Effects of a Heat Wave on Nocturnal Stomatal Conductance in Eucalyptus camaldulensis



1
School of Life Science and Engineering, Southwest University of Science and Technology, Mianyang 621010, China
2
Department of Crop and Forest Sciences and AGROTECNIO Center, Universitat de Lleida, 25198 Lleida
3
Department of Environmental Studies, University of California, Santa Cruz, CA 95064, USA, Spain
4
Hawkesbury Institute for the Environment, Western Sydney University, Richmond NSW 2753, Australia
*
Author to whom correspondence should be addressed.
Forests 2018, 9(6), 319; https://doi.org/10.3390/f9060319
Submission received: 5 March 2018
/
Revised: 10 May 2018
/
Accepted: 31 May 2018
/
Published: 3 June 2018
(This article belongs to the Special Issue Water and Gas Exchanges in Forests)
Abstract
:Nocturnal transpiration constitutes a significant yet poorly understood component of the global water cycle. Modeling nocturnal transpiration has been complicated by recent findings showing that stomata respond differently to environmental drivers over day- vs. night-time periods. Here, we propose that nocturnal stomatal conductance depends on antecedent daytime conditions. We tested this hypothesis across six genotypes of Eucalyptus camaldulensis Dehnh. growing under different CO2 concentrations (ambient vs. elevated) and exposed to contrasting temperatures (ambient vs. heat wave) for four days prior to the night of measurements, when all plants experienced ambient temperature conditions. We observed significant effects after the heat wave that led to 36% reductions in nocturnal stomatal conductance. The response was partly driven by changes in daytime stomatal behavior but additional factors may have come into play. We also observed significant differences in response to the heat wave across genotypes, likely driven by local adaptation to their climate of origin, but CO2 played no effect. Stomatal models may need to incorporate the role of antecedent effects to improve projections particularly after drastic changes in the environment such as heat waves.
1. Introduction
The opening of stomata during dark periods defies established conventions in both hydrology and plant physiology. Major assumptions in hydrometeorology include negligible nocturnal evapotranspiration [1], but sap flux studies record that, averaged across a range of species, nighttime transpiration amounts to 12% of daytime [2]. Including nocturnal transpiration into land surface models leads to reductions of up to 50% of the projected available soil moisture in semiarid environments [3]. Understanding the function of nocturnal water loss remains a conundrum because one of the organizing principles in plant physiology is that stomata operate optimally to minimize the rate of water lost per unit of assimilated carbon [4]. Consequently, stomata are theorized to remain closed overnight in C3 and C4 plants because there is no net photosynthetic CO2 assimilation at that time; however this contrasts with reality, where significant nocturnal stomatal opening across species seems to be the norm rather than the exception [5,6,7].
Understanding the drivers of stomatal function in the dark has proven more challenging than anticipated. This is partly because nighttime stomatal responses do not always follow the same pattern as daytime responses [8]. Some studies document that nocturnal stomatal conductance responds positively to elevated CO2 [9], which is opposite to the common diurnal response. Also, stomatal responses to vapor pressure deficit may be either neutral (i.e., no response) or positive, which again contradict the daytime pattern [10,11].
One of the possible mechanisms explaining differential day/night stomatal regulation is the effect of antecedent conditions. For instance, Easlon and Richards [12] found that nocturnal stomatal conductance in Vicia faba L. decreased after days with reduced rates of carbon assimilation induced by low CO2 concentrations. They associated this response with a potentially lower availability of starch, a compound that can be converted to sugars that serve as an osmoticant necessary for stomatal regulation. However, subsequent studies observed that nocturnal stomatal conductance was constant after exposing canopies of bean and cotton to different levels of solar radiation, which indicated a limited potential for a day/night carry-over effect driven by daytime assimilation in those species [13].
Heat waves are an environmental factor which may impact subsequent nocturnal conductance. Important stresses during the day may modify leaf anatomical or physiological properties and exert carry-over effects beyond the duration of the stress event. For instance, Duarte et al. [14] observed reduced nocturnal stomatal conductance in the weeks following a series of heat waves in Pseudotsuga menziesii (Mirb.) Franco, and they attributed this response to changes in stomatal occlusion or greater epicuticular wax. When such changes occur, which affect stomatal behavior as a whole (that is, day- and night- time stomatal behavior), one may expect that daytime and nighttime nocturnal stomatal conductance will vary accordingly in a coordinated fashion. However, other stomatal responses to heat waves are also possible. Some studies hypothesize, for instance, that nocturnal stomatal conductance favors stem water recharge [6]. If true, the high daytime transpiration that occurs during a heat wave, which depletes stem water capacitance, should lead to increased nocturnal stomatal conductance for xylem refilling. Alternatively, if assimilation is reduced during the heat wave [12], we should observe a reduction of nocturnal stomatal conductance, according to the hypothesis that daytime assimilation controls nighttime stomatal conductance.
The potential for antecedent daytime conditions affecting water fluxes in the following night, and responses during heat waves, are not well understood. To the best of our knowledge, the study with P. menziesii, a conifer [14], is the only study to measure nocturnal stomatal conductance after heat waves. However, elucidating the mechanisms explaining stomatal regulation in the dark is key towards improving stomatal models, which often show mixed results over nocturnal periods [14,15]. Furthermore, the increasing frequency of heat waves is more consistently projected to occur in the future [16]. Understanding the responses of nocturnal stomatal conductance to heat waves and potential carry-over effects thus remains an important research need.
Here, we sought to disentangle the possible effects of antecedent daytime conditions on nocturnal stomatal conductance by examining the response on the night immediately following a heat-wave event for genotypes of Eucalyptus camaldulensis grown in ambient and elevated CO2 concentrations. More specifically, we addressed the following questions: (1) Does nocturnal conductance change in response to heat waves, even after the heat wave ends? (2) Is the response of nocturnal conductance to heat waves affected by different growth CO2 concentrations? (3) How much does genotypic variability contribute to the response of nocturnal conductance to heat waves and CO2?
To address the first question, we hypothesized that antecedent heat-wave conditions would affect nocturnal stomatal conductance. To understand how that impact would occur, we wanted to test three mutually compatible hypotheses: (i) stress from the heat wave affects overall stomatal behavior, including nighttime conductance, and consequently day and night conductance would be correlated; (ii) heat-wave-induced changes in photosynthetic carbon assimilation correlate positively with nocturnal stomatal conductance (potentially via carbohydrate feedbacks); (iii) heat-wave-induced changes in daytime whole-plant transpiration correlate positively with nocturnal stomatal conductance (potentially through stem recharge). To address the second question, we hypothesized that heat-wave-induced effects would depend on CO2 concentrations because elevated CO2 often increases photosynthesis in the absence of nutrient limitations, and this could counter the negative effects of the heat wave. To address the third question, we used different genotypes because we found genetic control of nocturnal stomatal conductance in this species [17] and, consequently, we expected differences based on the climate of origin. Furthermore, previous studies have proposed examining responses across genotypes to increase the reproducibility of the results [18].
