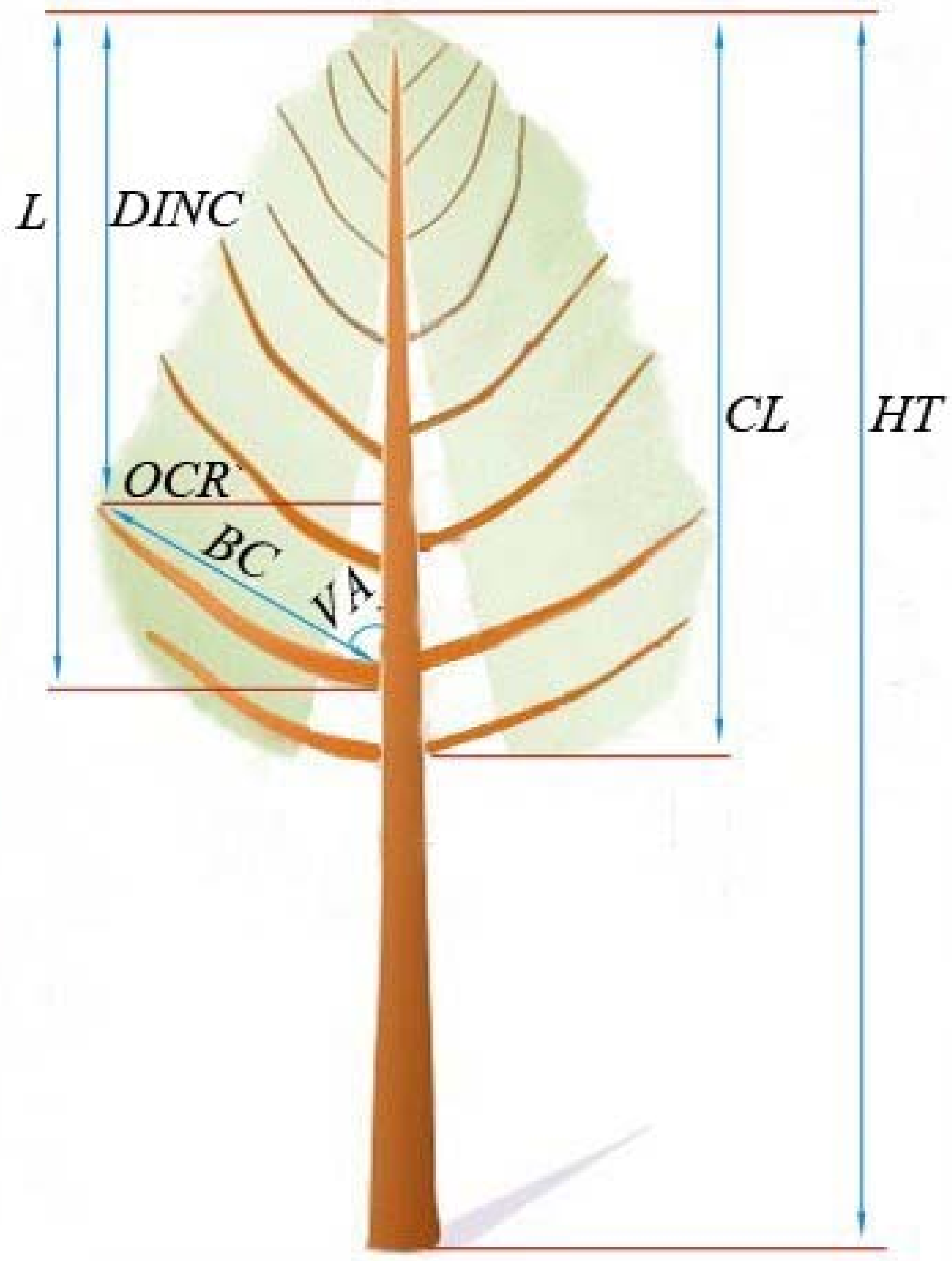
1. Introduction
Crown size exhibits an intimate relationship to species diversity and ecosystem stability; it is directly related to the growth and yield of individual trees within a specified stand and provides a reference for decision-making in forest management [
1]. The light conditions at different latitudes is the most decisive factor in determining the crown profile in comparison to other factors, such as the competition index, water and nutrient variations [
2], or genetics and the physical environment [
3]. However, the potential influence of inter-tree competition should not be ignored, as it can factor into determining the asymmetric shape of a tree’s crown [
4].
