
Variation in Trembling Aspen and White Spruce Wood Quality Grown in Mixed and Single Species Stands in the Boreal Mixedwood Forest

Abstract
:1. Introduction
2. Experimental Section
2.1. Sample Procurement
2.2. Wood Density
2.3. Cellulose Microfibril Angle (MFA)
2.4. Fibre Traits
2.5. Cell Wall Chemistry
2.6. Social Classes
2.7. Statistical Analysis
3. Results and Discussion
3.1. Wood Density
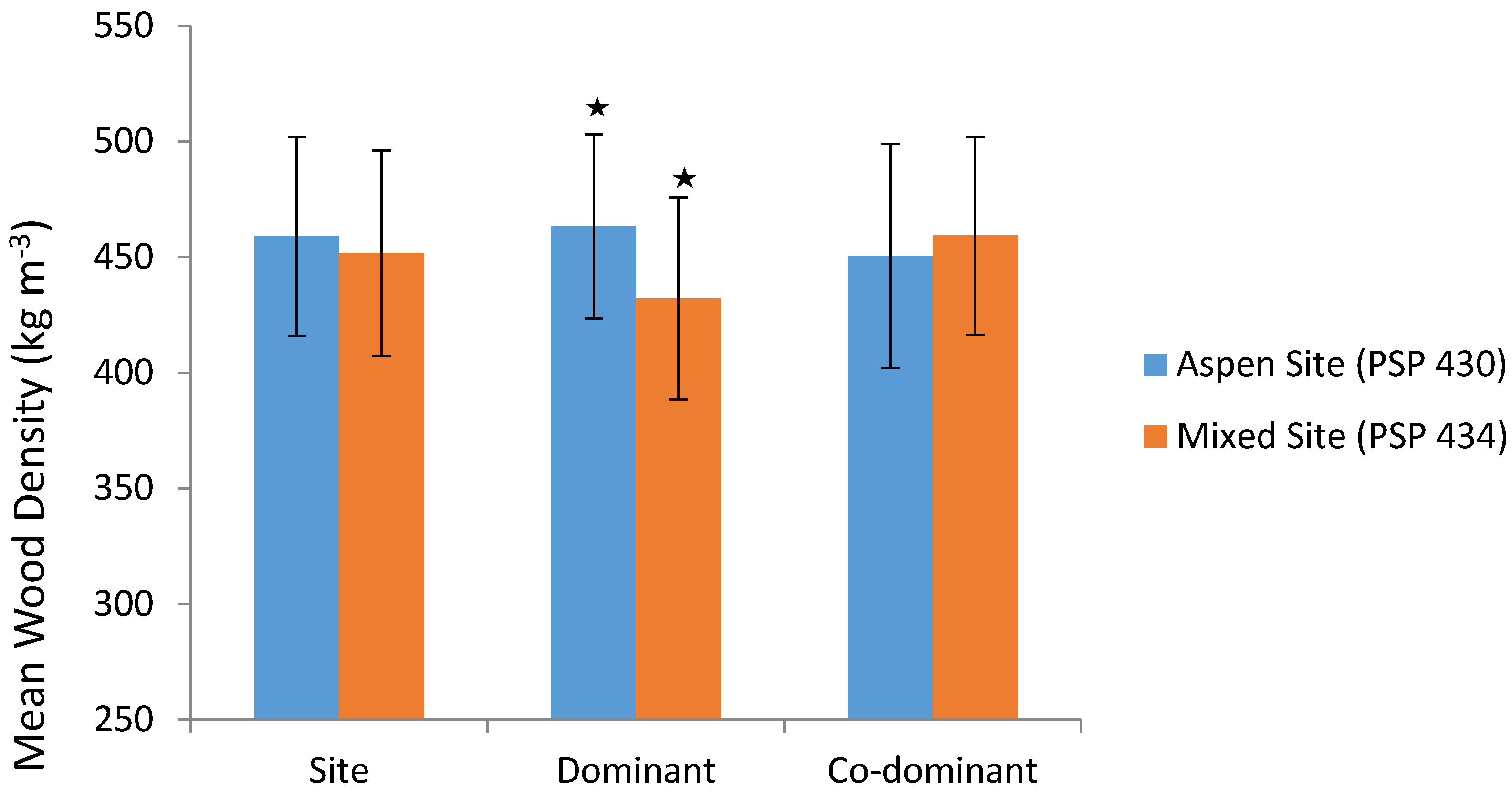
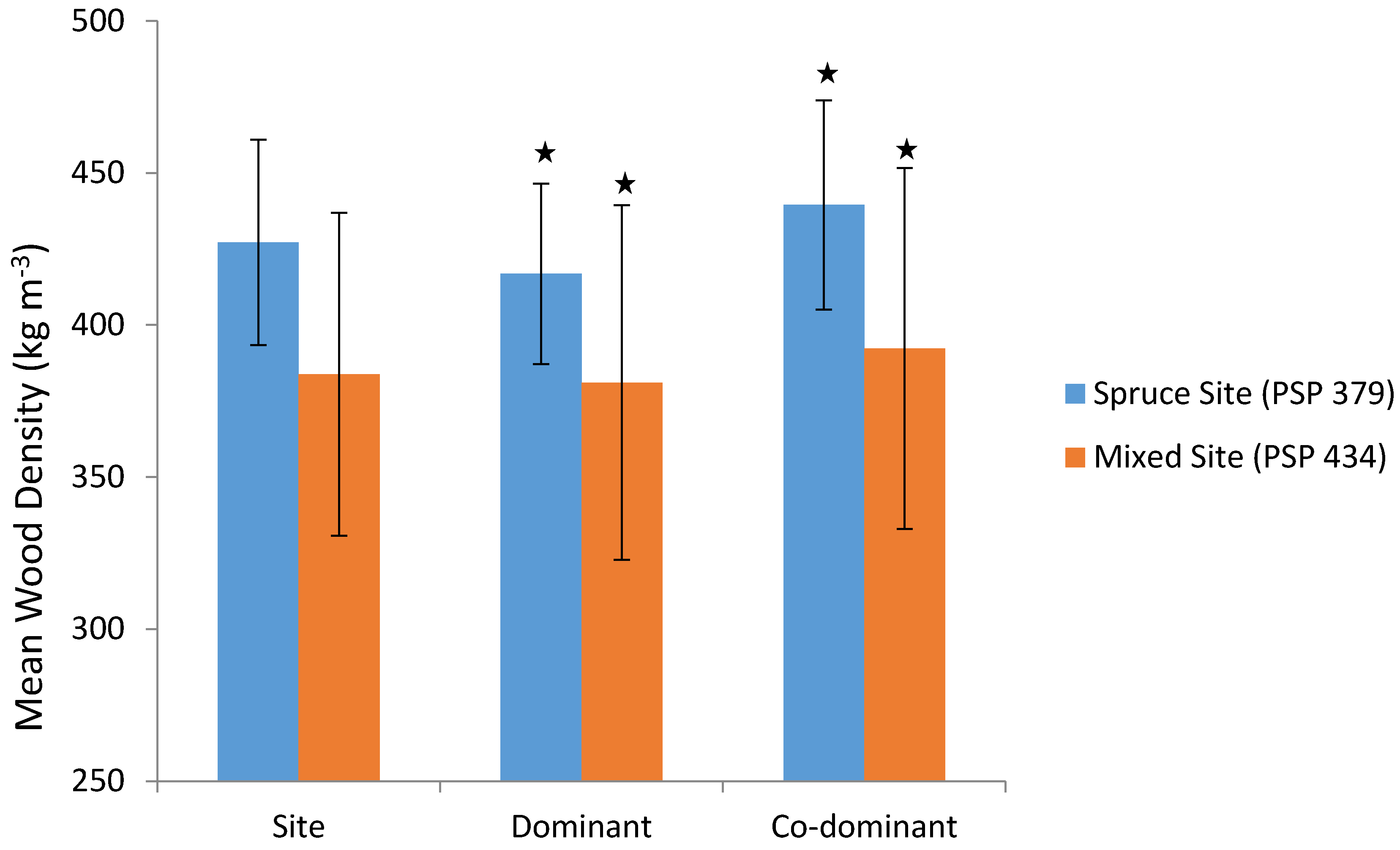
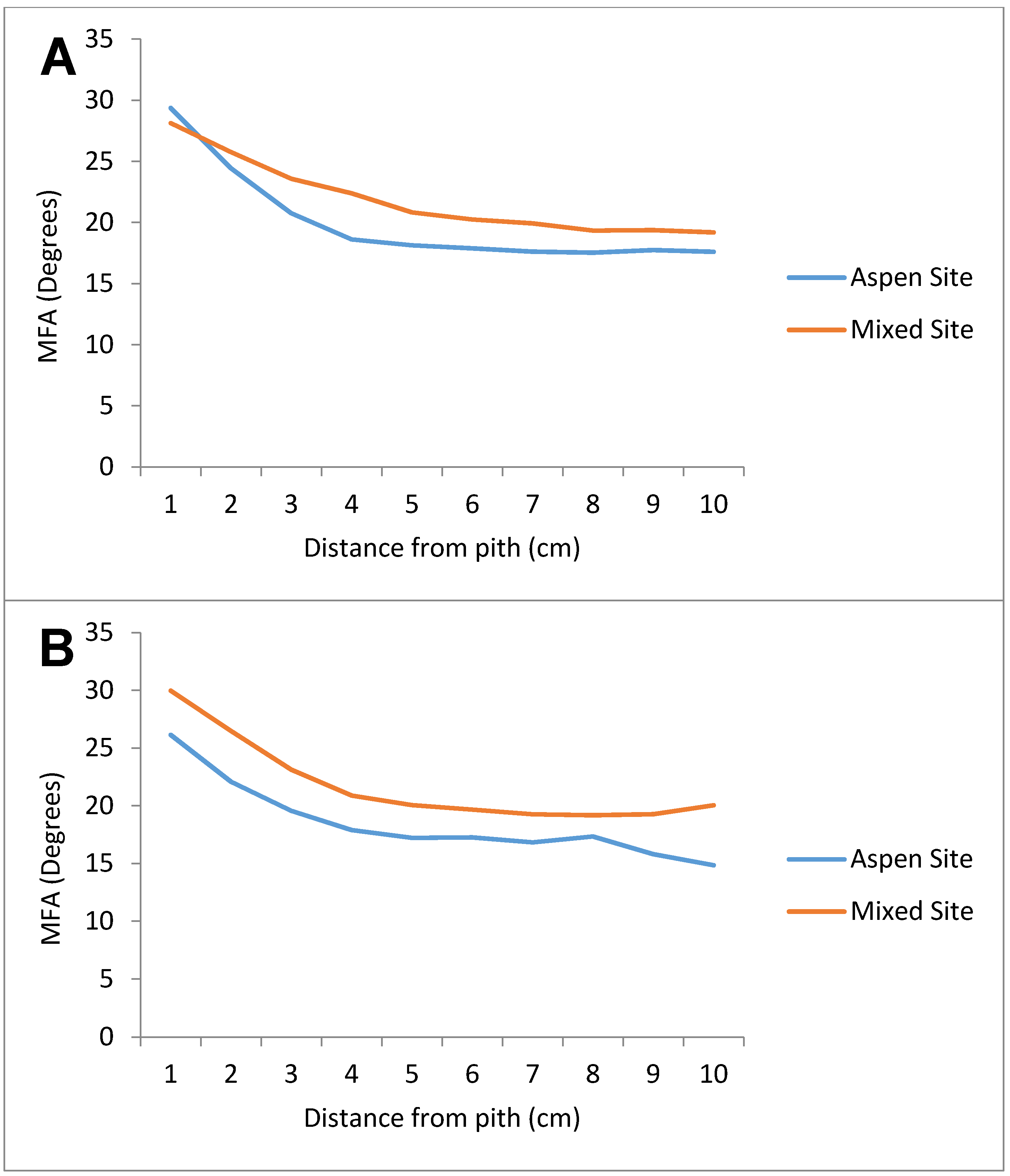
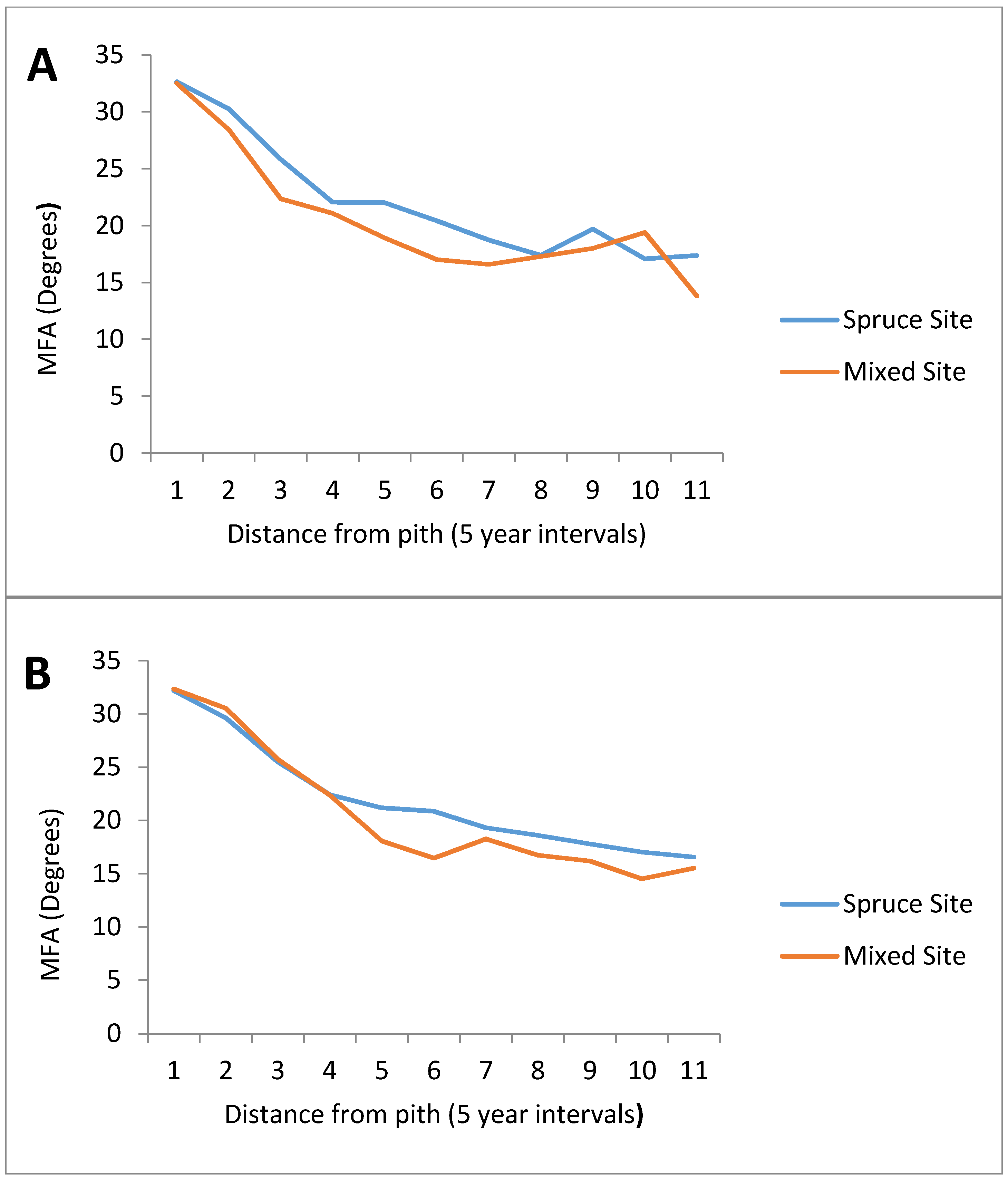
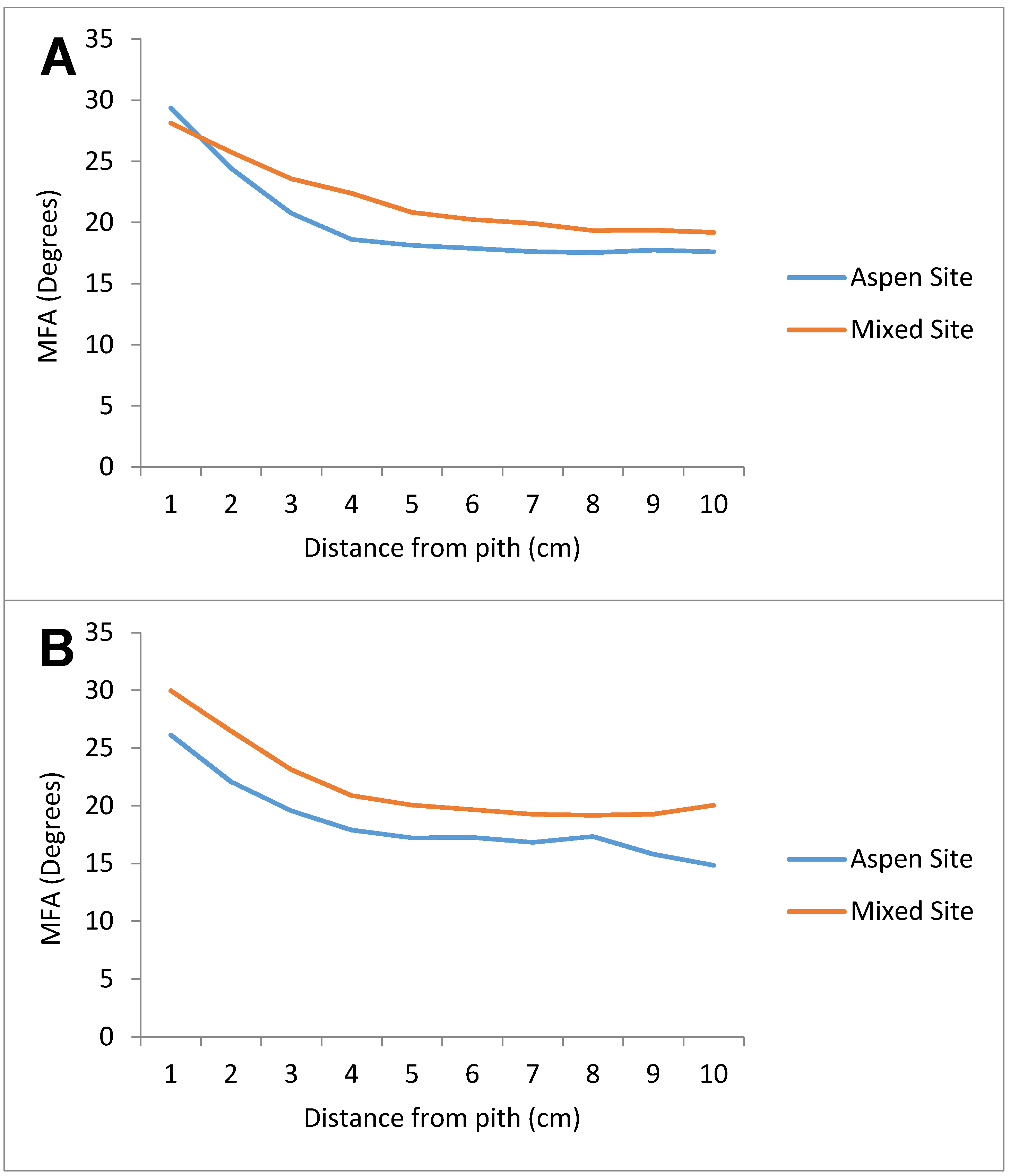
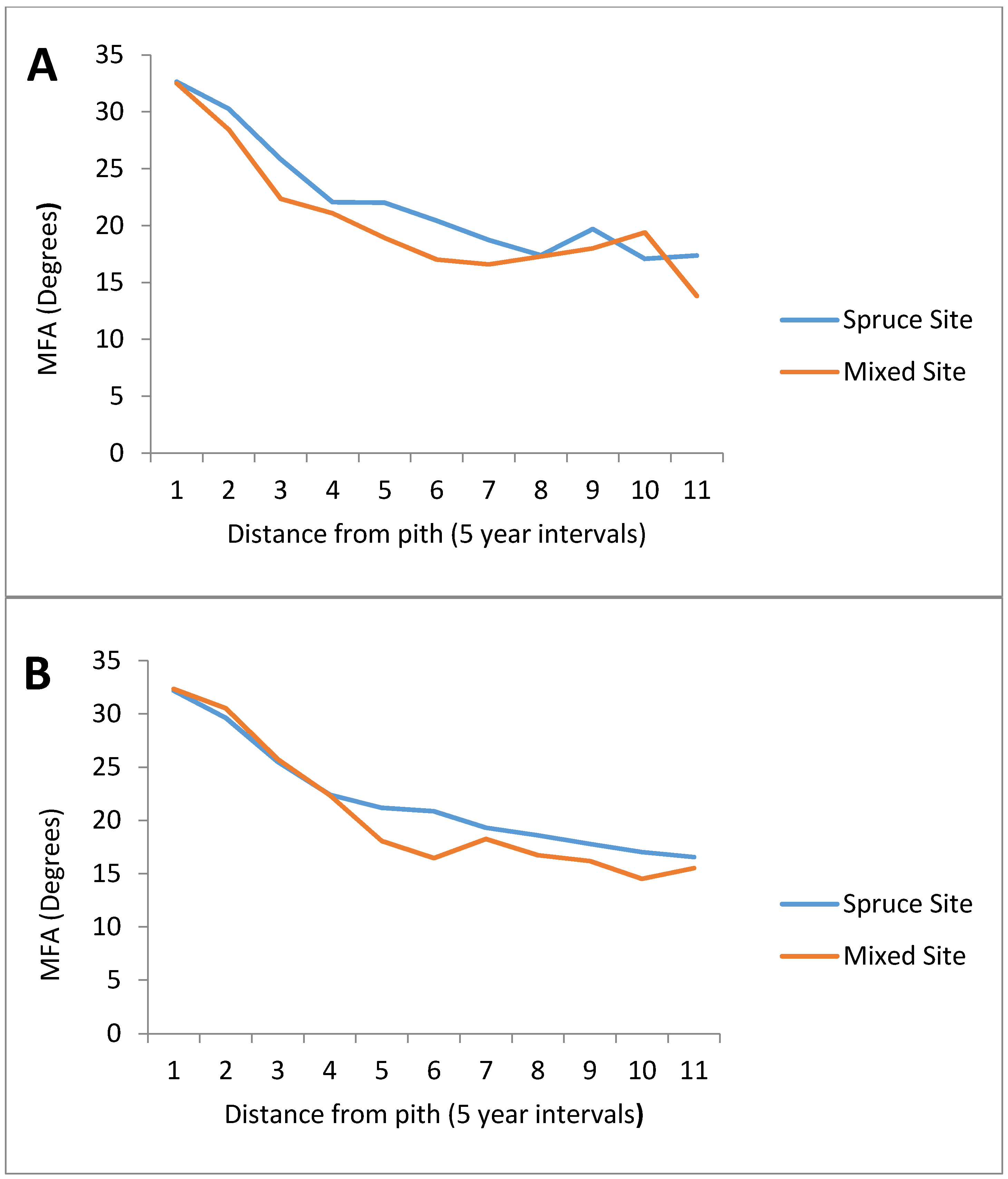
3.2. Microfibril Angle (MFA)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Distance (cm) | Aspen Site | Mixed Site |
|---|---|---|
| 0 | 28.59 | 29.46 |
| 1 | 23.84 | 26.28 |
| 2 | 20.50 | 23.25 |
| 3 | 18.47 | 21.30 |
| 4 | 17.90 | 20.27 |
| 5 | 17.79 | 19.83 |
| 6 | 17.53 | 19.45 |
| 7 | 17.64 | 19.23 |
| 8 | 17.76 | 19.30 |
| 9 | 17.45 | 19.60 |
| 10 | - | 19.55 |
| 11 | - | 18.71 |
| 12 | - | 18.74 |
| 13 | - | 17.49 |
| Age (Years) | Spruce Site | Mixed Site |
|---|---|---|
| 1 | 32.17 | 32.28 |
| 5 | 29.78 | 29.69 |
| 10 | 25.45 | 24.30 |
| 15 | 22.06 | 21.86 |
| 20 | 21.54 | 18.45 |
| 25 | 20.54 | 16.74 |
| 30 | 18.90 | 17.35 |
| 35 | 18.03 | 17.13 |
| 40 | 18.87 | 16.85 |
| 45 | 17.06 | 17.55 |
| 50 | 17.01 | 16.22 |
| 55 | 16.45 | - |
| 60 | 16.31 | - |
| 65 | 16.68 | - |
| 70 | 16.72 | - |
| 75 | 16.06 | - |
| 80 | 16.30 | - |
| 85 | 16.47 | - |
| 90 | 16.58 | - |
| 95 | 17.03 | - |
| 100 | 17.09 | - |
| 105 | 17.94 | - |
| 110 | 18.35 | - |
| 115 | 17.99 | - |
| 120 | 16.53 | - |
| 125 | 17.17 | - |
| 130 | 17.42 | - |
3.3. Fibre Traits
| Site/Social Class | Length (mm) | Width (µm) | Coarseness (mg·m−1) |
|---|---|---|---|
| Aspen Site (PSP 430) | 0.897 ± 0.102 * | 27.0 ± 1.4 * | 0.136 ± 0.028 * |
| Dominant | 0.893 ± 0.104 | 27.0 ± 1.5 † | 0.138 ± 0.029 |
| Co-Dominant | 0.909 ± 0.094 ‡ | 26.9 ± 1.2 ‡ | 0.132 ± 0.026 |
| Mixed Site (PSP 434) | 0.804 ± 0.071 * | 24.9 ± 1.2 * | 0.124 ± 0.024 * |
