Projected Future Distribution of Tsuga canadensis across Alternative Climate Scenarios in Maine, U.S


Abstract
:1. Introduction
2. Materials and Methods
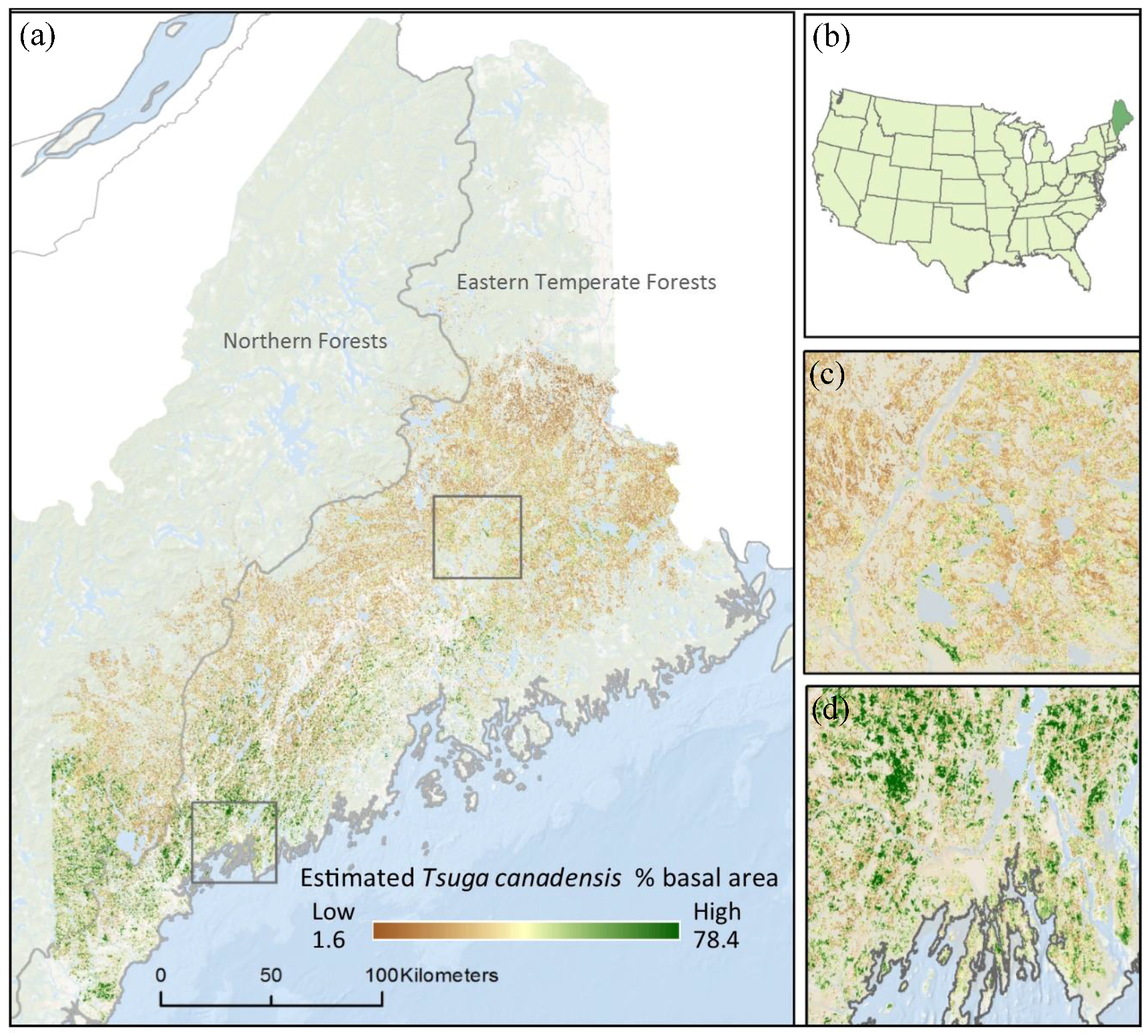
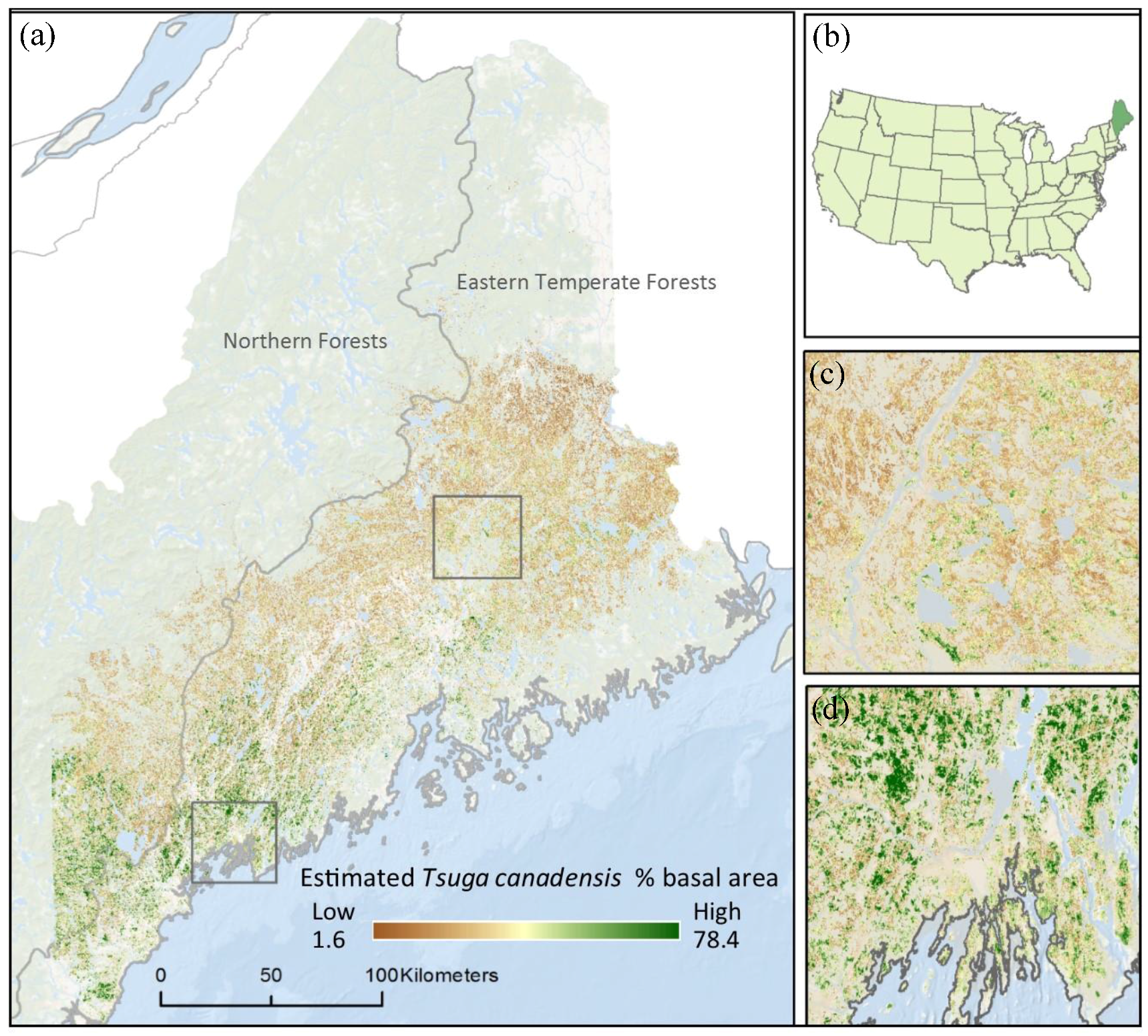
2.1. Study Area
2.2. Model Calibrations
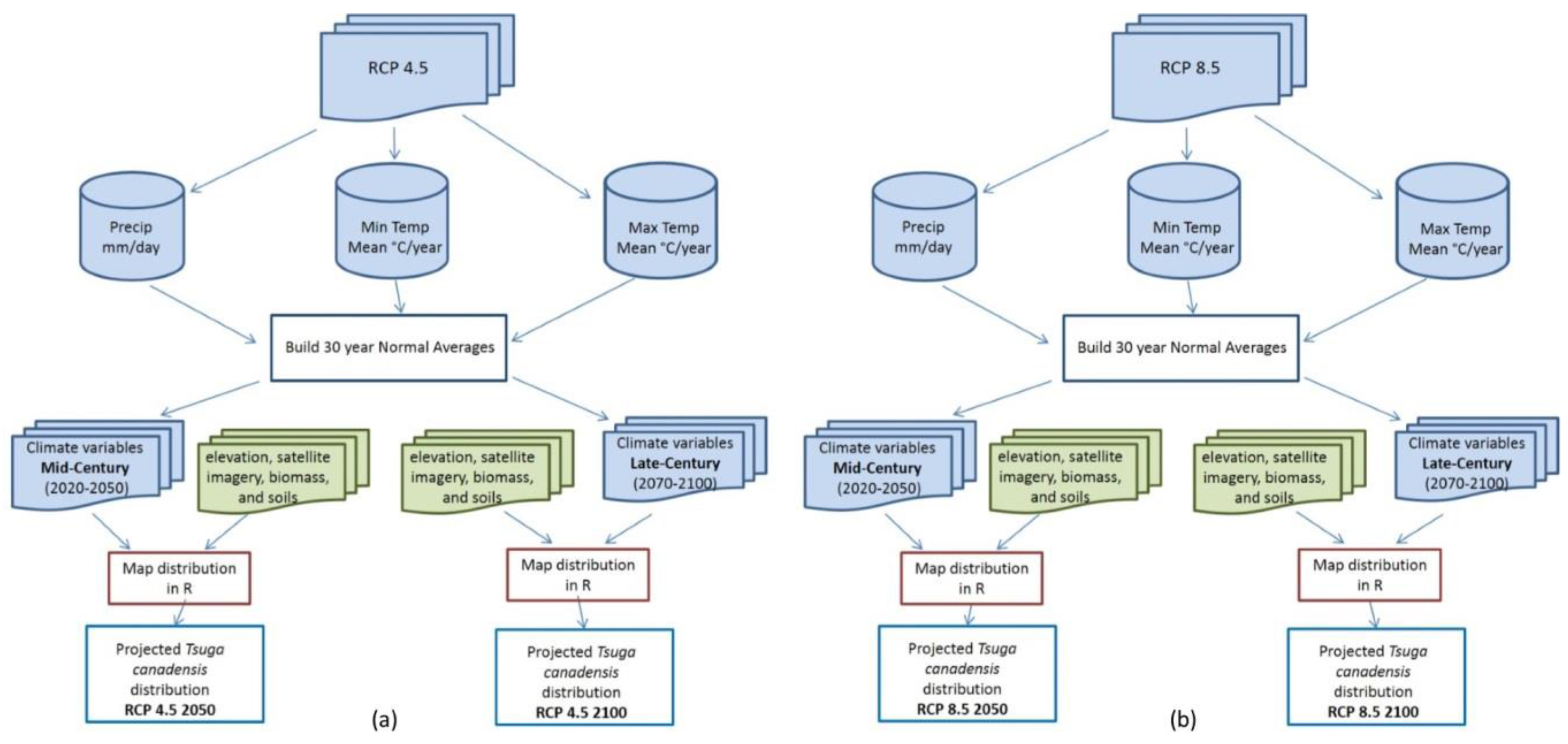
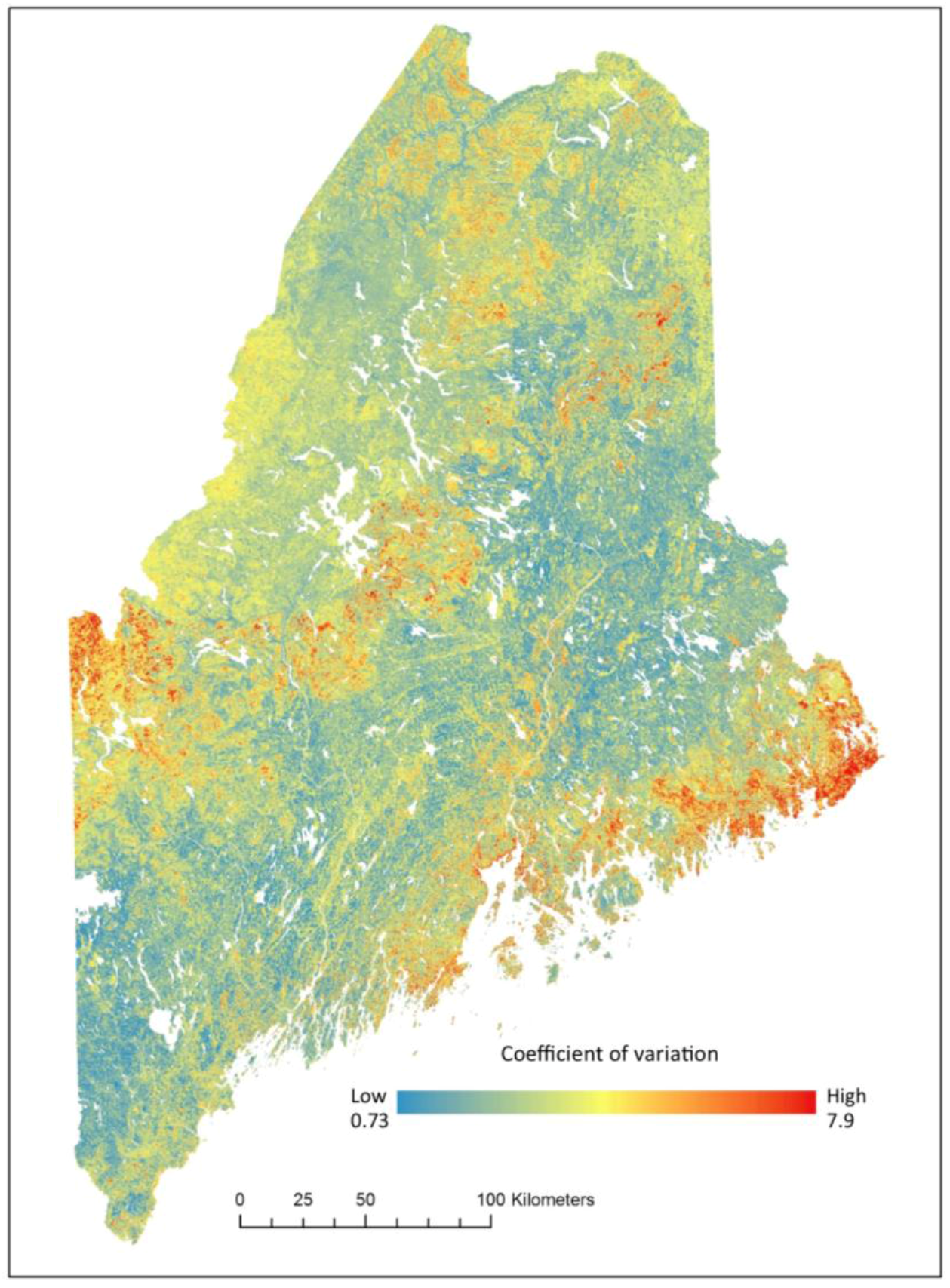
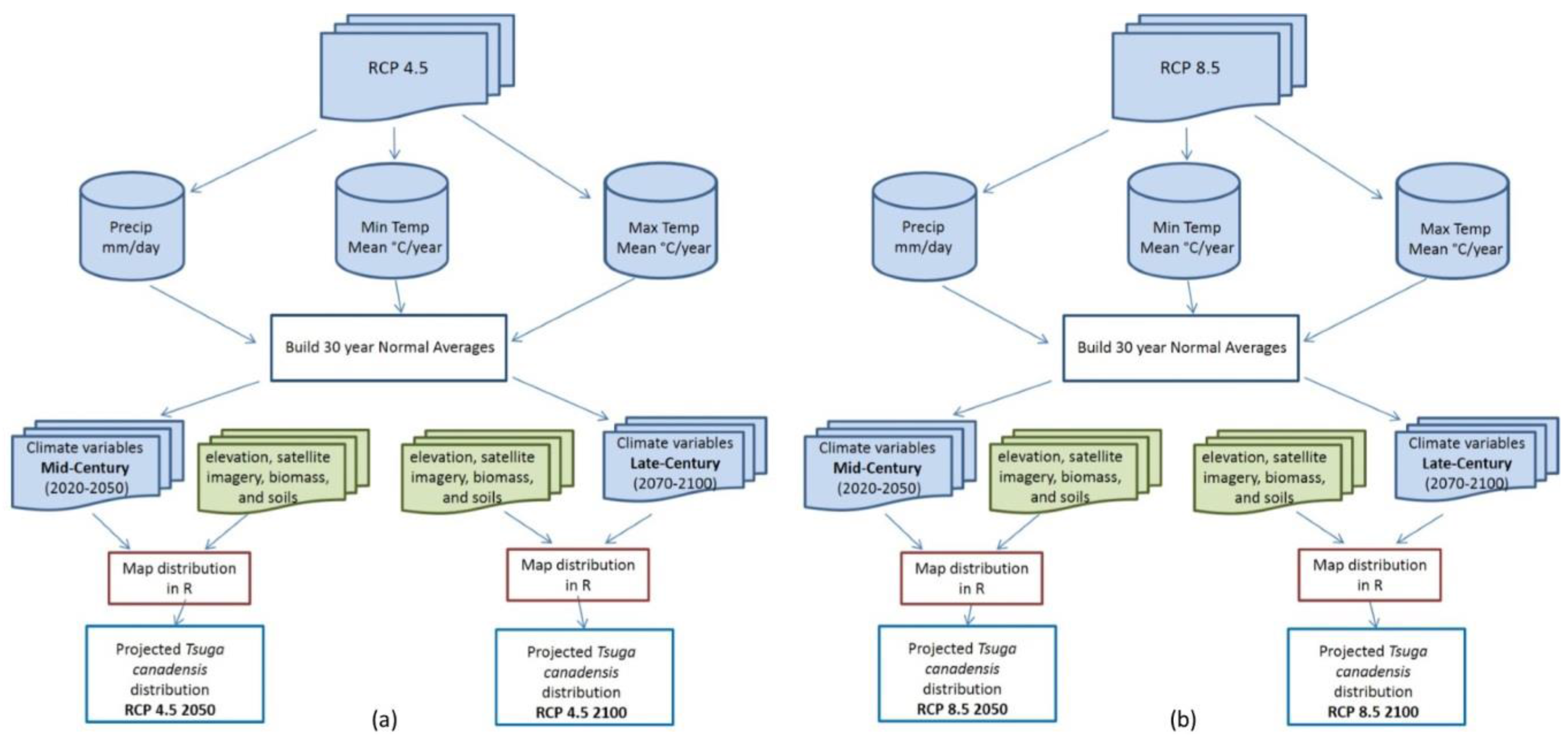
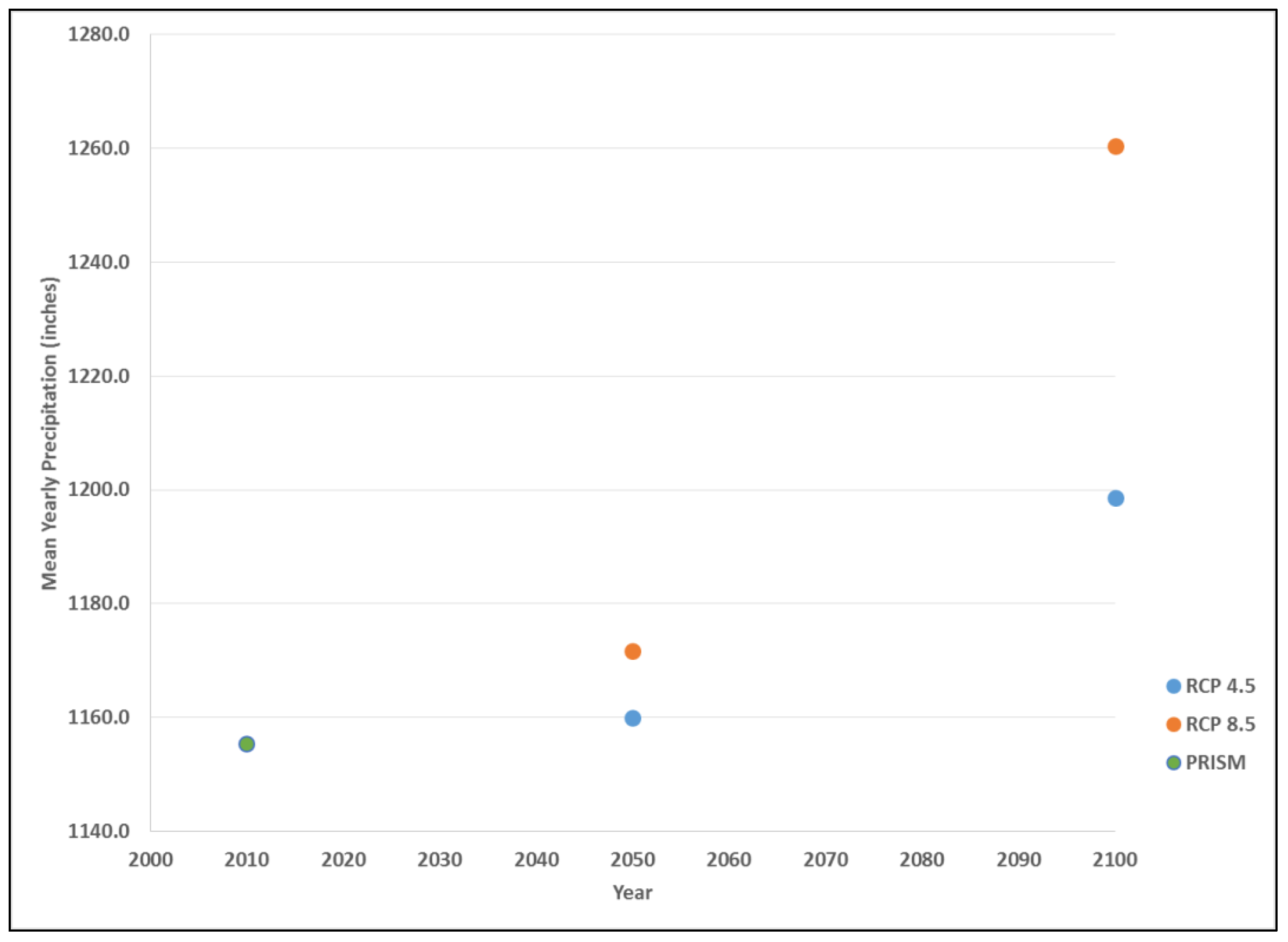
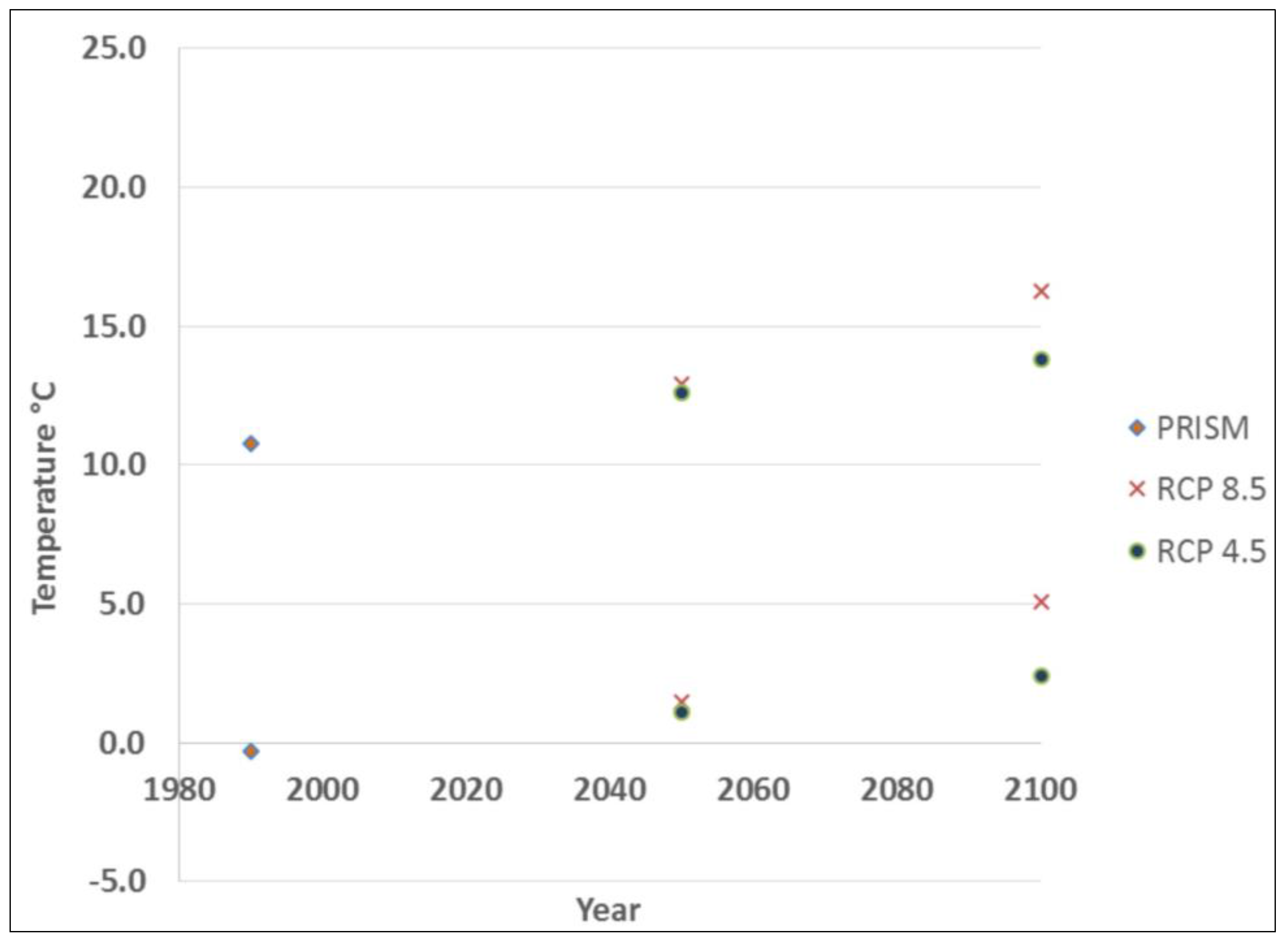
2.3. Data Processing
2.4. Model Processing
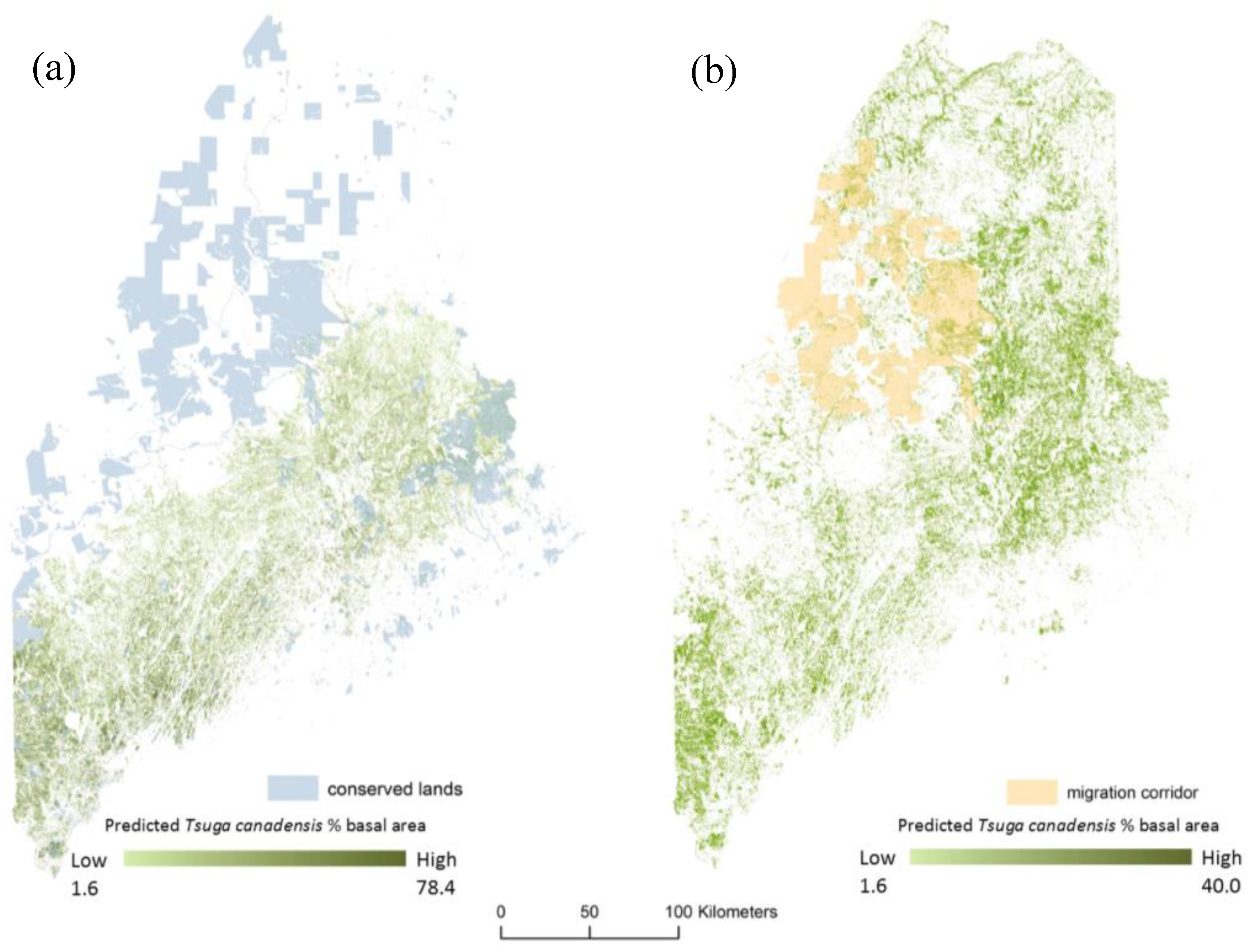
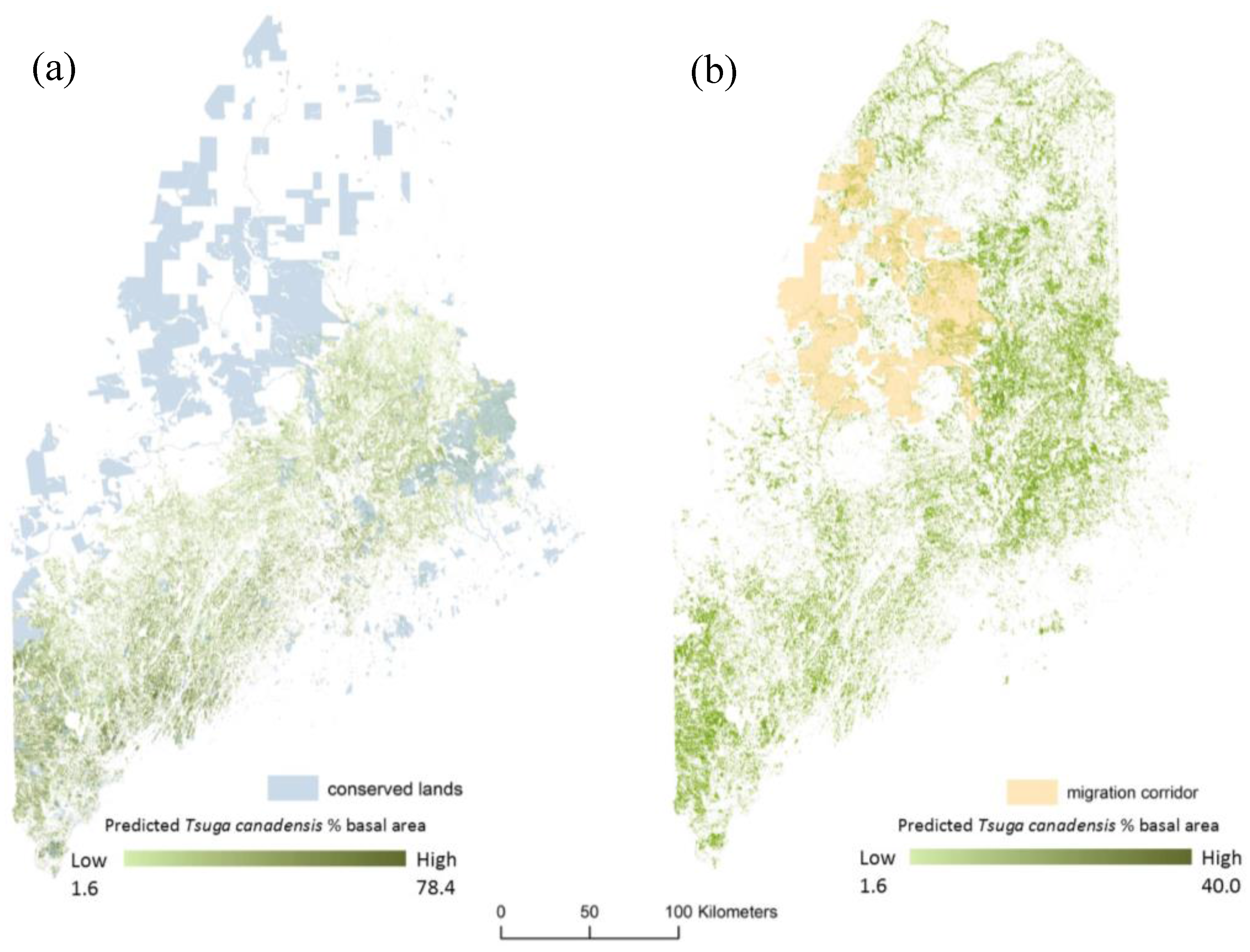
2.5. Migration Corridor
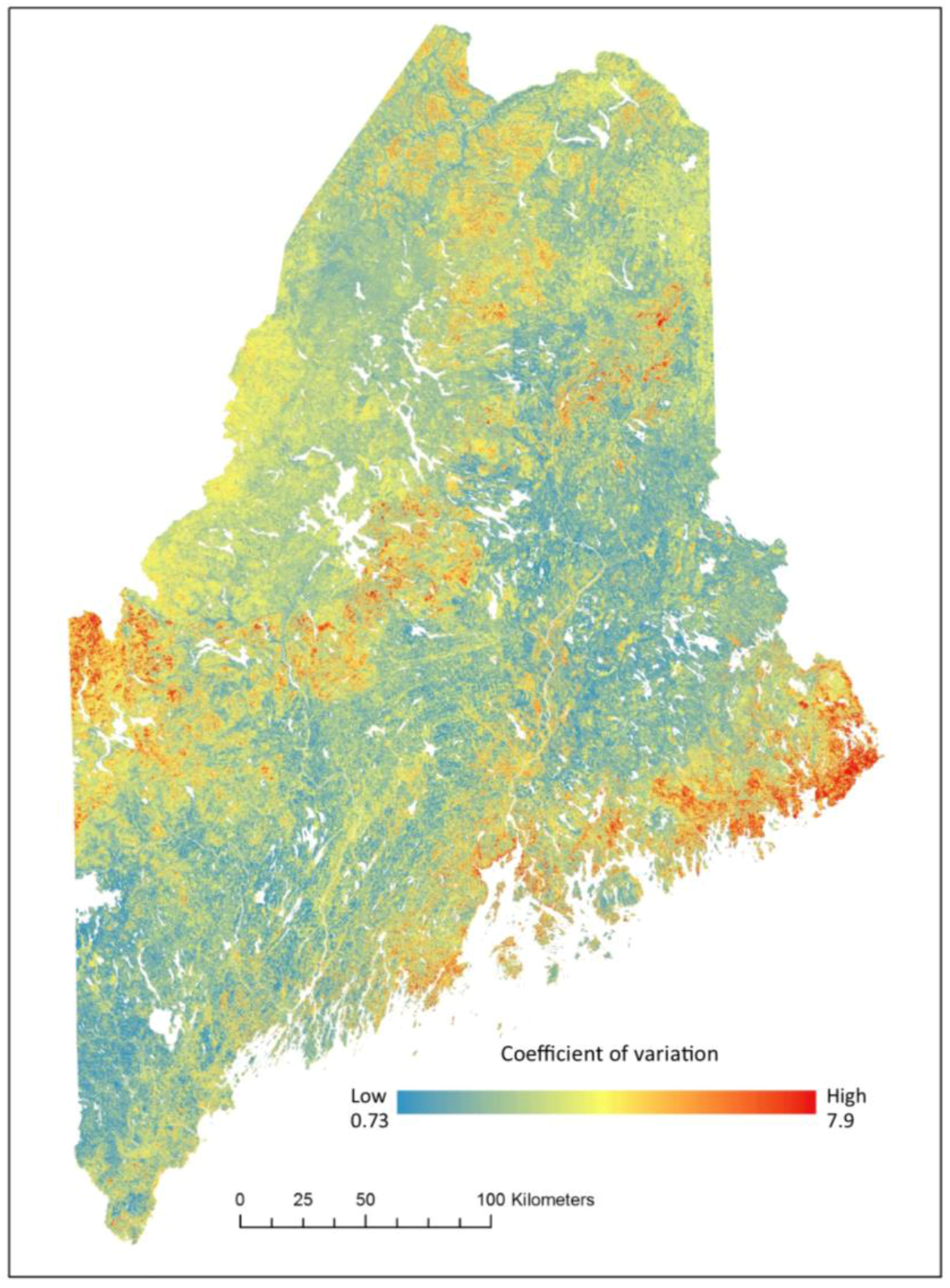
2.6. Validation
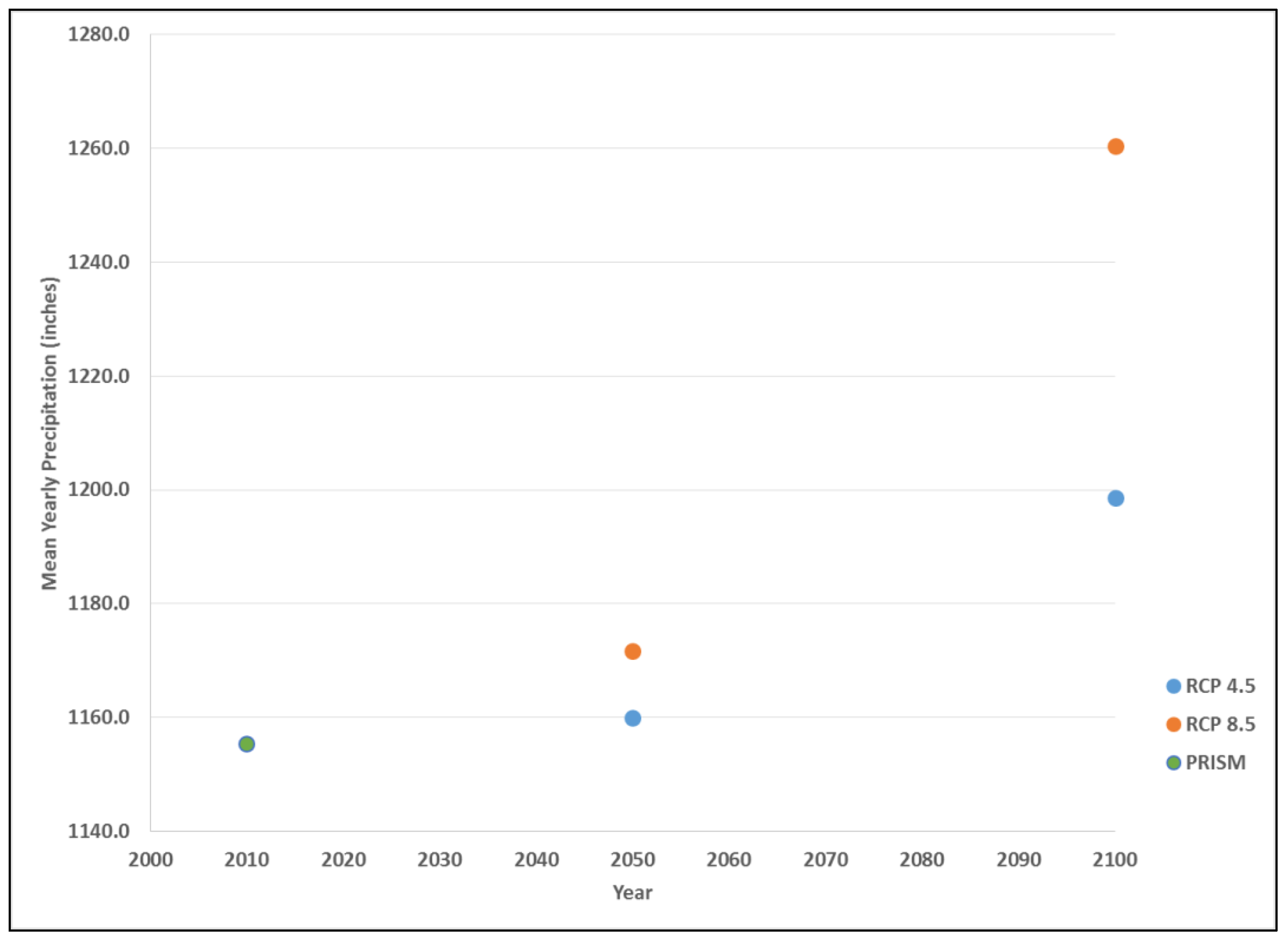
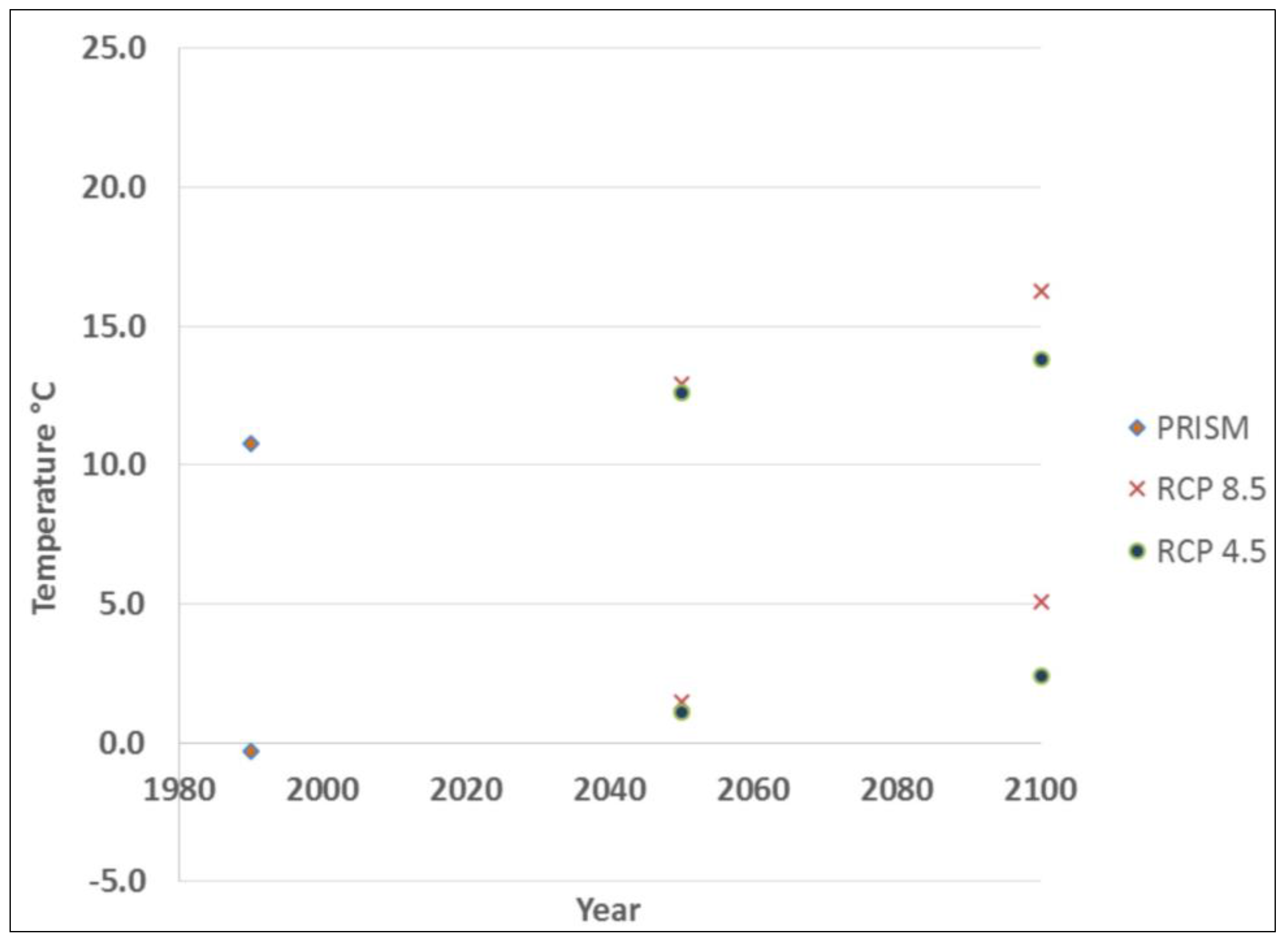
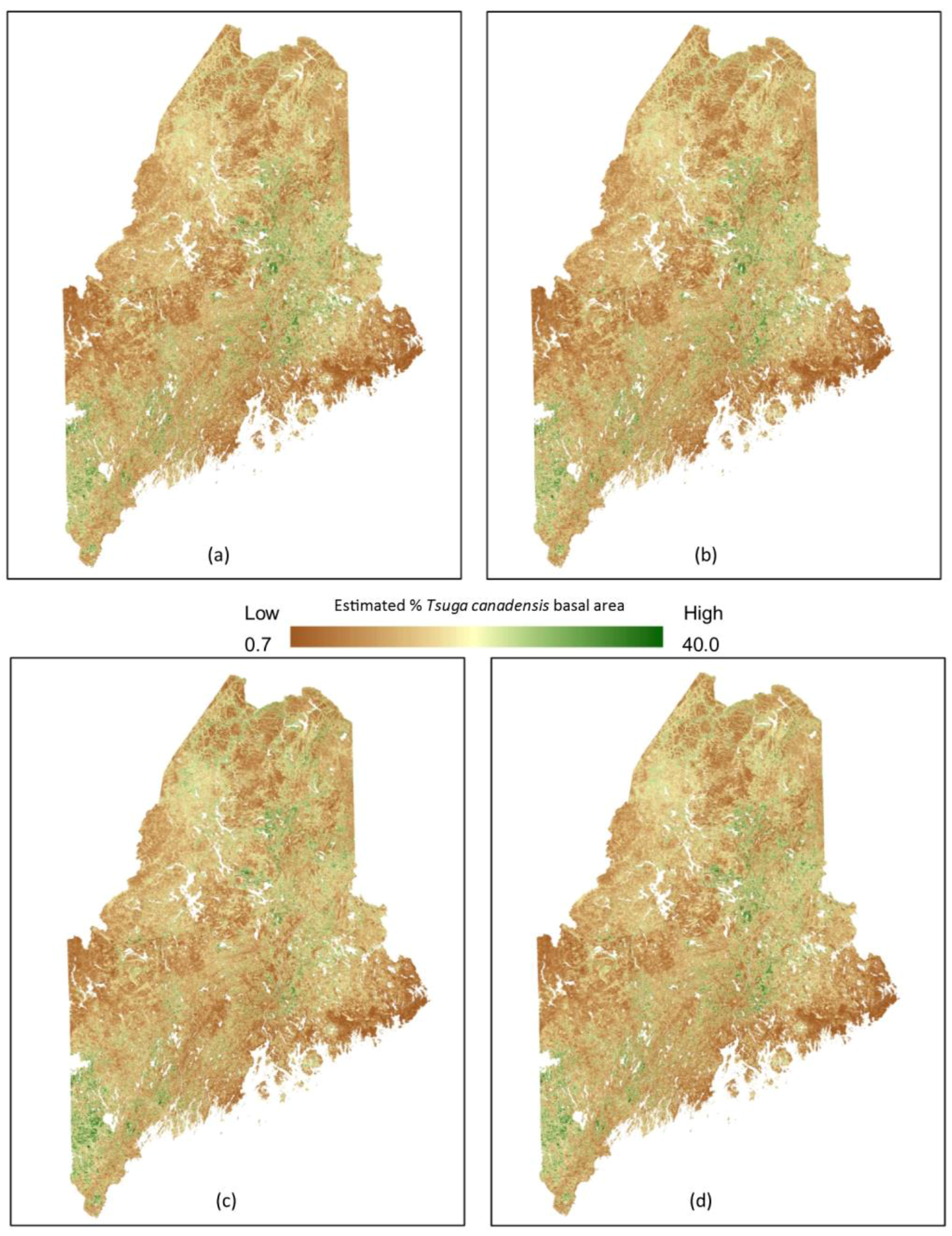
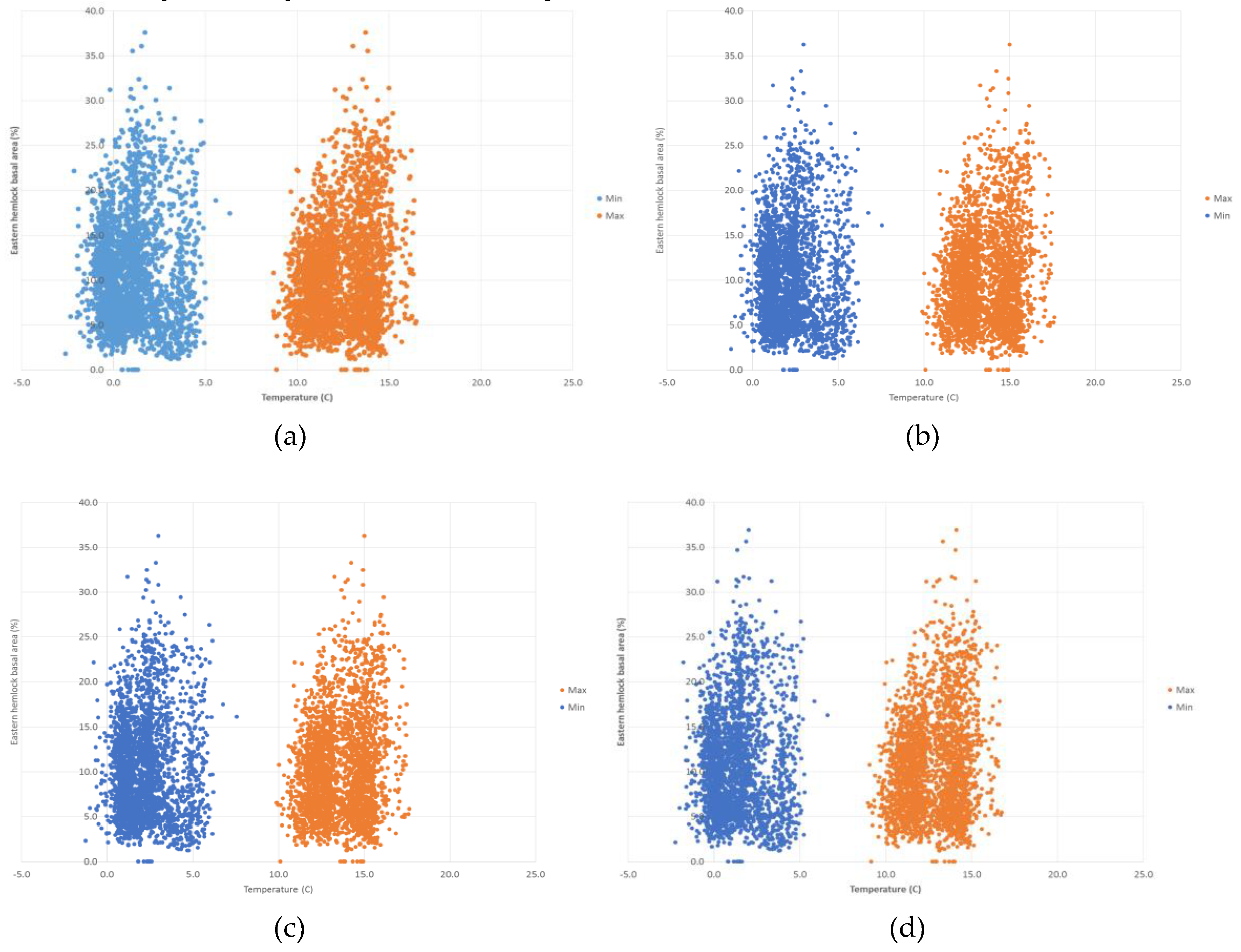
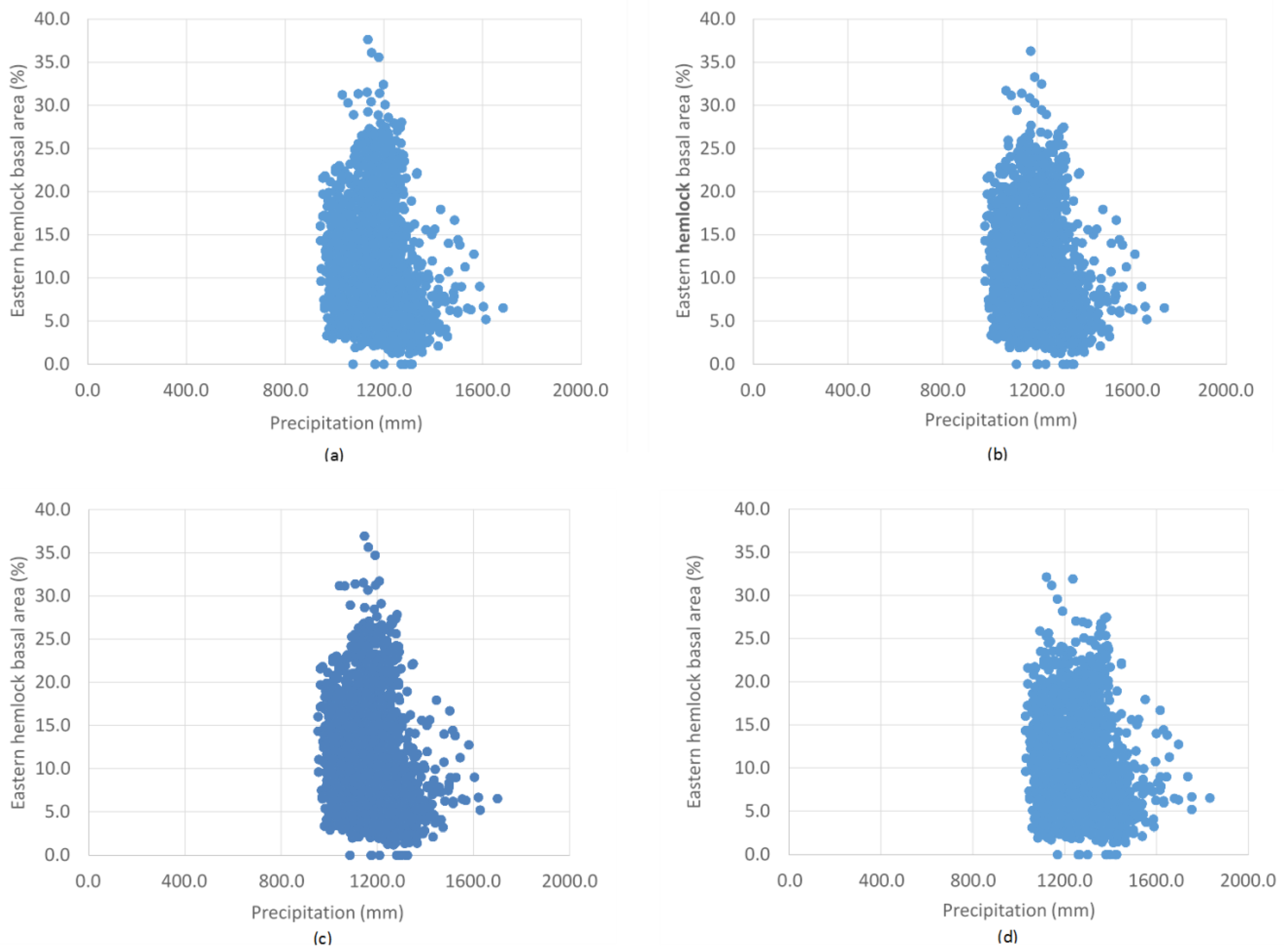
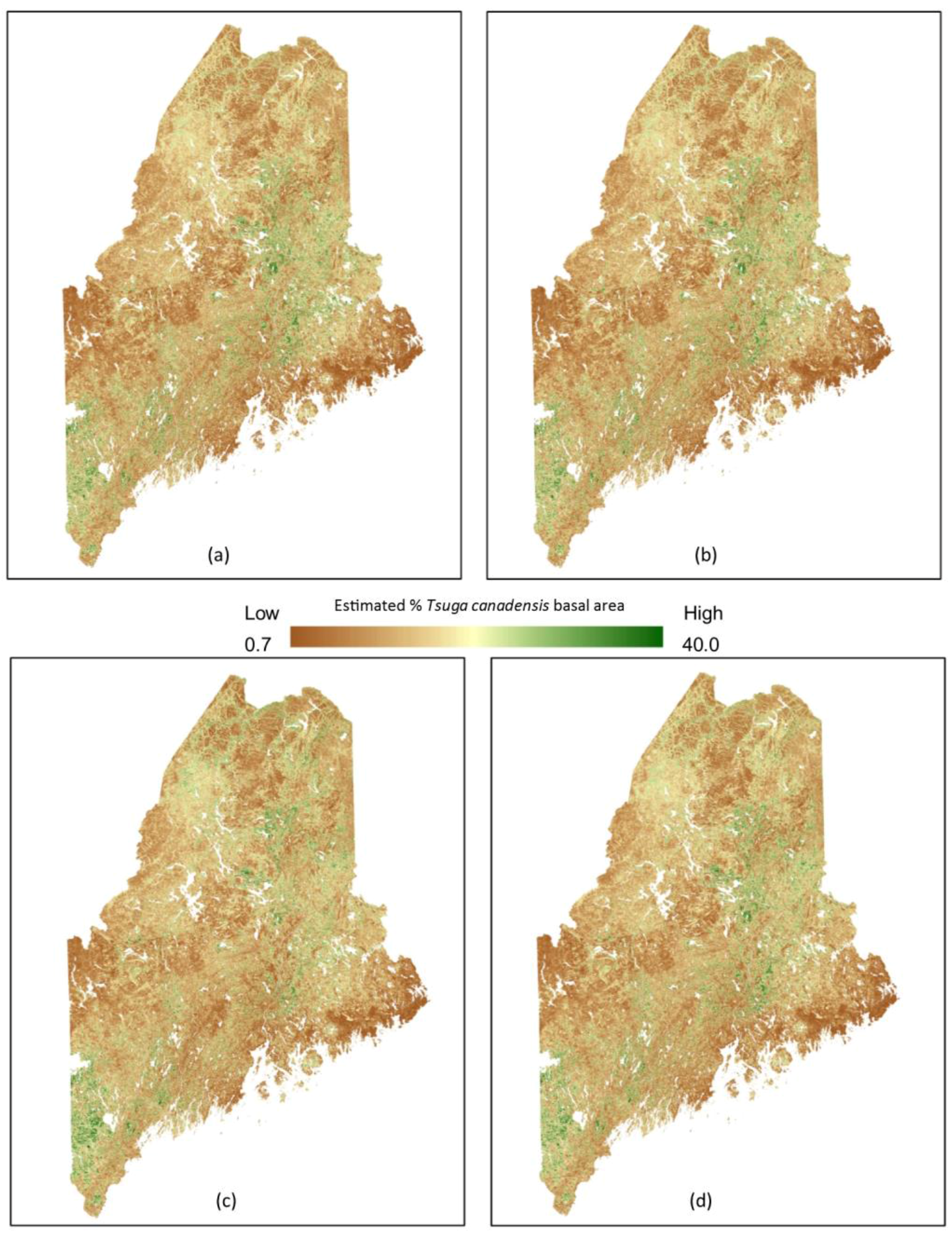
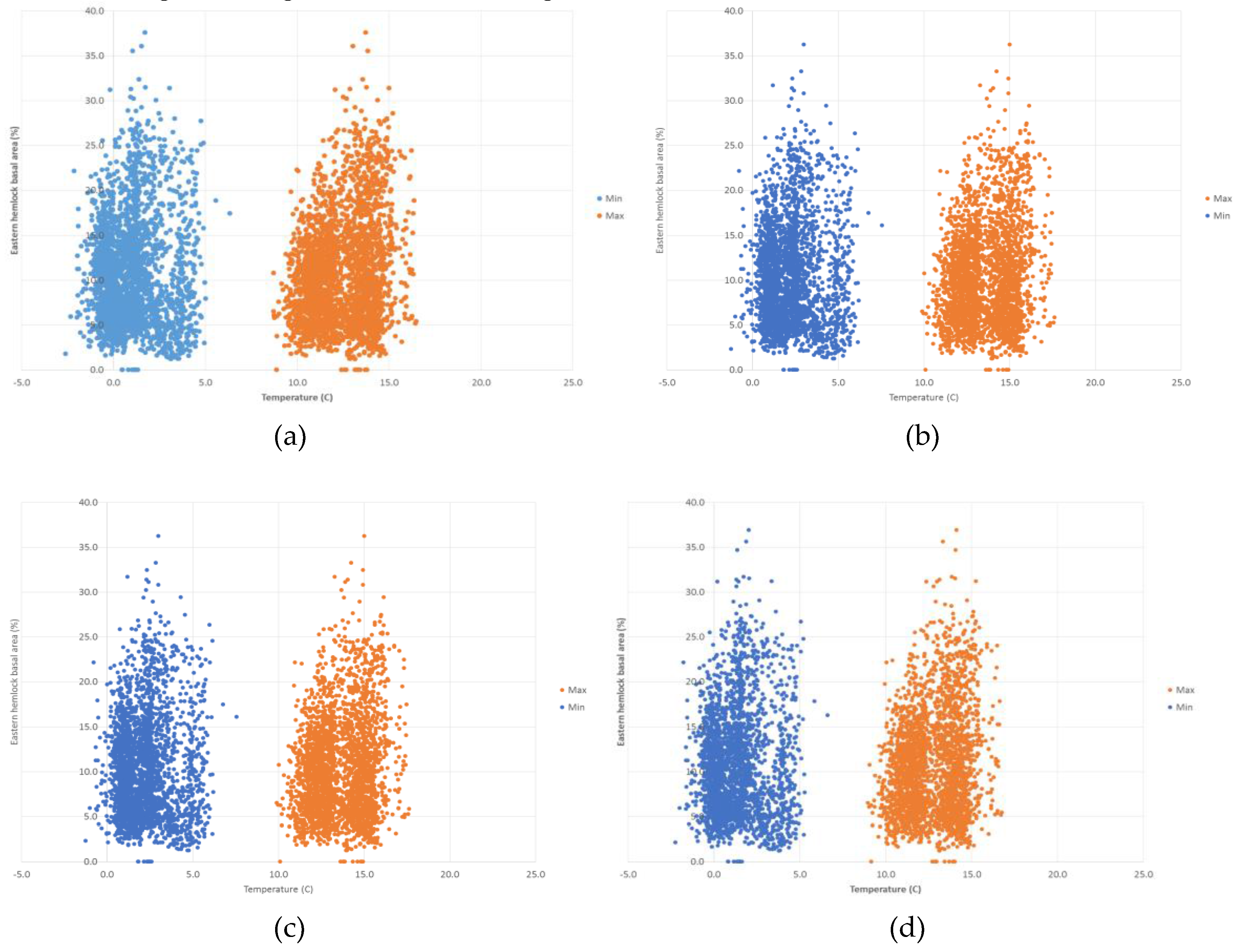
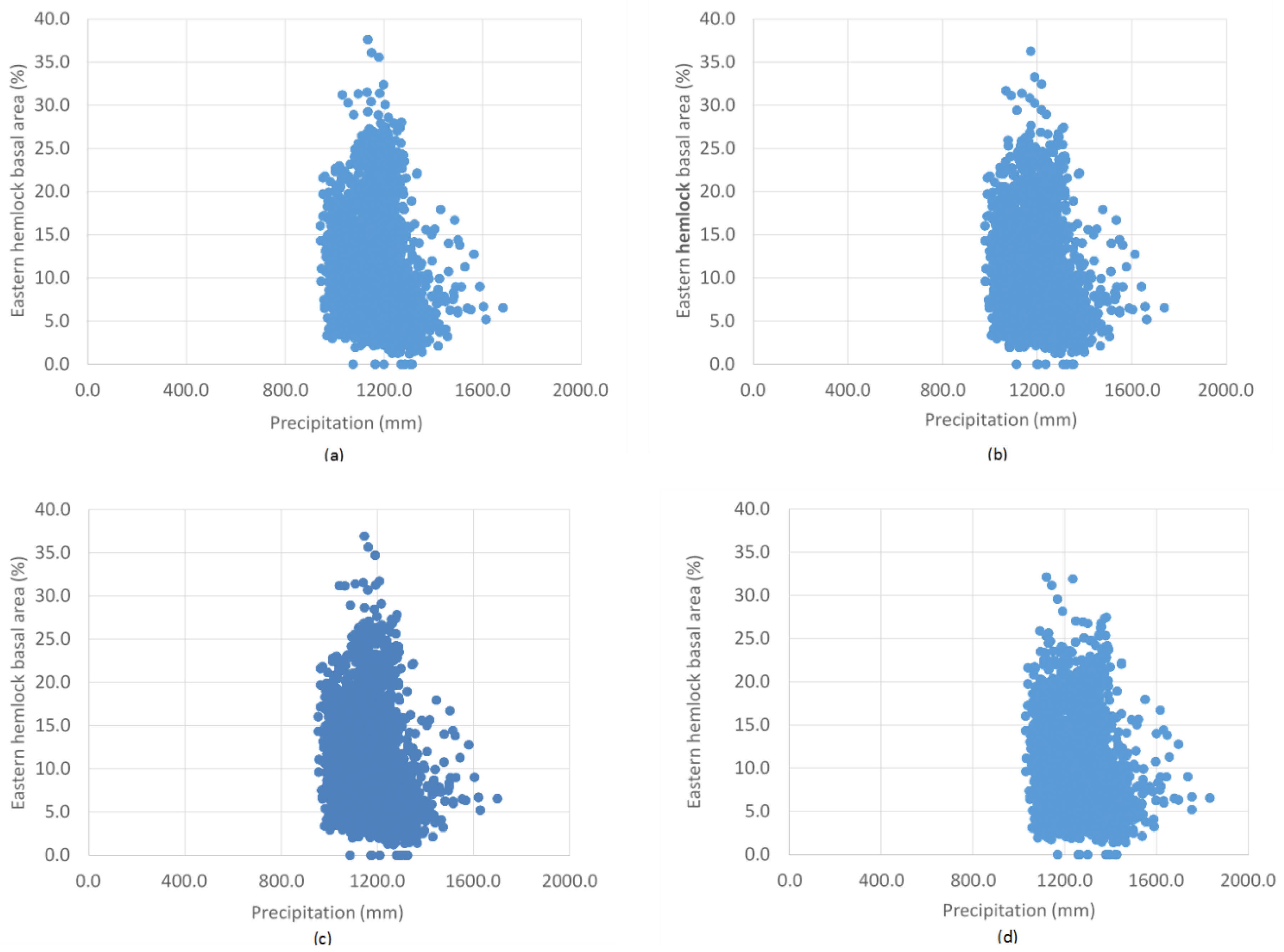
3. Results
