
Responses of Clonal Eucalypt Plantations to N, P and K Fertilizer Application in Different Edaphoclimatic Conditions

 , ,
, ,
Abstract
:1. Introduction
2. Material and Methods
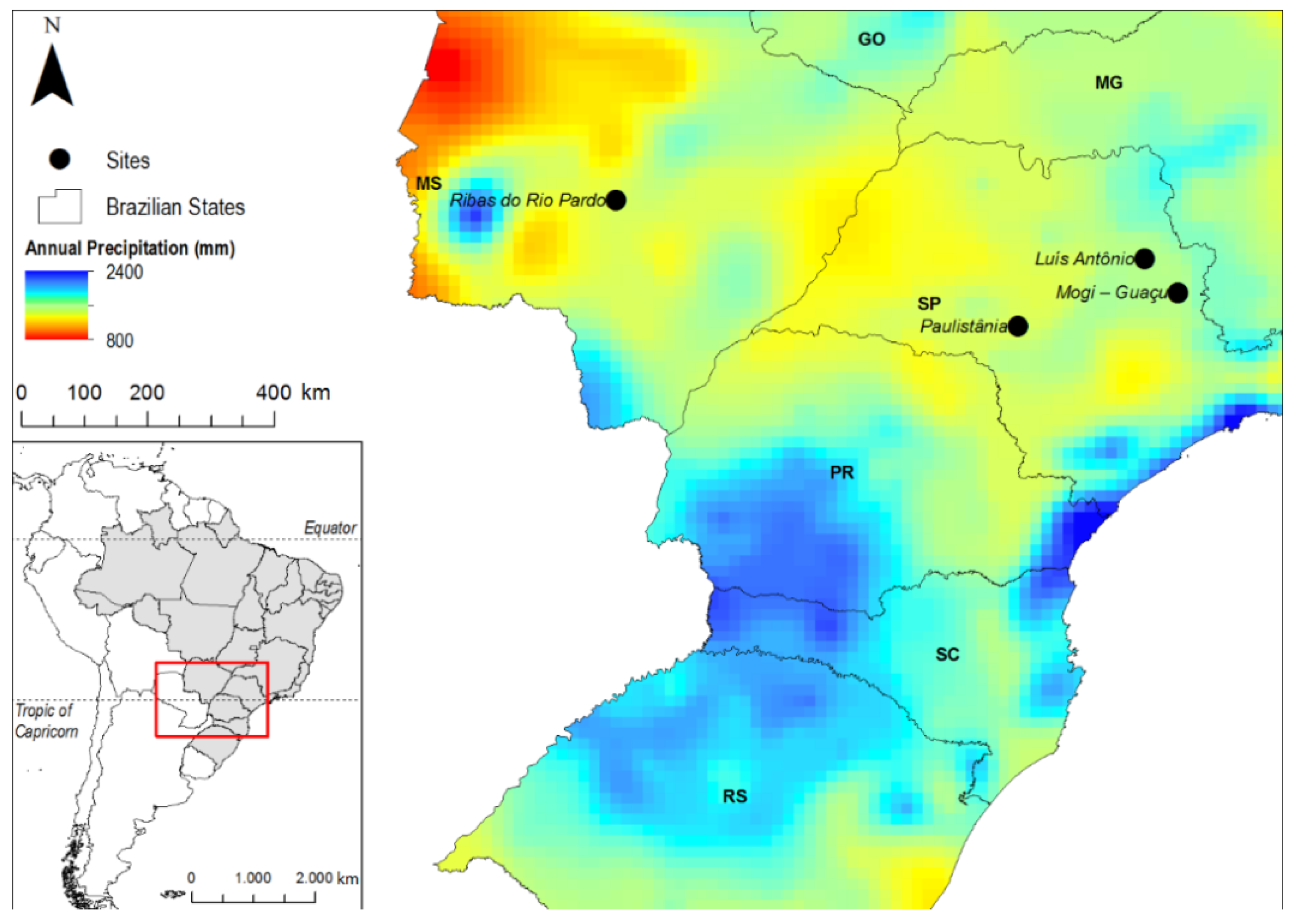
2.1. Site Description

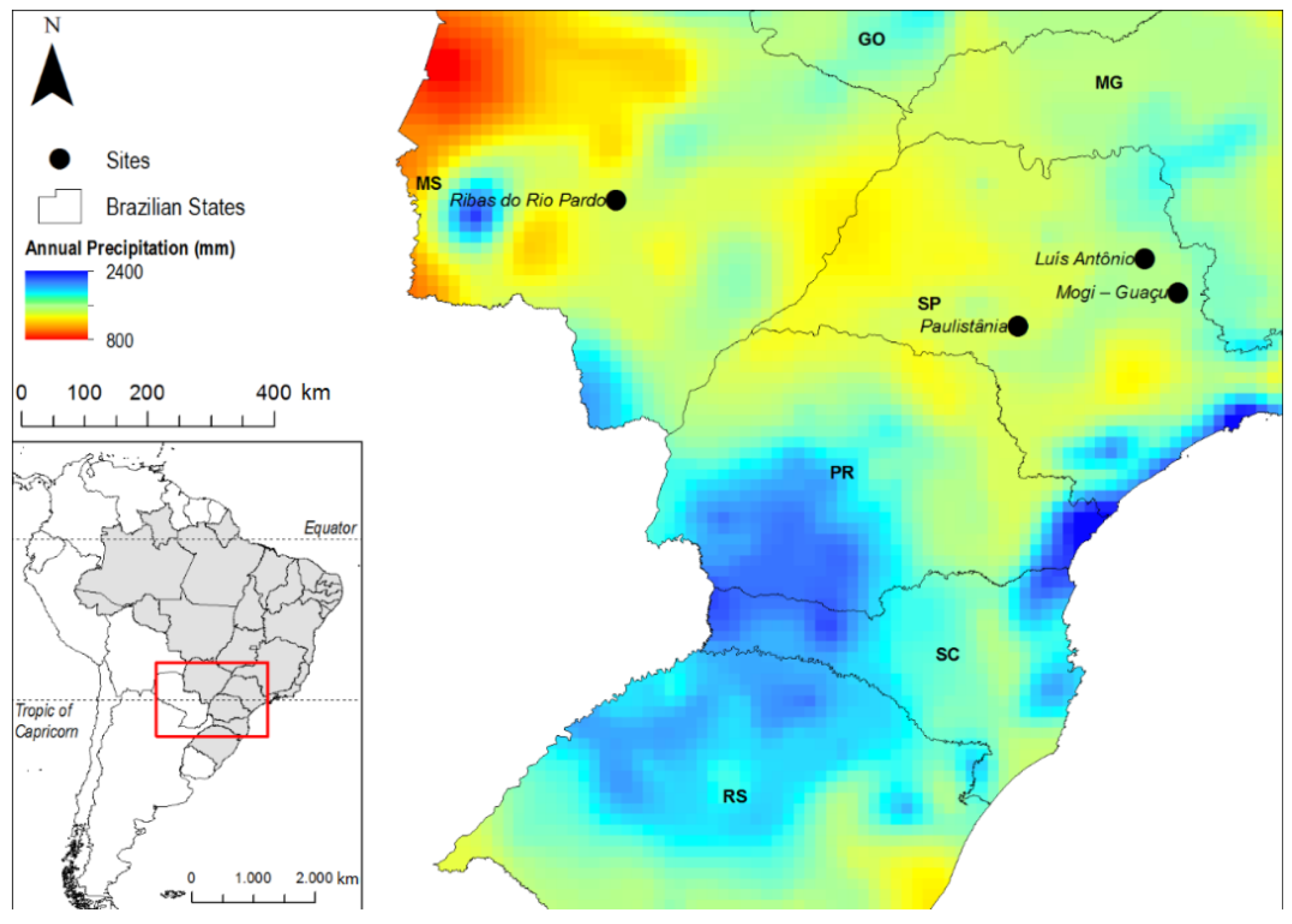{kind=link}
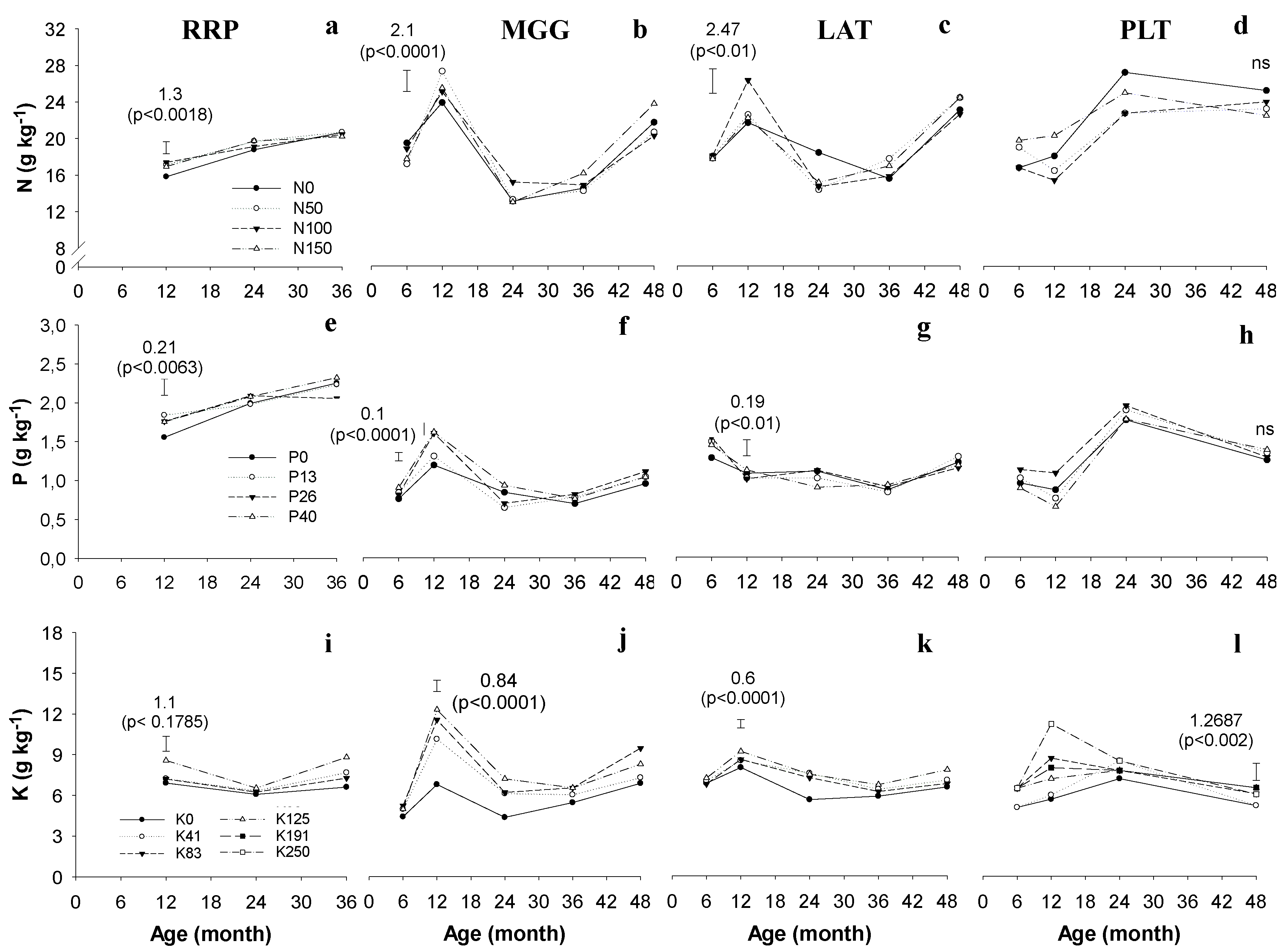{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Lat | Long | Alt | T | PP | WS 1,2 | WD 1,3 | WDM |
|---|---|---|---|---|---|---|---|---|
| S 4 | W 5 | m | °C | ____________mm____________ | ||||
| RRP | 20°45′ | 54°62′ | 530 | 24.5 | 1472 | 310 | 173 | 6 |
| PLT | 22°33′ | 49°19′ | 610 | 22.5 | 1387 | 355 | 86 | 6 |
| MGG | 22°04′ | 47°02′ | 591 | 22.0 | 1446 | 488 | 99 | 5 |
| LAT | 21°35′ | 47°31′ | 610 | 21.5 | 1213 | 311 | 118 | 6 |
| Site | Soil Type 1 | Clay 2 | Sand 2 | OM 3 | pH 4 | P-Resin 5 | Cation Exchange | |||
|---|---|---|---|---|---|---|---|---|---|---|
| Ca 5 | Mg 5 | K 5 | Al 6 | |||||||
| % | g·kg−1 | mg·kg−1 | ________mmolc·kg−1________ | |||||||
| RRP | Quartzipsamment | 7.3 | 90.8 | 12.9 | 4.0 | 1.4 | 1.4 | 1.0 | 0.3 | 6.8 |
| PLT | Rhodic Hapludox | 12.3 | 86.0 | 9.8 | 3.6 | 3.0 | 3.0 | 1.5 | 0.4 | 9.5 |
| LAT | Quartzipsamment | 8.9 | 90.7 | 19.4 | 4.1 | 8.7 | 16.3 | 5.8 | 0.4 | 4.1 |
| MGG | Typic Hapludox | 16.4 | 50.9 | 16.1 | 4.1 | 4.0 | 20.5 | 3.5 | 0.3 | 0.9 |
2.2. Experimental Design
2.3. Leaf Nutrient Content
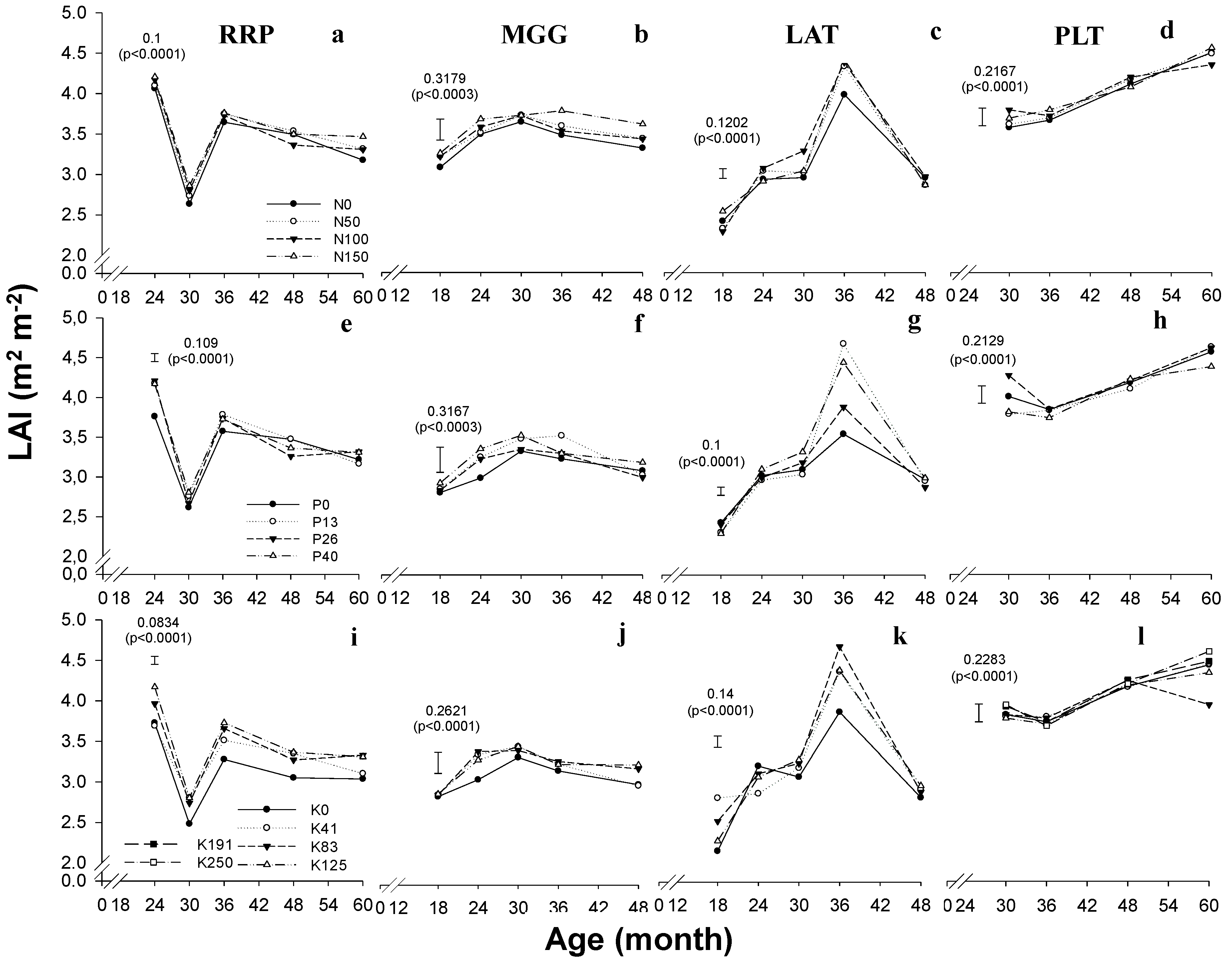
2.4. Leaf Area Index
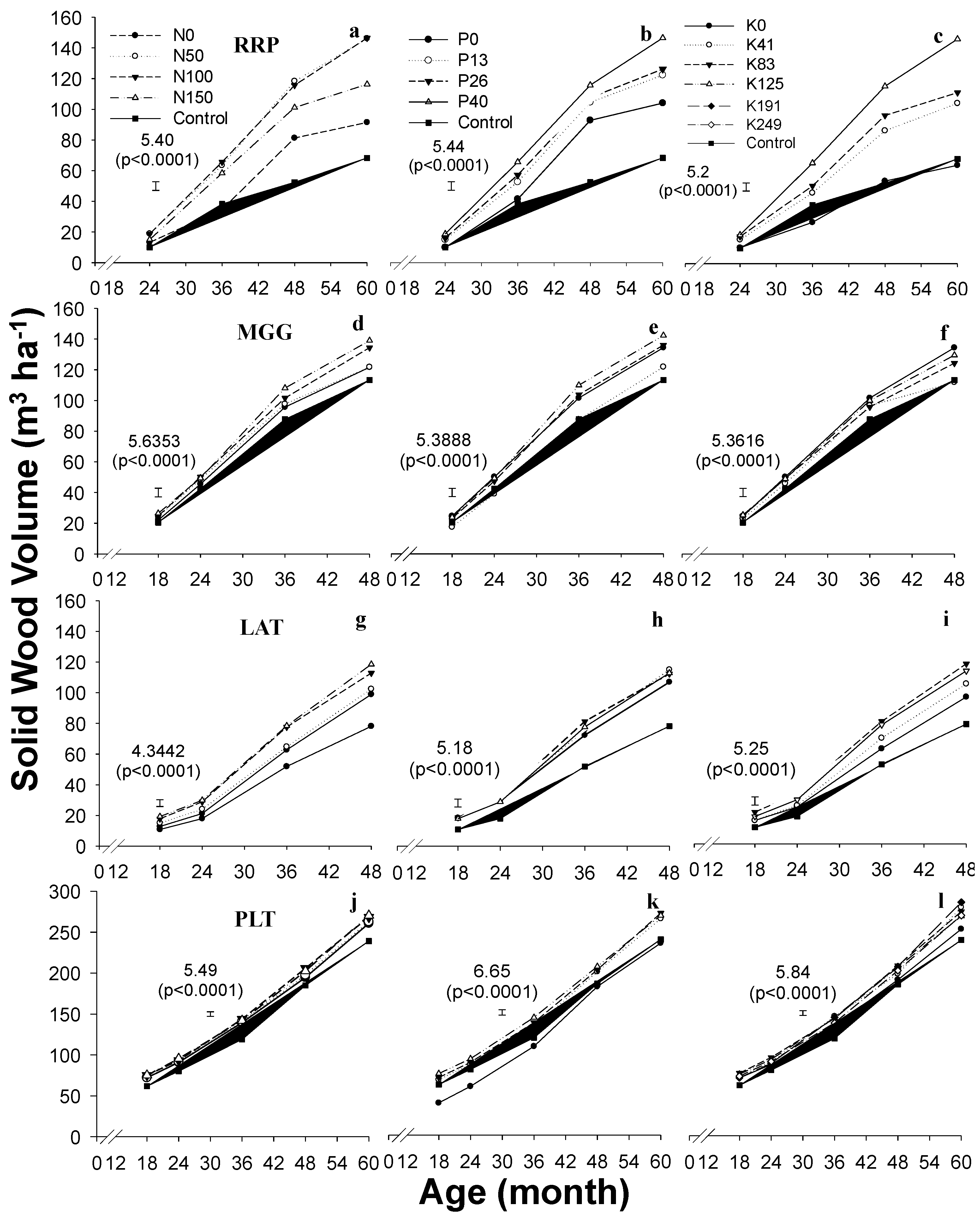
2.5. Tree Growth
2.6. Data Analysis
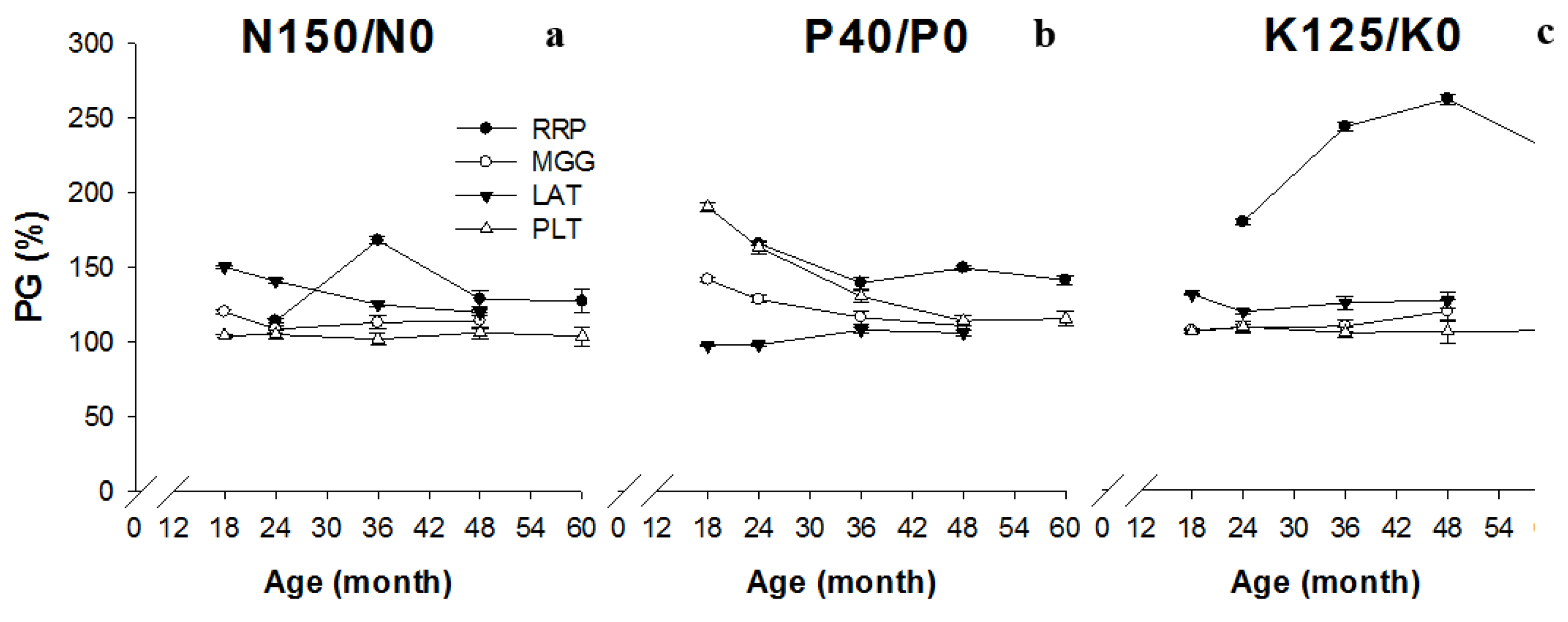
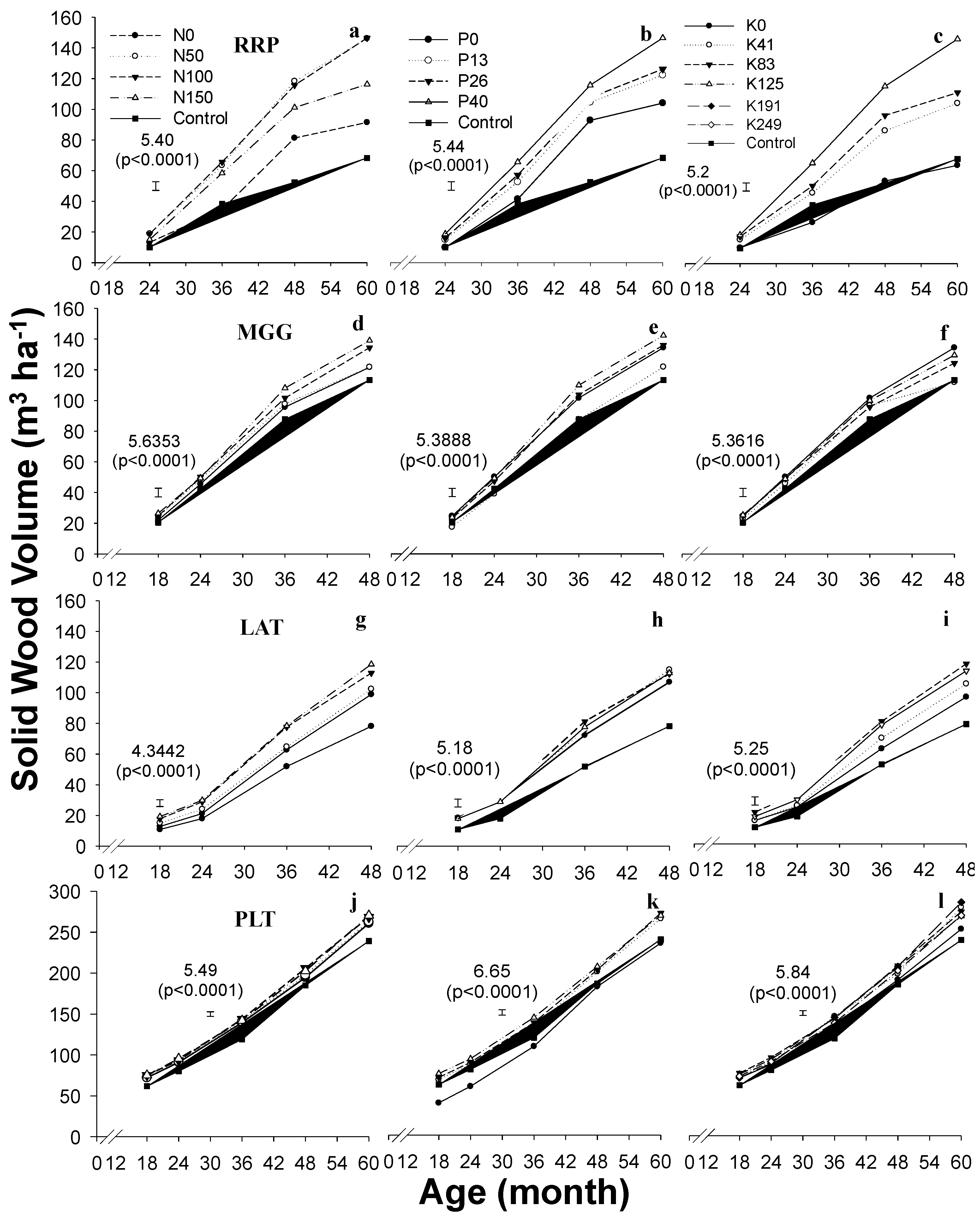
3. Results
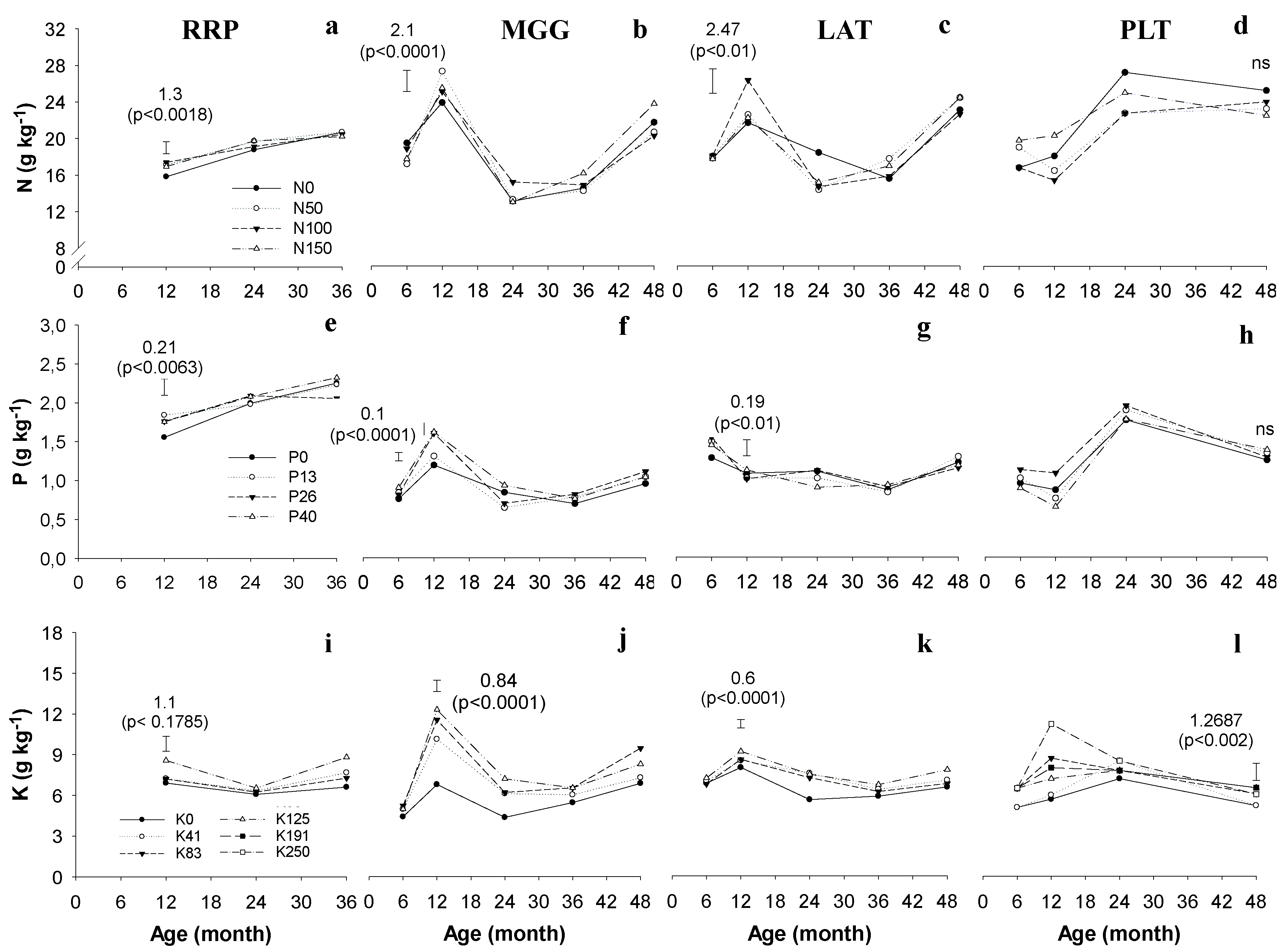

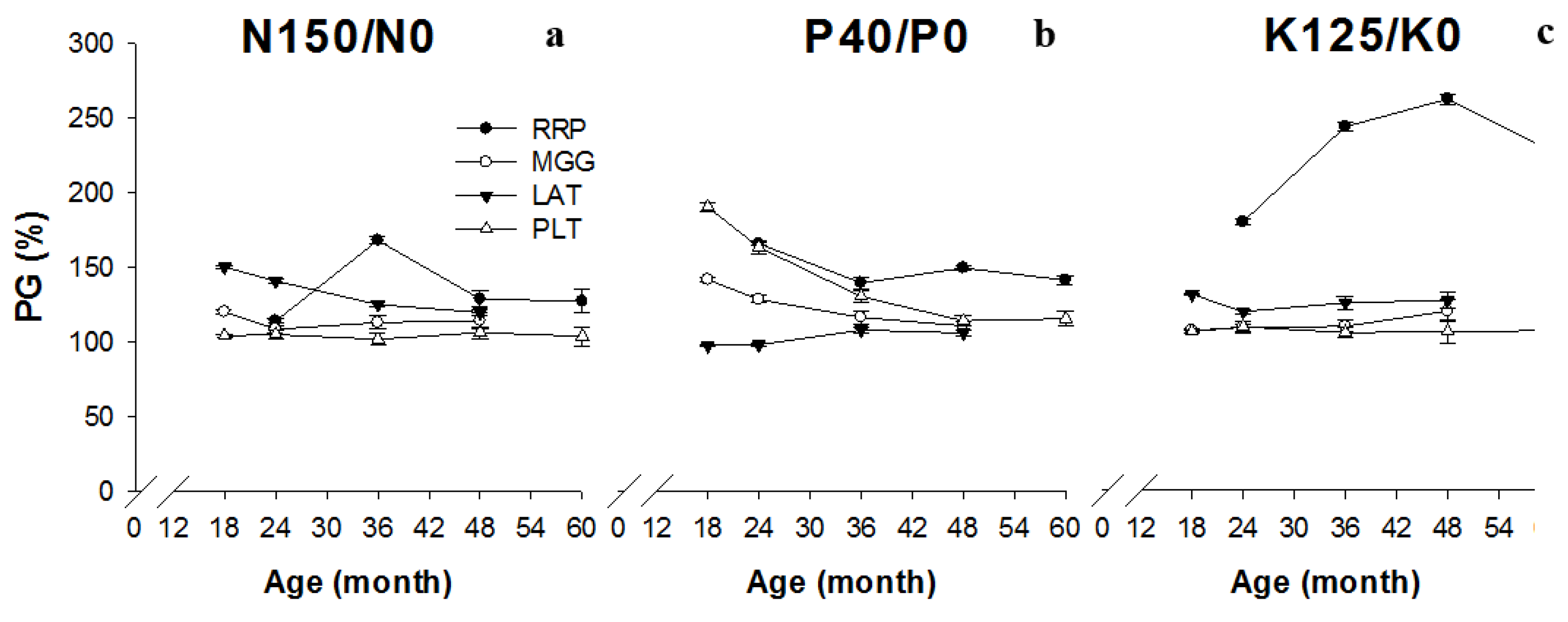