2. Materials and Methods
2.1. Plants and Growing Conditions
Seedlings from six different genotypes of E. camaldulensis subsp. camaldulensis were prepared from clonal hedges by the Commonwealth Scientific and Industrial Research Organization (see Table A1 for full details on provenances). The hedges were half-sib seedlings originating from provenances representing different geographic and climatic origins. After reaching an average height of 24.6 cm (±0.97; SE) and a basal diameter of 1.86 mm (±0.07), genotypes were transplanted into 6.9 L cylindrical pots and grown at the naturally lit (with 20% reduction of incident radiation) glasshouse facilities of Western Sydney University in Richmond, New South Wales, in southeastern Australia. No differences in height and diameter occurred between genotypes at experiment initiation (P > 0.05, ANOVA).
Each pot contained 7.5 kg of coarse textured soil (supplied by Australian Native Landscape, Richmond, NSW, Australia), with a pH of 6.5. To ensure that no nutrient limitations occurred, the plants were fertilized every fortnight with a commercial liquid fertilizer (500 mL Aquasol at 1.6 g L−1; 23% N, 4% P, 18% K, 0.05% Zn, 0.06% Cu, 0.013% Mo, 0.15% Mn, 0.06% Fe, 0.011% B; Yates Australia, Padstow, NSW, Australia). Plants were grown in ambient (400 ppm) or elevated (640 ppm) CO2 concentrations and were randomly assigned to one of two glasshouse bays per CO2 treatment within a randomized block design (n = 3–6). Pots were rotated between (monthly) and within (weekly) glasshouse bays to further reduce potential glasshouse bay effects on plant performance. Air temperatures (25:17 °C, average day:night) and relative humidity (45%:60%) were representative of average summer values in Richmond. The pots were daily watered to field capacity (until water drained from the bottom of the pot) and, consequently, soil moisture variation did not affect our results.
The heat wave was imposed in half of the plants after three months of growth, and air temperatures in the glasshouse were increased by 10 °C (day and night) for four days. We estimated the change in daytime relative humidity during the heat wave from the ratio of actual vapor pressure under ambient conditions divided by the saturation pressure during the heat wave [19]. Consequently, relative humidity during the heat wave was estimated at 25%. Vapour pressure deficit thus increased from 1.75 kPa during normal conditions to 4.23 kPa during the heat wave. At dusk of the fourth day/night transition, air temperatures were returned to pre-heat-wave conditions. Daytime measurements were collected on the fourth day of the heat wave and nighttime measurements on the night immediately following that day, when air temperatures were already back to normal. In other words, daytime measurements were collected with half of the plants experiencing a heat wave (and the other half not experiencing a heat wave), but nighttime measurements were performed immediately after the heat wave under ambient pre-heat-wave conditions and, consequently, all plants experienced the same environment. Further details on glasshouse design and set-up are given by [17,20].
2.2. Measurements and Statistical Analyses
Leaf gas exchange was measured with four cross-calibrated portable photosynthesis systems (LI-6400XT, Li-Cor Inc., Lincoln, NE, USA). Cross-calibration here means the systems had H2Ov and CO2 removed to set the zero and ambient gains on the infrared gas analyzers simultaneously, side-by-side, under the same physical conditions, and using the same lots of soda lime and Drierite. Nocturnal gas exchange was measured at 3:45 a.m. (predawn). Conditions inside the LI-6400XT cuvettes were set to match the growth conditions previously described. To test whether nighttime stomatal conductance (gn) followed daytime stomatal conductance (gd), we measured gn and gd. To test whether net assimilation (Anet) affects gn, we also measured Anet. To test whether larger diurnal transpiration favors larger gn, we multiplied leaf level transpiration by total leaf area, rendering total plant transpiration (Ep) assuming a “big leaf” model structure. It is well known that big leaf scaling approaches are fairly limited, especially within complex canopies, given that leaf–gas exchange is heterogeneous within canopies and varies as a function of factors such as leaf position or age [21], and this variation could also be under genetic control. These problems would have been minimized within our design because the plants were 3-months old and little time had elapsed for complexity in the canopy structure to be developed. Total plant leaf area was determined during plant harvest during the week following measurements using a leaf area meter (LI-3100C, Li-Cor Inc., Lincoln, NE, USA).
We first examined statistical patterns in gn, gd, Anet, and Ep during the treatments with linear mixed models that included sampling time, CO2 concentrations, genotype, and their interactions as fixed variables, and with glasshouse bay nested within heat-wave treatment as random variables. We then correlated gn with gd, Anet, and Ep to infer the potential for causal relationships among variables. These analyses were performed after examining whether the data conformed to assumptions of homoscedasticity and normality. All analyses were performed in the R software environment using base packages and lme4 [22].
3. Results
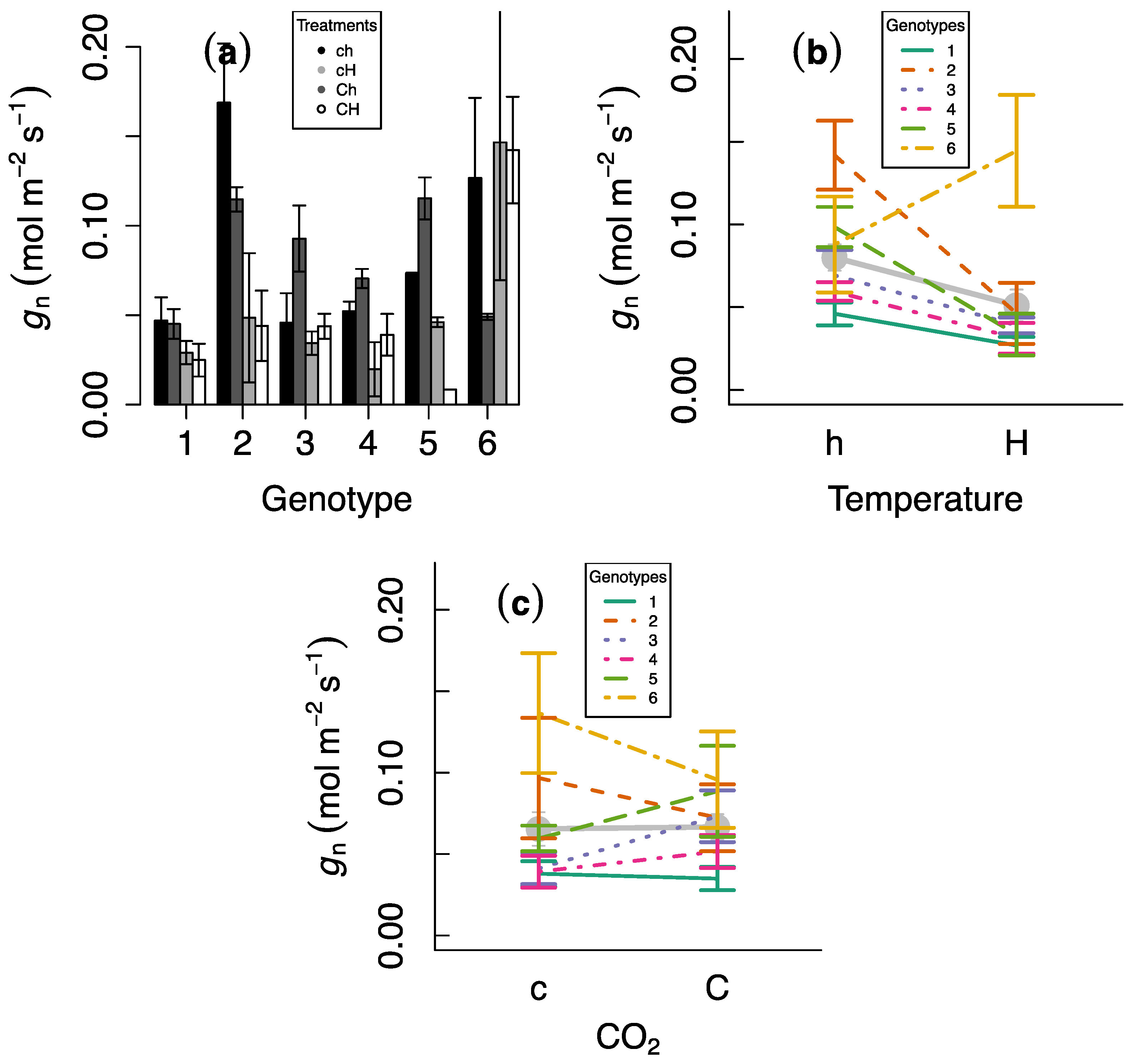
We observed significant differences in gn as a function of whether or not plants had been exposed to a heat wave on the previous days, indicating the existence of carry-over effects (Table 1). The main effect was a significant decline in gn after the heat wave of 36.1%, but responses varied across genotypes and gn increased in one genotype (Figure 1). There were no effects of CO2 on gn.