Various variables have been widely used to reflect the gross crown size, such as crown length or crown ratio [
5], crown width [
6], crown surface area, crown volume [
7], and leaf area and biomass distribution [
8]. Crown length and crown width focus on the crown features in vertical and horizontal directions, respectively. In comparison, crown volume, crown surface area, and leaf features are more effective variables in describing the spatial characteristics of the crown, and can reflect the competition level of an individual tree in forest stands; however, these characteristics are difficult to measure directly. Crown profile is another crown attribute that can reflect crown size and which can be modeled through easily-measured tree variables. Furthermore, the largest crown width, crown volume, and surface area can be calculated based on a crown profile model. A crown profile model can also be invaluable in evaluating inter-tree competition [
9], tree vigor, and biodiversity evaluation [
10]. Because tree and stand variables are continuously changing during the growth period, developing a crown profile model represents an efficient alternative to direct measurements, as it is costly and time consuming to measure crown profiles for all trees [
3].
Generally, crown profiles are modeled using two approaches. The first approach divides the entire crown into two sections: the “light crown” and the “shade crown”, with models developed separately for each of the two parts [
11]. The second method uses a single equation without a break point to model the entire crown [
9,
12,
13]. The disadvantage of the first approach is that the continuity of the entire crown is not considered; thus, the second approach has become increasingly popular. Today, the polynomial equation and variable exponent equation are frequently used in directly or indirectly modeling crown profiles [
14,
15,
16]. The polynomial equation was usually defective in the prediction accuracy of the maximum radius, and the variable exponent equation would express various shapes of the outer crown by changing the exponent of the function [
9].
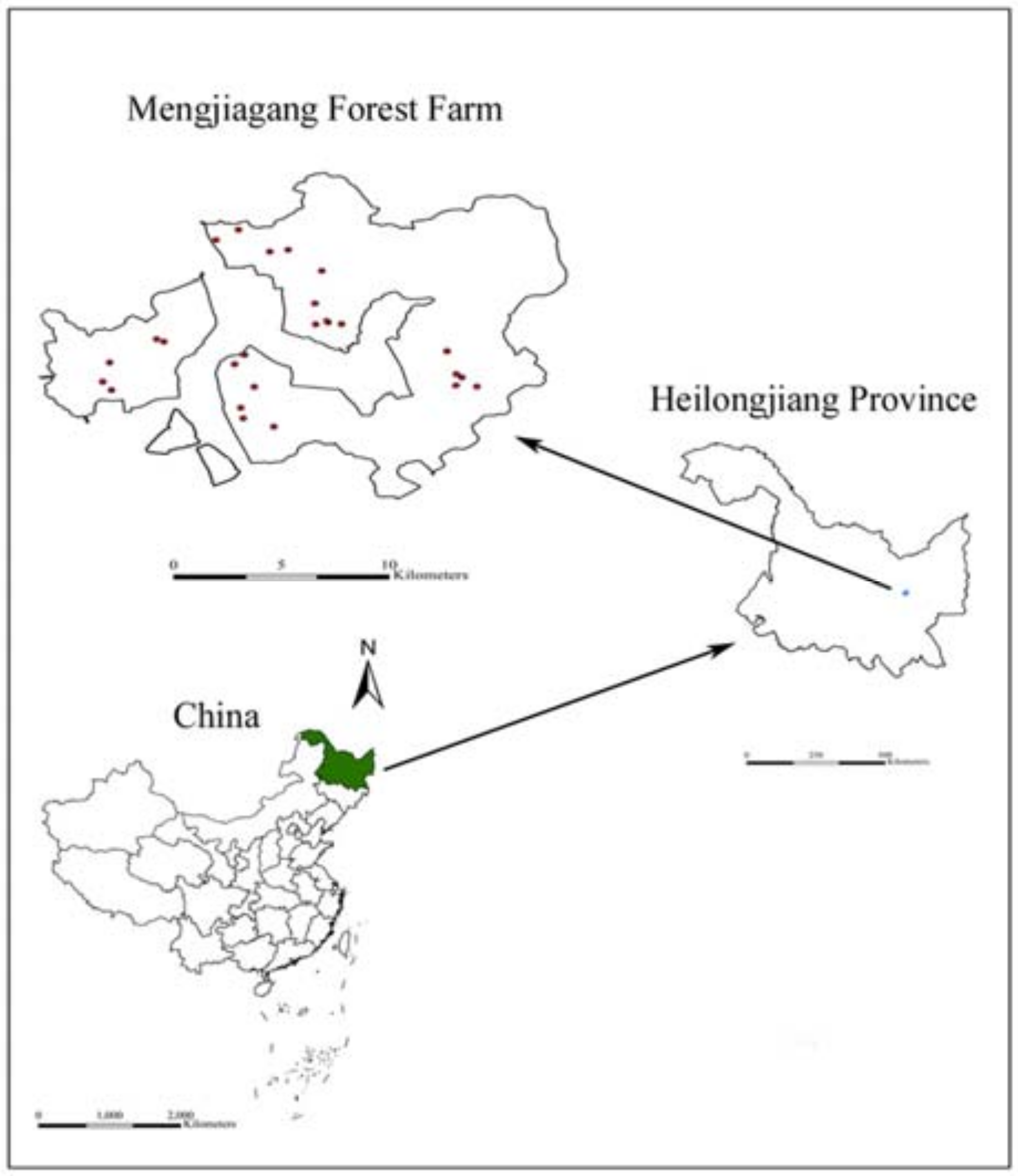
Heilongjiang Province is located in Northeast China, and has rich forest resources. The total area of forest in the Heilongjiang Province of Northeast China is approximately 19.63 million hectares, with a forest coverage of about 43.16%.
Larix olgensis Henry is one of the most important coniferous and afforestation tree species in Heilongjiang Province. The area planted with
Larix olgensis amounts to about 45% and 36% of the total forested planted tree area and standing volume, respectively [
17]. To date, there are limited studies on the outer crown profile of