| Nutrient | Site | Age (years) | NOR 1 | Equation | R2 | p |
|---|---|---|---|---|---|---|
| N | MGG | 2 | >150.0 | SV 2 = 22.3555 + 0.027 N | 0.91 | 0.05 |
| 4 | >150.0 | SV = 119.5067 + 0.1295 N | 0.88 | 0.01 | ||
| LAT | 2 | >150.0 | SV = 12.9599 + 0.4441 N | 0.98 | 0.01 | |
| 4 | >150.0 | SV = 97.78 + 0.1385 N | 0.97 | 0.01 | ||
| PLT | 2 | 55.0 | SV = 90.8829 + 0.01111 N + 0.001 N2 | 0.99 | 0.05 | |
| 4 | >150.0 | SV = 197.2356 + 0.0821 N | 0.99 | 0.01 | ||
| 5 | >150.0 | SV = 260.5 + 0.0683 N | 0.96 | 0.01 | ||
| RRP | 2 | ns 3 | ||||
| 4 | 83.0 | SV = 83.17 + 0.83 N − 0.005 N2 | 0.98 | 0.01 | ||
| 5 | 84.0 | SV = 92.6145 + 1.4245 N − 0.0085 N2 | 0.98 | 0.01 | ||
| P | MGG | 2 | - | SV= 39.1693 + 13.0978 P/(7,7766 + P) | 0.99 | 0.01 |
| 4 | 25.0 | SV = 121.527 + 1.591 P − 0.032 P2 | 0.99 | 0.01 | ||
| LAT | 2 | ns | ||||
| 4 | ns | |||||
| PLT | 2 | - | SV = 59.6622 + 43.7815 P/(9.2192 + P) | 0.98 | 0.05 | |
| 4 | - | SV = 185.2123 + 32.2774 P/(9.0247 + P) | 0.99 | 0.01 | ||
| 5 | - | SV = 234.5716 + 40.6509 P/(4.3426 + P) | 0.99 | 0.01 | ||
| RRP | 2 | - | SV = 9.8719 + 7.5309 P/(6.253 + P) | 0.99 | 0.01 | |
| 4 | - | SV = 92.8321 + 28.4579 P/(20.8669 + P) | 0.98 | 0.01 | ||
| 5 | >40.0 | SV = 105.568 + 0.468 P | 0.98 | 0.01 | ||
| K | MGG | 2 | 117.0 | SV = 46.14 + 0.07 K − 0.0003 K2 | 0.89 | 0.05 |
| 4 | 184.0 | SV = 112.275 + 0.2577 K − 0.0007 K2 | 0.99 | 0.05 | ||
| LAT | 2 | - | ns | |||
| 4 | 103.00 | SV = 94.7431 + 0.39 K − 0.0019 K2 | 0.91 | 0.05 | ||
| PLT | 2 | - | ns | |||
| 4 | - | ns | ||||
| 5 | - | ns | ||||
| RRP | 2 | 148.0 | SV = 10.4628 + 0.1182 K − 0.0004 K2 | 0.99 | 0.01 | |
| 4 | >125.0 | SV = 58.92 + 0.47 K | 0.95 | 0.01 | ||
| 5 | >125.0 | SV = 68.62 + 0.61 K | 0.94 | 0.01 |
4. Discussion
4.1. Leaf Nutrient Contents