| Dominant | 0.847 ± 0.062 | 25.3 ± 1.3 † | 0.127 ± 0.018 |
| Co-dominant | 0.788 ± 0.068 ‡ | 24.7 ± 1.1 ‡ | 0.123 ± 0.026 |
| Spruce Site (PSP 379) | 2.154 ± 0.245 ** | 38.1 ± 1.7 ** | 0.164 ± 0.025 ** |
| Dominant | 2.155 ± 0.244 †† | 38.7 ± 1.5 | 0.165 ± 0.025 †† |
| Co-Dominant | 2.196 ± 0.239 ‡‡ | 37.7 ± 1.7 ‡‡ | 0.163 ± 0.026 ‡‡ |
| Mixed Site (PSP 434) | 1.931 ± 0.192 ** | 37.2 ± 2.2 ** | 0.130 ± 0.019 ** |
| Dominant | 1.983 ± 0.196 †† | 38.3 ± 1.8 | 0.133 ± 0.021 †† |
| Co-dominant | 1.875 ± 0.189 ‡‡ | 36.2 ± 2.1 ‡‡ | 0.128 ± 0.018 ‡‡ |
3.4. Cell Wall Chemistry
| Site/Social Class | % Carbohydrate | % Lignin |
|---|---|---|
| Aspen Site (PSP 430) | 74.12 ± 6.44 | 22.62 ± 1.92 * |
| Dominant | 75.17 ± 4.63 | 22.32 ± 1.62 † |
| Co-Dominant | 71.38 ±9.65 | 23.41 ± 2.50 ‡ |
| Mixed Site (PSP 434) | 77.60 ± 3.38 | 20.39 ± 1.63 * |
| Dominant | 78.54 ± 1.52 | 19.91 ± 1.92 † |
| Co-dominant | 76.79 ± 4.39 | 20.81 ± 1.31 ‡ |
| Spruce Site (PSP 379) | 70.89 ± 2.50 | 25.57 ± 0.99 ** |
| Dominant | 67.16 ± 4.19 | 25.10 ± 0.77 †† |
| Co-Dominant | 68.98 ± 5.20 | 25.22 ± 0.70 ‡‡ |
| Mixed Site (PSP 434) | 70.11 ± 4.08 | 26.88 ± 0.93 ** |
| Dominant | 70.12 ± 4.71 | 26.85 ± 1.02 †† |
| Co-dominant | 70.10 ± 1.93 | 26.95 ± 0.76 ‡‡ |
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Lieffers, V.J.; Macmillan, R.B.; MacPherson, D.; Branter, K.; Stewart, J.D. Seminatural and intensive silvicultural systems for the boreal mixedwood forest. For. Chron. 1996, 72, 286–292. [Google Scholar] [CrossRef]
- Kelty, M.J. Comparative Productivity of Monocultures and Mixed Species Stands. In The Ecology and Silviculture of Mixed-Species Forests, 1st ed.; Kelty, M.J., Larson, B.C., Oliver, C.D., Eds.; Kluwer Academic Press: Dordrecht, The Netherlands, 1992; pp. 125–141. [Google Scholar]
- Man, R.; Lieffers, V.J. Are mixtures of aspen and white spruce more productive than single species stands? For. Chron. 1999, 75, 505–513. [Google Scholar] [CrossRef]