Larix olgensis plantations in China. To the best of our knowledge, most current crown profile models emphasize the hypothesis that the crown shape is symmetrical in all directions. In contrast, the shape of the crown in our study may be asymmetrical because of inter-tree competition. Thus, the goals of the present study were to: (1) select the best equation to develop the entire outer crown profile model for individual trees in planted stands of
Larix olgensis in Heilongjiang Province, Northeast China; (2) evaluate the differences in crown profiles between tree statuses (dominant, intermediate, and suppressed), assuming that the crown is symmetrical, and then between the four crown directions (i.e., north, east, south, and west) with the assumption of no differences existing between tree statuses, and then by considering the tree status and crown directions together.
4. Discussion
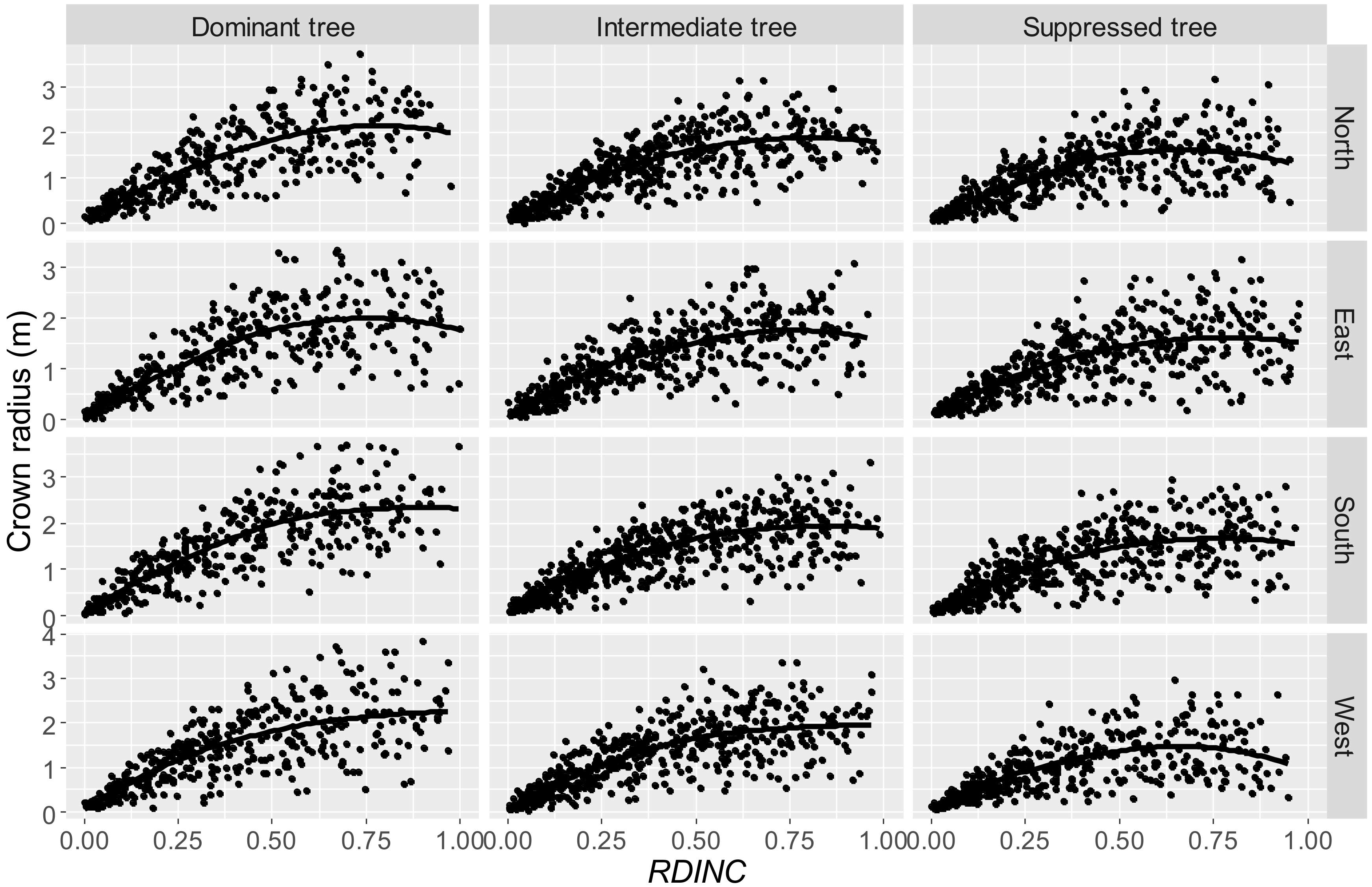
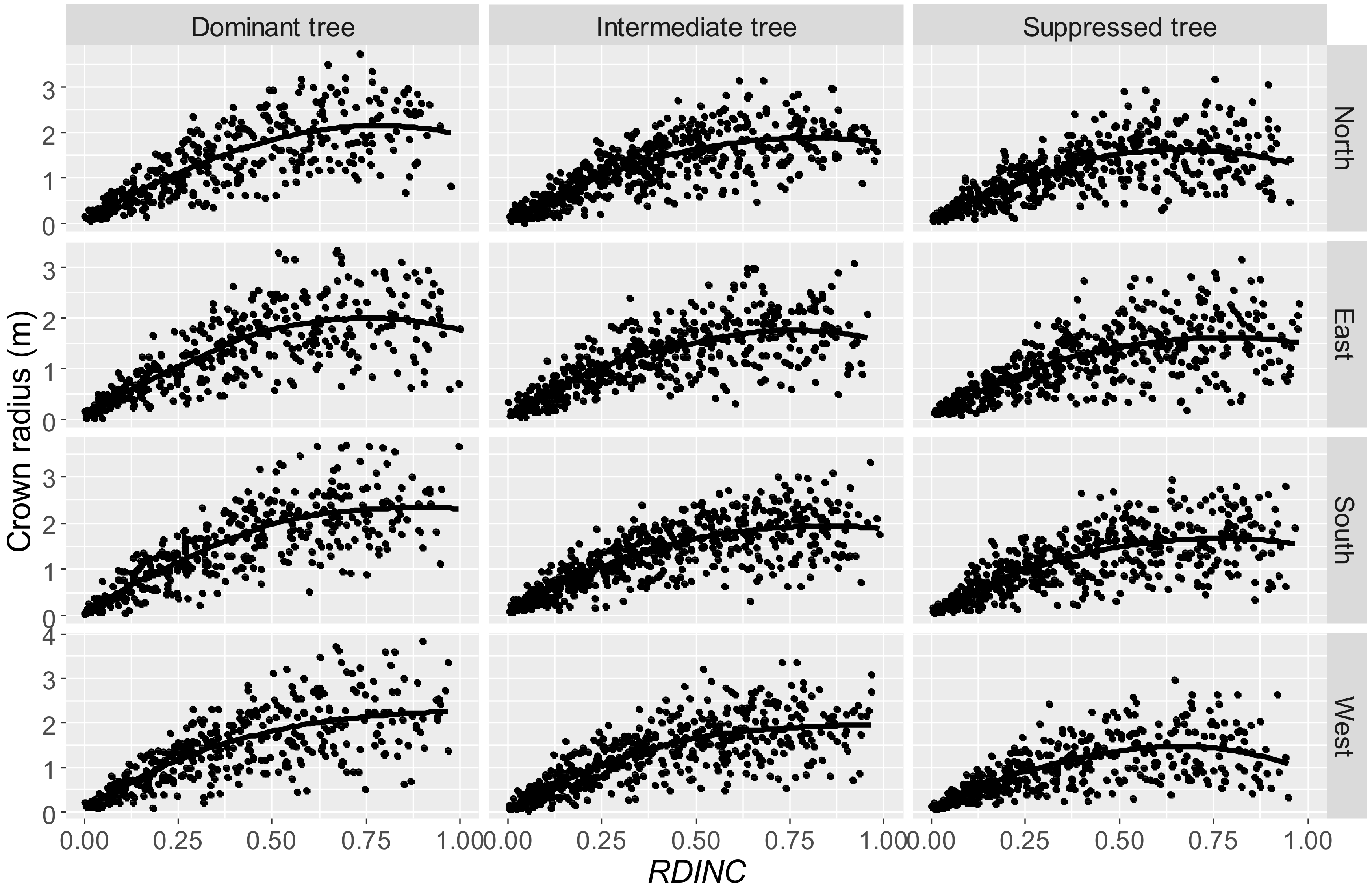
The data used in our study varied a great deal, and even the crown radii in different directions show large variation (
Figure 2). Tree variables explain the heterogeneity of the crown radius from different directions of dominant, intermediate, and suppressed trees. Thus, the final model (Equation 10) involved
DBH,
CR, and
HD.
DBH is always the most commonly used variable in the development of crown profile models [
9]. Crown width expressed the space an individual tree occupied and strongly influenced the crown profile, so
