Abstract
The success of some invasive tree species is attributed, in part, to high fecundity in the form of sexual propagules. If invasive trees produce more seed annually than co-occurring native trees, they will have a greater ability to disperse and establish across the landscape. In this study, seed production of female Ailanthus trees was investigated to determine (1) reproductive age limits; (2) annual and cumulative seed output; and (3) seed viability. Existing data on Ailanthus seed production were combined with a novel dataset to compare variability in seed production and explore relationships with tree diameter and age. Results from this study showed Ailanthus’ reproductive window is exceptional, spanning more than a century, with seed viability exceeding 65% from a 104-year-old individual. Germination studies and complementary tetrazolium assays also confirmed high propagule viability from a 7-year-old Ailanthus and supports tetrazolium assays as a proxy for germination studies. Not only can individual Ailanthus produce >1 million seeds annually, but a significant relationship exists between seed production and tree diameter. Using this relationship, cumulative seed production in individual Ailanthus can reach ca. 10 and 52 million seeds over a 40-year and 100-year period, respectively. This study provides a comprehensive investigation of various facets of the reproductive potential of Ailanthus.
1. Introduction
The invasive Ailanthus altissima (Mill.) Swingle (tree-of-heaven), a member of the mostly tropical Quassia family (Simaroubaceae), hereafter referred to as Ailanthus, was first introduced into the United States from England into Philadelphia, Pennsylvania in 1784. Early propagation by botanists and nurserymen was exclusively vegetative, which limited both the spread and widespread adoption of Ailanthus until ca. 1820, when seed-bearing (gynoecious) Ailanthus became established in the eastern United States [1]. By the late 1800s, Ailanthus’ ability to spread naturally from seed was widely acknowledged [2]. At that time, seed-producing females were strongly promoted in ornamental plantings [3] and their seed spread naturally along well-traveled railway corridors [1]. Since that time, sexual reproduction and subsequent movement of seed across the United States along transportation corridors has played an important role in shaping Ailanthus’ current population structure [4]. Ailanthus now occurs in more than 40 states, mostly as an urban and roadside weed, and is especially abundant in the Mid-Atlantic where it has resided for ca. 200 years [1,4,5]. Despite its long residency, widespread invasion of Ailanthus into eastern forests is relatively recent, with most invasions in Pennsylvania following salvage logging from widespread gypsy moth defoliations in the late twentieth century [1]. In Ohio, prescribed burning to enhance oak regeneration has also facilitated spread of Ailanthus in several areas of the state [6]. The impact of such large-scale invasions in forests and their adverse effects on native species richness and incidence have only been recently elucidated [7,8,9,10]. Invasions of Ailanthus along transportation corridors have long been recognized, but only recently have studies assessed the mechanisms of spread in these environments [11,12,13]. Researchers are now beginning to appreciate the role that roads play in facilitating forest invasions [13,14].
Previous studies on Ailanthus seed production have reported reproductive outputs from several hundred thousand [15,16] to several million seeds in a single year, with large inter-annual variation and tree size-dependent effects [17,18]. Martin and Canham [18] reported Ailanthus out-produced other exotic invasive counterparts (e.g., Acer platanoides L.) and co-occurring native tree species by several orders of magnitude, but without the periodic seed crop failures common among many native species. Similarly, Cabra-Rivas and Castro-Diez [16] reported that Ailanthus showed greater seed viability than the invasive Robinia pseudoacacia L. and native Fraxinus angustifolia Vahl in Spain. Moreover, Ailanthus is unexpectedly fecund even in environments presumably less favorable for invasion such as closed canopy forests [9,18] or as germinants following submergence in water [19,20]. Recent studies have shown certain morphological characteristics (e.g., samara width) affect water dispersal positively, but affect wind dispersal negatively [11].
The recent discovery that Ailanthus can produce seed beyond 100 years of age [1], coupled with the fact that almost no data exists regarding the species’ mean reproductive capacity and age-related seed production limitations, has raised questions about Ailanthus’ actual reproductive outputs.
In this study, the minimum and maximum age at which female Ailanthus trees produce seed and to what extent tree and seed age influences seed viability was evaluated. Secondly, the maximum and mean annual seed outputs for Ailanthus were quantified. Finally, the cumulative number of seeds produced during Ailanthus’ reproductive lifetime was modeled, and relationships between tree DBH (diameter at breast height, 1.4 m above soil line) and seed production were explored.