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Campbell, J.L.; Rustad, L.E.; Boyer, E.W.; Christopher, S.F.; Driscoll, C.T.; Fernandez, I.J.; Groffman, P.M.; Houle, D.; Kiekbusch, J.; Magill, A.H.; et al. Consequences of climate change for biogeochemical cycling in forests of northeastern North America. Can. J. For. Res. 2009, 39, 264–284. [Google Scholar] [CrossRef]
- Weiskittel, A.R.; Crookston, N.L.; Radtke, P.J. Linking climate, gross primary productivity, and site index across forests of the western United States. Can. J. For. Res. 2011, 41, 1710–1721. [Google Scholar] [CrossRef]
- Chiang, J.-M.; Iverson, L.R.; Prasad, A.; Brown, K.J. Effects of climate change and shifts in forest composition on forest net primary production. J. Integr. Plant Biol. 2008, 50, 1426–1439. [Google Scholar] [CrossRef] [PubMed]
- Renwick, K.M.; Rocca, M.E. Temporal context affects the observed rate of climate-driven range shifts in tree species. Glob. Ecol. Biogeogr. 2014, 24, 44–51. [Google Scholar] [CrossRef]
- Gottesman, A.J.; Keeton, W.S. Regeneration responses to management for old-growth characteristics in northern hardwood-conifer forests. Forests 2017, 8, 45. [Google Scholar] [CrossRef]
