Charcoal Increases Microbial Activity in Eastern Sierra Nevada Forest Soils
by
Zachary W. Carter
1, 
Benjamin W. Sullivan
1,
Robert G. Qualls
1,
Robert R. Blank
2,
Casey A. Schmidt
3 and
Paul S.J. Verburg
1,* 1
Department of Natural Resources and Environmental Science, University of Nevada, Reno, NV 89557, USA
2
Agricultural Research Service, United States Department of Agriculture, Reno, NV 89512, USA
3
Division of Hydrologic Sciences, Desert Research Institute, Reno, NV 89512, USA
*
Author to whom correspondence should be addressed.
Forests 2018, 9(2), 93; https://doi.org/10.3390/f9020093
Submission received: 10 January 2018
/
Revised: 7 February 2018
/
Accepted: 14 February 2018
/
Published: 16 February 2018
(This article belongs to the Special Issue Carbon, Nitrogen and Phosphorus Cycling in Forest Soils)
Abstract
:Fire is an important component of forests in the western United States. Not only are forests subjected to wildfires, but fire is also an important management tool to reduce fuels loads. Charcoal, a product of fire, can have major impacts on carbon (C) and nitrogen (N) cycling in forest soils, but it is unclear how these effects vary by dominant vegetation. In this study, soils collected from Jeffrey pine (JP) or lodgepole pine (LP) dominated areas and amended with charcoal derived from JP or LP were incubated to assess the importance of charcoal on microbial respiration and potential nitrification. In addition, polyphenol sorption was measured in unamended and charcoal-amended soils. In general, microbial respiration was highest at the 1% and 2.5% charcoal additions, but charcoal amendment had limited effects on potential nitrification rates throughout the incubation. Microbial respiration rates decreased but potential nitrification rates increased over time across most treatments. Increased microbial respiration may have been caused by priming of native organic matter rather than the decomposition of charcoal itself. Charcoal had a larger stimulatory effect on microbial respiration in LP soils than JP soils. Charcoal type had little effect on microbial processes, but polyphenol sorption was higher on LP-derived than JP-derived charcoal at higher amendment levels despite surface area being similar for both charcoal types. The results from our study suggest that the presence of charcoal can increase microbial activity in soils, but the exact mechanisms are still unclear.
1. Introduction
Wildfire is an important component of semi-arid forests of the western United States. Fire suppression and prevention during the last century have, however, contributed to increases in stand density, the number of small trees, the amount of understory shrub vegetation, and accumulation of forest floor, thereby increasing the risk of severe, stand-replacing wildfires [1]. Fire is also employed to reduce the risk of wildfire through broadcast burning or disposal of fuels following mechanical thinning through pile burning [2,3]. Fire typically reduces soil organic carbon (C) and nitrogen (N) pools due to volatilization of organic matter [4], thereby decreasing available substrate for primary decomposers. Bioavailable forms of soil N, including ammonium (NH4+) and nitrate (NO3−), can, however, increase due to chemical mineralization of organic material, deposition of ash, and lysis of soil microorganisms [5]. Elevated NH4+ levels can be sustained through subsequent mineralization of N. Following increases in NH4+, nitrification can be stimulated, resulting in an increase in soil NO3− [6]. Microbial biomass typically decreases after a fire because the transfer of heat into soils is large enough to kill most organisms in the top 5 to 10 cm [3,7]. The direct effects of fire on soil chemical and microbial processes depend on fire severity, with impacts being large in high-severity wildfires or pile burns and much less pronounced in low-severity broadcast burns [8,9]. Indirect effects of fire on microbial communities can be mediated through vegetation recovery following fire, with plant colonization being important in shaping post-fire microbial community structure in high-severity fires compared to low-severity fires [10].
In addition to vegetation affecting post-fire microbial processes, the presence of charcoal formed due to the incomplete combustion of woody materials may affect microbial processes following a fire [11,12,13,14]. Charcoal is highly resistant to decay and remains chemically active for up to a century [14,15]. Charcoal may affect microbial processes through (1) an increase in soil pH affecting microbial community structure; (2) adsorption of C compounds such as polyphenols that can inhibit mineralization and nitrification; (3) adsorption of labile C compounds that can act as substrate for microbial communities; and (4) an increase in water and nutrient availability [12,13,16,17,18]. In addition, microorganisms may enter pore spaces smaller than 20 μm in charcoal, thus avoiding predation by larger micro-arthropods [11,13,14,17].
The biochemical composition of the vegetation from which charcoal is derived constrains its final physical and chemical properties and thus the ability of charcoal to affect microbial processes [19,20]. Previous studies used a range of pyrogenic materials including field-collected charcoal [21], activated carbon [22], grasses [12], or biochar feedstock [11]. We are aware of only one study that compared charcoal derived from different tree species. This study found that charcoal derived from ponderosa pine (Pinus ponderosa Lawson & C. Lawson) had a higher sorption capacity for allelopathic substances than charcoal derived from Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco) due to a smaller tracheid diameter of the wood in ponderosa pine [23]. Despite these differences in chemistry, net nitrification rates were similar between charcoal types [23]. Other studies have shown, however, that chemical composition of charcoal can affect enzyme activity [15], thereby potentially affecting C and N fluxes in soils.
The objective of our study was to assess the response of soil microbes to charcoal that formed during wood combustion in semi-arid forests. We hypothesized that (1) microbial respiration and nitrification increases with increasing amounts of charcoal due to the increased sorption of labile C and nitrification-inhibiting compounds including polyphenols to the surface of charcoal particles, and (2) the effects of charcoal amendment on microbial activity depend on the source of charcoal, with microbial activity being greater in soils amended with charcoal that has greater surface area, porosity, and adsorption capacity.
To test our hypotheses, charcoal was derived from Jeffrey pine (Pinus jeffreyi Balf.) and lodgepole pine (Pinus contorta Douglas ex Loudon var. murrayana (Balf.) Engelm.), two tree species commonly found in semi-arid western forests. Charcoal was added to unburned soils collected from a site dominated by these two tree species and we measured physical and chemical properties of soils and charcoal, adsorption of polyphenolic substances, and microbial activity in amended and unamended soils. While our approach ignores the direct effects of fire on soils, this approach allowed us to separate effects of charcoal from other effects of fire on soil. In addition, our incubation conditions may represent low severity burns where direct impacts of fire on soils are small.
2. Materials and Methods
2.1. Field Site
Unburned soil and wood samples were collected from Little Valley, Nevada, approximately 30 km south of Reno, Nevada (39°14′23.76′′ N, 119°52′54.62′′ W). Little Valley is located in the Carson Range and the elevation ranges from 1900 to 2500 m. The overstory vegetation is dominated by Jeffrey pine and lodgepole pine. Two sites differing in overstory vegetation were selected adjacent to each other. Soils with a Jeffrey pine overstory were located on areas with a 10–15% slope and that had a sparse understory consisting of bitterbrush (Purshia tridentata (Pursh) DC.) and manzanita (Arctostaphylos patula Greene). Soils with a lodgepole pine overstory were located in an adjacent meadow on slopes of less than 5% and with an understory of mixed herbaceous species. Soils at both sites belonged to the Marla series (sandy, mixed, frigid Aquic Dystroxerepts; [24]). The climate at both sites is characterized by dry, warm summers and cold, wet winters. The mean annual air temperature is 5 °C and the mean annual precipitation is 87.5 cm with approximately half of the precipitation occurring as snow [25].
2.2. Soil Collection and Charcoal Production
We collected soil from the 0–10 cm mineral soil layer in ten locations at each site dominated by either Jeffrey pine (referred to as ‘JP soil’) or lodgepole pine (referred to as ‘LP soil’). Each sample contained approximately 1 kg of soil. All samples were composited by vegetation type. Prior to sampling, forest floor material was removed. After sampling, we air-dried, sieved (2 mm mesh size), and homogenized the samples. Root materials present in the mineral soil were discarded.
Branch material for charcoal production was collected from multiple slash piles present at the JP and LP sites. The branches were up to 2.5 cm in diameter. We produced charcoal following the procedure of Zackrisson et al. [14]. Branch material from JP and LP was allowed to oxidize at 450 °C for 15 min in a muffle furnace. Following this treatment, all material was charred. The charcoal was then gently crushed with a mortar and pestle and sieved using a 2 mm mesh sieve. Charcoal was mixed in with the soil.
We amended unburned JP and LP soils with charcoal using four amendment levels (0.5%, 1%, 2.5%, and 5% charcoal by weight), and two charcoal sources (JP and LP). In addition, we included unamended control soils resulting in a total of 20 soil/amendment combinations. The amendment levels were based on charcoal quantification in burn scars following pile burns. These analyses revealed that mineral soils in burn scars contained approximately 2.5% charcoal (Z. Carter, unpublished data). Unburned JP soils contained 0.03% charcoal while unburned LP soils contained 0.01% charcoal (Z. Carter, unpublished data).
2.3. Soil and Charcoal Characterization
Total %C and %N of soils and charcoal were quantified in triplicate using a Leco TruSpec CN analyzer after soil and charcoal types were ground for 48 h using a Mavco vial rotator. Soil pH of unamended and amended soils was determined in triplicate using a 1:2 soil to water (g g−1) ratio. Cation exchange capacity (CEC) was measured using the ammonium saturation method [26] at the Waters Agricultural Laboratories Inc., Camilla, GA, USA using a single CEC measurement for each soil and charcoal type. Surface area of charcoal was quantified using N2 adsorption isotherms at −196 °C at the United States Department of Agriculture-Agricultural Research Service Southern Regional Research Center using a Nova 2000 Surface Area Analyzer (Quantachrome Corp., Boynton Beach, FL, USA). Specific surface area, which included micropores >2 nm, was calculated from the adsorption isotherms using the Brunner-Emmett-Teller (BET) equation. Surface area was measured on crushed samples similar to what was added to the soil. Surface area measurements were performed in duplicate.
2.4. Polyphenol Adsorption
To determine the effects of charcoal on polyphenol adsorption, we extracted unamended JP and LP soils, JP soils amended with JP-derived charcoal, and LP soils amended with LP-derived charcoal with deionized water. We used the same treatment combinations used for the potential nitrification measurements (see below). Three replicates were used for each treatment (n = 3), resulting in a total of 30 samples. Ten grams of soil and 30 mL of deionized water were added to 50 mL centrifuge tubes, shaken for 24 h using an overhead shaker, centrifuged at 2500 rpm, decanted, and filtered using a 0.45 µm nylon fiber filter. Polyphenol concentration in the aqueous extract was measured using the Folin-Ciocalteu reagent [27]. Absorbance was measured using a Shimadzu spectrophotometer (Shimadzu, Columbia, MD, USA) at 760 nm wavelength in quartz cuvettes and compared to tannic acid standards.