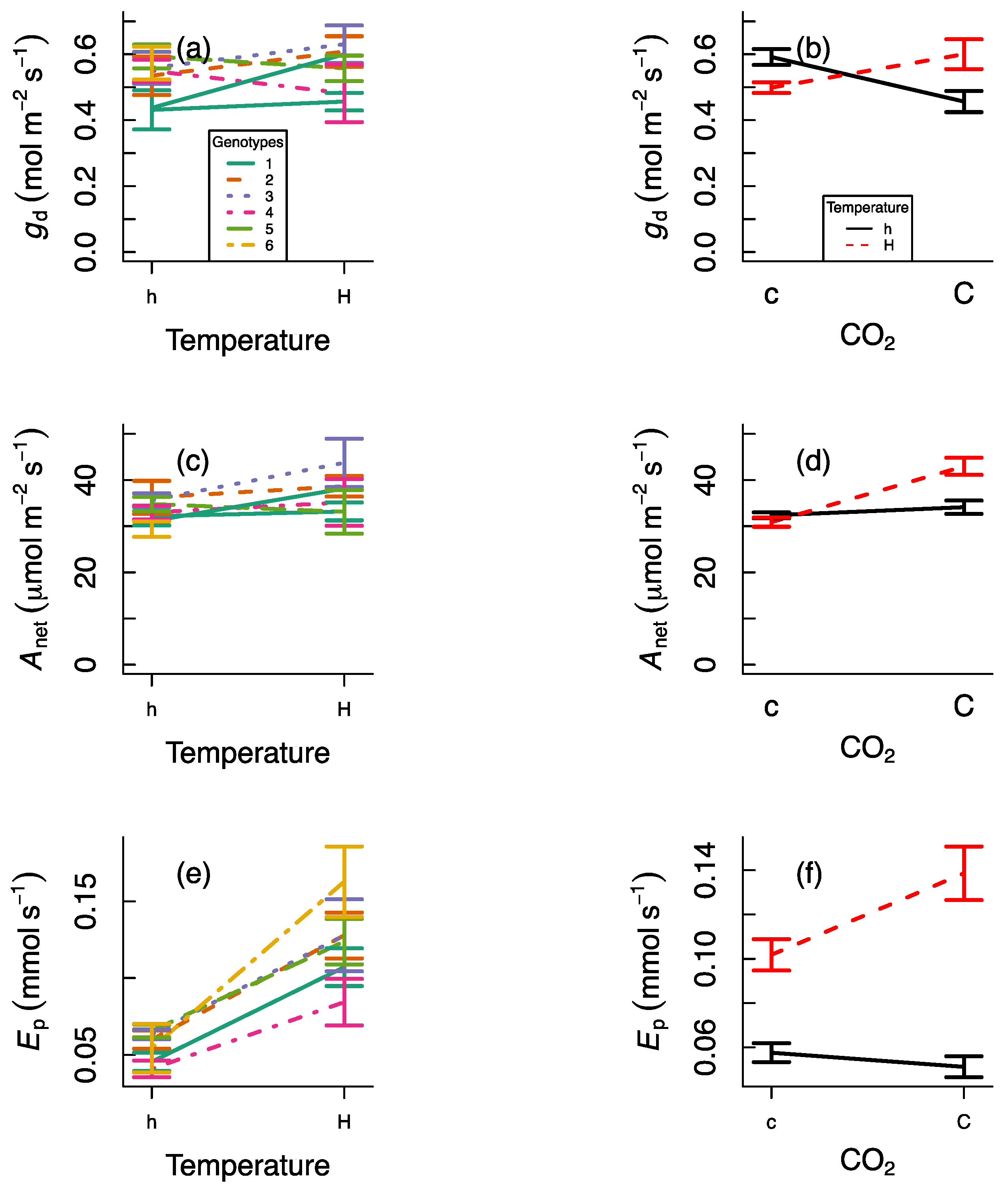
The response of daytime stomatal conductance and CO2 assimilation was also affected by the experimental treatments, although the effects were different for each of the traits (Table 1). gd varied significantly across genotypes (Figure 2a) and there was a significant CO2 × heat-wave interaction such that, during the heat wave, gd was higher in plants under elevated CO2 but lower under ambient CO2 (Figure 2b). Anet also varied significantly across genotypes (Figure 2c), and although there was also a CO2 × heat-wave interaction, in this case, there were no significant differences as a function of heat-wave exposure under ambient CO2, whereas there was a photosynthetic stimulation during the heat wave under elevated CO2 (Figure 2d). We also observed significant differences in Ep across genotypes (Figure 2e) and there was also a significant CO2 × heat-wave interaction. Ep was always higher during the heat wave, but Ep increased with elevated CO2 during the heat wave and it decreased with elevated CO2 under ambient temperature. Ep will be affected by transpiration rate and also by leaf area. Although different genotypes showed differences in leaf area and CO2 increased leaf area, there were no differences in leaf area as a function of heat wave exposure, as expected (Figure A1).
