Increased Forest Soil CO2 and N2O Emissions During Insect Infestation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Measurement
2.2. Incubation Experiment
2.3. Statistical Analyses
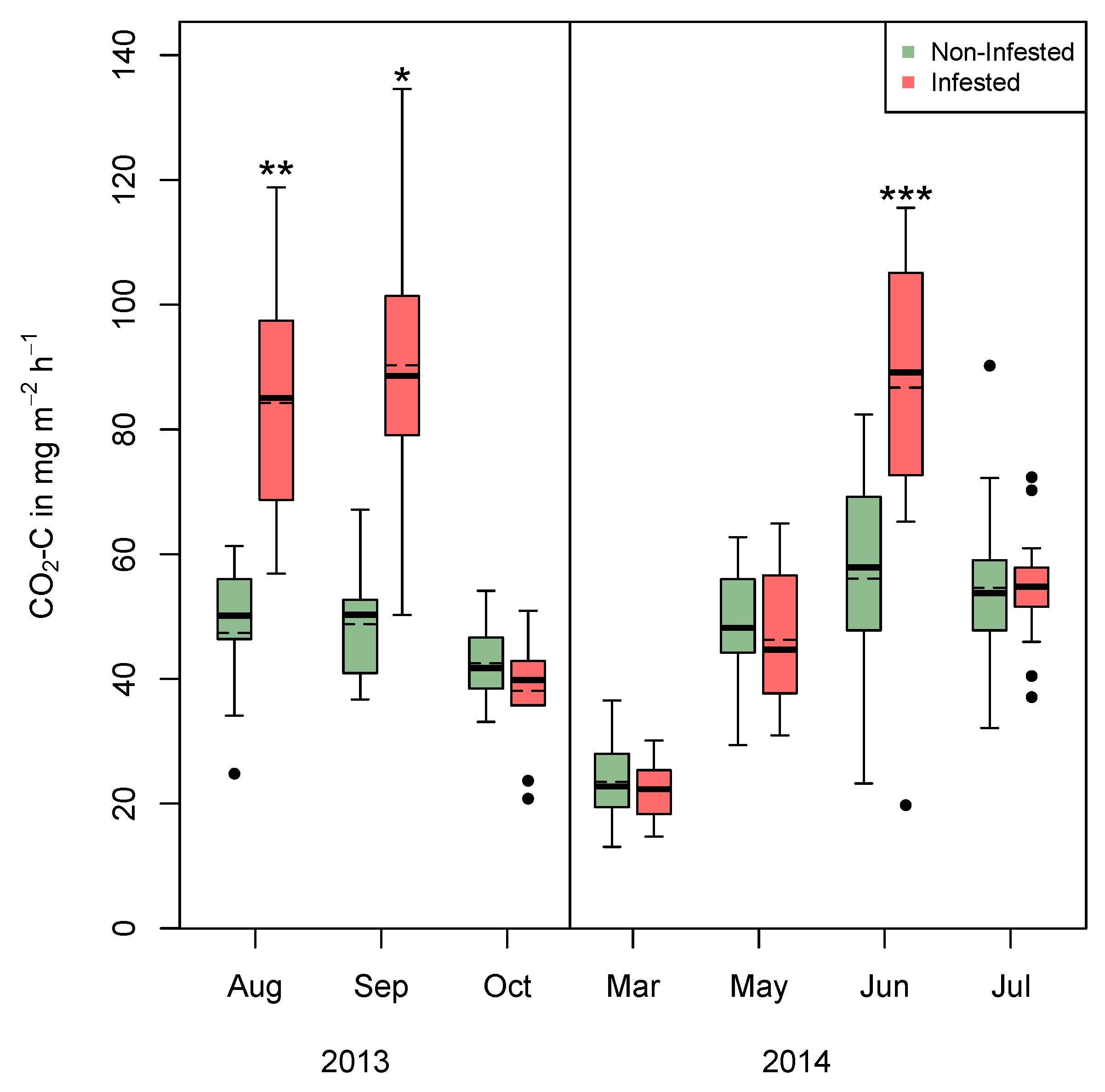
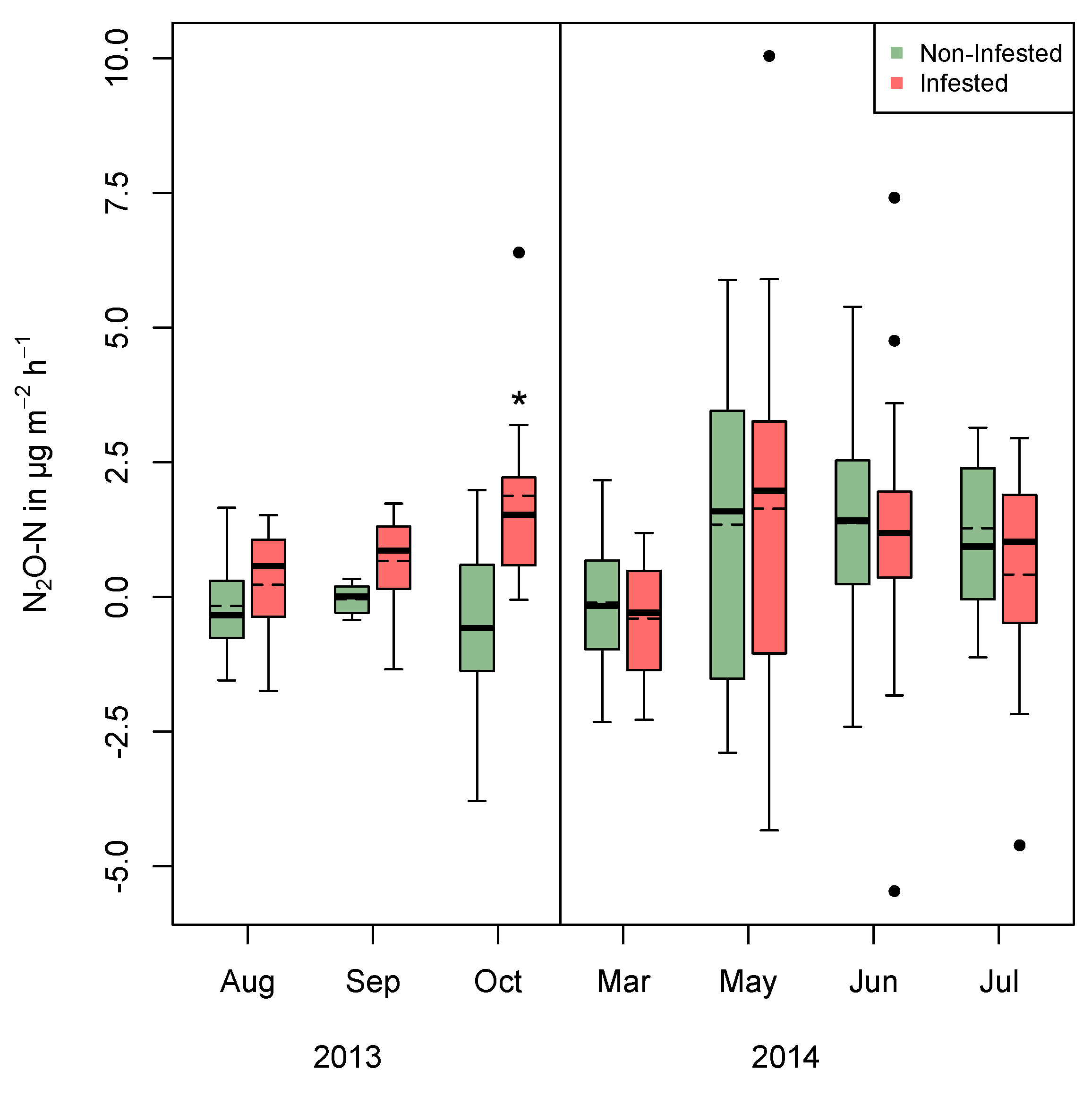
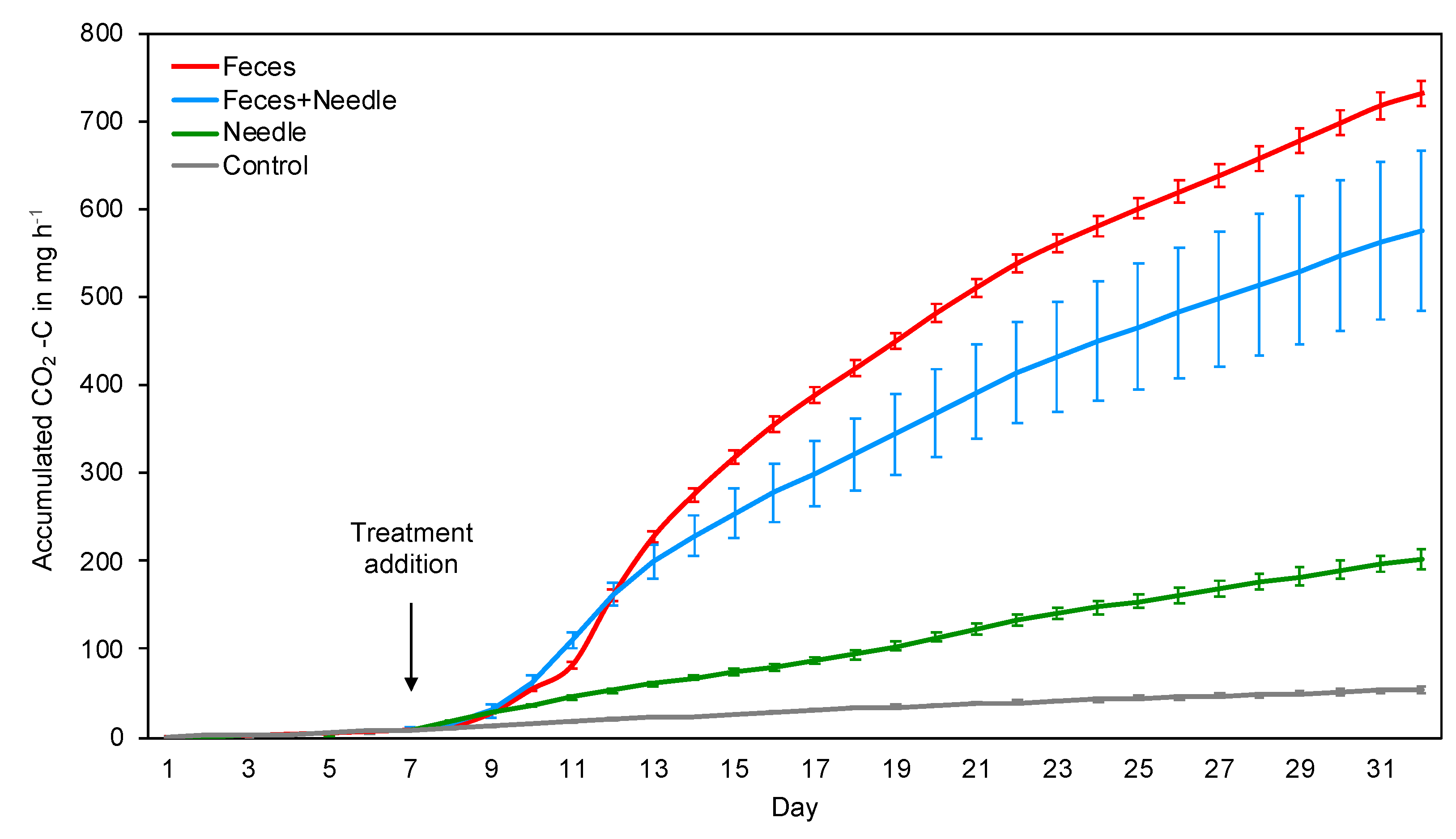
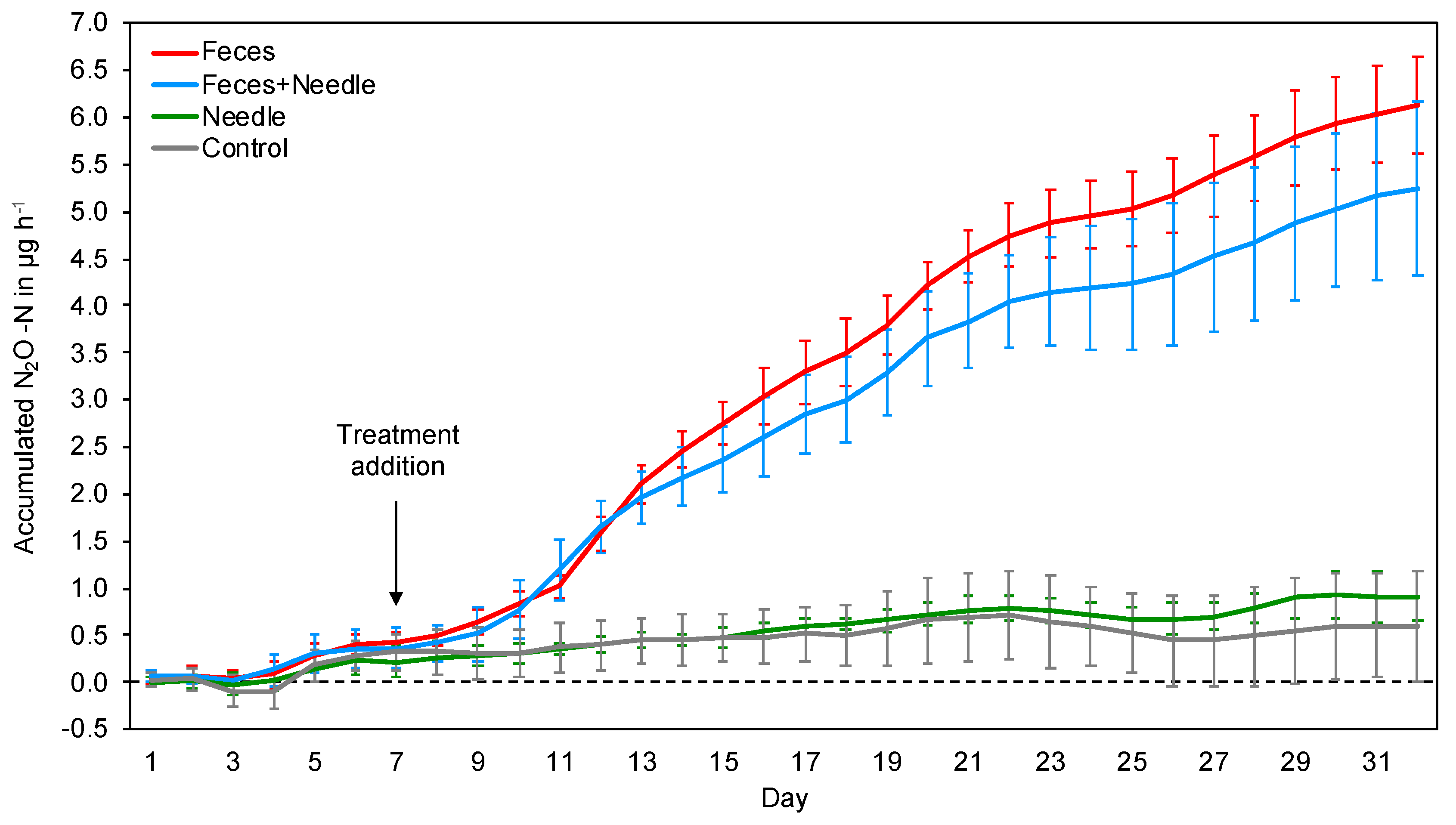
3. Results
3.1. Field Measurement
3.2. Incubation Experiment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kautz, M.; Meddens, A.J.H.; Hall, R.J.; Arneth, A. Biotic disturbances in Northern Hemisphere forests—A synthesis of recent data, uncertainties and implications for forest monitoring and modelling. Glob. Ecol. Biogeogr. 2017, 26, 533–552. [Google Scholar] [CrossRef]
- Kurz, W.A.; Dymond, C.C.; Stinson, G.; Rampley, G.J.; Neilson, E.T.; Carroll, A.L.; Ebata, T.; Safranyik, L. Mountain pine beetle and forest carbon feedback to climate change. Nature 2008, 452, 987–990. [Google Scholar] [CrossRef] [PubMed]
- Clark, K.L.; Skowronski, N.; Hom, J. Invasive insect impact forest carbon dynamics. Glob. Chang. Biol. 2010, 16, 88–101. [Google Scholar] [CrossRef]
- Dymond, C.C.; Neilson, E.T.; Stinson, G.; Porter, K.; MacLean, D.A.; Gray, D.R.; Campagna, M.; Kurz, W.A. Future Spruce Budworm Outbreak May Create a Carbon Source in Eastern Canadian Forests. Ecosystems 2010, 13, 917–931. [Google Scholar] [CrossRef]