DBH could also be used as a single independent variable in the crown width prediction models [
24]. Based on the analysis, crown radius increased significantly with the increase of
DBH. The crown shape of
Larix olgensis in our study was also influenced by
CR. The crown ratio is an indicator of stand density [
25], competition, and survival in the forest stand [
26], and has been used as the predictor in the crown profile model [
23].
CR strongly influenced the crown profile within the upper crown, and the crown radius increased with an increase in
CR.
The density expressed the crowding of the individual trees in the stands so that the crown shape would be affected, which conformed to the studies of previous research [
7]. Since
HD is the response to comprehensive management activities and reflects the slenderness of a tree as affected by forest management, it is usually included in the crown profile model [
9,
11,
15]. In our study, it was also a significant variable that influenced the crown profile. With respect to the simulation of the effect of
HD on crown shape in our study,
HD affected the crown shape of individual trees, even though it was not as pronounced when compared to other variables. The crown radius within the crown above the largest crown radius increased with the increase of
HD, and the crown radius decreased with the increase of
HD for the lower part, in which the largest crown radius occurred. Our study confirmed the findings of Sharma et al. [
1], where crown width decreased with the increase of the tree slenderness coefficient (
HD).
The dummy variable approach was used to model the crown profiles for different tree statuses (Step 1), different crown directions (Step 2), and tree status and crown directions in combination (Step 3). In our study, the results demonstrated that the crown profiles were affected by tree status (
Table 5), with tree status representing the reason why large error was produced using the traditional approach to predict the crown profiles for three tree statuses (
Table 6). Peltola et al. [
27] found that tree status affected the radial growth of individual trees, and that radial growth of trees was closely related to the crown. In addition, the light in the forest stand intercepted by larger trees was larger than that of the smaller trees, so the smaller trees had no chance to compete for light [
28]. The dominant trees occupied large enough spaces to utilize the light resources in the forest stand compared to intermediate and suppressed trees, which may be why the crown profiles of different tree statuses were different. Getzin and Wiegand [
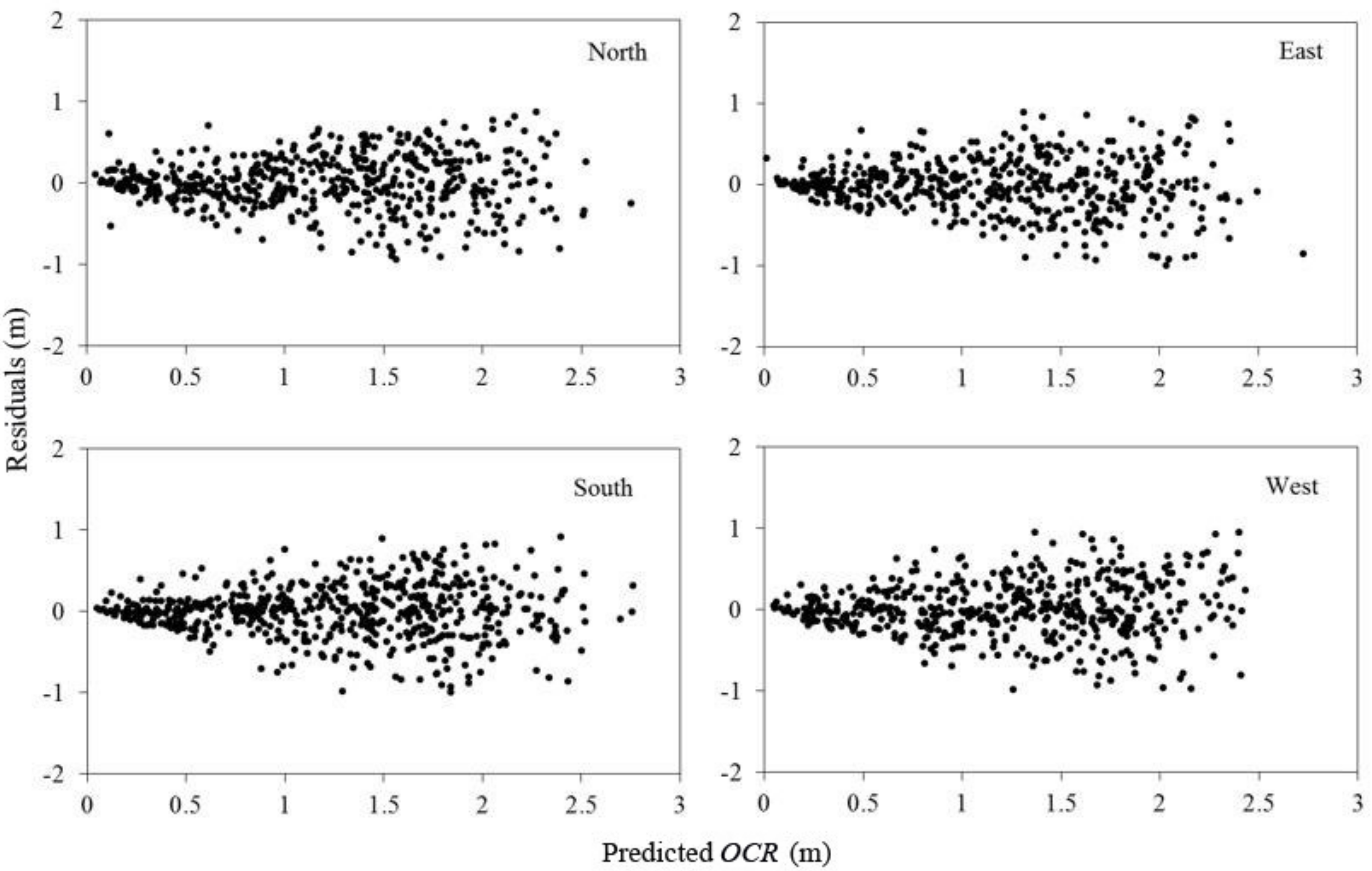
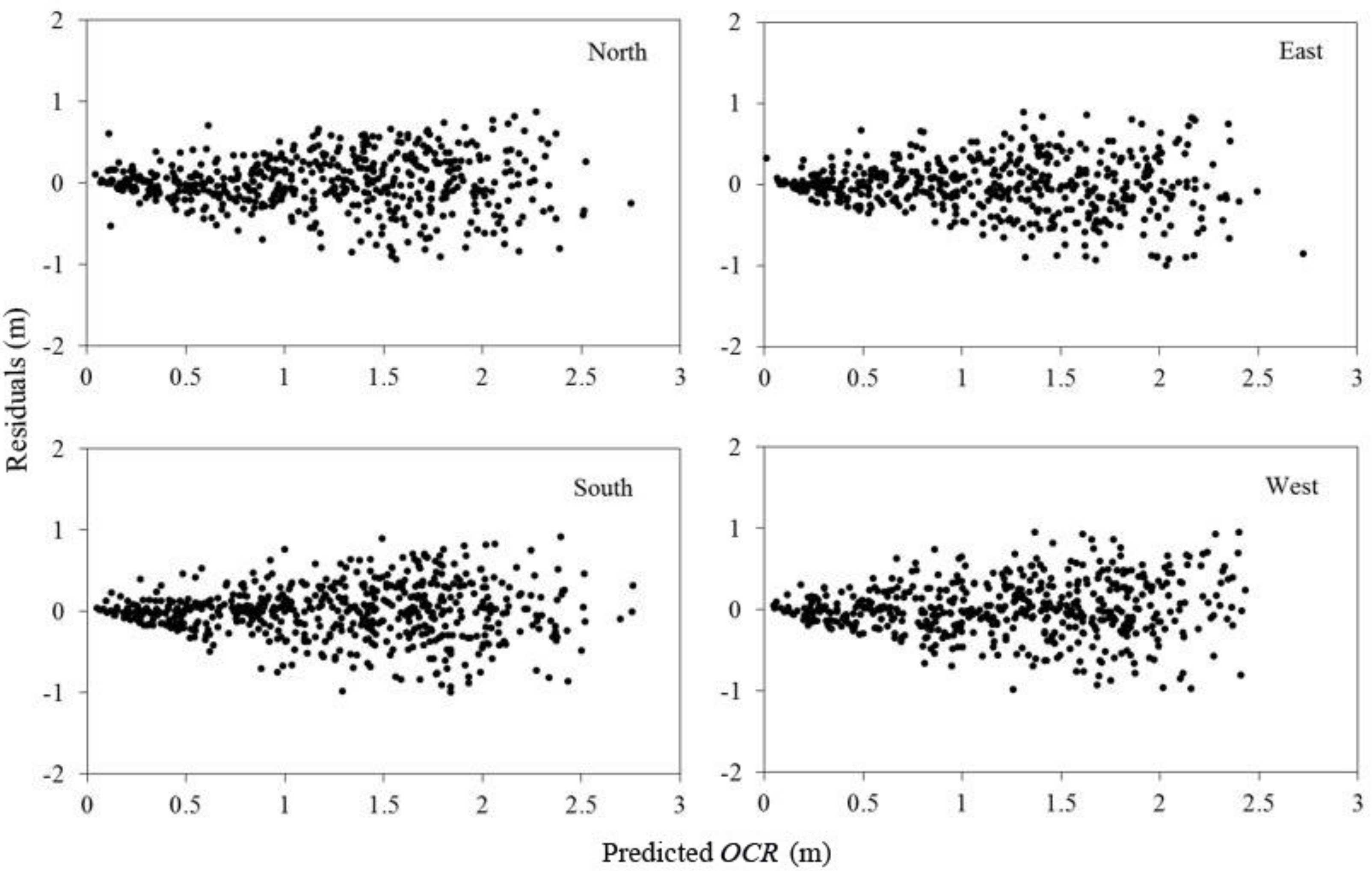