4.2. Leaf Area Index
4.3. Productivity of Plantations in Response to Fertilizer Application
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Goncalves, J.L.M.; Stape, J.L.; Laclau, J.P.; Bouillet, J.P.; Ranger, J. Assessing the effects of early silvicultural management on long-term site productivity of fast-growing eucalypt plantations: The Brazilian experience. South. For. 2008, 70, 105–118. [Google Scholar] [CrossRef]
- Goncalves, J.L.D.; Alvares, C.A.; Higa, A.R.; Silva, L.D.; Alfenas, A.C.; Stahl, J.; Ferraz, S.F.D.; Lima, W.D.P.; Brancalion, P.H.S.; Hubner, A.; et al. Integrating genetic and silvicultural strategies to minimize abiotic and biotic constraints in Brazilian eucalypt plantations. For. Ecol. Manag. 2013, 301, 6–27. [Google Scholar] [CrossRef]
- Brazilian Tree Industry (IBA). Relatório ibá 2015. Available online: http://iba.org/images/shared/iba_2015.pdf (accessed on 10 September 2015).
- Smethurst, P.; Baillie, C.; Cherry, M.; Holz, G. Fertilizer effects on LAI and growth of four Eucalyptus nitens plantations. For. Ecol. Manag. 2003, 176, 531–542. [Google Scholar] [CrossRef]
- Smethurst, P.; Holz, G.; Moroni, M.; Baillie, C. Nitrogen management in Eucalyptus nitens plantations. For. Ecol. Manag. 2004, 193, 63–80. [Google Scholar] [CrossRef]
- Pulito, A.P.; de Moraes Goncalves, J.L.; Smethurst, P.J.; Arthur Junior, J.C.; Alvares, C.A.; Tertulino Rocha, J.H.; Huebner, A.; de Moraes, L.F.; Miranda, A.C.; Kamogawa, M.Y.; et al. Available nitrogen and responses to nitrogen fertilizer in Brazilian eucalypt plantations on soils of contrasting texture. Forests 2015, 6, 973–991. [Google Scholar] [CrossRef]