- Hadden, D.; Grelle, A. Net CO2 emissions from a primary boreo-nemoral forest over a 10 year period. For. Ecol. Manag. 2017, 398, 164–173. [Google Scholar] [CrossRef]
- Grüning, M.M.; Simon, J.; Rennenberg, H.; L.M.-Arnold, A. Defoliating Insect Mass Outbreak Affects Soil N Fluxes and Tree N Nutrition in Scots Pine Forests. Front. Plant. Sci. 2017, 8, 954. [Google Scholar] [CrossRef]
- Stadler, B.; Müller, T.; Orwig, D.; Cobb, R. Hemlock Woolly Adelgid in New England Forests: Canopy Impacts Transforming Ecosystem Processes and Landscapes. Ecosystems 2005, 8, 233–247. [Google Scholar] [CrossRef]
- Müller, T.; Strobel, K.; Ulrich, A. Microorganisms in the phyllosphere of temperate forest ecosystems in a changing environment. In Microbial Ecology of Aerial Plant Surfaces; Chapter 4; Bailey, M.J., Lilley, A.K., Timms-Wilson, T.M., Spencer-Phillips, P.T.N., Eds.; CAB International: Wallingford, UK, 2006; p. 51. [Google Scholar] [CrossRef]
- Le Mellec, A.; Michalzik, B. Impact of a pine lappet (Dendrolimus pini) mass outbreak on C and N fluxes to the forest floor and soil microbial properties in a Scots pine forest in Germany. Can. J. For. Res. 2008, 38, 1829–1841. [Google Scholar] [CrossRef]
- Chapman, S.K.; Hart, S.C.; Cobb, N.S.; Whitham, T.G.; Koch, G.W. Insect Herbivory increases litter quality and decomposition: An extension of the acceleration hypothesis. Ecology 2003, 84, 2867–2876. [Google Scholar] [CrossRef]