29] described the asymmetric characteristics of tree crowns; however, the present crown profile models have seldom considered this aspect. In our study, the effect of crown directions alone on the crown profiles was further analyzed and the range of mean error was within ±0.0190 m, while the range of mean error for the traditional approach was ±0.0385 m (
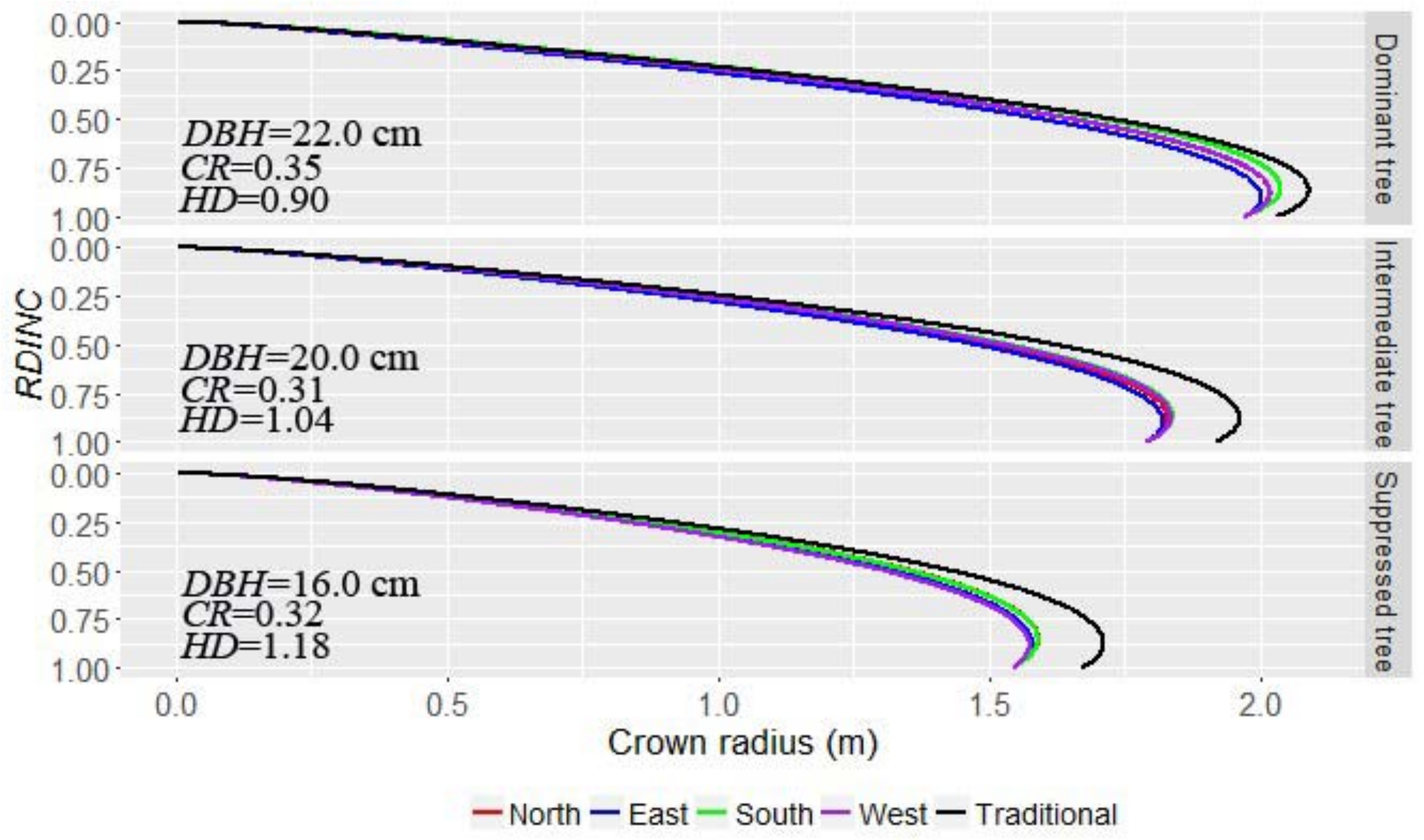
Table 7). We also analyzed the tree status and crown directions at the same time (Step 3), and results indicate that the north-, east-, south-, and west-orientated crown profiles for dominant, intermediate, and suppressed trees were different, which further demonstrated the asymmetries in the crown profiles (
Figure 5 and
Table 8). The crown radius in the south direction for dominant, intermediate, and suppressed trees was still the largest. This suggests that light was the main factor that affected crown shape, as the south side of the crown intercepts a relatively larger amount of solar radiation in Northeast China. Our results confirmed the study of Marshall et al. [
30], who revealed that individual trees in forest stands express intra-specific genetic variation both within and among populations, and that growth rates of individual trees were affected by the photosynthetic traits in the uniform environmental conditions. Thus, the light source was the main factor leading to the largest crown radius in the south direction, while the differences produced between other directions may derive from the unbalanced competition status from neighboring trees.
The physiological patterns of intermediate trees were distinct from either saplings or larger trees [
31]. Thomas and Ickes [
32] documented that leaf size peaked at intermediate trees in several tropical species. Despite the difference in climate for the trees growing between our study and that of the study by Thomas and Ickes [
32], it confirms that the growth state of intermediate trees is different from that of dominant trees. The crowns of intermediate trees have the potential to grow into the upper layer, even when faced with strong competition from other trees. For suppressed trees, the competition level was much stronger compared to that of dominant and intermediate trees. In addition, many biological processes are size-dependent, and the rate of the biological processes change during the growth of individual trees [
33]. Thus, as the main component of the individual tree, the growth and development of the crown may be affected and leads to the differences of crown shapes between tree statuses. However, the entire crown surface of individual trees modeled in our study was not continuous, and this is an issue we will seek to address in future research as we strive to lay the foundation for the development of competition indices.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}