2.5. Microbial Respiration
To quantify microbial respiration, 250 mL Pyrex media storage bottles were filled with 25 g of unamended or charcoal-amended soils and deionized water was added until the soil moisture level was at 60% of field capacity by weight similar to Kolb et al. [11] and DeLuca et al. [21]. For the incubation, JP and LP soils were amended with both JP and LP charcoal, resulting in a total of 20 treatment combinations. Soil moisture at field capacity was determined by saturating the unamended and amended soils and letting water drain for 24–48 h. Moist samples were weighed prior to and after oven-drying at 105 °C until a constant mass was reached. At the start of the incubation, we weighed and capped the incubation bottles, and bottles were stored in a dark incubator at 25 °C throughout the incubation. Samples were pre-incubated for one week before any measurements were taken to minimize artifacts in response to rewetting the soil. Microbial CO2 production was measured in all samples once a week throughout the 8-week incubation. Before each measurement, the incubation bottles were briefly opened to allow the headspace to equilibrate with atmospheric conditions. Subsequently, the bottles were closed and one headspace sample was taken at time 0 (baseline) and again after 24 h with a 250 µL syringe through a septum in the lid. The CO2 concentration was measured using a LI-COR LI-8100A infrared gas analyzer (LI-COR Biosciences, Lincoln, NE, USA) configured for bench top measurements. Microbial respiration rates were calculated as the increase in CO2 concentration during the 24 h measurement period. Following each respiration measurement, we weighed the bottles and re-established the target moisture content if needed. Except during the 24 h CO2 measurement period, lids were put on the jars loosely to allow for CO2/O2 exchange, thus preventing the headspace and soils from becoming anoxic while minimizing water loss. Three replicate samples were used for each soil-charcoal mixture (n = 3), resulting in a total of 60 samples.
2.6. Potential Nitrification Rate
Potential nitrification rates were determined for unamended JP and LP soils, JP soils amended with JP-derived charcoal, and LP soils amended with LP-derived charcoal. Nitrification potential was measured using a shaken soil slurry method [28] as modified by Hart et al. [29]. We mixed 15 g of soil in a 250 mL Erlenmeyer flask with 100 mL of deionized water, 1.5 mM NH4+, and 1 mM PO43− solution to ensure that nitrifiers were not limited by NH4+ and P availability. The slurries were shaken in an orbital shaker at 180 rpm to maintain aerobic conditions for a 24 h period at room temperature (21 °C). Subsamples (10 mL) were taken at 2, 4, 22, and 24 h from each slurry, which included both soil and solution in order to keep the ratio of soil to solution constant. The subsamples were centrifuged at 2500 rpm, decanted, filtered using a 1.5 µm glass fiber filter, and analyzed for NO3− concentration using a Lachat Autoanalyzer (Hach Company, Loveland, CO, USA). The nitrification rate was determined by obtaining the slope of the regression line between the NO3− concentration and time. The slope was subsequently corrected for temperature, water content, and soil mass in the sample. The 24-h potential nitrification was measured at the beginning of the incubation and after 4 and 8 weeks on different sets of samples. In between samplings, samples were incubated at 25 °C and maintained for water content as described for the microbial respiration measurements above. Three replicates were used for each treatment (n = 3), resulting in a total of 30 samples per time period. We did not include all possible treatment combinations because we were not able to extract all samples in the 24 h period without compromising the data quality. Because of the relatively slow growth of the nitrifying microbial community, and because the nitrifying community is dependent on substrate supply for maintenance and growth, the potential nitrification assay can be used to indicate relative differences in the size of the nitrifying community that are integrated over the relevant periods of the incubation (initial, 4 weeks, 8 weeks; [28,29,30]).
2.7. Data Analysis
Soil pH data were analyzed using Analysis of Variance (ANOVA) with soil type (JP and LP), charcoal type (JP and LP), and charcoal amendment level (0%, 0.5%, 1%, 2.5%, and 5%) as discrete independent variables. Polyphenol data were analyzed using ANOVA with soil type and charcoal amendment level as discrete independent variables. Microbial respiration was expressed as CO2 production per gram of soil C or total C (soil C plus charcoal C). Respiration data was analyzed using repeated-measures ANOVA with soil type, charcoal type, and amendment level as discrete independent variables. Potential nitrification rate data expressed as mg N kg−1 soil day−1 were analyzed separately within each time period using ANOVA with soil type and charcoal amendment level as discrete independent variables. We tested for differences in potential nitrification between time periods using student’s t-tests. Potential nitrification data were log-transformed for the ANOVA analyses to ensure normal distribution of the data, but untransformed data are presented for ease of interpretation. Following all ANOVA analyses, we conducted post-hoc tests to determine significant differences between treatments using a Tukey’s approach. For the respiration and potential nitrification measurements, post-hoc tests were performed for each measurement time separately. All statistical analyses were conducted with JMP® 13 (Version 13, SAS Institute Inc., Cary, NC, USA). Probability levels ≤ 0.05 were considered to be significant.
3. Results
3.1. Soil and Charcoal Characteristics
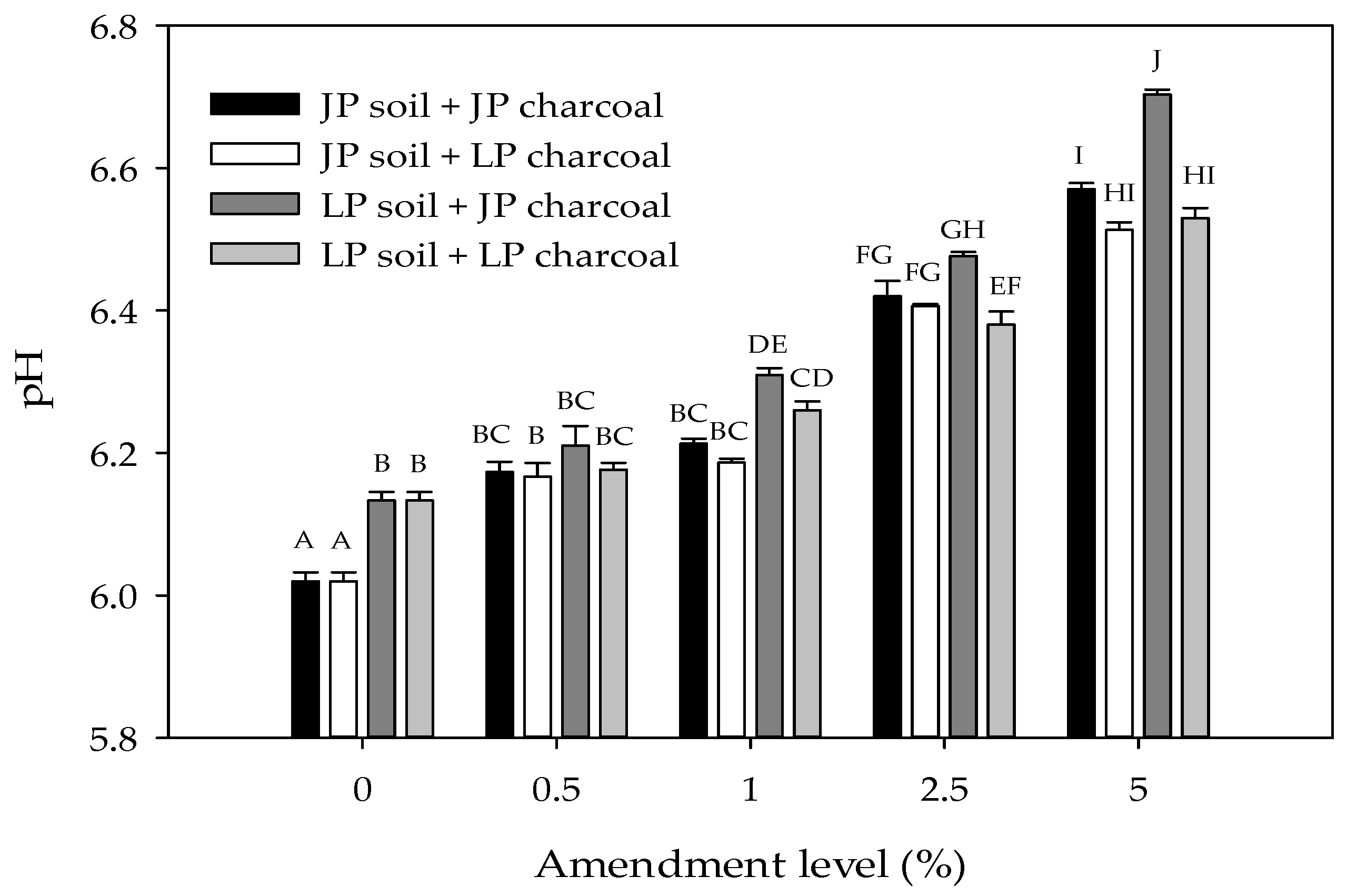
Total C, N, and pH were significantly higher in LP than in JP soils (Table 1 and Table 2; Figure 1). The addition of charcoal significantly increased the pH of both soils (Table 2). In LP soils, pH increased at all amendment levels, but in JP soils, pH was similar at the 0.5% and 1% amendment level. In addition, the effect of amendment level was strongest in soils amended with JP charcoal. Overall, pH was higher in soils amended with JP charcoal than in soils amended with LP charcoal. However, in JP soils, charcoal type did not affect pH, but in LP soils, pH was higher when amended with JP compared to LP charcoal. It should be noted though that differences in pH between soil and charcoal types were typically small (<0.1 unit; Figure 1). Surface area and CEC were similar in JP and LP-derived charcoal (Table 1).