- MacPherson, D.M.; Lieffers, V.J.; Blenis, P.V. Productivity of aspen with and without a spruce understory in Alberta’s boreal mixedwood forests. For. Chron. 2001, 77, 351–356. [Google Scholar] [CrossRef]
- Green, D.F.; Kneeshaw, D.D.; Messier, C.; Lieffers, V.; Cormier, D.; Doucet, R.; Coates, K.D.; Groot, A.; Grover, G.; Calogeropoulos, C. Modelling silvicultural alternatives for conifer regeneration in boreal mixedwood stands (aspen/white spruce/balsam fir). For. Chron. 2002, 78, 281–295. [Google Scholar] [CrossRef]
- Chen, H.Y.H.; Klinka, K.; Mathey, A.H.; Wang, X.; Varga, P.; Chourmouzis, C. Are mixed-species stands more productive than single-species stands: An empirical test of three forest types in British Columbia and Alberta. Can. J. For. Res. 2003, 33, 1227–1237. [Google Scholar] [CrossRef]
- Kelty, M.J. The role of species mixtures in plantation forestry. For. Ecol. Manag. 2006, 233, 195–204. [Google Scholar] [CrossRef]
- Filipescu, C.; Comeau, P.G. Competitive interactions between aspen and white spruce vary with stand age in boreal mixedwoods. For. Ecol. Manag. 2007, 247, 175–184. [Google Scholar] [CrossRef]
- Comeau, P.G.; Cosmin, N.F.; Kabzems, R.; Delong, C. Growth of white spruce underplanted beneath spaced and unspaced aspen stands in northeastern B.C—10 year results. For. Ecol. Manag. 2009, 257, 1087–1094. [Google Scholar] [CrossRef]
- Cortini, F.; Comeau, P.G.; Bokalo, M. Trembling aspen competition and climate effects on white spruce growth in boreal mixtures of Western Canada. For. Ecol. Manag. 2012, 277, 67–73. [Google Scholar] [CrossRef]
- Smith, D.M. The Principles of Silviculture, 1st ed.; John Wiley & Sons: New York, NY, USA, 1986. [Google Scholar]
- Kabzems, A.; Senyk, J.P. A comparison of actual and potential forest land production in Saskatchewan. For. Chron. 1967, 43, 257–264. [Google Scholar] [CrossRef]
- Day, R.J.; Bell, F.W. Development of Crop Plans for Hardwood and Conifer Stands on Boreal Mixedwood Sites. In Management and Utilization of Northern Wixedwoods; Samoil, J.K., Ed.; Minister of Supply and Services Canada: Edmonton, AB, Canada, 1988. [Google Scholar]