- Orwig, D.A.; Foster, D.R. Forest response to the introduced hemlock woolly adelgid in southern New England, USA. J. Torrey Bot. Soc. 1998, 125, 60–73. [Google Scholar] [CrossRef]
- Orwig, D.A.; Foster, D.R.; Mausel, D.L. Landscape patterns of hemlock decline in New England due to the introduced hemlock woolly adelgid. J. Biogeogr. 2002, 29, 1475–1487. [Google Scholar] [CrossRef]
- Ellison, A.M.; Bank, M.S.; Clinton, B.D.; Colburn, E.A.; Elliott, K.; Ford, C.R.; Foster, D.R.; Kloeppel, B.D.; Knoepp, J.D.; Lovett, G.M.; et al. Loss of foundation species: Consequences for the structure and dynamics of forested ecosystems. Front. Ecol. Environ. 2005, 3, 479–486. [Google Scholar] [CrossRef]
- Stadler, B.; Müller, T.; Orwig, D.; Cobb, R. Hemlock woolly adelgid in New England forests: Canopy impacts transforming ecosystem processes and landscapes. Ecosystems 2005, 8, 233–247. [Google Scholar] [CrossRef]
- Albani, M.; Moorcroft, P.R.; Ellison, A.M.; Orwig, D.A.; Foster, D.R. Predicting the impact of hemlock woolly adelgid on carbon dynamics of eastern United States forests. Can. J. For. Res. 2010, 40, 119–133. [Google Scholar] [CrossRef]