3.2. Polyphenol Adsorption
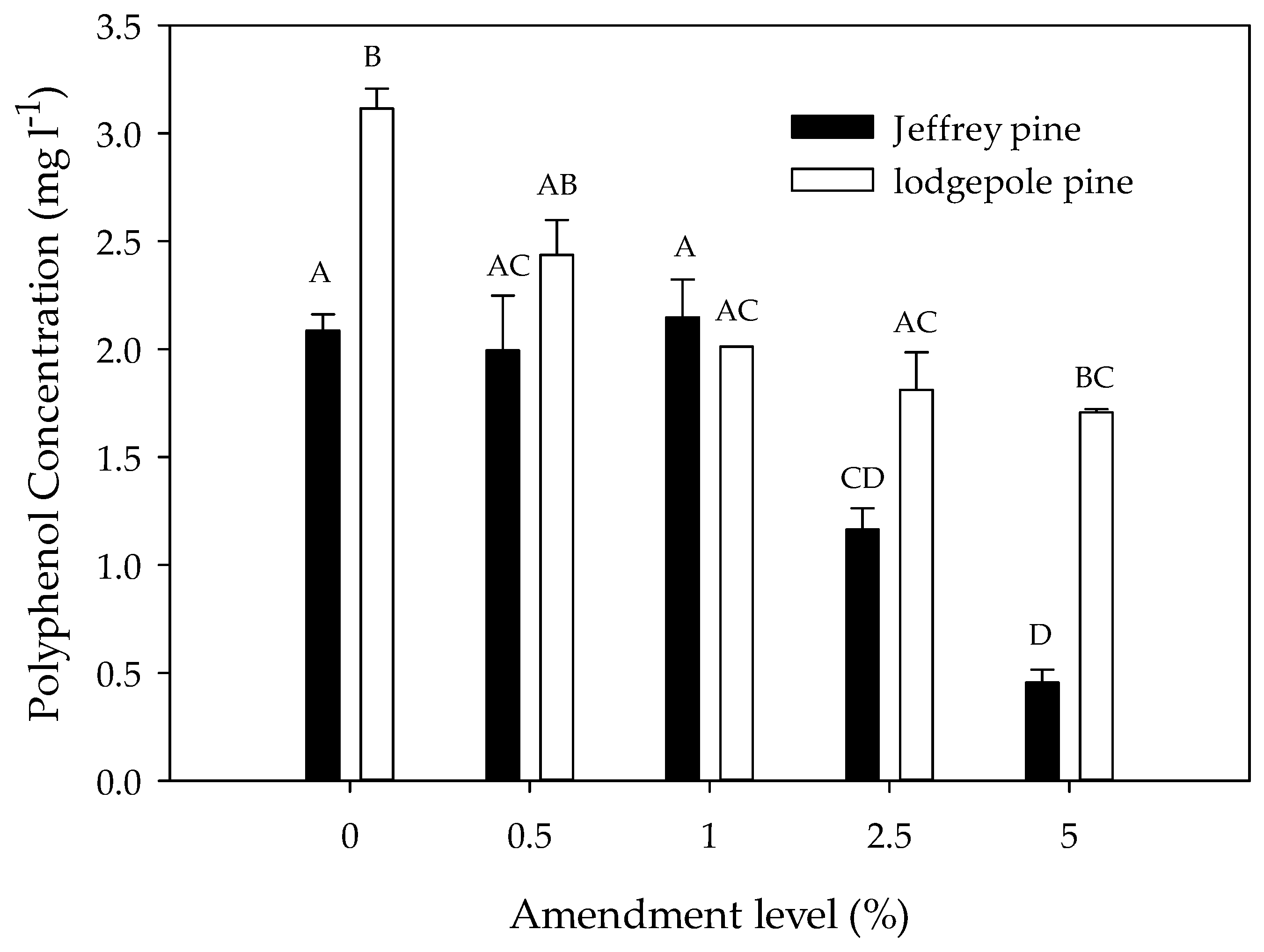
Average polyphenol concentrations in the soil extracts decreased with increasing charcoal amendment level in both JP and LP soils (Figure 2; Table 2). The post-hoc tests showed that polyphenol concentrations were similar at the 0%, 0.5%, and 1% amendment levels but consistently higher than at the 2.5% and 5% levels. The effect of amendment level varied by soil type; in LP soils, polyphenol concentrations were similar at the 0% and 0.5% amendment levels and significantly lower at the 1%, 2.5%, and 5% levels. In JP soils, polyphenol concentrations were similar at the 0%, 0.5%, and 1% levels and significantly lower at the 2.5% and 5% levels. The polyphenol concentrations were significantly higher in unamended and amended LP than in JP soils (Figure 2).
3.3. Microbial Respiration
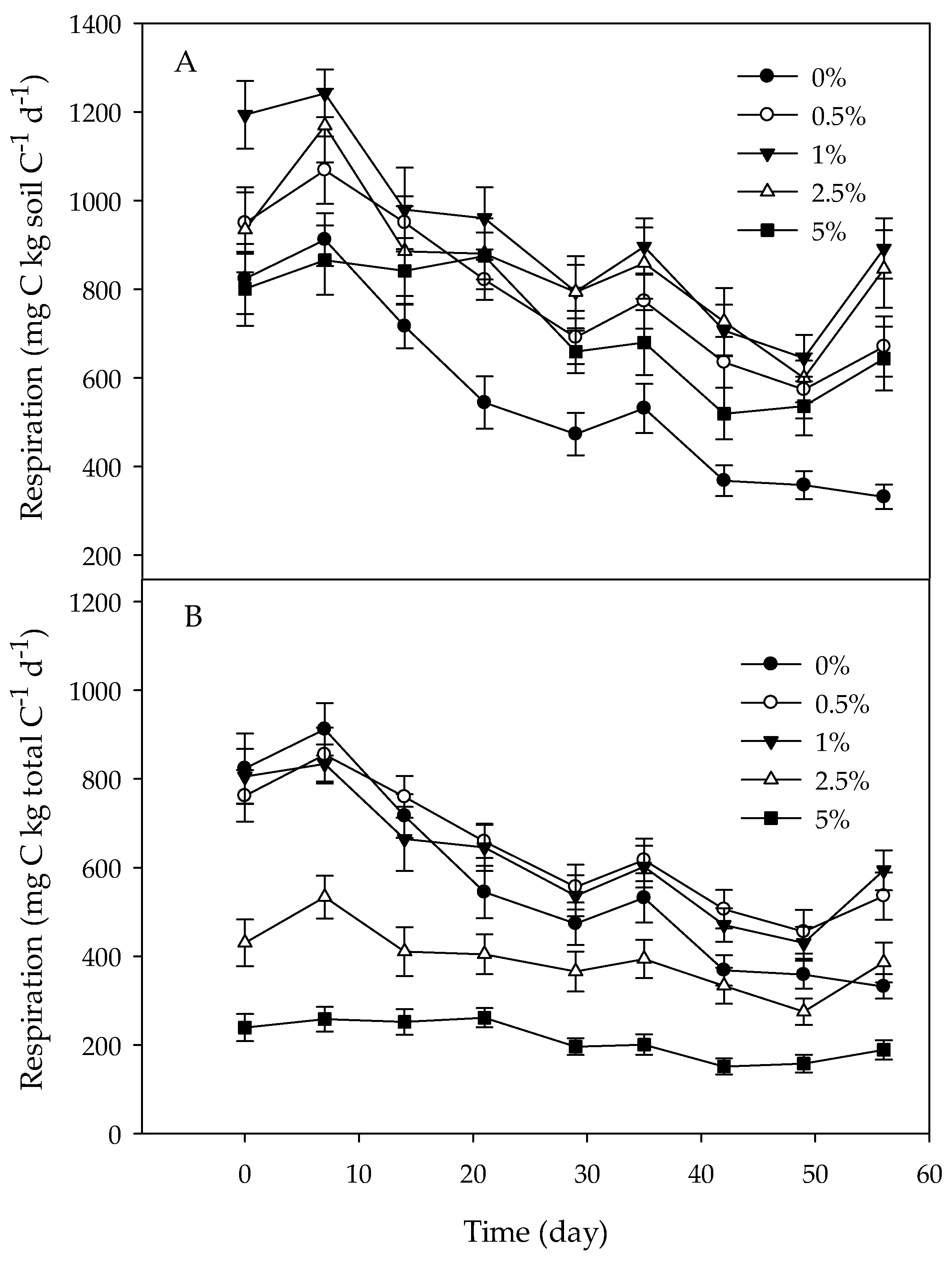
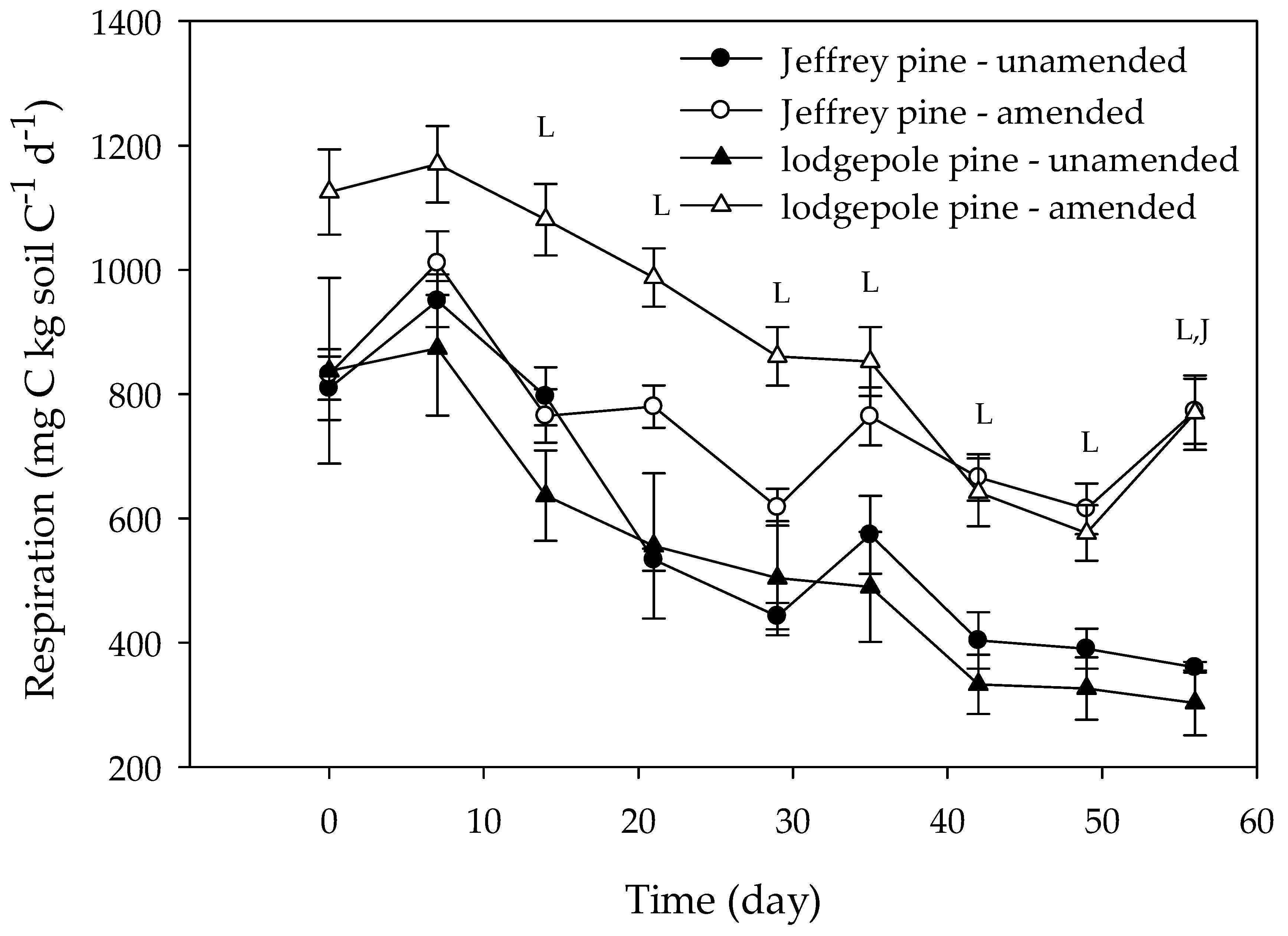
Overall, respiration expressed per gram of soil C was significantly higher in amended than in unamended soils (Figure 3 and Figure 4; Table 2 and Table 3), but respiration rates in the amended soils were consistently and significantly higher than in the unamended soils only at the 1% and 2.5% amendment levels (Figure 3; Table 2 and Table 3). The effect of charcoal amendment level on respiration depended on the soil type. In JP soils, respiration was stimulated most at the 2.5% amendment level, whereas in LP soils, respiration was highest at the 1% level. The effect of amendment on respiration varied with time. During the first three weeks, respiration was generally similar between unamended and amended soils (Figure 3; Table 3). After three weeks, respiration was generally higher in amended than in unamended soils but was similar among amendment levels. At the 5% amendment level, respiration was only significantly higher after 7 weeks. The temporal effects of amendment also varied with soil type. When all amendment levels were combined, respiration was significantly higher in amended than in unamended soils after day 14 in LP soils (Figure 4). In JP soils, amendment only had a significant effect on respiration on the last measurement day. Respiration rates in all treatments decreased during the incubation, but temporal patterns in respiration depended on soil type, with respiration declining more rapidly with time in LP than in JP soils. Overall, respiration rates were higher in LP soils than in JP soils.