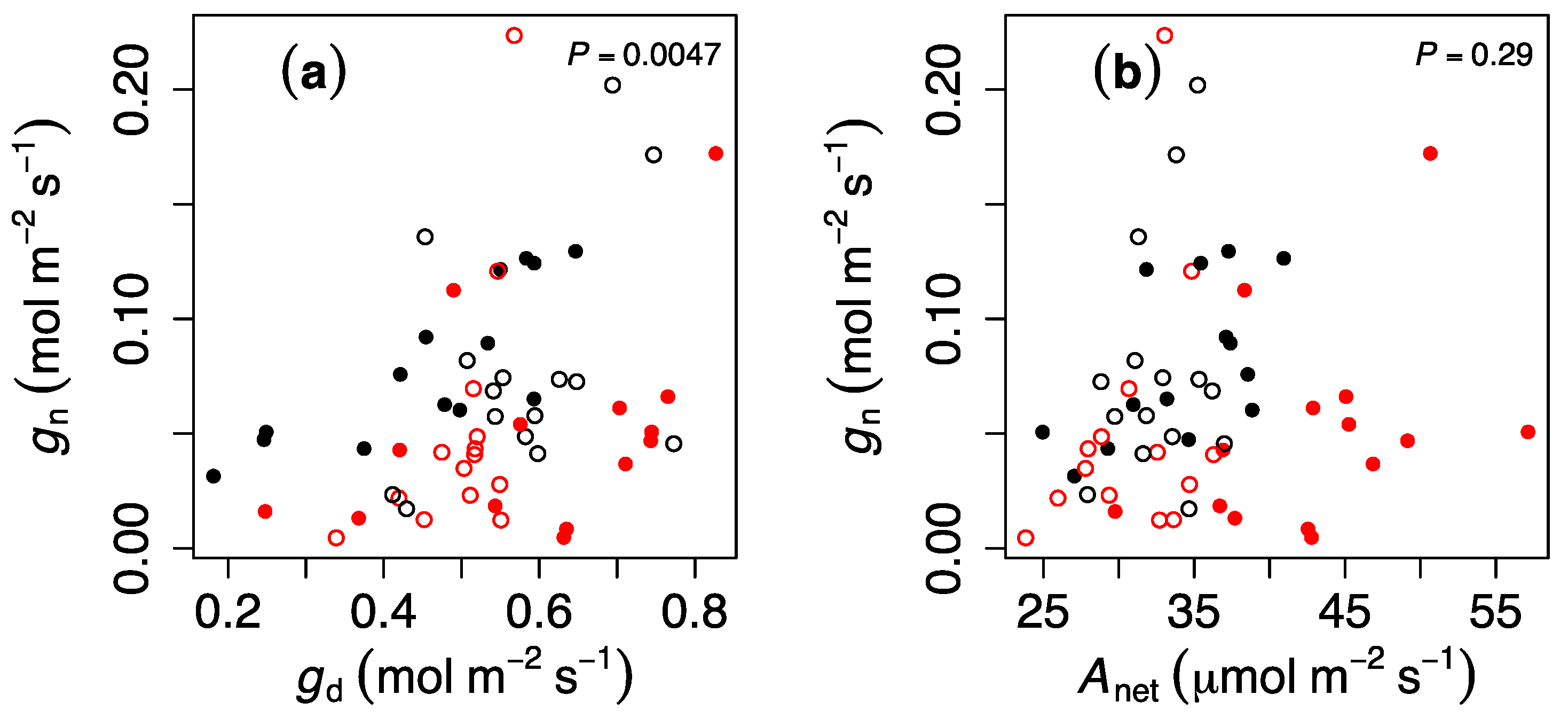
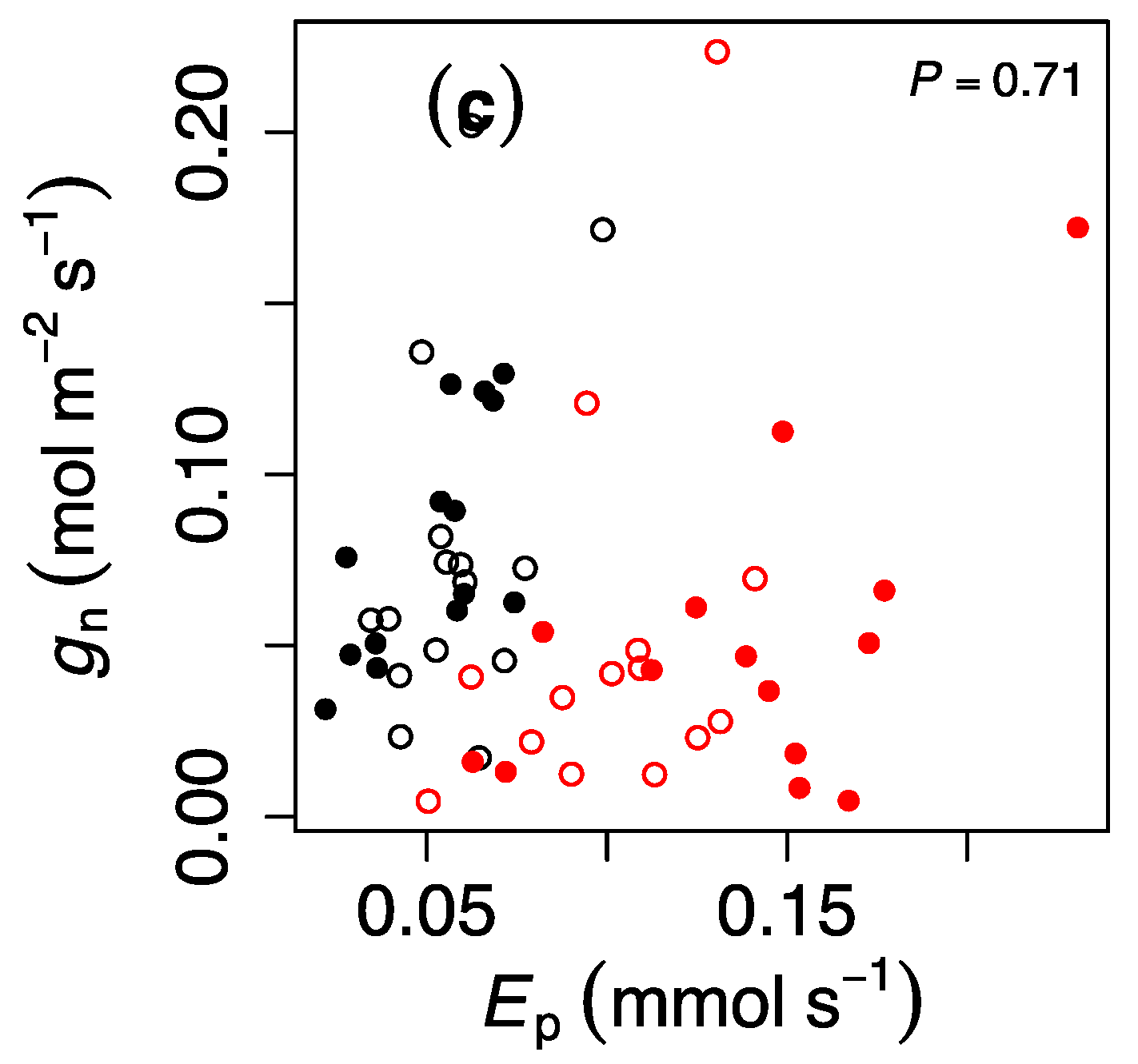
To understand the possible mechanisms explaining the pattern in gn, we correlated this parameter against gd, Anet, and Ep and only observed a significant and positive correlation between gn and gd (P = 0.0047, Figure 3), although the regression had limited explanatory power (R2 = 0.13).