- Rogers, B.M.; Jantz, P.; Goetz, S.J. Vulnerability of eastern US tree species to climate change. Glob. Chang. Biol. 2017, 23, 3302–3320. [Google Scholar] [CrossRef] [PubMed]
- Zolkos, S.; Jantz, P.; Cormier, T.; Iverson, L.; McKenney, D.; Goetz, S. Projected tree species redistribution under climate change: Implications for ecosystem vulnerability across protected areas in the eastern United States. Ecosystems 2015, 18, 202–220. [Google Scholar] [CrossRef]
- Iverson, L.R.; Prasad, A.M.; Matthews, S.N.; Peters, M. Estimating potential habitat for 134 eastern US tree species under six climate scenarios. For. Ecol. Manag. 2008, 254, 390–406. [Google Scholar] [CrossRef]
- Maria, K.J.; Louis, R.I.; Jon, F.; Stephen, D.H.; Matt, D.; Scott, T.; Brad, H.; Christopher, W.S. Assessing stand-level climate change risk using forest inventory data and species distribution models. J. For. 2017, 115, 222–229. [Google Scholar]
- International Institute for Applied Systems Analysis (IIASA). Encyclopedia of Operations Research and Management Science; IIASA: Laxenburg, Austria, 2013; pp. 805–806. [Google Scholar]