When respiration rates were expressed per gram of total C, respiration was similar between the 0%, 0.5%, and 1% charcoal addition levels, but respiration rates declined with increasing charcoal amendment level (Figure 3; Table 2 and Table 3). Similar as when expressed per gram of soil C, the effect of amendment level on respiration varied by soil type. In JP soils, respiration was lowest at the 2.5% and 5% amendment levels. In LP soils, respiration was lowest at the 5% level, while respiration rates were similar at all other amendment levels.
Regardless of how respiration was expressed, respiration rates were not significantly affected by charcoal type. The ANOVA, however, showed significant 3- and 4-way interactions that included charcoal type when respiration was expressed per gram of soil C indicating that charcoal type affected respiration depending on measurement time and soil type (Table 2). In JP soils, charcoal type had no effect on respiration. In LP soils, average respiration was higher in soils amended with LP-derived charcoal during the first two weeks, but in the following period, respiration was higher in soils amended with JP-derived charcoal. It should be noted though that post-hoc tests conducted for each individual time period did not indicate significant differences in respiration between LP soils amended with LP- or JP-derived charcoal. None of these interactions were present when respiration was expressed per gram of total C.
3.4. Potential Nitrification
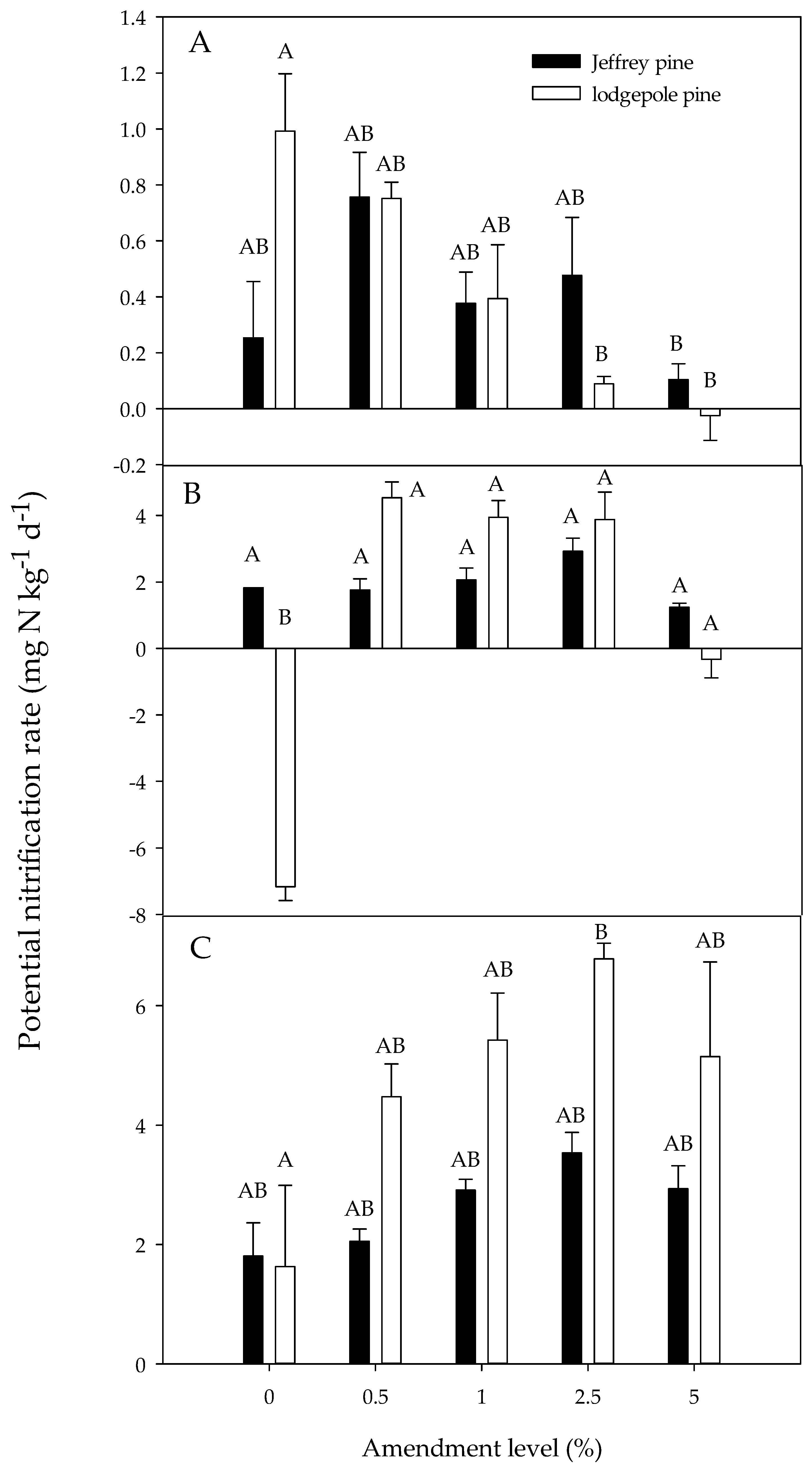
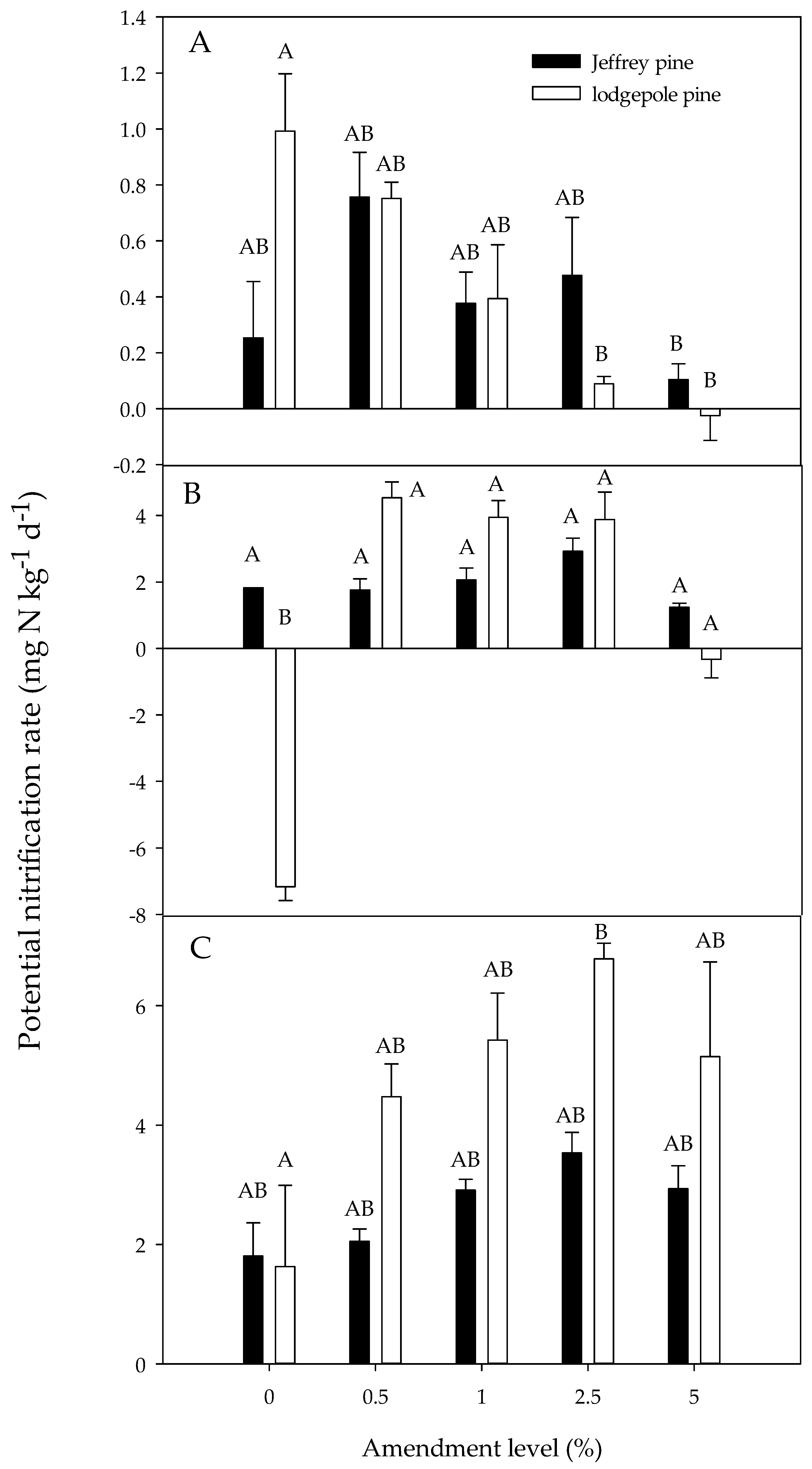
The potential nitrification rate was similar for day 0 and day 30 (p = 0.84) but significantly higher at day 64 compared to day 0 (p < 0.001) and day 30 (p < 0.001). Effects of amendment level and soil type depended on the measurement period. At day 0, potential nitrification was similar in the unamended soil and soils amended with 0.5%, 1%, and 2.5% charcoal, but it was significantly lower at the 5% amendment level (Figure 5; Table 4). The effects of amendment varied by soil type; in JP soils, nitrification was similar regardless of amendment level, but in LP soils, nitrification was significantly lower at the 2.5% and 5% amendment levels compared to the unamended soils (Figure 5; Table 4). Overall, potential nitrification rates were similar for JP and LP soils.