2. Materials and Methods
2.1. Seed Production
To study age- and size-dependent effects on seed reproduction as well as intra-specific variation within reproduction, 55 seed-bearing Ailanthus trees were sampled at various locations along transportation corridors throughout south-central Pennsylvania in August 2011 and 2012 (Table S1). Of the 55 sampled trees, 54 were destructively sampled (felled) for seed quantification and age determination. The location of the largest tree precluded removal and was instead cored with an increment borer to determine age [1]. Ten average-sized seed clusters were removed from this tree and six other additional destructively sampled forest trees for quantification. Total clusters were counted for total seed estimations, since it was impractical to collect all the seeds from these seven large trees. Small diameter seed-producing Ailanthus trees were targeted when present since there was insufficient/conflicting data on the minimum reproductive age of Ailanthus [21,22,23]. In addition, small diameter seed-producing trees were generally abundant along transportation corridors throughout the sampling area, thus providing a robust sampling population. Presumably some of these small diameter trees were sprouts arising from pre-existing root systems but could not be differentiated from seed-origin plants since roots were not excavated.
For each tree included in the study, one cross-section disk was removed at either DBH or ground level (for trees <1.4 m in height), or was sampled at DBH with an increment borer (Haglof Inc., Madison, MS, USA) to determine age. For felled trees, all available seed from the current year was collected into pre-labeled plastic bags. If seed was inadvertently released during felling, estimates of unrecovered seed were made in 5% intervals and recorded. For multi-stemmed trees (trees whose trunk forked below DBH), a cross-section was taken from each stem to compare ages and diameters of the conjoined stems. Diameters (cm) for the multi-stemmed trees were individually converted into basal area (BA = DBH2 × 0.00007854 (constant)), summed, and converted to a single DBH value (DBH =
). Basal area is defined as the cross-sectional area (m2) of a tree stem at DBH.
In the lab, Ailanthus seed from each tree was air-dried for up to three weeks, after which seeds were placed into paper bags and maintained at room temperature. Mean seed weight (mg) including the samara was determined for 25 randomly selected seeds from each tree. Seed with damaged samaras were excluded from weight measurements. Total dried seed was weighed and number of seeds per tree estimated by dividing total seed weight by mean seed weight for each tree. For trees where only 10 seed clusters were sampled (n = 7), mean seed count was determined and multiplied by the number of seed clusters estimated per tree.
In addition to trees sampled specifically for this study, data from 21 seed-bearing trees from France [15] and two historic seed-bearing trees from Pennsylvania [17], whose diameters and seed production were previously reported, were included in the master dataset to permit comparisons and analyses. Since ages of sampled Ailanthus were not reported for these two studies, only relationships between DBH and seed production were explored.
2.2. Germination Studies to Assess Seed Viability Based on Tree and Seed Age
For tree-age seed viability studies, 64 seeds with samaras per seed source were placed in a 15.2 × 25.4 cm plastic bag containing ca. 900 mL of potting soil mix (Sun Gro Horticulture Canada Ltd., Vancouver, BC, Canada) and 50 mL tap water. Seed was collected in late summer 2016 from two locations from Pennsylvania and five locations from West Virginia (Table 1, Table S1). Trees included in the study ranged from 7 to 104 years old (Table 1). Concurrently, 64 seeds with samaras per seed source for each of two seed-producing trees and 43 and 40 seeds per seed source for two additional seed-producing trees previously collected in Pennsylvania from 2008–2010 from female Ailanthus trees [1,24] were treated similarly (Table 1). For both germination studies, the seed/soil mix was maintained at 1.7 °C for 28 days, as previously reported [24], after which seeds were placed into seed flats containing additional potting mix, transferred to a greenhouse, and misted daily until initial germination was observed. Upon germination, individual seedling wells were labeled with date of germination. As new seedlings emerged, additional wells were labeled for up to 4-weeks, after which no additional seeds germinated. Total germinants were counted and divided by the number of total seeds evaluated.

Table 1.
Seed viability status for current and historic seed sources of Ailanthus altissima.
2.3. Tetrazolium Assay to Evaluate Seed Viability Based on Seed and Tree Age
As a complementary experiment to the germination study, a blue tetrazolium chloride (TZ) test [25] was performed on three of the same 2016 seed collections used in the germination studies. Tree ages were 7, 20, and 104 years. Tetrazolium chloride is a redox indicator commonly used to confirm cellular respiration and has been used frequently as a proxy for germination studies [25]. One hundred seeds from each of three seed collections sampled in 2016 were randomly selected. The experiment was replicated twice. Seeds encased in their samaras were soaked for 14 h in distilled sterile water, after which seeds were cut in half, longitudinally bisecting the seed radical (embryo). If a seed was determined to be deteriorated or damaged during bisectioning, it was excluded from the TZ test but included in the overall count. For seeds that appeared to be sound, half of the seed was placed into a 96-well microtiter plate containing 1% tetrazolium chloride solution (Alfa Aesar Haverhill, MA, USA). Individual wells were capped and wrapped in foil due to the light sensitivity of the assay. Each plate was incubated in a water bath at 40 °C for 3 h, after which seeds were removed and scored based on the intensity of staining to the seed radicals. Following the TZ test, seeds were classified as positive (staining of radical only or complete staining of radical and hypocotyl) or negative (no staining) (Figure 1).