- Ahmed, K.F.; Wang, G.; Silander, J.; Wilson, A.M.; Allen, J.M.; Horton, R.; Anyah, R. Statistical downscaling and bias correction of climate model outputs for climate change impact assessment in the U.S. northeast. Glob. Planet. Chang. 2013, 100, 320–332. [Google Scholar] [CrossRef]
- Hayhoe, K.; Wake, C.P.; Huntington, T.G.; Luo, L.; Schwartz, M.D.; Sheffield, J.; Wood, E.; Anderson, B.; Bradbury, J.; DeGaetano, A.; et al. Past and future changes in climate and hydrological indicators in the US Northeast. Clim. Dyn. 2007, 28, 381–407. [Google Scholar] [CrossRef]
- Dunckel, K.; Weiskittel, A.; Fiske, G.; Sader, S.A.; Latty, E.; Arnett, A. Linking remote sensing and various site factors for predicting the spatial distribution of eastern hemlock occurrence and relative basal area in Maine, USA. For. Ecol. Manag. 2015, 358, 180–191. [Google Scholar] [CrossRef]
- PRISM Climate Group. PRISM Climate Data; PRISM Climate Group: Oregon State University, Corvallis, OR, USA, 2004. [Google Scholar]
- Fiske, G. Python Code for netCDF Processing. Available online: http://www.prism.oregonstate.edu (accessed on 2 June 2017).
