima (Mill.) Swingle seedlings and show a strong association with this invasive species in Mediterranean woodlands. J. Torrey Bot. Soc. 2015, 142, 127–139. [Google Scholar] [CrossRef]
- Ripullone, F.; Lauteri, M.; Grassi, G.; Amato, M.; Borghetti, M. Variation in nitrogen supply changes water-use efficiency of Pseudotsuga menziesii and Populus × euroamericana; a comparison of three different approaches to determine water-use efficiency. Tree Physiol. 2004, 24, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Gutiérrez, C.; Dawson, T.E.; Nicolás, E.; Querejeta, J.I. Isotopes reveal contrasting water use strategies among coexisting plant species in a Mediterranean ecosystem. New Phytol. 2012, 196, 489–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badalamenti, E.; Gristina, L.; La Mantia, T.; Novara, A.; Pasta, S.; Lauteri, M.; Fernandes, P.; Correia, O.; Máguas, C. Relationship between recruitment and mother plant vitality in the alien species Acacia cyclops A. Cunn. ex G. Don. For. Ecol. Manag. 2014, 331, 237–244. [Google Scholar] [CrossRef] [Green Version]
- Brugnoli, E.; Farquhar, G.D. Photosynthetic fractionation of carbon isotopes. In Photosynthesis: Physiology and Metabolism; Leegood, R.C., Sharkey, T.D., Von Caemmerer, S., Eds.; Kluwer: Dordrecht, The Netherlands, 2000; pp. 399–434. [Google Scholar]
- Lauteri, M.; Brugnoli, E.; Spaccino, L. Carbon isotope discrimination in leaf soluble sugars and in whole plant dry matter in Helianthus annuus L. grown at different water conditions. In Physiological Ecology Series: “Stable Isotopes and Plant Carbon-Water Relations”; Ehleringer, J.R., Hall, A.E., Farquhar, G.D., Eds.; Academic Press: San Diego, CA, USA, 1993; pp. 93–108. [Google Scholar]
- Liu, C.G.; Wang, Y.J.; Pan, K.W.; Jin, Y.Q.; Li, W.; Zhang, L. Effects of phosphorus application on photosynthetic carbon and nitrogen metabolism, water use efficiency and growth of dwarf bamboo (Fargesia rufa) subjected to water deficit. Plant Physiol. Biochem. 2015, 96, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Mediavilla, S.; Escudero, A. Relative growth rate of leaf biomass and leaf nitrogen content in several Mediterranean woody species. Plant Ecol. 2003, 168, 321–332. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Physiol. Plant. Mol. Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Richardson, D.M.; Allsopp, N.; D’Antonio, C.M.; Milton, S.J.; Rejmanek, M. Plant invasions—The role of mutualisms. Biol. Rev. 2000, 75, 65–93. [Google Scholar] [CrossRef] [PubMed]
- Nuñez, M.A.; Dickie, I.A. Invasive belowground mutualists of woody plants. Biol. Invasions 2014, 16, 645–661. [Google Scholar] [CrossRef]
- Troelstra, S.R.; Wagenaar, R.; Smant, W.; Peters, B.A.M. Interpretation of bioassays in the study of interactions between soil organisms and plants: Involvement of nutrient factors. New Phytol. 2001, 150, 697–706. [Google Scholar] [CrossRef]
- Schulze, E.D.; Turner, N.C.; Nicolle, D.; Schumacher, J. Leaf and wood carbon isotope ratios, specific leaf areas and wood growth of Eucalyptus species across a rainfall gradient in Australia. Tree Physiol. 2006, 26, 479–492. [Google Scholar] [CrossRef] [PubMed]
- Warren, C.R.; Dreyer, E.; Tausz, M.; Adams, M.A. Ecotype adaptation and acclimation of leaf traits to rainfall in 29 species of 16-year-old Eucalyptus at two common gardens. Funct. Ecol. 2006, 20, 929–940. [Google Scholar] [CrossRef]