- Fernandez, J.Q.P.; Dias, L.E.; Barros, N.F.; Novais, R.F.; Moraes, E.J. Productivity of Eucalyptus camaldulensis affected by rate and placement of two phosphorus fertilizers to a Brazilian oxisol. For. Ecol. Manag. 2000, 127, 93–102. [Google Scholar] [CrossRef]
- Almeida, J.C.R.; Laclau, J.P.; Goncalves, J.L.D.; Ranger, J.; Saint-Andre, L. A positive growth response to NaCl applications in Eucalyptus plantations established on K-deficient soils. For. Ecol. Manag. 2010, 259, 1786–1795. [Google Scholar] [CrossRef]
- Bazani, J.H.; Gonçalves, J.L.M.; Rocha, J.H.T.; Melo, E.A.S.C.; Prieto, M. Nutrição fosfatada em plantações de eucalipto. Inf. Agron. 2014, 148, 11. [Google Scholar]
- Christina, M.; le Maire, G.; Battie-Laclau, P.; Nouvellon, Y.; Bouillet, J.-P.; Jourdan, C.; de Moraes Goncalves, J.L.; Laclau, J.-P. Measured and modeled interactive effects of potassium deficiency and water deficit on gross primary productivity and light-use efficiency in Eucalyptus grandis plantations. Glob. Chang. Biol. 2015, 21, 2022–2039. [Google Scholar] [CrossRef] [PubMed]
- Sette, C.R., Jr.; Filho, M.T.; Dias, C.T.S.; Laclau, J.P. Crescimento em diâmetro do tronco das árvores de Eucalyptus grandis W. Hill. Ex. Maiden e relação com as variáveis climáticas e fertilização mineral. Rev. Árvore 2010, 34, 979–990. [Google Scholar] [CrossRef]
- Forrester, D.I.; Medhurst, J.L.; Wood, M.; Beadle, C.L.; Valencia, J.C. Growth and physiological responses to silviculture for producing solid-wood products from Eucalyptus plantations: An australian perspective. For. Ecol. Manag. 2010, 259, 1819–1835. [Google Scholar] [CrossRef]
- Forrester, D.I.; Collopy, J.J.; Beadle, C.L.; Warren, C.R.; Baker, T.G. Effect of thinning, pruning and nitrogen fertiliser application on transpiration, photosynthesis and water-use efficiency in a young Eucalyptus nitens plantation. For. Ecol. Manag. 2012, 266, 286–300. [Google Scholar] [CrossRef]