4. Discussion
We observed that antecedent daytime environmental conditions affect nocturnal stomatal conductance. The response was largely consistent (but not uniform) across genotypes, but none of the previously hypothesized processes fully explain the response.
Current stomatal models assume nearly immediate response to environmental conditions and that antecedent environmental conditions exert no effect on stomata [23]. However, Resco de Dios et al. [24] proposed that stomatal models should consider circadian effects because the clock controlled up to 70% of the diurnal pattern in stomatal conductance in two crop species. They argued that circadian regulation acts as a “memory” such that the plant may anticipate predictable environmental changes (e.g., dawn/dusk transitions) based upon information from past environmental conditions. Here, we demonstrate that antecedent effects are an additional process whereby past environmental conditions may affect nocturnal stomatal behavior.
A crucial challenge will be to identify which daytime environmental processes affect the nocturnal stomatal response. Here, we observed that CO2 effects, which significantly affected gd, Anet, and Ep, did not have an impact on gn. We had previously identified the lack of an effect of elevated CO2 on gn in this species [19], but we did expect that gn would be affected by a CO2 × heat-wave interaction because elevated CO2 significantly affected gd responses.
We increased the temperature by 10 °C over four days, which also increased the vapor pressure deficit, and observed significant reductions in gn, averaged across genotypes, of 36% in the night after the heat wave (grey values in Figure 1b). This may be the first report of heat waves impacting gn in an angiosperm. Previous work on gn examined heat-wave responses in a conifer species [14] and in hardwoods, but only sap flux responses were assessed [25].
Why did we observe that gn was affected by antecedent heat waves but not by antecedent CO2? While further studies will be necessary to fully decipher this question, here we can offer some hypotheses. First, it should be noted that CO2 treatments were applied for the entire growing period, but the heat wave was imposed for only a few days and, consequently, the effect of these two environmental drivers is not fully comparable. Additionally, it could be argued that only drastically different environmental conditions may lead to carry-over effects. For instance, Easlon and Richards [12] observed that low daytime CO2 decreased gn, but in their experimental design, they completely scrubbed CO2 from the air to 0 ppm and, consequently, leaves may have been carbon starved when the night arrived. Others have similarly found that sucrose limitations explain variation in gn in some temperate trees [26]. Our CO2 treatments (400 and 640 ppm) would not have led to such dramatic effects and carbohydrate concentrations were generally not limiting [27]. By contrast, during our heat wave, we raised the temperatures by 10 °C, and this had important consequences for plant transpiration.
Duarte et al. [14] hypothesized (but did not measure) that changes in anatomical properties, such as stomatal occlusion or changes in epicuticular wax, could have occurred during the heat wave, and that led to carry-over effects in Douglas fir. A corollary from this hypothesis is that daytime and nighttime conductance should be proportionally reduced because physical changes in occlusion, for instance, would similarly affect daytime and nighttime conductance. Consistent with this hypothesis, we observed a significant correlation between gn and gd (Figure 3a), but it only explained 13% of the variation in gn, indicating that additional processes must be playing a role. However, a role for cuticular conductance here remains dubious since previous studies in Eucalypts report values of 0.003 mol m−2 s−1 [28], a value that is between 1 and 2 orders of magnitude smaller than the values of gn reported here.
Our findings are also not consistent with the notion that heat waves exert carry-over effects due to increased xylem hydraulic recharge because there was no correlation between gn and Ep. We note, however, that this hypothesis is not exempt from criticism. In fact, it could also be argued that stem recharge would actually benefit from stomatal closure (not opening), as transpiration lowers the leaf-level water potential and, consequently, breaks the equilibrium with soil water potential that favors recharge [29].
Previous studies had documented genetic variability in gn in a range of species [19,30,31], but we demonstrate, for the first time, genetic variability in gn responses to heat waves (Figure 1b). Eucalyptus camaldulensis is a riparian species, with deep roots in mature trees, that does not usually suffer from drought in its native range. Variability in gn may thus be advantageous because genotypes with higher gn may assimilate more C in the morning [19], with minimal danger of soil water scarcity in deep-rooted trees in riparian environments.
Genetic variability in the response to heat waves could be related to the climate of origin [20]. We did not perform detailed analyses based on climate of origin because we only had one genotype from each location. However, we note that the genotype that responded positively in gn to the heat wave (Genotype 6) originated from one of the locations with the highest mean annual maximum temperature (Table A1). If we assume that heat waves reach higher temperatures at locations with a higher maximum temperature, we could hypothesize that there is a higher capacity of gn to respond to heat waves in genotypes that experience more acute heat waves in their local environment. Of course, more data would be necessary to confirm this hypothesis.
Heat waves in nature often co-occur with water scarcity. Here, we kept any potential effect of soil moisture deficit at a minimum by diurnal irrigation to field capacity. We chose this approach to be able to disentangle heat-wave effects. However, soil water scarcity may exacerbate heat-wave effects and potentially increase differences in the response across genotypes. Further studies on gn should deepen on the role of multiple stressors acting concomitantly to modify the intraspecific responses.
An important open question is how to implement the impact of antecedent conditions into stomatal models. Statistical approaches have been developed to incorporate antecedent effects and to identify the relevant temporal scales [32]. These models rely on continued observations for prolonged periods of time, which is one of the major gaps in nocturnal conductance research, as most observations are available for just one or a few nights. Models incorporating the effect of antecedent conditions are empirical and, consequently, they would need to be coupled with further mechanistic studies that would allow separation of causal from casual correlations. In this experiment, we documented a significant impact of antecedent heat waves on gn and we were able to explain a significant but relatively small portion of the variance. Overall, we propose that long-term monitoring of gn and detailed mechanistic experiments combined with models for antecedent conditions should be at the forefront of our research efforts.
5. Conclusions
Studies on nocturnal stomatal conductance should routinely assess the role of antecedent environmental conditions, particularly those with extreme climate events such as heat waves. We observed carry-over effects from heat waves on gn that could be partly explained by changes in gd, but additional studies will be required to more fully elucidate the mechanisms. Genotypic variation in gn response to heat waves was also observed, suggesting local adaptation and potential importance in climate change scenarios where heat waves become more common.