Figure 1.
Ailanthus seed embryo challenged with blue tetrazolium chloride. A positive blue tetrazolium chloride (TZ) test resulted in (A) complete staining of radical and hypocotyl or (B) staining of radical only, whereas a negative TZ test resulted in (C) no staining.
2.4. Statistical Analyses
To examine age-DBH relationships and seed production-DBH relationships, two regression analyses were performed. For the age-diameter linear regression, datasets were analyzed individually and combined. External validation methods (R2) were used to assess model validity. Assumptions of linearity and homoscedasticity (equal variances) were tested and data was square root transformed to meet required assumptions. To analyze DBH-seed production relationships, data was Ln transformed, after which a goodness of fit test (Shapiro-Wilk W test) was used to test for normality. Statistical analyses were conducted using Minitab 17.1.0 (Minitab Inc., State College, PA, USA).
3. Results
3.1. Seed Production
Mean seed production ranged from 40–996,000 seeds per tree with a mean of ca. 70,000 seeds for all trees sampled from across the three studies (Figure 2 and Figure 3A). Mean seed production per tree was ca. 32,000 seeds (range: 40–686,000) for the 55 Pennsylvania trees included in the current study, ca. 86,000 seeds (range: 240–325,000) for 21 trees reported by Bory and Clair-Maczulajtys [15], and ca. 940,000 seeds (range: 880,000–996,000) for the two large Ailanthus trees reported by Illick and Brouse [17] (Figure 3A, Table S1).
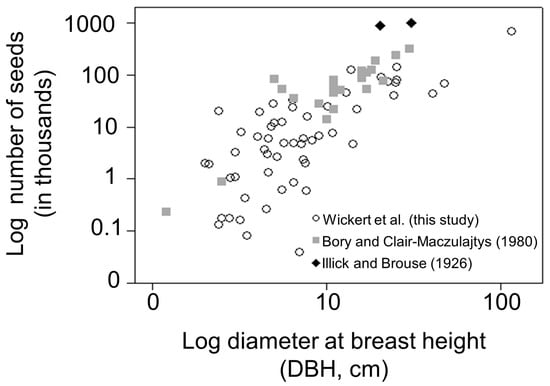
Figure 2.
Ailanthus altissima seed production versus diameter at breast height (DBH, cm) using the combined dataset.
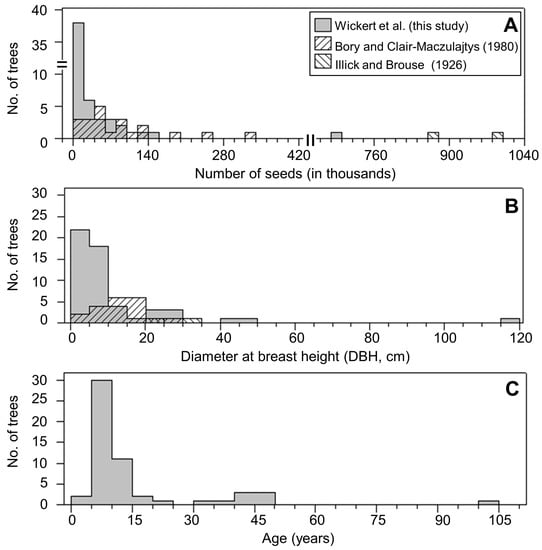
Figure 3.
Distribution of (A) seed counts, (B) diameter at breast height (DBH, cm), and (C) age for seed producing Ailanthus altissima from historic data sets and the current study.
Mean seed production per seed cluster for Ailanthus ranged from 32–1452 seeds, with a mean of 543 seeds per cluster for all 29 trees included in this and two previous studies. Mean seed production per cluster was 567 seeds (range: 91–1452, n = 70) for the seven sampled Pennsylvania trees included in the current study, 388 seeds (range: 32–500) for 21 trees reported by Bory and Clair-Maczulajtys [15], and 959 seeds (range: 703–1364, n = 4 clusters) for one tree reported by Illick and Brouse [17].
The greatest seed-producing trees in the combined dataset were from Pennsylvania. These include two trees reported by Illick and Brouse [17], which produced ca. 880,000 and 996,000 seeds per tree, respectively, and tree PA-62, located on the campus of Penn State University, which produced ca. 685,000 seeds in 2012 (Figure 2 and Figure 3A, Table S1).