- Goncalves, J. Silvicultural effects on the productivity and wood quality of eucalypt plantations. For. Ecol. Manag. 2004, 193, 45–61. [Google Scholar] [CrossRef]
- Bouillet, J.-P.; Laclau, J.-P.; de Moraes Goncalves, J.L.; Voigtlaender, M.; Gava, J.L.; Leite, F.P.; Hakamada, R.; Mareschal, L.; Mabiala, A.; Tardy, F.; et al. Eucalyptus and Acacia tree growth over entire rotation in single- and mixed-species plantations across five sites in Brazil and Congo. For. Ecol. Manag. 2013, 301, 89–101. [Google Scholar] [CrossRef]
- Jourdan, C.; Silva, E.V.; Goncalves, J.L.M.; Ranger, J.; Moreira, R.M.; Laclau, J.P. Fine root production and turnover in Brazilian Eucalyptus plantations under contrasting nitrogen fertilizer application regimes. For. Ecol. Manag. 2008, 256, 396–404. [Google Scholar] [CrossRef]
- Battie-Laclau, P.; Laclau, J.-P.; Domec, J.-C.; Christina, M.; Bouillet, J.-P.; Piccolo, M.D.C.; de Moraes Goncalves, J.L.; Moreira e Moreira, R.; Krusche, A.V.; Bouvet, J.-M.; et al. Effects of potassium and sodium supply on drought-adaptive mechanisms in Eucalyptus grandis plantations. New Phytol. 2014, 203, 401–413. [Google Scholar] [CrossRef] [PubMed]
- Jesus, G.L.; de Barros, N.F.; da Silva, I.R.; Neves, J.C.L.; Henriques, E.P.; Lima, V.C.; Fernandes, L.V.; Soares, E.M.B. Effect of nitrogen rates and sources on Eucalyptus yield and soil organic matter fractions in the cerrado region of Minas Gerais. Rev. Bras. Ciênc. Solo 2012, 36, 201–214. [Google Scholar] [CrossRef]
- Aguiar Ferreira, J.M.; Stape, J.L. Productivity gains by fertilisation in Eucalyptus urophylla clonal plantations across gradients in site and stand conditions. South. For. 2009, 71, 253–258. [Google Scholar]