- Evans, J.R. Photosynthesis and nitrogen relationships in leaves of C3 plants. Oecologia 1989, 78, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Ripullone, F.; Grassi, G.; Lauteri, M.; Borghetti, M. Photosynthesis–nitrogen relationships: Interpretation of different patterns between Pseudotsuga menziesii and Populus × euroamericana in a mini-stand experiment. Tree Physiol. 2003, 23, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Kattge, J.; Knorr, W.; Raddatz, T.; Wirth, C. Quantifying photosynthetic capacity and its relationship to leaf nitrogen content for global-scale terrestrial biosphere models. Glob. Chang. Biol. 2009, 15, 976–991. [Google Scholar] [CrossRef]
- Ali, A.A.; Xu, C.G.; Rogers, A.; McDowell, N.G.; Medlyn, B.E.; Fisher, R.A.; Wullschleger, S.D.; Reich, P.B.; Vrugt, J.A.; Bauerle, W.L.; et al. Global-scale environmental control of plant photosynthetic capacity. Ecol. Appl. 2015, 25, 2349–2365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handley, L.L.; Azcon, R.; Lozano, J.M.R.; Scrimgeour, C.M. Plant δ15N associated with arbuscular mycorrhization, drought and nitrogen deficiency. Rapid Commun. Mass Spectrom. 1999, 13, 1320–1324. [Google Scholar] [CrossRef]
- Querejeta, J.I.; Allen, M.F.; Caravaca, F.; Roldán, A. Differential modulation of host plant δ13C and δ18O by native and nonnative arbuscular mycorrhizal fungi in a semiarid environment. New Phytol. 2006, 169, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Handley, L.L.; Daft, M.J.; Wilson, J.; Scrimgeour, C.M.; Ingleby, K.; Sattar, M.A. Effects of the ecto- and VA-mycorrhizal fungi Hydnagium carneum and Glomus clarum on the δ15N and δ13C values of Eucalyptus globulus and Ricinus communis. Plant Cell Environ. 1993, 16, 375–382. [Google Scholar] [CrossRef]
- Brueck, H.; Senbayram, M. Low nitrogen supply decreases water-use efficiency of oriental tobacco. J. Plant Nutr. Soil Sci. 2009, 172, 216–223. [Google Scholar] [CrossRef]
- Livingston, N.J.; Guy, R.D.; Sun, Z.J.; Ethier, G.J. The effects of nitrogen stress on the stable carbon isotope composition, productivity and water use efficiency of white spruce (Picea glauca (Moench) Voss) seedlings. Plant Cell Environ. 1999, 22, 281–289. [Google Scholar] [CrossRef]
- Cabrera-Bosquet, L.; Molero, G.; Bort, J.; Nogués, S.; Araus, J.L. The combined effect of constant water deficit and nitrogen supply on WUE, NUE and Δ13C in durum wheat potted plants. Ann. Appl. Biol. 2007, 151, 277–289. [Google Scholar] [CrossRef]
- Bruck, H.; Jureit, C.; Hermann, M.; Schulz, A.; Sattelmacher, B. Effects of water and nitrogen supply on water use efficiency and carbon isotope discrimination in edible canna (Canna edulis Ker-Gawler). Plant Biol. 2001, 3, 326–334. [Google Scholar] [CrossRef]
- Guehl, J.M.; Fort, C.; Ferhi, A. Differential response of leaf conductance, carbon isotope discrimination and water-use efficiency to nitrogen deficiency in maritime pine and pedunculate oak plants. New Phytol. 1995, 131, 149–157. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Fan, Y.; Long, J.; Wei, R.; Kjelgren, R.; Gong, C.; Zhao, J. Effects of soil water and nitrogen availability on photosynthesis and water use efficiency of Robinia pseudoacacia seedlings. J. Environ. Sci. 2013, 25, 585–595. [Google Scholar] [CrossRef]
- Cabrera-Bosquet, L.; Molero, G.; Nogués, S.; Araus, J.L. Water and nitrogen conditions affect the relationships of Δ13C and Δ18O to gas exchange and growth in durum wheat. J. Exp. Bot. 2009, 60, 1633–1644. [Google Scholar] [CrossRef] [PubMed]