- British Columbia Forest Inventory Division. Whole Stem Cubic Metre Volume Equations and Tables: Centimetre Diameter Class Merchantable Volume Factors; Forest Inventory Division, British Columbia Forest Services—Department of Forests: Victoria, BC, Canada, 1976. [Google Scholar]
- Hart, J.F.; de Araujo, F.; Thomas, B.R.; Mansfield, S.D. Wood quality and growth characterization across intra- and inter-specific hybrid aspen clones. Forests 2013, 4, 786–807. [Google Scholar] [CrossRef]
- Egler, F.E. Vegetation Science Concepts: I. Initial floristics composition: A factor in old field vegetation development. Plant Ecol. 1954, 4, 412–417. [Google Scholar] [CrossRef]
- Loo, J.A.; Tauer, C.G.; McNew, R.W. Genetic variation in the time of transition from juvenile to mature wood in loblolly pine (Pinus taeda L.). Silvae Genet. 1985, 34, 14–19. [Google Scholar]
- Aubry, C.A.; Adams, W.T.; Fahey, T.D. Determination of relative economic weights for multitrait selection in coastal Douglas-fir. Can. J. For. Res. 1998, 28, 1164–1170. [Google Scholar] [CrossRef]
- Pot, D.; Chantre, G.; Rozenberg, P.; Rodrigues, J.C.; Jones, G.L.; Pereira, H.; Hannrup, B.; Calahan, C.; Plomion, C. Genetic control of pulp and timber properties in maritime pine (Pinus pinaster Ait). Ann. For. Sci. 2002, 59, 563–579. [Google Scholar] [CrossRef]
- Vargas-Hernandez, J.; Adams, W.T.; Krahmer, R.L. Family variation in age trends of wood density traits in young coastal Douglas-fir. Wood Fiber Sci. 1994, 26, 229–236. [Google Scholar]
- Louzada, J.; Fonseca, F.M.A. The heritability of wood density components in Pinus pinaster and the implications for tree breeding. Ann. For. Sci. 2002, 59, 867–873. [Google Scholar] [CrossRef]
- Yanchuk, A.D.; Dancik, B.P.; Micko, M.M. Intraclonal variation in wood density of trembling aspen in Alberta. Wood Fibre Sci. 1983, 15, 387–394. [Google Scholar]
- Karki, T. Variation of wood density and shrinkage in European aspen (Populus tremula). Holz. Roh. Wekstoff 2001, 59, 79–84. [Google Scholar] [CrossRef]