- Mello, H.A.; Mascarenhas Sobrinho, J.; Simões, J.W.; do Couto, H.T.Z. Resultados da aplicação de fertilizantes minerais na produção de madeira de Eucalyptus saligna sm. Em solos de cerrado do estado de são paulo. IPEF 1970, 1, 20. [Google Scholar]
- Silva, P.H.M.; Poggiani, F.; Libardi, P.L.; Goncalves, A.N. Fertilizer management of eucalypt plantations on sandy soil in Brazil: Initial growth and nutrient cycling. For. Ecol. Manag. 2013, 301, 67–78. [Google Scholar] [CrossRef]
- Smethurst, P.J. Forest fertilizer application: Trends in knowledge and practice compared to agriculture. Plant Soil 2010, 335, 83–100. [Google Scholar] [CrossRef]
- Gava, J.L.; Goncalves, J.L.M.; Shibata, F.Y.; Corradini, L. Eficiência relativa de fertilizantes fosfatados no crescimento inicial de eucalipto cultivado em solos do cerrado. Rev. Bras. Ciênc. Solo 1997, 21, 497–504. [Google Scholar] [CrossRef]
- Lopes, A.S.; Cox, F.R. A survey of the fertility status of surface soils under cerrado vegetation in Brazil. Soil. Sci. Soc. Am. J. 1977, 41, 742–747. [Google Scholar] [CrossRef]
- Vendrame, P.R.S.; Brito, O.R.; Guimaraes, M.F.; Martins, E.S.; Becquer, T. Fertility and acidity status of latossolos (oxisols) under pasture in the Brazilian cerrado. An. Acad. Bras. Cienc. 2010, 82, 1085–1094. [Google Scholar] [CrossRef] [PubMed]
- Valenti, M.W.; Cianciaruso, M.V.; Batalha, M.A. Seasonality of litterfall and leaf decomposition in a cerrado site. Braz. J. Biol. 2008, 68, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Thornthwaite, C.W.; Mather, J.R. The Water Balance; Drexel Institute of Technology, Laboratory of Climatology: Centerton, NJ, USA, 1955; p. 104. [Google Scholar]