At day 30, potential nitrification rates were significantly higher in amended than the unamended soils, but rates were similar for the 0.5%, 1%, 2.5%, and 5% amendment levels (Figure 5; Table 4). The potential nitrification rate for the unamended LP soils appeared to be anomalously low, however, given that rates for unamended soils at day 0 and day 64 were much higher than for day 30. Similar to the observed patterns at day 0, the effect of charcoal addition at day 30 varied by soil type, with potential nitrification being similar for unamended and amended JP soils, but the anomalously low value for the unamended LP soils at day 30 caused the amended LP soils to have a significantly higher nitrification rate. Overall, potential nitrification was significantly higher in JP than LP soils, but when the low value for the LP soils was removed, no significant differences between soil types were observed.
At day 64, potential nitrification rates in the 2.5% charcoal amendment treatments exceeded the unamended treatment, but this was mainly caused by patterns observed in LP soils; potential nitrification rates were similar in unamended and amended JP soils, but in LP soils rates were higher in the 2.5% amendment treatment compared to the unamended soils. At day 64, potential nitrification rates were significantly higher in LP than in JP soils.
4. Discussion
Our study showed that the addition of charcoal to soils with low background levels of charcoal had mixed effects on microbial processes. Microbial respiration generally increased at intermediate amendment levels, but potential nitrification rates showed few effects of charcoal addition. Several mechanisms may explain our observed patterns. First, any increases in respiration upon addition of charcoal could have been caused by priming of native organic matter. This priming could be due to charcoal providing a refuge for microbes, thereby allowing them to avoid predation by larger soil fauna [14]. Previous studies have shown mixed effects of charcoal on priming and noted that conflicting reports regarding effects of priming may be due to soil types used, length of the incubation, and pre-incubation conditions, but no specifics were given how these factors may have affected priming [31]. Whether or not priming was important in our study could have been addressed by using isotopically labeled charcoal and measuring the isotopic signature of the respired CO2. However, creating uniformly labeled charcoal would require a very lengthy continuous exposure to 13C-enriched or depleted CO2, which would be challenging. In addition, we could have used a soil that contained organic matter derived from C4 vegetation to create a differential C signature between SOM and charcoal. We chose to use native soils originating from areas associated with the vegetation used to produce charcoal in order to include native microbial populations.
Second, decomposition of charcoal itself may have contributed to the increase in microbial respiration [12,32]. If this were the case, respiration expressed per gram of total C should have increased with increasing amounts of charcoal. Instead, respiration per gram of total C decreased with increasing charcoal amendment level, indicating that, even if charcoal itself decomposed, it had a lower quality of C than the native organic matter in the soil [18] and was not the main reason why respiration increased at intermediate amendment levels. Similar to priming, previous studies have shown inconsistent results with regard to the decomposability of charcoal. For instance, one study showed that charcoal derived from ponderosa pine was very resistant against decomposition by fungi [15], but a second study showed that the same charcoal type can decompose especially when charcoal had not been exposed to light or ultraviolet radiation [31]. The interaction between charcoal amendment level and soil type (Figure 3; Table 3) showed that charcoal had a larger stimulatory effect in LP than in JP soils, further indicating that other mechanisms than decomposition of charcoal itself were responsible for the observed patterns of respiration.
Third, charcoal may have adsorbed labile C compounds that were subsequently used as a substrate by microorganisms [14,17,22,33,34], thereby causing an increase in respiration. Our polyphenol data showed that polyphenol concentrations were lower in charcoal-amended than in unamended soils (Figure 2). Sorption of polyphenols was higher in JP soils at the higher amendment levels, but respiration did not increase at these higher levels, thus indicating that sorption of C compounds by itself may not explain patterns in respiration. Despite increased sorption of polyphenols, the effects of charcoal amendments on potential nitrification were small. When in solution, polyphenols can either immobilize N through complexation or cause enzymatic inhibition of nitrification, but when removed from solution, nitrification can be stimulated [17,21,35]; however, our data did not show consistent increases in nitrification potential at higher amendment levels. In addition, overall potential nitrification rates were higher in LP soils despite polyphenol sorption being lower, suggesting that the relationship between polyphenols and potential nitrification was tenuous at best.
Finally, the addition of charcoal caused an increase in soil pH (Figure 1), potentially making conditions more favorable for the microbial community and/or shifting the microbial community structure. Still, the changes in pH upon charcoal addition were relatively modest (around 0.1 unit) and unlikely to be ecologically significant. Other studies have shown, however, that the presence of charcoal can directly affect the composition of the microbial community. For instance, a recent study showed that the presence of activated charcoal increased the relative abundance of an unidentified bacterium and an Actinomycetales and decreased the relative abundance of a Flavobacterium, suggesting that charcoal can have an important impact on microbial community structure [36]. Still, overall, no single mechanism appeared to explain the observed patterns in respiration and potential nitrification in our study. We conducted a multiple regression analysis and the only significant correlation we observed was between polyphenol concentration and pH (r2 = 0.52; p < 0.001). We did not find significant correlations between pH and/or polyphenol concentration and potential nitrification rates or microbial respiration.
In general, charcoal type did not differentially affect microbial respiration in the amended soils, but the ANOVA showed that for LP soils, charcoal type affected respiration depending on the time period. It appeared that LP soils amended with LP-derived charcoal initially had higher respiration rates than soils amended with JP-derived charcoal, but this pattern was reversed in the later stages of the incubation. It is not entirely clear what mechanisms were responsible for these patterns, given that chemical and physical properties of JP- and LP-derived charcoal were very similar and this effect was only present in LP soils. Still, the different charcoal types differentially affected soil pH and polyphenol sorption, suggesting that other properties besides surface area and CEC may affect charcoal behavior in soils. The interaction between charcoal type, soil type, and time was not present when respiration was expressed per gram of total (soil C plus charcoal C), potentially because differences between soil types became smaller as relative differences in total C decreased with increasing charcoal content. Our results support previous studies showing that charcoal type does not have a major impact on microbial processes. A study comparing microbial activity in soils amended with charcoal originating from Empetrum nigrum twigs and forest humus material and activated C mixed with pumice found that both charcoal types stimulated basal respiration similarly [17]. In addition, another study found that net nitrification rates were similar in charcoal derived from ponderosa pine bark compared to other charcoal types despite physico-chemical characteristics being different [23]. This last study suggested that the temperature at which charcoal is produced is more important in determining microbial responses than source material. The lack of response of microbial processes to differences in charcoal composition in our study and other studies is somewhat surprising given that recent studies have shown that changes in chemical composition of charcoal can affect enzyme activity [15,31]. It is important to note that we did not cross all soil and charcoal type treatments for the polyphenol and potential nitrification measurements, as soils were amended with their respective charcoal types. As a result, it is difficult to separate the effects of soil type versus charcoal type on potential nitrification and/or polyphenol sorption. Particularly the polyphenol data showed differential behavior of charcoal/soil type that can most likely be ascribed to charcoal type rather than soil type. Polyphenol sorption was much higher at the higher amendment levels in JP soils than in LP soils despite surface area being similar between charcoal types, suggesting differences in reactivity between the charcoal types.
Charcoal appeared to have a larger stimulatory effect on microbial respiration in LP than in JP soils (Figure 3), which may be related to the type of organic matter present in the soil. Both under- and overstory vegetation differed between soil types, so it is likely that, in addition to differences in %C and %N, the organic matter chemistry was different between vegetation types as well. Despite these potential differences, respiration in unamended soils expressed per gram of soil C was similar in unamended JP and LP soils (Figure 4). Potential nitrification rates at the end of the incubation were higher in LP than in JP soils, however, (Figure 5; Table 4) indicating a higher capacity of LP soils to support nitrifying microorganisms. The higher nitrification rate in LP soils was not due to lower concentrations of potentially inhibitory compounds, since polyphenol concentrations were higher in LP than in JP unamended soils (Figure 5), again suggesting that relationships between polyphenol concentrations and nitrification are weak. The higher potential nitrification rates may have been related to potentially higher rates of N mineralization and thus increased NH4+ availability in LP soils.
The results from our study are different from previous studies that showed that respiration, NO3− concentrations, and net nitrification rates increase with increasing charcoal concentration even when higher amendment levels were used [11,21,22]. In some studies, lower levels of charcoal addition were used of up to 1% [21,22], but Kolb et al. [11] used amendment levels up to 10% and observed the largest stimulation of microbial respiration at the 5% and 10% amendment levels. The reasons for the discrepancies between our study and previous studies are not entirely clear. Potentially, the type of charcoal used varied between studies. Previous studies used activated C [20] or charcoal derived from mixtures of manure and woody materials [11] although our study and other studies suggest that charcoal type may not have a large effect on microbial processes. Potentially, particle size distribution of the charcoal and associated surface area may have varied between studies. Some studies used a mixture of course (>2mm) and fine (<2 mm) charcoal [11], fine (<2 mm) charcoal only [21], or unknown particle size [22]. Finally, incubation times varied between studies ranging from 21 days [32] to about 100 days [11] and several studies, including ours, show that effects of amendments can change over time. Particularly our potential nitrification data showed temporal effects and results could have changed had we extended the incubation period.
5. Conclusions
In our study, charcoal was added to soils that had not recently been subjected to fire, similar to the approach taken in other studies [11,21,22]. By amending unburned soils with charcoal, we were able to separate the effects of charcoal from effects of fire on soils, but one potential drawback is that the results of this study may not easily apply to field conditions. However, our incubation conditions may be representative for low-severity burns where direct impacts of fire on soils are typically small [8]. Overall, the presence of charcoal appeared to stimulate microbial respiration and may benefit the recovery of microbial populations following fire events. The degree of stimulation depended on the amendment level and at high levels of charcoal (>2.5%) presence of charcoal did not affect microbial respiration. Presence of charcoal had limited effects on potential nitrification rates. Charcoal had a larger stimulatory effect on microbial respiration in LP soils than JP soils. Charcoal type had little effect on microbial processes, but polyphenol sorption was higher on LP-derived than JP-derived charcoal at higher amendment levels despite surface area being similar for both charcoal types. In addition to the direct effects of charcoal on microbial processes, charcoal may indirectly affect microbial activity by increasing the water holding capacity and nutrient availability of the soil [34,37]. These indirect effects are likely to have a positive impact on microbial processes, but these aspects were not included in our study. Despite our results showing generally positive effects of charcoal on microbial activity, the exact mechanisms responsible for these effects are still somewhat poorly understood.
Acknowledgments
Funding for this work was provided by the Department of Natural Resources and Environmental Science of the University of Nevada, Reno, and the Whittell Graduate Fellowship awarded to Zachary Carter. We thank Jonathan Birkel who assisted with the field and laboratory work. We thank two anonymous reviewers for their valuable comments that greatly helped improve the paper.