- Belovsky, G.E.; Slade, J.B. Insect herbivory accelerates nutrient cycling and increases plant production. Proc. Natl. Acad. Sci. USA 2000, 97, 14412–14417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frost, C.F.; Hunter, M.D. Insect herbivory and frass deposition affect soil nutrient dynamics and export in oak mesocosms. Ecology 2004, 85, 3335–3347. [Google Scholar] [CrossRef]
- Le Mellec, A.; Habermann, M.; Michalzik, B. Canopy herbivory altering C to N ratios and soil input patterns of different organic matter fractions in a Scots pine forest. Plant Soil. 2009, 325, 255–262. [Google Scholar] [CrossRef]
- Fogal, W.H.; Slansky, F. Contribution of feeding by European pine sawfly larvae to litter production and element flux in Scots pine plantations. Can. J. For. Res. 1985, 15, 484–487. [Google Scholar] [CrossRef]
- Lovett, G.M.; Ruesink, A.E. Carbon and nitrogen mineralization from decomposing gypsy moth frass. Oecologia 1995, 104, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Zimmer, M.; Topp, W. The role of coprophagy in nutrient release from feces of phytophagous insects. Soil Biol. Biochem. 2002, 34, 1093–1099. [Google Scholar] [CrossRef]
- Stremińska, M.A.; Błaszczyk, M.; Kolk, A. Microbial abundance and some of their physiological activities in soil organic horizon of pine forest affected by insect herbivory. Pol. J. Environ. Stud. 2006, 15, 905–914. [Google Scholar]