- Soil Survey Staff. Keys to Soil Taxonomy; USDA, National Resources Conservation Service, National Soil Survey Center: Lincoln, NE, USA, 2014; Volume 12. [Google Scholar]
- Empresa Brasileira de Pesquisa Agropecuária—EMBRAPA. Manual de Métodos de Análise de Solo; EMBRAPA: Rio de Janeiro, Brazil, 1999; p. 212. [Google Scholar]
- Van Raij, B.; Andrade, J.C.; Cantarella, H.; Quaggio, J.A. Análise Química Para Avaliação da Fertilidade de Solos Tropicais; Instituto Agronômico de Campinas: Campinas, Brazil, 2001; p. 285. [Google Scholar]
- Laclau, J.P.; Ranger, J.; Goncalves, J.L.D.; Maquere, V.; Krusche, A.V.; M’Bou, A.T.; Nouvellon, Y.; Saint-Andre, L.; Bouillet, J.P.; Piccolo, M.D.; et al. Biogeochemical cycles of nutrients in tropical Eucalyptus plantations main features shown by intensive monitoring in Congo and Brazil. For. Ecol. Manag. 2010, 259, 1771–1785. [Google Scholar] [CrossRef]
- Malavolta, E.; Vitti, G.C.; Oliveira, A.S. Avaliação do Estado Nutricional das Plantas; Princípios e Aplicações Potafos: Piracicaba, Brazil, 1997; p. 319. [Google Scholar]
- Schleppi, P.; Conedera, M.; Sedivy, I.; Thimonier, A. Correcting non-linearity and slope effects in the estimation of the leaf area index of forests from hemispherical photographs. Agric. For. Meteorol. 2007, 144, 236–242. [Google Scholar] [CrossRef]
- Campoe, O.C.; Stape, J.L.; Teixeira Mendes, J.C. Can intensive management accelerate the restoration of Brazil’s Atlantic forests? For. Ecol. Manag. 2010, 259, 1808–1814. [Google Scholar] [CrossRef]
- Neto, C.G.; Hakamada, R.; Otto, M.S.G.; Stape, J.L.; Silva, S.R. Calibração de dois métodos indiretos para estimativa do índice de área foliar em plantações de Eucalyptus. Sci. For. (accepted).