Author Contributions
Z.C. and P.V. conceived and designed the experiments; Z.C. performed the experiments; Z.C. and P.V. analyzed the data; B.S., R.Q., R.B., and C.S. contributed to the data collection and provided analytical capabilities; Z.C. and P.V. wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Stephens, S.L.; Collins, B.M. Fire regimes of mixed-conifer forests in the north-central Sierra Nevada at multiple spatial scales. Northwest Sci. 2004, 78, 12–23. [Google Scholar]
- Moghaddas, E.E.Y.; Stephens, S.L. Thinning, burning, and thin-burn fuel treatment effects on soil properties in a Sierra Nevada mixed-conifer forest. For. Ecol. Manag. 2007, 250, 156–166. [Google Scholar] [CrossRef]
- Jiménez Esquilín, A.E.; Stromberger, M.E.; Massman, W.J.; Frank, J.M.; Shepperd, W.D. Microbial community structure and activity in a Colorado Rocky Mountain forest soil scarred by slash pile burning. Soil Biol. Biochem. 2007, 39, 1111–1120. [Google Scholar] [CrossRef]
- Johnson, D.W.; Miller, W.W.; Susfalk, R.B.; Murphy, J.D.; Dahlgren, R.A.; Glass, D.W. Biogeochemical cycling in forest soils of the eastern Sierra Nevada Mountains, USA. For. Ecol. Manag. 2009, 258, 2249–2260. [Google Scholar] [CrossRef]
- Prieto-Fernández, Á.; Carballas, M.; Carballas, T. Inorganic and organic N pools in soils burned or heated: Immediate alterations and evolution after forest wildfires. Geoderma 2004, 121, 291–306. [Google Scholar] [CrossRef]
- Canfield, D.E.; Glazer, A.N.; Falkowski, P.G. The evolution and future of Earth’s nitrogen cycle. Science 2010, 330, 192–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choromanska, U.; DeLuca, T.H. Prescribed fire alters the impact of wildfire on soil biochemical properties in a Ponderosa pine forest. Soil Sci. Soc. Am. J. 2001, 65, 232–238. [Google Scholar] [CrossRef]
- Certini, G. Effects of fire on properties of forest soils: A review. Oecologia 2005, 143, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Holden, S.R.; Rogers, B.M.; Treseder, K.R.; Randerson, J.T. Fire severity influences the response of soil microbes to a boreal forest fire. Environ. Res. Lett. 2016, 11, 035004. [Google Scholar] [CrossRef]
- Knelman, J.E.; Graham, E.B.; Trahan, N.A.; Schmidt, S.K.; Nemergut, D.R. Fire severity shapes plant colonization effects on bacterial community structure, microbial biomass, and soil enzyme activity in secondary succession of a burned forest. Soil Biol. Biochem. 2015, 90, 161–168. [Google Scholar] [CrossRef]
- Kolb, S.E.; Fermanich, K.J.; Dornbush, M.E. Effect of charcoal quantity on microbial biomass and activity in temperate soils. Soil Sci. Soc. Am. J. 2009, 73, 1173–1181. [Google Scholar] [CrossRef]
- Smith, J.L.; Collins, H.P.; Bailey, V.L. The effect of young biochar on soil respiration. Soil Biol. Biochem. 2010, 42, 2345–2347. [Google Scholar] [CrossRef]
- Wardle, D.A.; Nilsson, M.C.; Zackrisson, O. Fire-derived charcoal causes loss of forest humus. Science 2008, 320, 629. [Google Scholar] [CrossRef] [PubMed]
- Zackrisson, O.; Nilsson, M.C.; Wardle, D.A. Key ecological function of charcoal from wildfire in the Boreal forest. Oikos 1996, 77, 10–19. [Google Scholar] [CrossRef]
- Gibson, C.; Berry, T.D.; Wang, R.; Spencer, J.A.; Johnston, C.T.; Jiang, Y.; Bird, J.A.; Filley, T.R. Weathering of pyrogenic organic matter induces fungal oxidative enzyme response in single culture inoculation experiments. Org. Geochem. 2016, 92, 32–41. [Google Scholar] [CrossRef]
- Alexander, M. Introduction to Soil Microbiology, 2nd ed.; Wiley: New York, NY, USA, 1977; 467p. [Google Scholar]
- Pietikäinen, J.; Kiikkilä, O.; Fritze, H. Charcoal as a habitat for microbes and its effect on the microbial community of the underlying humus. Oikos 2000, 89, 231–242. [Google Scholar] [CrossRef]
- DeLuca, T.H.; Aplet, G.H. Charcoal and carbon storage in forest soils of the Rocky Mountain West. Front. Ecol. Environ. 2008, 6, 18–24. [Google Scholar] [CrossRef]
- Keech, O.; Carcaillet, C.; Nilsson, M.C. Adsorption of allelopathic compounds by wood-derived charcoal: The role of wood porosity. Plant Soil. 2005, 272, 291–300. [Google Scholar] [CrossRef]
- Hatton, P.J.; Chatterjee, S.; Filley, T.; Dastmalchi, K.; Plante, A.F.; Abiven, S.; Ga, X.; Masiello, C.A.; Leavitt, S.W.; Nadelhoffer, K.J.; et al. Tree taxa and pyrolysis temperature interact to control the efficacy of pyrogenic organic matter formation. Biogeochemistry 2016, 130, 103–116. [Google Scholar] [CrossRef]
- DeLuca, T.H.; MacKenzie, M.D.; Gundale, M.J.; Holben, W.E. Wildfire-produced charcoal directly influences nitrogen cycling in ponderosa pine forests. Soil Sci. Soc. Am. J. 2006, 70, 448–453. [Google Scholar] [CrossRef]
- Berglund, L.M.; DeLuca, T.H.; Zackrisson, O. Activated carbon amendments to soil alters nitrification rates in Scots pine forests. Soil Biol. Biochem. 2004, 36, 2067–2073. [Google Scholar] [CrossRef]
- Gundale, M.J.; DeLuca, T.H. Temperature and source material influence ecological attributes of ponderosa pine and Douglas-fir charcoal. For. Ecol. Manag. 2006, 231, 86–93. [Google Scholar] [CrossRef]
- Soil Survey Staff. Keys to Soil Taxonomy, 12th ed.; United States Department of Agriculture-National Resources Conservation Service: Washington, DC, USA, 2014; pp. 1–360.
- Johnson, B.G.; Johnson, D.W.; Miller, W.W.; Carroll-Moore, E.M.; Board, D.I. The effects of slash pile burning on soil and water macronutrients. Soil Sci. 2011, 176, 413–425. [Google Scholar] [CrossRef]
- Sumner, M.E.; Miller, W.P. Cation exchange capacity and exchange coefficients. In Methods of Soil Analysis. Part 3. Chemical Methods; Sparks, D.L., Ed.; Soil Science Society of America: Madison, WI, USA, 1996; pp. 1201–1230. [Google Scholar]
- Rice, E.W.; Baird, R.B.; Eaton, A.D.; Clesceri, L.S. Standard Methods for Examination of Water and Wastewater, 22nd ed.; American Water Works Association: Washington, DC, USA, 2012. [Google Scholar]
- Belser, L.W.; Mays, E.L. Specific inhibition of nitrite oxidation by chlorate and its use in assessing nitrification in soils and sediments. Appl. Environ. Microb. 1980, 39, 505–510. [Google Scholar]
- Hart, S.C.; Stark, J.M.; Davidson, E.A.; Firestone, M.K. Nitrogen mineralization, immobilization, and nitrification. In Methods of Soil Analysis: II. Microbiological and Biochemical Properties; Weaver, R.W., Ed.; Soil Science Society of America: Madison, WI, USA, 1994; pp. 985–1018. [Google Scholar]
- Sullivan, B.W.; Selmants, P.C.; Hart, S.C. New evidence that high potential nitrification rates occur in soils during dry sesasons: Are microbial communities metabolically active during dry seasons? Soil Biol. Biochem. 2012, 53, 28–31. [Google Scholar] [CrossRef]
- Wang, R.; Gibson, C.D.; Berry, T.D.; Jiang, Y.; Bird, J.A.; Filley, T.R. Photooxidation of pyrogenic organic matter (PyOM) reduces the reactive, labile C pool and the apparent soil oxidative microbial enzyme response. Geoderma 2017, 293, 10–18. [Google Scholar] [CrossRef]
- Gundale, M.J.; DeLuca, T.H. Charcoal effects on soil solution chemistry and growth of Koeleria macrantha in the ponderosa pine/Douglas-fir ecosystem. Biol. Fertil. Soils 2007, 43, 303–311. [Google Scholar] [CrossRef]
- DeLuca, T.H.; Nilsson, M.C.; Zackrisson, O. Nitrogen mineralization and phenol accumulation along a fire chronosequence in northern Sweden. Oecologia 2002, 133, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Wardle, D.A.; Zackrisson, O.; Nilsson, M.C. The charcoal effect in Boreal forests: Mechanisms and ecological consequences. Oecologia 1998, 115, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Ball, P.N.; MacKenzie, M.D.; DeLuca, T.H.; Holben, W.E. Wildfire and charcoal enhance nitrification and ammonium-oxidizing bacterial abundance in dry montane forest soils. J. Environ. Qual. 2010, 39, 1243–1253. [Google Scholar] [CrossRef] [PubMed]
- Nolan, N.E.; Kulmatiski, A.; Beard, K.H.; Norton, J.M. Activated carbon decreases invasive plant growth by mediating plant–microbe interactions. AoB Plants 2015, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Glaser, B.; Lehmann, J.; Zech, W. Ameliorating physical and chemical properties of highly weathered soils in the tropics with charcoal—A review. Biol. Fertil. Soils 2002, 35, 219–230. [Google Scholar] [CrossRef]
Figure 1.
Soil pH as a function of charcoal type and amendment level in Jeffrey pine (JP) and lodgepole pine (LP) soils. Error bars represent standard error of the mean pH (n = 3). Different letters indicate significant differences (p < 0.05) across soil types, charcoal types, and amendment levels.
Figure 1.
Soil pH as a function of charcoal type and amendment level in Jeffrey pine (JP) and lodgepole pine (LP) soils. Error bars represent standard error of the mean pH (n = 3). Different letters indicate significant differences (p < 0.05) across soil types, charcoal types, and amendment levels.