Author Contributions
Conceptualization, V.R.D., M.E.L. and D.T.T.; Methodology, V.R.D., M.E.L., R.A.S. and D.T.T.; Formal Analysis, V.R.D.; Investigation, V.R.D., M.E.L., R.A.S. and D.T.T.; Writing-Original Draft Preparation, V.R.D., M.E.L., R.A.S. and D.T.T..; Writing-Review & Editing, V.R.D., M.E.L., R.A.S. and D.T.T.; Funding Acquisition, D.T.T.
Funding
This study was funded by a Science Industry Endowment Fund (project code RP04–122) and supported by the Hawkesbury Institute for the Environment, Western Sydney University. VRD was partly funded from a Ramón y Cajal Fellowship (RYC-2012–10970).
Acknowledgments
The manuscript benefitted from the comments of two anonymous reviewers.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table A1.
Climate of origin for the different genotypes used in this study. For each location, we provide mean annual precipitation (MAP, mm), mean annual air temperature (MAT, °C), mean annual maximum air temperature (MXT, °C), and the mean annual temperature range (ATR, °C) from meteorological station data for the years 1971–2010.
Table A1.
Climate of origin for the different genotypes used in this study. For each location, we provide mean annual precipitation (MAP, mm), mean annual air temperature (MAT, °C), mean annual maximum air temperature (MXT, °C), and the mean annual temperature range (ATR, °C) from meteorological station data for the years 1971–2010.
| Genotype | Location | Latitude | Longitude | MAP (mm) | MAT (°C) | MXT (°C) | ATR (°C) |
|---|---|---|---|---|---|---|---|
| 1 | YASS RIVER | 34.53 | 149.02 | 675 | 14.0 | 28.4 | 25.8 |
| 2 | OVENS VALLEY | 36.36 | 146.47 | 653 | 15.0 | 30.5 | 26.8 |
| 3 | COONAWARRAW | 37.2 | 140.42 | 646 | 14.5 | 30.0 | 24.0 |
| 4 | NYNGAN | 31.33 | 147.11 | 481 | 19.2 | 34.2 | 27.6 |
| 5 | CONDOBOLIN | 33.06 | 147.09 | 459 | 17.6 | 33.9 | 28.9 |
| 6 | BARMAH SF | 35.5 | 145.07 | 403 | 16.4 | 33.0 | 28.2 |
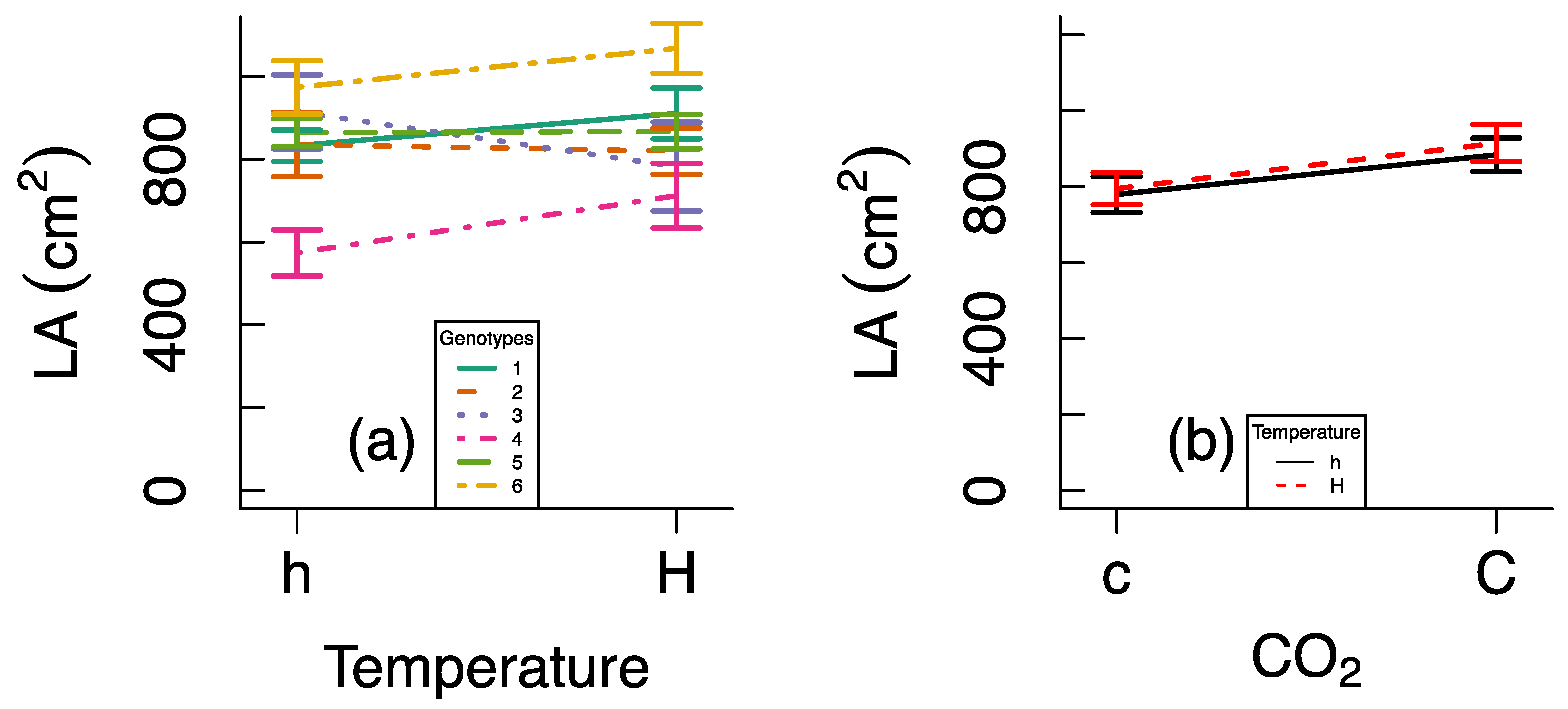
Figure A1.
Variation in leaf area (LA) across genotypes, heat waves, and CO2 concentrations. Data are presented as reaction norms for six genotypes as a function of temperature (a) and as reaction norms for temperature as a function of CO2 treatment (b). Error bars indicate standard errors.
Figure A1.
Variation in leaf area (LA) across genotypes, heat waves, and CO2 concentrations. Data are presented as reaction norms for six genotypes as a function of temperature (a) and as reaction norms for temperature as a function of CO2 treatment (b). Error bars indicate standard errors.