- Ritchie, M.; Tilman, D.; Knops, J. Herbivore effects on plant and nitrogen dynamics in oak savanna. Ecology 1998, 79, 165–177. [Google Scholar] [CrossRef]
- Hillstrom, M.; Meehan, T.D.; Kelly, K.; Lindroth, R.L. Soil carbon and nitrogen mineralization following deposition of insect frass and greenfall from forests under elevated CO2 and O3. Plant Soil. 2010, 336, 75–85. [Google Scholar] [CrossRef]
- L.-M.-Arnold, A.; Grüning, M.; Simon, J.; Reinhardt, A.B.; Lamersdorf, N.; Thies, C. Forest defoliator pests alter carbon and nitrogen cycles. R. Soc. Open Sci. 2016, 3, 160361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morehouse, K.; Johns, T.; Kaye, J.; Kaye, M. Carbon and nitrogen cycling immediately following bark beetle outbreaks in southwestern ponderosa pine forests. For. Ecol. Manag. 2008, 255, 2698–2708. [Google Scholar] [CrossRef]
- Matthes, J.H.; Lang, A.K.; Jevon, F.V.; Russel, S.J. Tree Stress and Mortality from Emerald Ash Borer Does Not Systematically Alter Short-Term Soil Carbon Flux in a Mixed Northeastern U.S. Forest. Forests 2018, 9, 37. [Google Scholar] [CrossRef]
- Mikkelson, K.M.; Brouillard, B.M.; Bokman, C.M.; Sharp, J.O. Ecosystem resilience and limitations revealed by soil bacterial community dynamics in a bark beetle- impacted forest. mBio 2017, 8, e01305-17. [Google Scholar] [CrossRef] [PubMed]