- Greer, G.K.; Dietrich, M.A.; Lincoln, J.M. Ailanthus altissima stimulates legume nodulation in Trifolium pratense via root exudates: A novel mechanism facilitating invasion? Int. J. Plant Sci. 2016, 177, 400–408. [Google Scholar] [CrossRef]
- Evans, J.R.; Seemann, J.R. The allocation of protein nitrogen in the photosynthetic apparatus: Costs, consequences, and control. In Photosynthesis; Briggs, W.R., Ed.; A.R. Liss: New York, NY, USA, 1989; pp. 183–205. [Google Scholar]
- Makino, A. Rubisco and nitrogen relationships in rice. Leaf photosynthesis and plant growth. Soil Sci. Plant Nutr. 2003, 49, 319–327. [Google Scholar] [CrossRef]
- Kumar, P.A.; Parry, M.A.J.; Mitchell, R.A.C.; Ahmad, A.; Abrol, Y.P. Photosynthesis and nitrogen-use efficiency. In Photosynthetic Nitrogen Assimilation and Associated Carbon and Respiratory Metabolism; Foyer, C.H., Noctor, G., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands; Boston, MA, USA; London, UK, 2002; pp. 23–34. [Google Scholar]
- Walters, M.B.; Reich, P.B. Response of Ulmus americana seedlings to varying nitrogen and water status. 1 Photosynthesis and growth. Tree Physiol. 1989, 5, 159–172. [Google Scholar] [CrossRef] [PubMed]
- Hobbie, E.A.; Hӧgberg, P. Nitrogen isotopes link mycorrhizal fungi and plants to nitrogen. New Phytol. 2012, 196, 367–382. [Google Scholar] [CrossRef] [PubMed]
- Evans, R.D. Physiological mechanisms influencing plant nitrogen isotope composition. Trends Plant Sci. 2001, 6, 121–126. [Google Scholar] [CrossRef]
- Craine, J.M.; Elmore, A.J.; Aidar, M.P.M.; Bustamante, M.; Dawson, T.E.; Hobbie, E.A.; Kahmen, A.; Mack, M.C.; McLauchlan, K.K.; Michelsen, A.; et al. Global patterns of foliar nitrogen isotopes and their relationships with climate, mycorrhizal fungi, foliar nutrient concentrations, and nitrogen availability. New Phytol. 2009, 183, 980–992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azcón-G.-Aguilar, R.; Handley, L.L.; Scrimgeour, C.M. The δ15N of lettuce and barley are affected by AM status and external concentration of N. New Phytol. 1998, 138, 19–26. [Google Scholar]
- Michelsen, A.; Schmidt, I.K.; Jonasson, S.; Quarmby, C.; Sleep, D. Leaf 15N abundance of subarctic plants provides field evidence that ericoid, ectomycorrhizal and non- and arbuscular mycorrhizal species access different sources of soil nitrogen. Oecologia 1996, 105, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Hodge, A.; Storer, K. Arbuscular mycorrhiza and nitrogen: Implications for individual plants through to ecosystems. Plant Soil 2015, 386, 1–19. [Google Scholar] [CrossRef]
- Corrêa, A.; Cruz, C.; Ferrol, N. Nitrogen and carbon/nitrogen dynamics in arbuscular mycorrhiza: The great unknown. Mycorrhiza 2015, 25, 499–515. [Google Scholar] [CrossRef] [PubMed]
- Johnson, N.C.; Graham, J.; Smith, F. Functioning of mycorrhizal associations along the mutualism–parasitism continuum. New Phytol. 1997, 135, 575–586. [Google Scholar] [CrossRef]
- Püschel, D.; Janoušková, M.; Hujslová, M.; Slavíková, R.; Gryndlerová, H.; Jansa, J. Plant–fungus competition for nitrogen erases mycorrhizal growth benefits of Andropogon gerardii under limited nitrogen supply. Ecol. Evol. 2016, 6, 4332–4346. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Soil Treatment | Height (cm) | Stem Diameter (cm) | ||||