Mean diameter of Ailanthus trees sampled in Pennsylvania from 2011–2012 (11.3 cm) was similar to Ailanthus sampled from France in ca. 1980 (13.1 cm). However, significantly larger diameter trees were included in the Pennsylvania study (max DBH = 115 cm) compared to trees sampled across France (max DBH = 30 cm) (Figure 3B, Table S1). Trees reported by Illick and Brouse [17] had diameters of 20.3 and 30.5 cm (Figure 3B). The results of this study support an upward trending relationship between seed production and tree DBH (Figure 2). Indeed, the top 10 highest seed-producing Ailanthus trees from the combined dataset averaged 30.0 cm DBH (range 13–115 cm) and produced ca. 4 million seeds compared to the 10 lowest seed-producing Ailanthus, which accounted for ca. 2300 seeds, with a mean DBH of 3.8 cm (range 2.4–7.6 cm) (Figure 2, Table S1).
The mean age of the seed producing Ailanthus trees sampled across south-central Pennsylvania was 17 years, with a range from 4–104 years (Figure 3C, Table S1). The oldest tree reported here, PA-62, was exceptional in both seed production and age, with an estimated 685,000 seeds produced in 2012 at 99 years of age. Seed collected in both 2008 and 2016 from PA-62 were used in the germination assays.
Regression analysis revealed a significant positive linear relationship between Ailanthus age and DBH, with DBH being a strong single predictor of age for both individual and combined datasets (Figure 4). DBH accounted for 76% and 83% of the observed variability in ages for trees sampled for this study and a previous study by Kasson et al. [1], respectively, and 87% of variability in the combined analysis (Figure 4). Using these results, the ages of the Pennsylvania trees reported by Illick and Brouse [17] were estimated to be 21 years old (20.3 cm DBH) and 29 years old (30.5 cm DBH) (Table S1). Estimated ages for French trees reported by Bory and Clair-Maczulajtys [15] averaged 15 years with a range of 4–29 years, the latter of which was the predicted age for the Ailanthus tree that produced their commonly cited 325,000 seeds (Table S1).
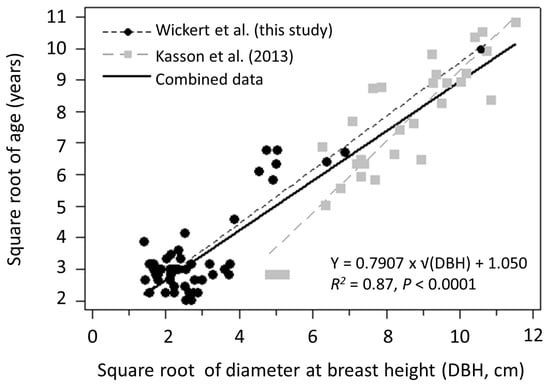
Figure 4.
Relationship between age and diameter at breast height (DBH, cm) of open-grown Ailanthus altissima. Equation for combined data is given above. Equation for Wickert et al. (this study) and Kasson et al. [1], respectively, are as follows: Y = 0.8517 × + 1.027, R2 = 0.76, p < 0.0001; and Y = 1.1266 × − 1.964, R2 = 0.83, p < 0.0001.
Regression analysis of the combined Ailanthus dataset revealed a significant relationship (R2 = 0.66) between Ln number of seeds and Ln DBH (cm), indicating that DBH alone accounted for ca. 66% of the variability in seed production (Figure 5). Although the model was developed to estimate Ailanthus seed production for trees with a specific DBH, the model can also provide estimates of cumulative seed production for the lifetime of an individual Ailanthus. For example, the lifetime seed production of a 50 cm DBH Ailanthus is estimated to be ca. 10 million seeds. Because age-DBH relationships were also explored, it is assumed this estimate of 10 million seeds encompasses a reproductive window of ages 4–43 years (Figure 4). Since the average growth of these trees per year was not determined, cumulative seed production was estimated per year and summed. The lifetime seed production of a 115 cm Ailanthus, such as tree PA-62, is estimated to be around 52 million seeds.
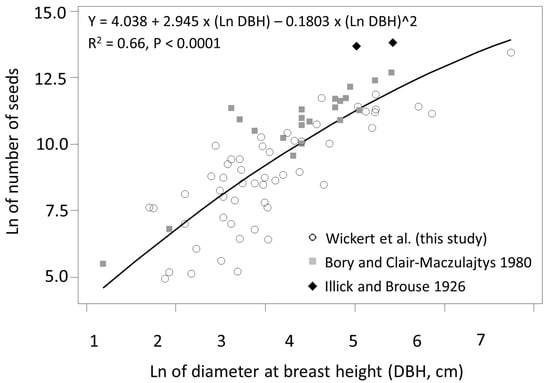
Figure 5.
Relationship between number of seeds produced by Ailanthus altissima and diameter at breast height (DBH, cm). Cumulative seed production for an individual A. altissima over its lifetime, as it grew from 1–49 cm in diameter, was estimated to be ca. 10 million seeds.