- Brouillard, B.M.; Mikkelson, K.M.; Bokman, C.M.; Berryman, E.M.; Sharp, J.O. Extent of localized tree mortality influences soil biogeochemical response in a beetle-infested coniferous forest. Soil Biol. Biochem. 2017, 114, 309–318. [Google Scholar] [CrossRef]
- Russel, C.A.; Kosola, K.R.; Paul, E.A.; Robertson, G.P. Nitrogen cycling in poplar stands defoliated by insects. Biogeochemistry 2004, 68, 365–381. [Google Scholar] [CrossRef]
- Lindroth, R.L.; Dearing, D.M. Herbivory in a World of Elevated CO2. In A History of Atmospheric CO2 and Its Effects on Plants, Animals, and Ecosystems. Ecological Studies (Analysis and Synthesis); Baldwin, I.T., Caldwell, M.M., Heldmaier, G., Jackson, R.B., Lange, O.L., Mooney, H.A., Schulze, E.-D., Sommer, U., Ehleringer, J.R., Dearing, M.D., et al., Eds.; Springer: New York, NY, USA, 2005; Volume 177, pp. 468–486. [Google Scholar]
- Verkaik, E.; Jongkind, A.G.; Berendse, F. Short-term and long-term effects of tannins on nitrogen mineralization and litter decomposition in kauri (Agathis australis (D.Don) Lindl.) forests. Plant Soil. 2006, 287, 337–345. [Google Scholar] [CrossRef]
- Lovett, G.M.; Christenson, L.M.; Groffman, P.M.; Jones, C.G.; Hart, J.E.; Mitchell, M.J. Insect Defoliation and Nitrogen Cycling in Forests. BioScience 2002, 52, 335–341. [Google Scholar] [CrossRef]
- Hunter, M.D.; Linnen, C.R.; Reynolds, B.C. Effects of endemic densities of canopy herbivores on nutrient dynamics along a gradient in elevation in the southern Appalachians. Pedobiologia 2003, 47, 231–244. [Google Scholar] [CrossRef]