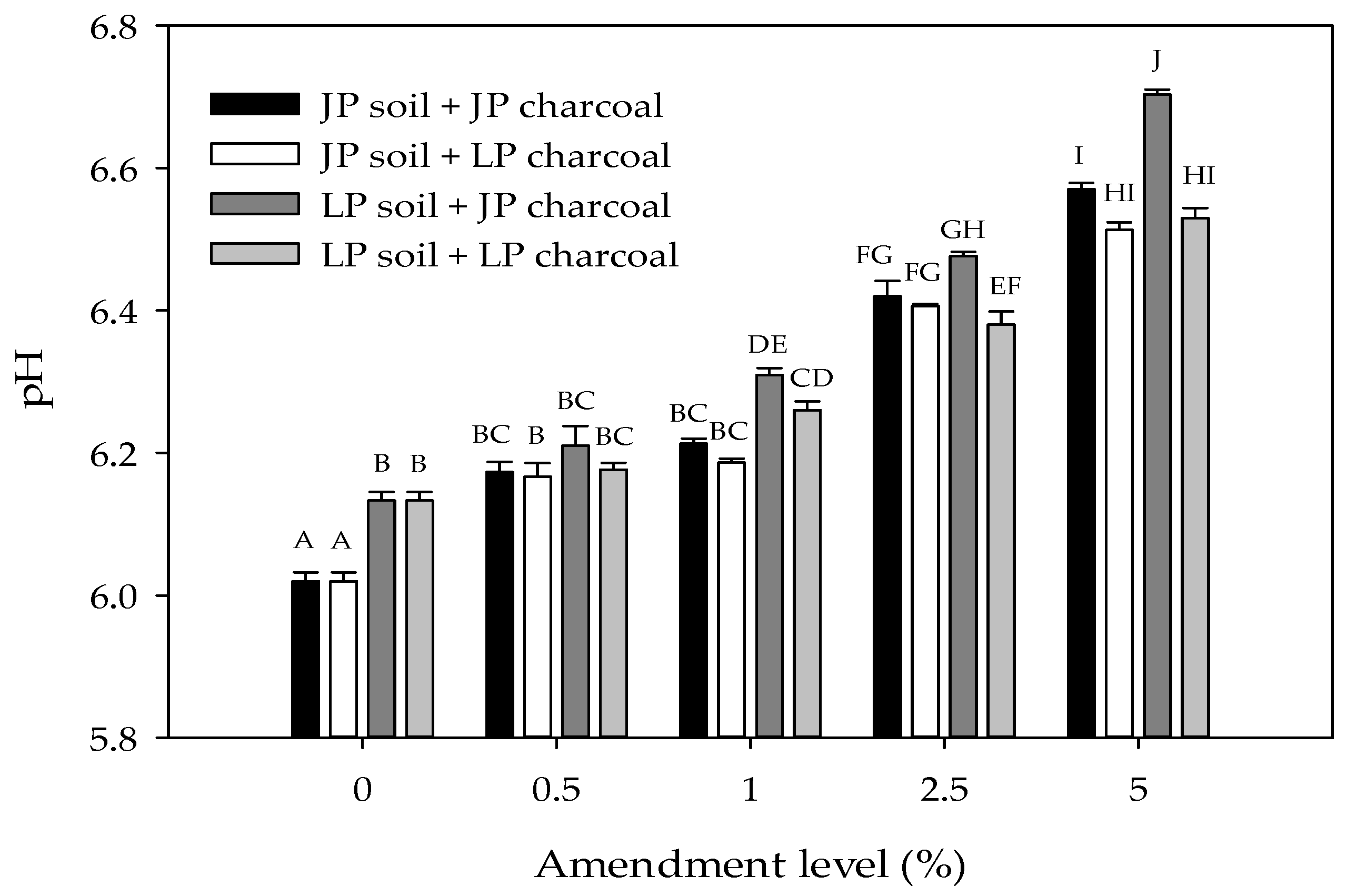
Figure 2.
Polyphenol concentration in soil extracts as a function of charcoal amendment level (% by mass) in Jeffrey pine and lodgepole pine soils. Error bars represent standard error of the mean polyphenol concentration (n = 3). Different letters indicate significant differences (p < 0.05) across soil types and amendment levels.
Figure 2.
Polyphenol concentration in soil extracts as a function of charcoal amendment level (% by mass) in Jeffrey pine and lodgepole pine soils. Error bars represent standard error of the mean polyphenol concentration (n = 3). Different letters indicate significant differences (p < 0.05) across soil types and amendment levels.
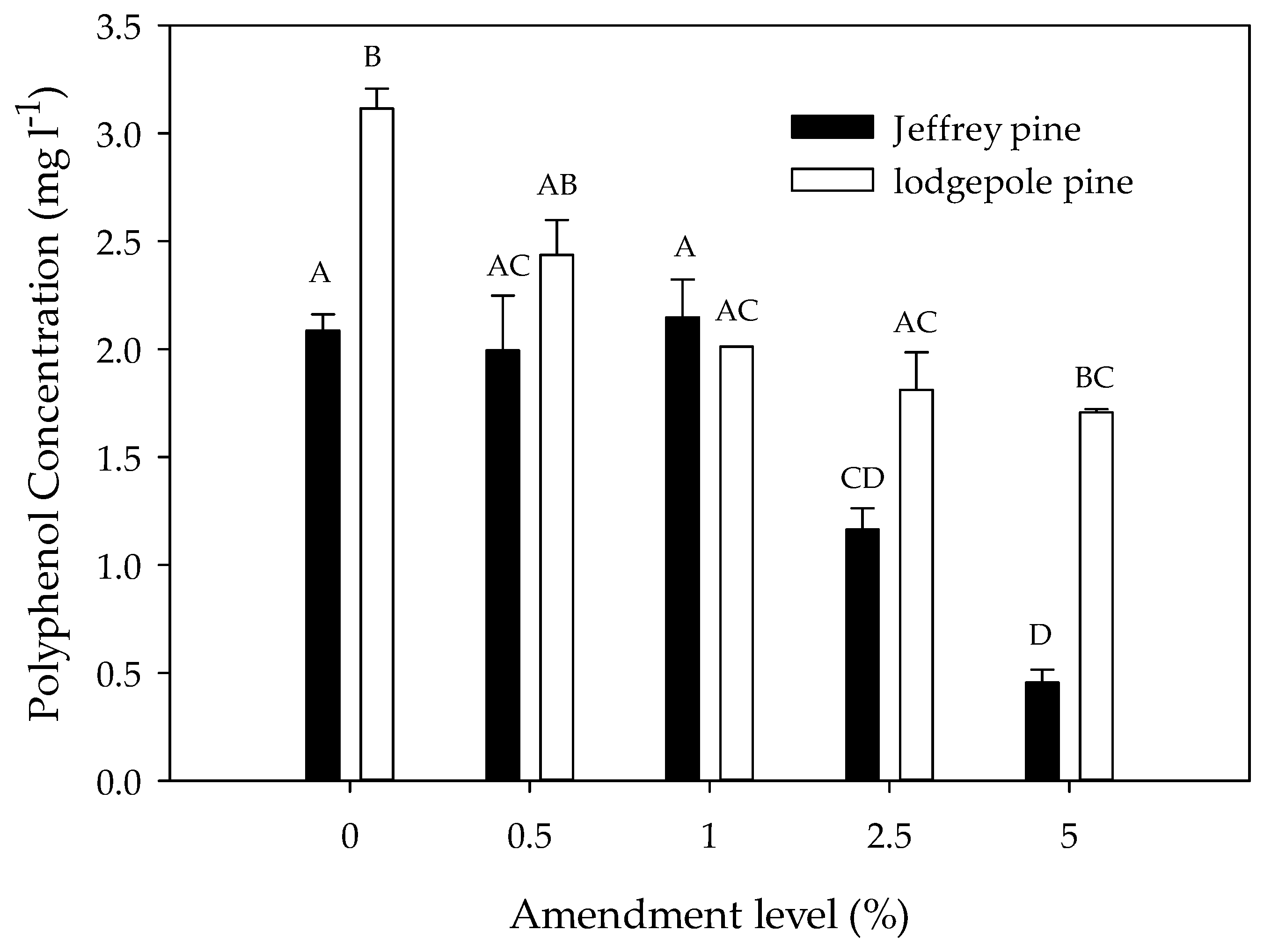
Figure 3.
Microbial respiration expressed per g soil C (A) or total C (soil C plus charcoal C); (B) during a two-month long incubation. Data are pooled by soil type and charcoal type to emphasize the effect of amendment level. Day 0 represents the first measurement day following a one-week pre-incubation period. Error bars represent standard errors of the mean (n = 12). Results of post-hoc tests assessing differences between amendment levels for each measurement time are shown in Table 3.
Figure 3.
Microbial respiration expressed per g soil C (A) or total C (soil C plus charcoal C); (B) during a two-month long incubation. Data are pooled by soil type and charcoal type to emphasize the effect of amendment level. Day 0 represents the first measurement day following a one-week pre-incubation period. Error bars represent standard errors of the mean (n = 12). Results of post-hoc tests assessing differences between amendment levels for each measurement time are shown in Table 3.

Figure 4.
Microbial respiration expressed per gram of soil C in unamended and amended Jeffrey and lodgepole pine soils. Values for amended soils were pooled by amendment level and charcoal type to emphasize how soil type affects the response to charcoal amendment. Day 0 represents the first measurement day following a one-week pre-incubation period. Error bars represent standard errors of the mean (n = 6 for unamended soils, n = 24 for amended soils). The letters at different measurement times indicate significant (p < 0.05) differences between unamended and amended Jeffrey pine soils (J) or unamended and amended lodgepole pine soils (L) for that particular measurement time.
Figure 4.
Microbial respiration expressed per gram of soil C in unamended and amended Jeffrey and lodgepole pine soils. Values for amended soils were pooled by amendment level and charcoal type to emphasize how soil type affects the response to charcoal amendment. Day 0 represents the first measurement day following a one-week pre-incubation period. Error bars represent standard errors of the mean (n = 6 for unamended soils, n = 24 for amended soils). The letters at different measurement times indicate significant (p < 0.05) differences between unamended and amended Jeffrey pine soils (J) or unamended and amended lodgepole pine soils (L) for that particular measurement time.