3.2. Germination Studies to Assess Seed Viability Based on Seed and Tree Age
Germination rates for seed collected in 2016 ranged from 1.6–78.1% (Table 1). No age-related trends were apparent, as both the oldest and youngest seed-producing trees had similarly high germination rates of 65.6% and 78.1%, respectively. For historic seed sources collected from 2008–2012, germination rates ranged from 0–26.5% (Table 1). More recently collected seed appeared to have higher germination success compared to seed collected in 2009 and 2008, although only three seed sources were compared. Nevertheless, seed collected 7–9 years prior to this study and stored under ideal lab conditions (dry and cool) did germinate.
3.3. Tetrazolium Assay to Evaluate Seed Viability Based on Seed and Tree Age
Results for the seed collected in 2016 using the TZ test supported the results of the germination study, with 22%, 70%, and 62% viability as compared to 16%, 78%, and 66% germination for seed sources WV-02, PA-61, and PA-62, respectively (Table 1). Viability among replicates for individual seed sources were similar, with a range of 17–26% viability for WV-02, 67–72% viability for PA-61, and 55–69% viability for PA-62 (Table 1). These results indicate the TZ test provides a robust evaluation of seed viability and may serve as a suitable substitute for germination studies of A. altissima.
4. Discussion
4.1. Seed Production
Although annual seed production for Ailanthus trees sampled in this study and from the two historic reports varied considerably, seed production was generally positively related to tree diameter, which validates previous studies (Figure 2; [15,23]). Martin and Canham [18] estimated mean seed production of a 30-cm DBH Ailanthus tree in a forested stand at ca. 2 million seeds (range: 564,000–3,793,000). However, these estimates (that resulted from inverse modeling) did not address the issue of which individual tree was contributing seeds and, therefore, cannot be directly compared with mean seed production reported herein. Nevertheless, their data generally support the current study’s results; large Ailanthus trees can produce >1 million seeds per year [18]. This far exceeds the 325,000 seeds per tree per year reported by Bory and Clair-Maczulajtys [15].
In comparison to Ailanthus, Martin and Canham [18] reported that the invasive Acer platanoides can produce ca. 27,000 seeds annually, but with a dramatic decline in seed production over time, suggesting that seed inundation likely contributes less to the invasiveness of this species compared to Ailanthus. Not surprisingly, few other invasive perennial woody species in the United States have annual seed production that surpasses Ailanthus. One exception, velvet tree (Miconia calvescens DC.) in Hawaii, can produce 5–8 million seeds annually and is listed in the top 100 world’s worst invasive alien species (http://www.issg.org/worst100_species.html) [26,27]. The broad-leaved paperbark tree (Melaleuca quinquenervia (Cav.) S.T. Blake), which has invaded the Florida Everglades, also can produce ca. 1 million seeds annually and store up to 20 million seeds produced over successive years in small woody seed pods until conditions favor germination [28]. Saltcedar (Tamarix spp. L.), which has devastated freshwater riparian ecosystems throughout the southwestern United States, can produce up to ca. 500,000 seeds per tree per year [29]. However, unlike paperbark tree seeds, saltcedar seeds are exceptionally short-lived (ca. <5 weeks) and must come in contact with suitable moisture shortly after dispersal to ensure germination [29]. This suggests, as with Acer platanoides, saltcedar deploys other strategies to facilitate invasion [29].
Seed data from Ailanthus sampled in Pennsylvania indicate these populations had >30% more seeds per cluster compared to trees reported from southern France despite having less total seeds/tree, which may be in part explained by strong positive height-seed production relationships in southern France’s Mediterranean climate [23]. The greater seed production among larger trees may reflect greater crown volume to capture sunlight and a correspondingly larger root biomass to absorb soil moisture and nutrients [30]. In addition, previous studies by Kowarik and Säumel [23] showed strong positive height-seed production relationships, which were not explored in this study. Regardless of the factors driving seed production, the results emphasize the significant numbers of seeds and resulting seedlings that a few dozen large trees could produce in a landscape, especially following a major site disturbance downwind/downstream of large seed-producing Ailanthus trees.
Some of the young roadside seed-bearing Ailanthus trees sampled in this study were likely vegetative sprouts arising from pre-existing root systems, probably due to wounding during roadside mowing or trimming. However, the growth form and isolated nature of other young Ailanthus saplings suggests that such trees were likely of seed-origin, supporting the fact that Ailanthus saplings as young as four years of age can produce seed.