- Bejer, B. The Nun Moth in European Spruce Forests. In Dynamics of Forest Insect Populations, Population Ecology (Theory and Application); Berryman, A.A., Ed.; Springer: Boston, MA, USA, 1988; pp. 211–231. [Google Scholar]
- Beule, L.; Grüning, M.M.; Karlovsky, P.; l-M.-Arnold, A. ; Changes of Scots Pine Phyllosphere and Soil Fungal Communities during Outbreaks of Defoliating Insects. Forests 2017, 8, 316. [Google Scholar] [CrossRef]
- Lessard, R.; Rochette, P.; Topp, E.; Pattey, E.; Desjardins, R.; Beaumont, G. Methane and carbon fluxes from poorly drained adjacent cultivated and forest sites. Can. J. Soil Sci. 1994, 74, 139–146. [Google Scholar] [CrossRef]
- Loftfield, N.; Flessa, H.; Augustin, J.; Beese, F. Automated gas chromatographic system for rapid analysis of the atmospheric trace gases methane, carbon dioxide, and nitrous oxide. J. Environ. Qual. 1997, 26, 560–564. [Google Scholar] [CrossRef]
- Heinrichs, H.; Brumsack, H.-J.; Loftfield, N.; König, N. Verbessertes Druckaufschlußsystem für biologische und anorganische Materialien. J. Soil Sci. Plant Nutr. 1986, 149, 350–353. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing, R Foundation for Statistical Computing, Vienna, Austria. 2017. Available online: https://www.r-project.org (accessed on 10 August 2018).