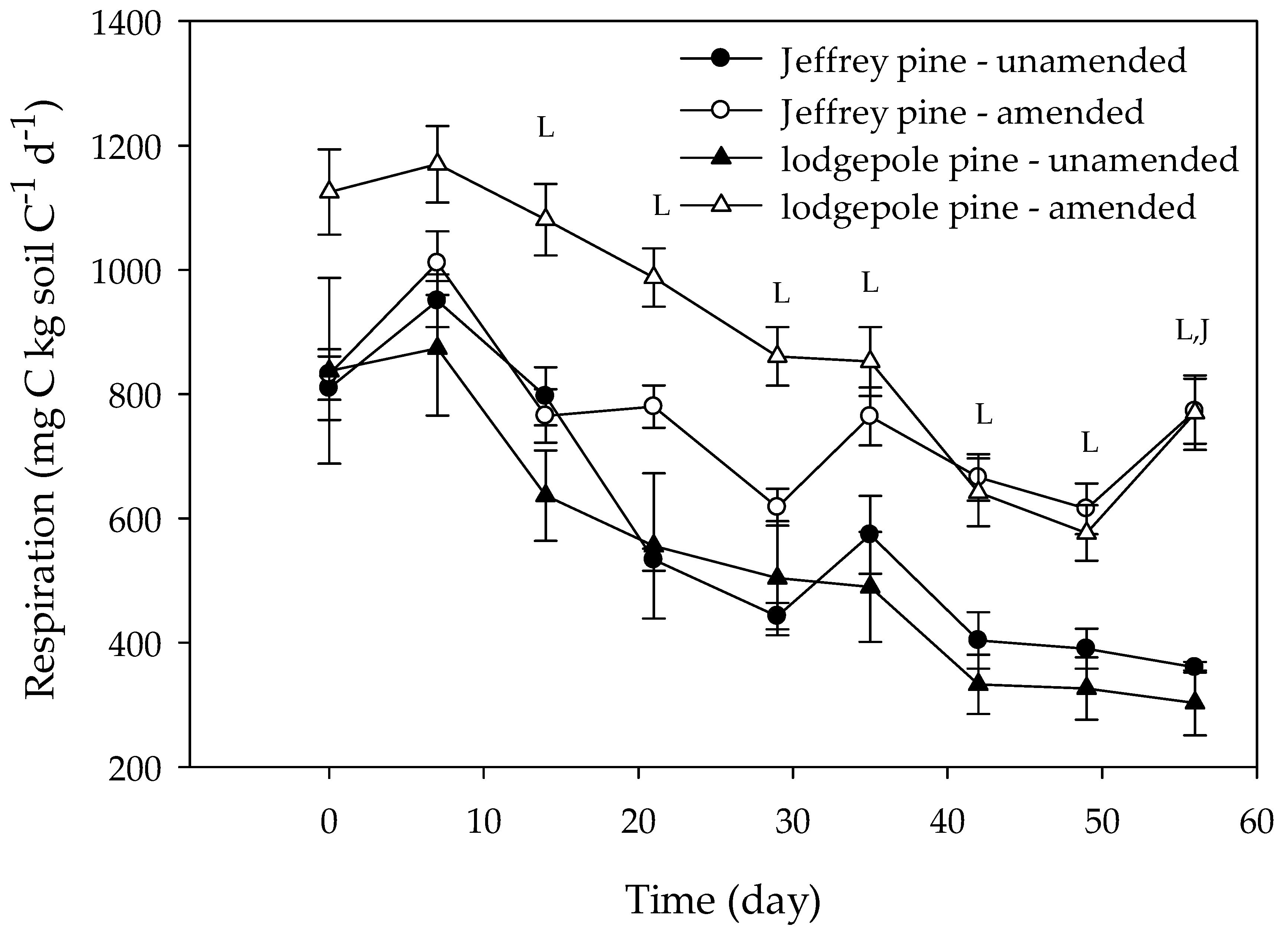
Figure 5.
Potential nitrification rate as a function of the charcoal amendment level for both Jeffrey pine and lodgepole pine soil treatments at day 0 (A), day 30 (B), and day 64 (C). Day 0 represents the first measurement day following a one-week pre-incubation period. Error bars represent standard error of the mean (n = 3). Different letters indicate significant differences (p < 0.05) across soil types and amendment levels.
Figure 5.
Potential nitrification rate as a function of the charcoal amendment level for both Jeffrey pine and lodgepole pine soil treatments at day 0 (A), day 30 (B), and day 64 (C). Day 0 represents the first measurement day following a one-week pre-incubation period. Error bars represent standard error of the mean (n = 3). Different letters indicate significant differences (p < 0.05) across soil types and amendment levels.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Physical and chemical properties of Jeffrey pine (JP) and lodgepole pine (LP) soils and charcoal used in this study. The pH, total C, and total N measurements were done in triplicate, surface area was measured in duplicate, and no replicates were used for cation exchange capacity (CEC) measurements.
Table 1.
Physical and chemical properties of Jeffrey pine (JP) and lodgepole pine (LP) soils and charcoal used in this study. The pH, total C, and total N measurements were done in triplicate, surface area was measured in duplicate, and no replicates were used for cation exchange capacity (CEC) measurements.
| Site | pH | Total C | Total N | CEC | Surface Area |
|---|---|---|---|---|---|
| (%) | (%) | meq 100 g−1 | m2 g−1 | ||
| JP Soil | 6.02 ± 0.02 | 1.59 ± 0.02 | 0.06 ± 0.002 | 9.8 | - |
| LP Soil | 6.13 ± 0.01 | 2.24 ± 0.21 | 0.08 ± 0.006 | 11.2 | - |
| JP charcoal | - | 93.0 ± 0.2 | - | 26.5 | 468.5 ± 14.5 |
| LP charcoal | - | 92.9 ± 5.7 | - | 27.7 | 449.1 ± 7.0 |
Table 2.
Results from the ANOVA (degrees of freedom, df; F-values, F; and probabilities, p) for pH, polyphenol sorption, and respiration expressed per g soil C or total C (soil C plus charcoal C). Significant F and p values are highlighted in bold.
Table 2.
Results from the ANOVA (degrees of freedom, df; F-values, F; and probabilities, p) for pH, polyphenol sorption, and respiration expressed per g soil C or total C (soil C plus charcoal C). Significant F and p values are highlighted in bold.
| pH | Polyphenol | Respiration | Respiration | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Per g Soil C | Per g Total C | |||||||||||
| df | F | p | df | F | p | df | F | p | df | F | p | |
| Amendment Level (AL) | 4 | 574.997 | <0.001 | 4 | 21.845 | <0.001 | 4 | 6.997 | <0.001 | 4 | 24.021 | <0.001 |
| Charcoal Type (CT) | 1 | 37.358 | <0.001 | - | - | - | 1 | <0.001 | 0.980 | 4 | 0.030 | 0.863 |
| Soil Type (ST) | 1 | 69.938 | <0.001 | 1 | 32.040 | <0.001 | 1 | 4.772 | 0.035 | 1 | 6.630 | 0.014 |
| Time (T) | - | - | - | - | - | - | 8 | 66.556 | <0.001 | 8 | 79.612 | <0.001 |
| AL × CT | 4 | 6.923 | <0.001 | - | - | - | 4 | 0.264 | 0.990 | 4 | 0.191 | 0.942 |
| AL × ST | 4 | 6.333 | <0.001 | 4 | 4.107 | 0.016 | 4 | 3.618 | 0.013 | 4 | 3.035 | 0.028 |
| AL × T | - | - | - | - | - | - | 32 | 2.109 | 0.002 | 32 | 4.215 | <0.001 |
| CT × ST | 1 | 11.25 | 0.002 | - | - | - | 1 | 0.021 | 0.886 | 1 | 0.080 | 0.780 |
| CT × T | - | - | - | - | - | - | 8 | 1.461 | 0.209 | 8 | 1.313 | 0.271 |
| T × ST | - | - | - | - | - | - | 8 | 5.880 | <0.001 | 8 | 5.702 | <0.001 |
| AL × CT × ST | 4 | 2.095 | 0.100 | - | - | - | 32 | 0.925 | 0.459 | 32 | 0.341 | 0.848 |
| AL × CT × T | - | - | - | - | - | - | 32 | 1.344 | 0.128 | 32 | 0.974 | 0.516 |
| AL × ST × T | - | - | - | - | - | - | 32 | 1.656 | 0.027 | 32 | 2.160 | <0.001 |
| CT × ST × T | - | - | - | - | - | - | 8 | 3.292 | <0.001 | 8 | 1.611 | 0.160 |
| CT × T × ST × AL | - | - | - | - | - | - | 32 | 1.735 | 0.017 | 32 | 1.097 | 0.349 |
Table 3.
Results from post-hoc tests assessing differences in microbial respiration between amendment levels at each measurement time. Day 0 represents the first measurement day following a one-week pre-incubation period. Different letters indicate significant (p < 0.05) differences between amendment levels for each measurement day.
Table 3.
Results from post-hoc tests assessing differences in microbial respiration between amendment levels at each measurement time. Day 0 represents the first measurement day following a one-week pre-incubation period. Different letters indicate significant (p < 0.05) differences between amendment levels for each measurement day.
| Amendment | Day | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 7 | 14 | 21 | 29 | 35 | 42 | 49 | 56 | ||
| 0% | A | A | A | A | A | A | A | A | A | |
| 0.5% | AB | AB | AB | B | AB | AB | B | B | B | |
| Per g soil C | 1% | B | B | B | B | B | B | B | B | B |
| 2.5% | AB | AB | AB | B | B | B | B | B | B | |
| 5% | A | A | AB | B | AB | AB | AB | B | B | |
| 0% | A | A | A | AB | AB | AB | AB | AB | CD | |
| 0.5% | A | A | A | A | A | A | A | A | AB | |
| Per g total C | 1% | A | A | A | A | A | A | AB | A | A |
| 2.5% | B | B | B | BC | B | BC | B | BC | BC | |
| 5% | B | C | B | C | C | C | C | C | D |
Table 4.
Results from ANOVA (degrees of freedom, df; F-values, F; and probabilities, p) for potential nitrification for each measurement period. Day 0 represents the first measurement day following a one-week pre-incubation period. Significant F and p values are given in bold.
Table 4.
Results from ANOVA (degrees of freedom, df; F-values, F; and probabilities, p) for potential nitrification for each measurement period. Day 0 represents the first measurement day following a one-week pre-incubation period. Significant F and p values are given in bold.
| Day 0 | Day 30 | Day 60 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| df | F | p | df | F | p | df | F | p | |
| Amendment Level | 4 | 5.941 | 0.026 | 4 | 7.569 | 0.001 | 4 | 3.444 | 0.028 |
| Soil Type | 1 | 0.177 | 0.679 | 1 | 8.414 | 0.010 | 1 | 8.478 | 0.009 |
| Amend. Level × Soil Type | 4 | 3.224 | 0.034 | 4 | 7.230 | 0.002 | 4 | 0.838 | 0.518 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Carter, Z.W.; Sullivan, B.W.; Qualls, R.G.; Blank, R.R.; Schmidt, C.A.; Verburg, P.S.J. Charcoal Increases Microbial Activity in Eastern Sierra Nevada Forest Soils. Forests 2018, 9, 93. https://doi.org/10.3390/f9020093
AMA Style
Carter ZW, Sullivan BW, Qualls RG, Blank RR, Schmidt CA, Verburg PSJ. Charcoal Increases Microbial Activity in Eastern Sierra Nevada Forest Soils. Forests. 2018; 9(2):93. https://doi.org/10.3390/f9020093
Chicago/Turabian StyleCarter, Zachary W., Benjamin W. Sullivan, Robert G. Qualls, Robert R. Blank, Casey A. Schmidt, and Paul S.J. Verburg. 2018. "Charcoal Increases Microbial Activity in Eastern Sierra Nevada Forest Soils" Forests 9, no. 2: 93. https://doi.org/10.3390/f9020093
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.