In this study, it was determined DBH was a strong predictor of seed production from Ailanthus trees sampled throughout Pennsylvania. Tree DBH-seed/fruit production relationships for other tree species have been reported [30,31,32,33]. Studies on big-leaf mahogany (Swietenia macrophylla King) in the neotropical forests of southern Mexico [30] and Brazil nut (Bertholletia excelsa Humb. and Bonpl.) in northwestern Brazil [33] not only showed that nut production increased with stem diameter, but large trees were more consistent nut producers with limited seed crop failures. Furthermore, Kainer et al. [33] reported that DBH alone explained >50% of the variability in seed production among individuals. In North America, Ares and Brauer [31] and Healy et al. [32] reported nut yield for black walnut (Juglans nigra L.) in south-central United States (R2 = 0.55) and northern red oak (Quercus rubra L.) in Massachusetts (R2 = 0.54), respectively, was significantly and positively related to tree DBH. These estimates may be conservative in that large Ailanthus trees seem to have fewer seed crop failures [18].
4.2. Seed Viability
Although seed viability was not assessed for the 4–5-year-old Ailanthus included in this study, viability of such seed has been previously reported for small diameter sprouts [23]. Nevertheless, seed viability was confirmed from at least one 7-year old tree included in this study with a 78.1% germination rate and 70% seed viability based on a tetrazolium test. Two slightly older trees also included in this study, ages 12 and 16, had much lower germination rates (<20%) despite being part of the same 2016 collections from the same geographic region. These observations support there being other factors (i.e., genetics, microenvironment) that influence seed viability [9,34,35]. The oldest tree assessed in this study (PA-62), a 104-year-old female located on the campus of Penn State University, had a high germination rate of 65.6% and 62% seed viability based on the TZ test. Overall, the tetrazolium test appears to be a reliable proxy for germination based on the reported sampling sizes and replications.
Previous seed germination studies from PA-62 reported similarly high germination rates [1]. Coupled with the ca. 700,000 seeds produced annually by this tree, almost half a million viable offspring are produced each year. This is in sharp contrast to a 114-year-old Ailanthus (HSAa29) in Philadelphia observed with seed production 3–4 orders of magnitude lower than the PA-62 in 2011 [1]. Observations in 2017 of this same tree in Philadelphia indicate that this tree may have stopped producing seed altogether. Although viability of seed from the Philadelphia Ailanthus was not determined, low seed production may suggest fecundity is more variable in advanced age and influenced by other mitigating factors [36]. For example, pollarding (pruning of upper branches to promote dense foliage and branches) was observed in HSAa29, which has been associated with losses in seed production and diameter growth in both strychnine tree (Strychnos nux-vomica L.) and Eucalyptus spp. L’Heritier in India [37,38]. Furthermore, the inherent costs associated with maintenance of sexual reproduction may eventually give way to exclusively asexual reproduction in older trees, which was evidenced by the numerous vegetative sprouts found growing under tree HSAa29.
5. Conclusions
The remarkable success of Ailanthus invasion into the United States is due in part to prolific seed production and prolonged fecundity. In this study, it was established that Ailanthus seed production can begin as early as 4–5 years and can extend >100 years, at which time seed viability can exceed 65%. Total lifetime seed production is dependent upon the individual lifespan of the tree, but females that live 40 years will easily produce 10 million seeds during their lifetime, whereas exceptionally fecund long-lived trees may produce in excess of >50 million seeds. Seed viability varied somewhat among individuals, but seed from both the youngest and oldest individuals assayed in this study (ages 7 and 104 years) had viability >65%. The prolonged reproductive window, as noted in at least two aging Ailanthus trees in Pennsylvania (104 and 114 years as of 2017), ensures the continued reproductive success of this species, since old female trees can produce tens of millions of seeds over their lifetime. Thus, Ailanthus not only has an advantage over competitors by having an early reproductive start at 4–5 years of age, but the consistent, prolific annual seed production ensures that adequate Ailanthus seed will be present during any year that a site is disturbed, and that such sites can be readily colonized by Ailanthus seedlings.
Supplementary Materials
The following are available online at www.mdpi.com/1999-4907/8/7/226/s1, Table S1: Tree-level data for gynoecious Ailanthus altissima included in the study.
Acknowledgments
Special thanks to Annie Ye, Toby Grapner, and Jim Savage who assisted with seed collection, sorting, and counting. Thanks to Christa Quint and Ida Holaskova for advice on statistics. Additional thanks to Pennsylvania Department of Transportation (PennDOT), Pennsylvania Game Commission, and Army Corps of Engineers, who provided sampling locations. Funding for K.L.W. was provided by USDA APHIS (Agreement No. 16-8254-2074-CA) and USDA Forest Service, Northeastern Area, State and Private Forestry, Forest Health Technology Enterprise Team (FHTET), Morgantown, West Virginia (Cooperative Agreement 16-CA-11420004-296). M.T.K. was supported by funds appropriated under Hatch Project NE-1333.