- Christenson, L.M.; Lovett, G.M.; Mitchell, M.J.; Groffman, P.M. The fate of nitrogen in gypsy moth frass deposited to an oak forest floor. Oecologia 2002, 131, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Fielding, D.J.; Trainor, E.; Zhang, M. Diet influences rates of carbon and nitrogen mineralization from decomposing grasshopper frass and cadavers. Biol. Fertil. Soils. 2013, 49, 537–544. [Google Scholar] [CrossRef]
- Baranchicov, Y.N.; Perevoznikova, V.D.; Vishnyakova, Z.V. Carbon Emission by Soils in Forests Damaged by the Siberian Moth. Russ. J. Ecol. 2002, 33, 398–401. [Google Scholar] [CrossRef]
- Janssens, I.A.; Dieleman, W.; Luyssaert, S.; Subke, J.-A.; Reichstein, M.; Ceulemans, R.; Ciais, P.; Dolman, A.J.; Grace, J.; Matteucci, G.; et al. Reduction of forest soil respiration in response to nitrogen deposition. Nat. Geosci. 2010, 3, 315–322. [Google Scholar] [CrossRef]
- Kaukonen, M.; Ruotsalainen, A.L.; Wöli, P.R.; Männistö, M.L.; Setälä, H.; Saravesi, K.; Huusko, K.; Markkola, A. Moth herbivory enhances resource turnover in subarctic mountain birch forests? Ecology 2013, 94, 267–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grüning, M.M.; Beule, L.; Meyer, S.; Karlovsky, P.; L.M.-Arnold, A. The Abundance of Fungi, Bacteria and Denitrification Genes during Insect Outbreaks in Scots Pine Forests. Forests 2018, 9, 497. [Google Scholar] [CrossRef]
- Brady, N.C.; Weil, R.R. The Nature and Properties of Soils, 11th ed.; Prentice Hall: Upper Saddle River, NJ, USA, 1996; p. 740, ISBN-10: 1292162236. [Google Scholar]
- Sinsabaugh, R.L. Phenol oxidase, peroxidase and organic matter dynamics of soil. Soil Biol. Biochem. 2010, 42, 391–404. [Google Scholar] [CrossRef]
- Jungkunst, H.F.; Fiedler, S.; Stahr, K. N2O emissions of a mature Norway spruce (Picea abies) stand in the Black Forest (southwest Germany) as differentiated by the soil pattern. J. Geophys Res. 2004, 109, D07302. [Google Scholar] [CrossRef]
- Butterbach-Bahl, K.; Baggs, E.M.; Dannenmann, M.; Kiese, R.; Zechmeister-Boltenstern, S. Nitrous oxide emissions from soils: How well do we understand the processes and their controls? Phil. Trans. R. Soc. B. 2013, 368, 20130122. [Google Scholar] [CrossRef] [PubMed]
- Jung, P.; Lunderstädt, J. Effect of feces from pine-fed larvae of Dendrolimus pini L. (Lep., Lasiocampidae) on the germination and development of pine, birch and oak seeds. J. Appl. Ent. 2000, 124, 253–258. [Google Scholar] [CrossRef]
- Riek, W.; Stähr, F. Eigenschaften Typischer Waldböden im Nordostdeutschen Tiefland unter Besonderer Berücksichtigung des Landes Brandenburg. Hinweise für die Waldbewirtschaftung; Eberswalder Forstliche Schriftenreihe, Ministerium für ländliche Entwicklung, Umwelt und Verbraucherschutz des Landes Brandenburg: Eberswalde, Germany, 2004; Volume 19, pp. 1–180. [Google Scholar]
- Groffman, P.M.; Butterbach-Bahl, K.; Fulweiler, R.W.; Gold, A.J.; Morse, J.L.; Stander, E.K.; Tague, C.; Tonitto, C.; Vidon, P. Challenges to incorporating spatially and temporally explicit phenomena (hotspots and hot moments). Biogeochemistry 2009, 93, 49–77. [Google Scholar] [CrossRef]