- Avramidis, S.; Mansfield, S.D. On some physical properties of six aspen clones. Holzforschung 2005, 59, 54–55. [Google Scholar]
- Manfield, S.D.; Weineisen, H. Wood fiber quality and kraft pulping efficiencies of trembling aspen (Populus tremuloides Michx) clones. J. Wood Chem. Technol. 2007, 27, 135–151. [Google Scholar] [CrossRef]
- Micko, M.M.; Wang, E.; Taylor, F.; Yanchuk, A. Determination of wood specific gravity in standing white spruce using a pilodyn tester. Forest Chron. 1982, 58, 178–180. [Google Scholar] [CrossRef]
- Taylor, F.W.; Wang, E.I.C.; Yanchuk, A.; Micko, M. Specific gravity and tracheid length variation of white spruce in Alberta. Can. J. For. Res. 1982, 12, 561–566. [Google Scholar] [CrossRef]
- Corriveau, A.; Beaulieu, J.; Mothe, F. Wood density of natural white spruce populations in Quebec. Can. J. For. Res. 1987, 17, 675–682. [Google Scholar] [CrossRef]
- Bamber, R.K.; Burley, J. The Wood Properties of Radiata Pine; Commonwealth Agriculture Bureau: Slough, UK, 1983. [Google Scholar]
- Butterfield, B.G. Wood Anatomy in Relation to Wood Quality. In Wood Quality and Its Biological Basis; Barnett, J.R., Jeronimidis, G., Eds.; Blackwell Publishing Ltd.: Oxford, UK, 2003; pp. 30–83. [Google Scholar]
- Cave, I.D.; Walker, J.C.F. Stiffness of wood in fast-grown plantation softwoods: The influence of microfibril angle. For. Prod. J. 1994, 44, 43–48. [Google Scholar]
- Cramer, S.; Kretschemann, D.; Lakes, R.; Schmidt, T. Earlywood and latewood elastic properties in loblolly pine. Holzforschung 2005, 59, 531–538. [Google Scholar] [CrossRef]
- Evans, R.; Ilic, J. Rapid prediction of wood stiffness from microfibril angle and density. For. Prod. J. 2001, 51, 53–57. [Google Scholar]
- Anagnost, S.E.; Mark, R.E.; Hanna, R.B. Variation of microfibril angle within individual fibres. Wood Fiber Sci. 2002, 34, 337–349. [Google Scholar]