- Walker, W.S.; Kellndorfer, J.M.; LaPoint, E.; Hoppus, M.; Westfall, J. An empirical InSAR-optical fusion approach to mapping vegetation canopy height. Remote Sens. Environ. 2007, 109, 482–499. [Google Scholar] [CrossRef]
- Briemen, L. Random Forests. Mach. Learn. 2001, 45, 5–23. [Google Scholar]
- Liaw, A.; Weiner, M. Classification and regression by random forest. R News 2002, 2, 18–22. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Freeman, E. ModelMap: An R Package for Modeling and Map Production Using Random Forest and Stochastic Gradient Boosting; USDA Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 2009. [Google Scholar]
- Maine Office of GIS. Maine Office of GIS Data Catalog. Available online: http://www.maine.gov/megis (accessed on 2 June 2017).
- Baccini, A.; Goetz, S.J.; Walker, W.S.; Laporte, N.T.; Sun, M.; Sulla-Menashe, D.; Hackler, J.; Beck, P.S.A.; Dubayah, R.; Friedl, M.A.; et al. Estimated carbon dioxide emissions from tropical deforestation improved by carbon-density maps. Nat. Clim. Chang. 2012, 2, 182–185. [Google Scholar] [CrossRef]
- Coops, N.C.; Waring, R.H.; Plowright, A.; Lee, J.; Dilts, T.E. Using remotely-sensed land cover and distribution modeling to estimate tree species migration in the Pacific northwest region of north America. Remote Sens. 2016, 8, 65. [Google Scholar] [CrossRef]