References
- Wang, K.; Dickinson, R.E. A review of global terrestrial evapotranspiration: Observation, modeling, climatology, and climatic variability. Rev. Geophys. 2012, 50, RG2005. [Google Scholar] [CrossRef]
- Forster, M.A. How significant is nocturnal sap flow? Tree Physiol. 2014, 34, 757–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lombardozzi, D.L.; Zeppel, M.J.B.; Fisher, R.A.; Tawfik, A. Representing nighttime and minimum conductance in clm4.5: Global hydrology and carbon sensitivity analysis using observational constraints. Geosci. Model Dev. 2017, 10, 321–331. [Google Scholar] [CrossRef]
- Cowan, I.R.; Farquhar, G.D. Stomatal function in relation to leaf metabolism and environment. In Integration of Activity in the Higher Plant; Jennings, D.H., Ed.; Cambridge University Press: Cambridge, UK, 1977. [Google Scholar]
- Caird, M.A.; Richards, J.H.; Donovan, L.A. Nighttime stomatal conductance and transpiration in C3 and C4 plants. Plant Physiol. 2007, 143, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Zeppel, M.J.; Lewis, J.D.; Phillips, N.G.; Tissue, D.T. Consequences of nocturnal water loss: A synthesis of regulating factors and implications for capacitance, embolism and use in models. Tree Physiol. 2014, 34, 1047–1055. [Google Scholar] [CrossRef] [PubMed]
- Musselman, R.C.; Minnick, T.J. Nocturnal stomatal conductance and ambient air quality standards for ozone. Atm Env. 2000, 34, 719–733. [Google Scholar] [CrossRef]
- Ogle, K.; Lucas, R.W.; Bentley, L.P.; Cable, J.M.; Barron-Gafford, G.A.; Griffith, A.; Ignace, D.; Jenerette, G.D.; Tyler, A.; Huxman, T.E.; et al. Differential daytime and night-time stomatal behavior in plants from North American deserts. New Phytol. 2012, 194, 464–476. [Google Scholar] [CrossRef] [PubMed]
- Zeppel, M.J.B.; Lewis, J.D.; Chaszar, B.; Smith, R.A.; Medlyn, B.E.; Huxman, T.E.; Tissue, D.T. Nocturnal stomatal conductance responses to rising [CO2], temperature and drought. New Phytol. 2012, 193, 929–938. [Google Scholar] [CrossRef] [PubMed]
- Barbour, M.M.; Cernusak, L.A.; Whitehead, D.; Griffin, K.; Turnbull, M.; Tissue, D.T.; Farquhar, G.D. Nocturnal stomatal conductance and implications for modelling δ 18O of leaf-respired CO2 in temperate tree species. Funct. Plant Biol. 2005, 32, 1107–1121. [Google Scholar] [CrossRef]
- Resco de Dios, V.; Diaz-Sierra, R.; Goulden, M.L.; Barton, C.V.; Boer, M.M.; Gessler, A.; Ferrio, J.P.; Pfautsch, S.; Tissue, D.T. Woody clockworks: Circadian regulation of night-time water use in eucalyptus globulus. New Phytol. 2013, 200, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Easlon, H.M.; Richards, J.H. Photosynthesis affects following night leaf conductance in vicia faba. Plant Cell Environ. 2009, 32, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Resco de Dios, V.; Roy, J.; Ferrio, J.P.; Alday, J.G.; Landais, D.; Milcu, A.; Gessler, A. Processes driving nocturnal transpiration and implications for estimating land evapotranspiration. Sci. Rep. 2015, 5, 10975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duarte, A.G.; Katata, G.; Hoshika, Y.; Hossain, M.; Kreuzwieser, J.; Arneth, A.; Ruehr, N.K. Immediate and potential long-term effects of consecutive heat waves on the photosynthetic performance and water balance in douglas-fir. J. Plant Physiol. 2016, 205, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Barbour, M.M.; Buckley, T.N. The stomatal response to evaporative demand persists at night in ricinus communis plants with high nocturnal conductance. Plant Cell Environ. 2007, 30, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Field, C.B.; Barros, V.; Stocker, T.F.; Dahe, Q.; Dokken, D.J.; Ebi, K.L.; Mastrandrea, M.D.; Mach, K.J.; Plattner, G.-K.; Allen, S.K.; et al. Managing the Risks of Extreme Events and Disasters to Advance Climate Change Adaptation—Special Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2012; p. 582. [Google Scholar]
- De Dios, V.R.; Loik, M.E.; Smith, R.A.; Aspinwall, M.J.; Tissue, D.T. Genetic variation in circadian regulation of nocturnal stomatal conductance enhances plant fitness. Plant Cell Environ. 2016, 39, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Milcu, A.; Puga-Freitas, R.; Ellison, A.M.; Blouin, M.; Scheu, S.; Freschet, G.T.; Rose, L.; Barot, S.; Cesarz, S.; Eisenhauer, N.; et al. Genotypic variability enhances the reproducibility of an ecological study. Nat. Ecol. Evol. 2018, 2, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Jones, H. Plants and Microclimate: A Quantitative Approach to Environmental Plant Physiology, 3rd ed.; Cambridge University Press: Cambridge, UK, 2014; p. 423. [Google Scholar]
- Loik, M.E.; Resco de Dios, V.; Smith, R.; Tissue, D.T. Relationships between climate of origin and photosynthetic responses to an episodic heat wave depend on growth CO2 concentration for eucalyptus camaldulensis var. Camaldulensis. Funct. Plant Biol. 2017, 44, 1053–1066. [Google Scholar] [CrossRef]
- De Pury, D.G.G.; Farquhar, G.D. Simple scaling of photosynthesis from leaves to canopies without the errors of big-leaf models. Plant Cell Environ. 1997, 20, 537–557. [Google Scholar] [CrossRef] [Green Version]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Lme4: Linear mixed-effects models using eigen and s4. R package version 1.1-7. 2014. Available online: Http://cran.R-project.Org/package=lme4 (accessed on 20 February 2018).