- Wimmer, R.; Downes, G.M.; Evans, R. Temporal variation of microfibril angle in Eucalyptus nitens grown in different irrigation regimes. Tree Physiol. 2002, 22, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Erickson, H.D.; Arima, T. Douglas-fir wood quality studies, Part II: Effects of age and stimulated growth on fibril angle and chemical constituents. Wood Sci. Technol. 1974, 8, 255–265. [Google Scholar] [CrossRef]
- Donaldson, L.A. Within- and between-tree variation in microfibril angle in Pinus radiata. N. Z. J. For. Sci. 1992, 22, 77–86. [Google Scholar]
- Ivkovich, M.; Namkoong, G.; Koshy, M. Genetic variation in wood properties of interior spruce. II. Fibre characteristics. Can. J. For. Res. 2002, 32, 2128–2139. [Google Scholar] [CrossRef]
- Hannrup, B.; Cahalan, C.; Chantre, G.; Grabner, M.; Karlsson, B.; le Bayon, I.; Jones, G.L.; Muller, U.; Pereira, H.; Rodrigues, J.; et al. Genetic parameters of growth and wood quality traits in Picea Abies. Scand. J. For. Res. 2004, 19, 14–29. [Google Scholar] [CrossRef]
- Jozsa, L.A.; Middleton, G.R. A Discussion of Wood Quality Attributes and Their Practical Implications; Special Publication No. SP-34; Forintek Canada Corp.: Vancouver, Canada, 1995. [Google Scholar]
- Downes, G.M.; Nyakuengama, J.G.; Evans, R.; Northway, R.; Blakemor, P.; Dickson, R.L.; Lausberg, M. Relationship between wood density, microfibril angle and stiffness in thinned and fertilized Pinus radiate. IAWA J. 2002, 23, 253–265. [Google Scholar] [CrossRef]
- Donaldson, L.A. Microfibril angle: Measurement, variation and relationships—A review. IAWA J. 2008, 29, 345–386. [Google Scholar] [CrossRef]
- Makinen, H.; Saranpaa, P.; Linder, S. Effect of growth rate on fibre characteristics in Norway spruce (Picea abies (L.) Karst). Holzforschung 2002, 56, 449–460. [Google Scholar] [CrossRef]
- Seth, R.; Kingsland, M.A. The reinforcing properties of softwood kraft pulps. Pulp Pap. Can. 1990, 91, 68–75. [Google Scholar]
- Ericsson, T.; Fries, A. Genetic analysis of fibre size in a full-sib Pinus sylvestris L. progeny test. Scand. J. For. Res. 2004, 19, 7–13. [Google Scholar] [CrossRef]
- Hannrup, B.; Ekberg, I.; Persson, A. Genetic correlations among wood growth, growth capacity and stem traits in Pinus sylvestris. Scand. J. For. Res. 2000, 15, 161–170. [Google Scholar] [CrossRef]