- Ashcroft, M.B.; King, D.H.; Raymond, B.; Turnbull, J.D.; Wasley, J.; Robinson, S.A. Moving beyond presence and absence when examining changes in species distributions. Glob. Chang. Biol. 2017, 23, 2929–2940. [Google Scholar] [CrossRef] [PubMed]
- Godman, R.M.; Lancaster, K. Eastern Hemlock. Available online: https://www.na.fs.fed.us/spfo/pubs/silvics_manual/Volume_1/tsuga/canadensis.htm (accessed on 2 June 2017).
- Sittaro, F.; Paquette, A.; Messier, C.; Nock, C.A. Tree range expansion in eastern North America fails to keep pace with climate warming at northern range limits. Glob. Chang. Biol. 2017, 23, 3292–3301. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tsuga canadensis (% basal area) | Year | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 2010 | 2050 | 2100 | ||||||||
| RCP 4.5 | RCP 8.5 | RCP 4.5 | RCP 8.5 | |||||||
| km2 | m2 ha−1 | km2 | m2 ha−1 | km2 | m2 ha−1 | km2 | m2 ha−1 | km2 | m2 ha−1 | |
| Not present | 64,319 | 0.804 | 2 | 0 | 2 | 0 | 1 | 0 | 0 | 0 |
| 1–15 | 6689 | 0.084 | 64,722 | 0.809 | 65,220 | 0.815 | 66,465 | 0.831 | 68,484 | 0.856 |
| 16–30 | 6703 | 0.084 | 14,392 | 0.180 | 13,933 | 0.174 | 12,770 | 0.160 | 10,820 | 0.135 |
| >30 | 1622 | 0.020 | 217 | 0.003 | 178 | 0.002 | 97 | 0.001 | 29 | 0.000 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dunckel, K.; Weiskittel, A.; Fiske, G. Projected Future Distribution of Tsuga canadensis across Alternative Climate Scenarios in Maine, U.S. Forests 2017, 8, 285. https://doi.org/10.3390/f8080285
Dunckel K, Weiskittel A, Fiske G. Projected Future Distribution of Tsuga canadensis across Alternative Climate Scenarios in Maine, U.S. Forests. 2017; 8(8):285. https://doi.org/10.3390/f8080285
Chicago/Turabian StyleDunckel, Kathleen, Aaron Weiskittel, and Greg Fiske. 2017. "Projected Future Distribution of Tsuga canadensis across Alternative Climate Scenarios in Maine, U.S" Forests 8, no. 8: 285. https://doi.org/10.3390/f8080285
APA StyleDunckel, K., Weiskittel, A., & Fiske, G. (2017). Projected Future Distribution of Tsuga canadensis across Alternative Climate Scenarios in Maine, U.S. Forests, 8(8), 285. https://doi.org/10.3390/f8080285