- Buckley, T.N. Modeling stomatal conductance. Plant Physiol. 2017, 174, 572–582. [Google Scholar] [CrossRef] [PubMed]
- De Dios, V.R.; Gessler, A.; Ferrio, J.P.; Alday, J.G.; Bahn, M.; del Castillo, J.; Devidal, S.; García-Muñoz, S.; Kaylerd, Z.; Landais, D.; et al. Circadian rhythms have significant effects on leaf-to-canopy gas exchange under field conditions. GigaScience 2016, 5, 43. [Google Scholar] [CrossRef] [PubMed]
- Pfautsch, S.; Adams, M.A. Water flux of eucalyptus regnans: Defying summer drought and a record heat wave in 2009. Oecologia 2013, 172, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Kupper, P.; Ivanova, H.; Sõber, A.; Rohula-Okunev, G.; Sellin, A. Night and daytime water relations in five fast-growing tree species: Effects of environmental and endogenous variables. Ecohydrology 2017, e1927. [Google Scholar] [CrossRef]
- Blackman, C.J.; Aspinwall, M.J.; Resco de Dios, V.; Smith, R.; Tissue, D.T. Leaf photosynthetic, economics and hydraulic traits are decoupled among genotypes of a widespread species of eucalypt grown under ambient and elevated CO2. Funct. Ecol. 2016, 30, 1491–1500. [Google Scholar] [CrossRef]
- Phillips, N.G.; Lewis, J.D.; Logan, B.A.; Tissue, D.T. Inter- and intra-specific variation in nocturnal water transport in eucalyptus. Tree Physiol. 2010, 30, 586–596. [Google Scholar] [CrossRef] [PubMed]
- Bucci, S.J.; Scholz, F.G.; Goldstein, G.; Meinzer, F.C.; Hinojosa, J.A.; Hoffmann, W.A.; Franco, A.C. Processes preventing nocturnal equilibration between leaf and soil water potential in tropical savanna woody species. Tree Physiol. 2004, 24, 1119–1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoppach, R.; Claverie, E.; Sadok, W. Genotype-dependent influence of night-time vapour pressure deficit on night-time transpiration and daytime gas exchange in wheat. Funct. Plant Biol. 2014, 41, 963–971. [Google Scholar] [CrossRef]
- Christman, M.A.; Richards, J.H.; McKay, J.K.; Stahl, E.A.; Juenger, T.E.; Donovan, L.A. Genetic variation in arabidopsis thaliana for night-time leaf conductance. Plant Cell Environ. 2008, 31, 1170–1178. [Google Scholar] [CrossRef] [PubMed]
- Ogle, K.; Barber, J.J.; Barron-Gafford, G.A.; Bentley, L.P.; Young, J.M.; Huxman, T.E.; Loik, M.E.; Tissue, D.T. Quantifying ecological memory in plant and ecosystem processes. Ecol. Lett. 2015, 18, 221–235. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Experimental responses of nocturnal stomatal conductance. (a) Variation across genotypes as a function of heat-wave exposure and CO2 concentrations. (b) Reaction norms of heat-wave responses across genotypes. (c) Reaction norms of CO2 responses across genotypes. Grey in (b,c) indicates average values for all genotypes and the other colors indicate individual genotypes. h, H, c and C indicate no heat-wave exposure, heat-wave exposure, ambient CO2, and elevated CO2, respectively. Error bars indicate standard errors.
Figure 1.
Experimental responses of nocturnal stomatal conductance. (a) Variation across genotypes as a function of heat-wave exposure and CO2 concentrations. (b) Reaction norms of heat-wave responses across genotypes. (c) Reaction norms of CO2 responses across genotypes. Grey in (b,c) indicates average values for all genotypes and the other colors indicate individual genotypes. h, H, c and C indicate no heat-wave exposure, heat-wave exposure, ambient CO2, and elevated CO2, respectively. Error bars indicate standard errors.

Figure 2.
Experimental responses of daytime stomatal conductance (gd; a,b), net photosynthesis (Anet; c,d), and whole plant transpiration (Ep; e,f) in response to no heat-wave exposure (h), heat-wave exposure (H), ambient CO2 (c), and elevated CO2 (C). Data are presented as reaction norms for six genotypes as a function of temperature (figures a,c,e) and as reaction norms for temperature as a function of CO2 treatment (figures b,d,f). Error bars indicate standard errors.
Figure 2.
Experimental responses of daytime stomatal conductance (gd; a,b), net photosynthesis (Anet; c,d), and whole plant transpiration (Ep; e,f) in response to no heat-wave exposure (h), heat-wave exposure (H), ambient CO2 (c), and elevated CO2 (C). Data are presented as reaction norms for six genotypes as a function of temperature (figures a,c,e) and as reaction norms for temperature as a function of CO2 treatment (figures b,d,f). Error bars indicate standard errors.

Figure 3.
Relationship between nocturnal stomatal conductance (gn) and (a) daytime stomatal conductance (gd,), (b) net assimilation (Anet,), and (c) whole plant transpiration (Ep). Black and red values indicate ambient temperature and heat wave, respectively, and open and closed symbols indicate ambient and elevated CO2 concentrations, respectively. P-values indicate the result of least-squares fitting. Each value represents a single genotype.
Figure 3.
Relationship between nocturnal stomatal conductance (gn) and (a) daytime stomatal conductance (gd,), (b) net assimilation (Anet,), and (c) whole plant transpiration (Ep). Black and red values indicate ambient temperature and heat wave, respectively, and open and closed symbols indicate ambient and elevated CO2 concentrations, respectively. P-values indicate the result of least-squares fitting. Each value represents a single genotype.


Table 1.
Results of linear mixed models on the effects of CO2, genotype, heat wave, and their interactions on nocturnal stomatal conductance (gn), daytime stomatal conductance (gd), leaf assimilation rates (Anet), and whole plant transpiration (Ep), with glasshouse bay nested within CO2 concentration as random variables. P-values reflect the results of Wald tests on linear mixed model that included glasshouse bay nested within heat-wave treatment as random variables. Significant (P < 0.05) values are in bold text.
Table 1.
Results of linear mixed models on the effects of CO2, genotype, heat wave, and their interactions on nocturnal stomatal conductance (gn), daytime stomatal conductance (gd), leaf assimilation rates (Anet), and whole plant transpiration (Ep), with glasshouse bay nested within CO2 concentration as random variables. P-values reflect the results of Wald tests on linear mixed model that included glasshouse bay nested within heat-wave treatment as random variables. Significant (P < 0.05) values are in bold text.
| gn | gd | Anet | Ep | |||||
|---|---|---|---|---|---|---|---|---|
| χ2 | P | χ2 | P | χ2 | P | χ2 | P | |
| CO2 | 0.001 | 0.99 | 0.09 | 0.75 | 27.42 | <0.001 | 1.560 | 0.21 |
| Genotype | 41.01 | <0.001 | 19.83 | 0.003 | 17.92 | 0.006 | 30.72 | <0.001 |
| Heat wave | 11.85 | <0.001 | 1.66 | 0.20 | 6.23 | 0.01 | 116.88 | <0.001 |
| C × G | 8.67 | 0.12 | 7.68 | 0.26 | 4.99 | 0.54 | 4.33 | 0.50 |
| C × H | 0.07 | 0.79 | 13.45 | <0.001 | 17.02 | <0.001 | 16.97 | <0.001 |
| G × H | 28.63 | <0.001 | 6.45 | 0.26 | 6.04 | 0.30 | 10.41 | 0.06 |
| C × G × H | 6.99 | 0.22 | 4.63 | 0.46 | 2.39 | 0.79 | 5.44 | 0.56 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Resco de Dios, V.; Loik, M.E.; Smith, R.A.; Tissue, D.T. Effects of a Heat Wave on Nocturnal Stomatal Conductance in Eucalyptus camaldulensis. Forests 2018, 9, 319. https://doi.org/10.3390/f9060319
AMA Style
Resco de Dios V, Loik ME, Smith RA, Tissue DT. Effects of a Heat Wave on Nocturnal Stomatal Conductance in Eucalyptus camaldulensis. Forests. 2018; 9(6):319. https://doi.org/10.3390/f9060319
Chicago/Turabian StyleResco de Dios, Víctor, Michael E. Loik, Renee A. Smith, and David T. Tissue. 2018. "Effects of a Heat Wave on Nocturnal Stomatal Conductance in Eucalyptus camaldulensis" Forests 9, no. 6: 319. https://doi.org/10.3390/f9060319
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.