Author Contributions
K.L.W., D.D.D. and M.T.K. designed the studies. K.L.W., E.S.O. and M.T.K. conducted the research. K.L.W. and M.T.K. wrote the manuscript.
Conflicts of Interest
The authors report no conflicts of interest.
References
- Kasson, M.T.; Davis, M.D.; Davis, D.D. The invasive Ailanthus altissima in Pennsylvania: A case study elucidating species introduction, migration, invasion, and growth patterns in the northeastern US. Northeast. Nat. 2013, 20, 1–60. [Google Scholar]
- Gray, A. Manual of the Botany of the Northern United States; Ivison, Blakeman, Taylor, and Co.: New York, NY, USA, 1868. [Google Scholar]
- Barry, P.; Smith, J.J.E. Arboriculture Notes-No. II: Bartram’s Garden, Philadelphia. Hortic. J. Rural Art Rural Taste 1885, 5, 371–375. [Google Scholar]
- Aldrich, P.R.; Briguglio, J.S.; Kapadia, S.N.; Morker, M.U.; Rawal, A.; Kalra, P.; Greer, G.K. Genetic structure of the invasive tree Ailanthus altissima in Eastern United States cities. J. Bot. 2010, 2010, 795735. [Google Scholar]
- McAvoy, T.J.; Snyder, A.L.; Johnson, N.; Salom, S.M.; Kok, L.T. Road survey of the invasive tree-of-heaven (Ailanthus altissima) in Virginia. Invasive Plant Sci. Manag. 2012, 5, 506–512. [Google Scholar] [CrossRef]
- Rebbeck, J.; Kloss, A.; Bowden, M.; Coon, C.; Hutchinson, T.F.; Iverson, L.; Guess, G. Aerial detection of seed-bearing female Ailanthus altissima: A cost-effective method to map an invasive tree in forested landscapes. For. Sci. 2015, 61, 1068–1078. [Google Scholar] [CrossRef]
- Gaertner, M.; Den Breeyen, A.; Hui, C.; Richardson, D.M. Impacts of alien plant invasions on species richness in Mediterranean-type ecosystems: A meta-analysis. Prog. Phys. Geogr. 2009, 33, 319–338. [Google Scholar] [CrossRef]
- Harris, P.T.; Cannon, G.H.; Smith, N.E.; Muth, N.Z. Assessment of plant community restoration following tree-of-heaven (Ailanthus altissima) control by Verticillium albo-atrum. Biol. Invasions 2013, 15, 1–7. [Google Scholar] [CrossRef]
- Kota, N.L.; Landenberger, R.E.; McGraw, J.B. Germination and early growth of Ailanthus and tulip poplar in three levels of forest disturbance. Biol. Invasions 2007, 9, 197–211. [Google Scholar] [CrossRef]
- Motard, E.; Muratet, A.; Clair-Maczulajtys, D.; Machon, N. Does the invasive species Ailanthus altissima threaten floristic diversity of temperate peri-urban forests? C. R. Biol. 2011, 334, 872–879. [Google Scholar] [CrossRef] [PubMed]
- Planchuelo, G.; Catalán, P.; Delgado, J.A. Gone with the wind and the stream: Dispersal in the invasive species Ailanthus altissima. Acta Oecol. 2016, 73, 31–37. [Google Scholar] [CrossRef]
- Planchuelo, G.; Catalán, P.; Delgado, J.A.; Murciano, A. Estimating wind dispersal potential in Ailanthus altissima: The need to consider the three-dimensional structure of samaras. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2017, 151, 316–322. [Google Scholar] [CrossRef]
- Von der Lippe, M.; Bullock, J.M.; Kowarik, I.; Knopp, T.; Wichmann, M. Human-mediated dispersal of seeds by the airflow of vehicles. PLoS ONE 2013, 8, e52733. [Google Scholar] [CrossRef]
- Mortensen, D.A.; Rauschert, E.S.; Nord, A.N.; Jones, B.P. Forest roads facilitate the spread of invasive plants. Invasive Plant Sci. Manag. 2009, 2, 191–199. [Google Scholar] [CrossRef]
- Bory, G.; Clair-Maczulajtys, D. Production, dissemination and polymorphism of seeds in Ailanthus altissima. Reuve Generale de Botanique 1980, 88, 297–311. [Google Scholar]
- Cabra-Rivas, I.; Castro-Díez, P. Comparing the sexual reproductive success of two exotic trees invading Spanish riparian forests vs. a native reference. PLoS ONE 2016, 11, e0160831. [Google Scholar] [CrossRef] [PubMed]
- Illick, J.S.; Brouse, E.F. The Ailanthus tree in Pennsylvania; Pennsylvania Department of Forests and Waters: Harrisburg, PA, USA, 1926. [Google Scholar]
- Martin, P.H.; Canham, C.D. Dispersal and recruitment limitation in native versus exotic tree species: Life-history strategies and Janzen-Connell effects. Oikos 2010, 119, 807–824. [Google Scholar] [CrossRef]