- Via, B.K.; Stine, M.; Shipe, T.F.; So, C.L.; Groom, L. Genetic improvement of fiber length and coarseness based on paper product performance and material variability: A review. IAWA J. 2004, 25, 401–414. [Google Scholar] [CrossRef]
- Yanchuk, A.D.; Micko, M.M. Radial variation of wood density and fibre length in trembling aspen. IAWA Bull. 1990, 11, 211–215. [Google Scholar] [CrossRef]
- Beaulieu, J. Genetic variation in tracheid length and relationships with growth and wood traits in eastern white spruce (Picea glauca). Wood Fiber Sci. 2003, 35, 609–616. [Google Scholar]
- Bailey, I.W.; Shepard, H.N. Sanio’s laws for the variation in size of coniferous tracheids. Bot. Gaz. 1915, 60, 66–71. [Google Scholar] [CrossRef]
- Seth, R. Fibre Quality Factors in Papermaking. I. The Importance of Fibre Length and Strength. In Material Interactions Relevant to Pulp, Paper, and Wood Industries; Caulfield, D.F., Passaretti, J.D., Sobczynski, S.F., Eds.; Material Research Society: Pittsburgh, PA, USA, 1990; pp. 125–141. [Google Scholar]
- Seth, R. Fibre Quality Factors in Papermaking. II. The Importance of Fibre Coarseness. In Material Interactions Relevant to Pulp, Paper, and Wood Industries; Caulfield, D.F., Passaretti, J.D., Sobczynski, S.F., Eds.; Material Research Society: Pittsburgh, PA, USA, 1990; pp. 142–161. [Google Scholar]
- Mansfield, S.D.; Kibblewhite, R.P. Reinforcing the potential of different eucalypt: Softwood blends during separate and co-refining. Appita J. 2000, 53, 385–392. [Google Scholar]
- Brown, G.R.; Bassoni, D.L.; Gill, G.P.; Fontana, J.R.; Wheeler, N.C.; Megraw, R.A.; Davis, M.F.; Sewell, M.M.; Tuskan, G.A.; Neale, D.B. Identification of quantitative trait loci influencing wood property traits in loblolly pine (Pinus taeda L.). III. QTL verification and candidate gene mapping. Genetics 2003, 164, 1537–1546. [Google Scholar]