|---|---|---|---|---|---|---|
| Year | 2012 | 2015 | 2016 | 2012 | 2015 | 2016 |
| Natural (NT) | 11.0 a | 57.2 a | 75.1 a | 0.6 a | 0.9 a | 1.0 a |
| Sterilized (ST) | 9.9 b | 61.2 a | 76.3 a | 0.4 b | 0.6 b | 0.9 a |
| Non Mycorrhizal (NM) | 7.7 c | 59.0 a | 76.2 a | 0.3 c | 0.6 b | 0.8 b |
| Soil Treatment | Leaf Number (N) | Leaf Area (cm2) | Leaf Mass Area (g m−2) | Specific Leaf Area (cm2 g−1) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Year | 2012 | 2015 | 2016 | 2012 | 2015 | 2016 | 2012 | 2015 | 2016 | 2012 | 2015 | 2016 |
| Natural (NT) | 9.9 a ± 0.6 | 10.2 a ± 1.1 | 10.4 a ± 0.5 | 160.82 a | 1539.90 a | 1607.01 a | 22.5 a | 51.8 a | 32.0 a | 448.7 a | 199.4 b | 326.1 a |
| Sterilized (ST) | 8.4 a ± 0.3 | 9.8 a ± 0.6 | 10.2 a ± 0.7 | 87.35 b | 1389.73 a | 1516.84 a | 22.4 a | 35.8 b | 29.2 a | 456.0 a | 284.5 a | 350.4 a |
| Non Mycorrhizal (NM) | 7.3 b ± 0.2 | 8.6 a ± 1.0 | 9.2 a ± 1.2 | 48.39 c | 1325.63 a | 1480.11 a | 24.1 a | 41.4 b | 25.5 a | 419.8 a | 261.6 a | 404.4 a |
| Soil Treatment | RGR (Days−1) | ||
|---|---|---|---|
| Year | 2012 | 2015 | 2016 |
| Natural (NT) | 0.0347 a ± 0.0007 | 0.0026 b ± 0.0003 | 0.0004 a ± 0.0002 |
| Sterilized (ST) | 0.0279 b ± 0.0016 | 0.0030 ba ± 0.0001 | 0.0005 a ± 0.0001 |
| Non Mycorrhizal (NM) | 0.0233 c ± 0.0024 | 0.0035 a ± 0.0006 | 0.0004 a ± 0.0001 |
| Soil Treatment | δ13C (‰) | δ15N (‰) | N (%) | NxLMA (g m−2) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Year | 2012 | 2015 | 2016 | 2012 | 2015 | 2016 | 2012 | 2015 | 2016 | 2012 | 2015 | 2016 |
| Natural (NT) | −30.6 a | −26.4 a | −27.2 a | 8.2 a | 9.2 a | 7.7 a | 4.0 a | 1.0 a | 1.4 a | 0.9 a | 0.5 a | 0.4 a |
| Sterilized (ST) | −31.6 b | −28.5 b | −29.4 b | 7.9 a | 9.5 a | 8.1 a | 3.4 b | 1.4 a | 1.7 a | 0.8 b | 0.5 a | 0.5 a |
| Non Mycorrhizal (NM) | −31.2 b | −27.8 b | −29.7 b | 7.7 a | 7.7 b | 8.6 a | 3.5 b | 1.0 a | 1.7 a | 0.8 ab | 0.4 a | 0.4 a |
| Treatment | Year | δ13C vs. N×LMA | δ13C vs. %N |
|---|---|---|---|
| Natural | 2012 | 0.14 | 0.06 |
| Sterilized | 2012 | 0.20 | 0.00 |
| Non-mycorrhizal | 2012 | 0.28 | 0.03 |
| Natural | 2015 | 0.16 | 0.60 |
| Sterilized | 2015 | 0.09 | 0.57 |
| Non-mycorrhizal | 2015 | 0.45 | 0.00 |
| Natural | 2016 | 0.42 | 0.10 |
| Sterilized | 2016 | 0.16 | 0.14 |
| Non-mycorrhizal | 2016 | 0.02 | 0.35 |
| All treatments | 2012 | 0.33 | 0.11 |
| All treatments | 2015 | 0.05 | 0.57 |
| All treatments | 2016 | 0.03 | 0.14 |
| All treat. all years | 2012–2015–2016 | 0.34 | 0.67 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Badalamenti, E.; Ciolfi, M.; Lauteri, M.; Quatrini, P.; Mantia, T.L. Effects of Arbuscular Mycorrhizal Fungi on the Vegetative Vigor of Ailanthus altissima (Mill.) Swingle Seedlings under Sustained Pot Limitation. Forests 2018, 9, 409. https://doi.org/10.3390/f9070409
Badalamenti E, Ciolfi M, Lauteri M, Quatrini P, Mantia TL. Effects of Arbuscular Mycorrhizal Fungi on the Vegetative Vigor of Ailanthus altissima (Mill.) Swingle Seedlings under Sustained Pot Limitation. Forests. 2018; 9(7):409. https://doi.org/10.3390/f9070409
Chicago/Turabian StyleBadalamenti, Emilio, Marco Ciolfi, Marco Lauteri, Paola Quatrini, and Tommaso La Mantia. 2018. "Effects of Arbuscular Mycorrhizal Fungi on the Vegetative Vigor of Ailanthus altissima (Mill.) Swingle Seedlings under Sustained Pot Limitation" Forests 9, no. 7: 409. https://doi.org/10.3390/f9070409