- Kaproth, M.A.; McGraw, J.B. Seed viability and dispersal of the wind-dispersed invasive Ailanthus altissima in aqueous environments. For. Sci. 2008, 54, 490–496. [Google Scholar]
- Kowarik, I.; Säumel, I. Water dispersal as an additional pathway to invasions by the primarily wind-dispersed tree Ailanthus altissima. Plant Ecol. 2008, 198, 241–252. [Google Scholar] [CrossRef]
- Feret, P.P. Notes: Early flowering in Ailanthus. For. Sci. 1973, 19, 237–239. [Google Scholar]
- Hegi, G. Illustrierte Flora von Mitteleuropa; J.F. Lehmann: Munich, Germany, 1906; Volume 4. [Google Scholar]
- Kowarik, I.; Säumel, I. Biological flora of Central Europe: Ailanthus altissima (Mill.) Swingle. Perspect. Plant Ecol. 2007, 8, 207–237. [Google Scholar] [CrossRef]
- Kasson, M.T.; O’Neal, E.S.; Davis, D.D. Expanded host range testing for Verticillium nonalfalfae: Potential biocontrol agent against the invasive Ailanthus altissima. Plant Dis. 2015, 99, 823–835. [Google Scholar] [CrossRef]
- Cottrell, H.J. Tetrazolium salt as a seed germination indicator. Nature 1947, 159, 748. [Google Scholar] [CrossRef] [PubMed]
- Mason, R.A.; Cooke, J.; Moles, A.T.; Leishman, M.R. Reproductive output of invasive versus native plants. Glob. Ecol. Biogeogr. 2008, 17, 633–640. [Google Scholar] [CrossRef]
- Meyer, J.Y. Observations on the reproductive biology of Miconia calvescens DC. (Melastomataceae), an alien invasive tree on the island of Tahiti (South Pacific Ocean). Biotropica 1998, 30, 609–624. [Google Scholar] [CrossRef]
- Swearingen, J.M. Plant Conservation Alliance, Alien Plant Working Group: Melaleuca quinquenervia (Cav.) Blake. Available online: https://www.nps.gov/plants/alien/fact/mequ1.htm (accessed on 15 May 2017).
- Di Tomaso, J.M. Impact, biology, and ecology of saltcedar (Tamarix spp.) in the southwestern United States. Weed Technol. 1998, 12, 326–336. [Google Scholar]
- Snook, L.K.; Cámara-Cabrales, L.; Kelty, M.J. Six years of fruit production by mahogany trees (Swietenia macrophylla King): Patterns of variation and implications for sustainability. For. Ecol. Manag. 2005, 206, 221–235. [Google Scholar] [CrossRef]
- Ares, A.; Brauer, D. Growth and nut production of black walnut in relation to site, tree type and stand conditions in south-central United States. Agrofor. Syst. 2004, 63, 83–90. [Google Scholar] [CrossRef]
- Healy, W.M.; Lewis, A.M.; Boose, E.F. Variation of red oak acorn production. For. Ecol. Manag. 1999, 116, 1–11. [Google Scholar] [CrossRef]
- Kainer, K.A.; Wadt, L.H.; Staudhammer, C.L. Explaining variation in Brazil nut fruit production. For. Ecol. Manag. 2007, 250, 244–255. [Google Scholar] [CrossRef]
- Constán-Nava, S.; Bonet, A. Genetic variability modulates the effect of habitat type and environmental conditions on early invasion success of Ailanthus altissima in Mediterranean ecosystems. Biol. Invasions 2012, 14, 2379–2392. [Google Scholar] [CrossRef]
- Delgado, J.A.; Jimenez, M.D.; Gomez, A. Samara size versus dispersal and seedling establishment in Ailanthus altissima (Miller) Swingle. J. Environ. Biol. 2009, 30, 183–186. [Google Scholar] [PubMed]
- Moore, J.E.; Lacey, E.P. A comparison of germination and early growth of four early successional tree species of the southeastern United States in different soil and water regimes. Am. Midl. Nat. 2009, 162, 388–394. [Google Scholar] [CrossRef]
- Nicholson, J.W. Results of coppicing, pollarding and pruning experiments to stimulate Strychnos nux-vomica fruit production. Indian For. 1937, 63, 588–597. [Google Scholar]
- Varghese, M.; Ravi, N.; Kamalakannan, R.; Harwood, C.E. Effect of silvicultural treatments on growth, fertility and capsule traits in seedling seed orchards of Eucalyptus camaldulensis and E. tereticornis. New For. 2009, 37, 99–107. [Google Scholar] [CrossRef]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).