- Gonçalves, J.L.M. Fertilização de plantação de eucalipto. In Anais do 2° Encontro Brasileiro de Silvicultura; Gonçalves, J.L.M., Pulito, A.P., Arthur Júnior, J.C., Silva, L.D., Eds.; PTSM; IPEF; ESALQ; FUPEF: Piracicaba, Brazil, 2011; pp. 85–114. [Google Scholar]
- Tuner, J.; Lambert, M.J. Nutrient cycling in age sequences of two Eucalyptus plantation species. For. Ecol. Manag. 2008, 255, 1701–1712. [Google Scholar] [CrossRef]
- Misra, R.K.; Turnbull, C.R.A.; Cromer, R.N.; Gibbons, A.K.; LaSala, A.V.; Ballard, L.M. Below- and above-ground growth of Eucalyptus nitens in a young plantation II. Nitrogen and phosphorus. For. Ecol. Manag. 1998, 106, 295–306. [Google Scholar] [CrossRef]
- Battaglia, M.; Cherry, M.L.; Beadle, C.L.; Sands, P.J.; Hingston, A. Prediction of leaf area index in eucalypt plantations: Effects of water stress and temperature. Tree Physiol. 1998, 18, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Chen, J.M.; Jin, G.; Qi, Y. Estimating seasonal variations of leaf area index using litterfall collection and optical methods in four mixed evergreen-deciduous forests. Agric. For. Meteorol. 2015, 209, 36–48. [Google Scholar] [CrossRef]
- Xavier, A.C.; Soares, J.V.; Almeida, A.C. Variação do índice de área foliar em clones de eucalipto ao longo de seu ciclo de crescimento. Rev. Árvore 2002, 26, 7. [Google Scholar] [CrossRef]
- Otto, M.S.G.; Hubbard, R.M.; Binkley, D.; Stape, J.L. Dominant clonal Eucalyptus grandis × urophylla trees use water more efficiently. For. Ecol. Manag. 2014, 328, 117–121. [Google Scholar] [CrossRef]
- Barros, N.F.; Novais, R.F.; Neves, J.C.L. Recomendação de fertilizantes minerais em plantios de eucalipto. In Nutrição e Fertilização Florestal; Gonçalves, J.L.M., Benedetti, V., Eds.; IPEF: Piracicaba, Brazil, 2000; pp. 270–283. [Google Scholar]
- Epstein, E.; Bloom, A.J. Nutrição Mineral de Plantas: Princípios E Perspectivas; Editora Planta: Londrina, Brazil, 2006; p. 403. [Google Scholar]
- Xu, D.; Dell, B.; Malajczuk, N.; Gong, M. Effects of P fertilisation on productivity and nutrient accumulation in a E. grandis x urophylla plantation in southern China. For. Ecol. Manag. 2002, 161, 89–100. [Google Scholar] [CrossRef]
- Xu, D.; Dell, B.; Yang, Z.; Malajczuk, N.; Gong, M. Effects of phosphorus application on productivity and nutrient accumulation of a Eucalyptus urophylla plantation. J. Trop. For. Sci. 2005, 17, 447–461. [Google Scholar]
- Novais, R.F.; Barros, N.F.; Neves, J.C.L. Interpretação de análise química do solo para o crescimento e desenvolvimento de Eucalyptus spp.:Níveis críticos de implantação e manutenção. Rev. Árvore 1986, 10, 105–111. [Google Scholar]
- Almeida, J.C.R. Nutrição, crescimento, eficiência de uso de água e de nutrientes em povoamentos de Eucalyptus grandis fertilizados com potássio e sódio. In Tese—Escola Superior de Agricultura Luiz de Queiroz; Universidade de São Paulo: Piracicaba, Brazil, 2009. [Google Scholar]
- Laclau, J.-P.; da Silva, E.A.; Lambais, G.R.; Bernoux, M.; le Maire, G.; Stape, J.L.; Bouillet, J.-P.; de Moraes Goncalves, J.L.; Jourdan, C.; Nouvellon, Y. Dynamics of soil exploration by fine roots down to a depth of 10 m throughout the entire rotation in Eucalyptus grandis plantations. Front. Plant Sci. 2013, 4, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Stape, J.L.; Binkley, D.; Ryan, M.G.; Fonseca, S.; Loos, R.A.; Takahashi, E.N.; Silva, C.R.; Silva, S.R.; Hakamada, R.E.; Ferreira, J.M.D.; et al. The Brazil Eucalyptus potential productivity project: Influence of water, nutrients and stand uniformity on wood production. For. Ecol. Manag. 2010, 259, 1684–1694. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melo, E.A.S.C.d.; Gonçalves, J.L.d.M.; Rocha, J.H.T.; Hakamada, R.E.; Bazani, J.H.; Wenzel, A.V.A.; Arthur, J.C., Jr.; Borges, J.S.; Malheiros, R.; Lemos, C.C.Z.d.; et al. Responses of Clonal Eucalypt Plantations to N, P and K Fertilizer Application in Different Edaphoclimatic Conditions. Forests 2016, 7, 2. https://doi.org/10.3390/f7010002
Melo EASCd, Gonçalves JLdM, Rocha JHT, Hakamada RE, Bazani JH, Wenzel AVA, Arthur JC Jr., Borges JS, Malheiros R, Lemos CCZd, et al. Responses of Clonal Eucalypt Plantations to N, P and K Fertilizer Application in Different Edaphoclimatic Conditions. Forests. 2016; 7(1):2. https://doi.org/10.3390/f7010002
Chicago/Turabian StyleMelo, Eduardo Aparecido Sereguin Cabral de, José Leonardo de Moraes Gonçalves, José Henrique Tertulino Rocha, Rodrigo Eiji Hakamada, José Henrique Bazani, Andrea Virginia Athayde Wenzel, José Carlos Arthur, Jr., Jarbas Silva Borges, Rogério Malheiros, Cristiane Camargo Zani de Lemos, and et al. 2016. "Responses of Clonal Eucalypt Plantations to N, P and K Fertilizer Application in Different Edaphoclimatic Conditions" Forests 7, no. 1: 2. https://doi.org/10.3390/f7010002
APA StyleMelo, E. A. S. C. d., Gonçalves, J. L. d. M., Rocha, J. H. T., Hakamada, R. E., Bazani, J. H., Wenzel, A. V. A., Arthur, J. C., Jr., Borges, J. S., Malheiros, R., Lemos, C. C. Z. d., Ferreira, E. V. d. O., & Ferraz, A. D. V. (2016). Responses of Clonal Eucalypt Plantations to N, P and K Fertilizer Application in Different Edaphoclimatic Conditions. Forests, 7(1), 2. https://doi.org/10.3390/f7010002