- Aber, J.; McDowell, W.; Nadelhoffer, K.; Magill, A.; Berntson, G.; Kamakea, M.; McNulty, S.; Currie, W.; Rustad, L.; Fernandez, I. Nitrogen Saturation in Temperate Forest Ecosystems: Hypotheses revisited. BioScience 1998, 48, 921–934. [Google Scholar] [CrossRef]
- Stevenson, F.J.; Cole, M.A. Cycles of Soils: Carbon, Nitrogen, Phosphorus, Sulfur, Micronutrients, 2nd ed.; Wiley: Hoboken, NJ, USA, 1999; p. 448. ISBN 978-0-471-32071-5. [Google Scholar]
- Kosola, K.R.; Dickmann, D.I.; Parry, E.A.; Parry, D. Repeated insect defoliation effects on growth, nitrogen acquisition, carbohydrates, and root demography of poplars. Oecologia 2001, 129, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Simmons, M. J.; Lee, T. D.; Ducey, M. J.; Elkinton, J. S.; Boettner, G. H.; Dodds, K. J. Effects of Invasive Winter Moth Defoliation on Tree Radial Growth in Eastern Massachusetts, USA. Insects 2004, 5, 301–318. [Google Scholar] [CrossRef] [PubMed]
- Ayres, M.P.; Lombardero, M.J. Assessing the consequences of global change for forest disturbance from herbivores and pathogens Sci. Total Environ. 2000, 262, 263–286. [Google Scholar] [CrossRef]
- Dale, V.H.; Joyce, L.A.; McNulty, S.; Neilson, R.P.; Ayres, M.P.; Flannigan, M.D.; Hanson, P.J.; Irland, L.C.; Lugo, A.E.; Peterson, C.J.; et al. Climate change and forest disturbances. BioScience 2001, 51, 723–734. [Google Scholar] [CrossRef]
- Flower, C.E.; Gonzales-Meler, M.A. Responses of temperate forest productivity to insect and pathogen disturbances. Ann. Rev. Plant. Biol. 2015, 66, 547–569. [Google Scholar] [CrossRef] [PubMed]
- Wenk, M.; Apel, K.H. Die Regenerationsfähigkeit von durch Fraß des Kiefernspinners (Dendrolimus pini L.) und der Nonne (Lymantria monacha L.) geschädigten Kiefernbeständen in Brandenburg. In Die Kiefer im nordostdeutschen Tiefland—Ökologie und Bewirtschaftung, Kätzel, R., Möller, K., Löffler, S., Engel, J., Liero, K., Eds.; Eberswalder Forstliche Schriftenreihe; Ministerium für ländliche Entwicklung, Umwelt und Verbraucherschutz des Landes Brandenburg: Eberswalde, Germany, 2007; Volume 32, pp. 280–287. [Google Scholar]
- Mitton, J.B.; Ferrenberg, S.M. Mountain Pine Beetle Develops an Unprecedented Summer Generation in Response to Climate Warming. Am. Nat. 2012, 179, E163–E171. [Google Scholar] [CrossRef] [PubMed]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grüning, M.M.; Germeshausen, F.; Thies, C.; L.-M.-Arnold, A. Increased Forest Soil CO2 and N2O Emissions During Insect Infestation. Forests 2018, 9, 612. https://doi.org/10.3390/f9100612
Grüning MM, Germeshausen F, Thies C, L.-M.-Arnold A. Increased Forest Soil CO2 and N2O Emissions During Insect Infestation. Forests. 2018; 9(10):612. https://doi.org/10.3390/f9100612
Chicago/Turabian StyleGrüning, Maren Marine, Franziska Germeshausen, Carsten Thies, and Anne L.-M.-Arnold. 2018. "Increased Forest Soil CO2 and N2O Emissions During Insect Infestation" Forests 9, no. 10: 612. https://doi.org/10.3390/f9100612
APA StyleGrüning, M. M., Germeshausen, F., Thies, C., & L.-M.-Arnold, A. (2018). Increased Forest Soil CO2 and N2O Emissions During Insect Infestation. Forests, 9(10), 612. https://doi.org/10.3390/f9100612