- Baucher, M.; Halpin, C.; Petit-Conil, M.; Boerjan, W. Lignin: Genetic engineering and impact on pulping. Crit. Rev. Biochem. Mol. Biol. 2003, 38, 305–350. [Google Scholar] [CrossRef] [PubMed]
- Chantre, G.; Rozenberg, P.; Baonza, V.; Macchioni, N.; Le Turcq, A.; Rueff, M.; Petit-Conil, M.; Heois, B. Genetic selection within Douglas-fir (Pseudotsuga. menziesii) in Europe for papermaking uses. Ann. For. Sci. 2002, 59, 583–593. [Google Scholar] [CrossRef]
- Stewart, J.J.; Kadla, J.F.; Mansfield, S.D. The influence of lignin chemistry and ultrastructure on the pulping efficiency of clonal aspen (Populus tremuloides Michx). Holzforschung 2006, 60, 111–122. [Google Scholar] [CrossRef]
- Rowell, R.M.; Pettersen, R.; Han, J.S.; Rowell, J.S.; Tshabalala, M.A. Cell Wall Chemistry. In Handbook of Wood Chemistry and Wood Composites; Rowell, R.M., Ed.; Taylor & Francis Group: Boca Raton, FL, USA, 2005; pp. 35–77. [Google Scholar]
- Willfor, S.; Sundber, A.; Hemming, J.; Holmbom, B. Polysaccharides in some industrially important softwood species. Wood Sci. Technol. 2005, 39, 245–258. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Araujo, F.; Hart, J.F.; Mansfield, S.D. Variation in Trembling Aspen and White Spruce Wood Quality Grown in Mixed and Single Species Stands in the Boreal Mixedwood Forest. Forests 2015, 6, 1628-1648. https://doi.org/10.3390/f6051628
De Araujo F, Hart JF, Mansfield SD. Variation in Trembling Aspen and White Spruce Wood Quality Grown in Mixed and Single Species Stands in the Boreal Mixedwood Forest. Forests. 2015; 6(5):1628-1648. https://doi.org/10.3390/f6051628
Chicago/Turabian StyleDe Araujo, Francis, James F. Hart, and Shawn D. Mansfield. 2015. "Variation in Trembling Aspen and White Spruce Wood Quality Grown in Mixed and Single Species Stands in the Boreal Mixedwood Forest" Forests 6, no. 5: 1628-1648. https://doi.org/10.3390/f6051628
APA StyleDe Araujo, F., Hart, J. F., & Mansfield, S. D. (2015). Variation in Trembling Aspen and White Spruce Wood Quality Grown in Mixed and Single Species Stands in the Boreal Mixedwood Forest. Forests, 6(5), 1628-1648. https://doi.org/10.3390/f6051628