1. Introduction
A major land-use change in the British Isles in the previous century was the expansion of the forest area through the establishment of extensive planted forests of non-native conifers in the upland zones of both Britain and Ireland [
1,
2]. The main silvicultural system applied in these forests was patch clear-felling combined with artificial regeneration [
3]. There were some early critics of this simple silvicultural regime who called for more species diversification in planted conifer forests and for the use of silvicultural systems that provided greater variation in stand structure [
4,
5,
6,
7]. Nevertheless, for many years the practical impact of such criticisms was limited because the main focus of forestry activity, and of underpinning research, was to support the extensive afforestation programmes in both Britain and Ireland. These programmes had the aims of increasing the amount of forest cover and of producing usable timber at a reasonable rate of return [
8].
During the last two decades of the twentieth century there was increasing awareness of the importance of other types of ecosystem service provided by forests such as landscape, recreation and biodiversity [
2]. There was recognition that meeting these multi-functional objectives in planted forests would require a move away from the simple structures and single species stands characteristic of patch clear-felling regimes towards more irregular forests composed of a range of tree sizes and of a mixture of species [
9]. Such varied structures are considered to be beneficial for biodiversity [
10] and to be preferred by visitors [
11]. The development of irregular forests with mixed species composition is also proposed as one way of adapting British forests to projected climate change [
12,
13]. Therefore, the implementation of more complex and diversified forests is now explicitly supported in forest policy documents in various countries of the British Isles such as Ireland, Scotland and Wales [
14,
15,
16]. One means of achieving this change is through increased adoption of Continuous Cover Forestry (CCF), an approach to forest management which seeks to manage the forest ecosystem rather than purely the trees, which uses natural processes as the basis of stand management, which works with site limitations, and which seeks to create diverse stand structures with a range of species [
17,
18]. The approach is implemented by silvicultural systems which avoid the use of clear-felling and thus maintain a continuity of woodland conditions across the site (hence “continuous cover”).
Although Malcolm
et al. [
9] estimated that up to half the area of conifer planted forest in Britain could be suitable for transformation to CCF based on knowledge of soils and wind climate, they believed that perhaps only 5% of these forests were being managed using this approach. Despite the ongoing support for CCF in country policy documents, precise figures on the areas that have been placed under this type of management are difficult to obtain. A recent review of the adoption of CCF in British forestry identified some 60 thousand ha of woodlands actively managed under this approach (around 3% of forest cover) [
19]. However, this review also noted a much larger area in public forests where there were aspirations to transform forests to irregular structures in the near future: the area involved amounted to around 20% of public forests in Great Britain [
19, unpublished data]. In the Irish republic, Vitkova
et al. [
20] found just over 10 thousand ha being managed under CCF (less than 2% of the forest area) and much of this had only been designated in the last 15 years. Examination of these reviews suggests that adoption of CCF has tended to be concentrated in mixed woodlands in more sheltered regions as well as in broadleaved woodlands in public forests in Ireland. By contrast, there are relatively few examples of CCF being implemented in the single species conifer plantations established in the last century in upland Britain, and in particular in the extensive forests dominated by Sitka spruce (
Picea sitchensis (Bong.) Carr.).
Sitka spruce was introduced into the British Isles in 1831 from north-west America by David Douglas [
21]. In its natural range the species is mainly confined to coastal forests between Alaska and northern California where individual trees reach the largest dimensions recorded for any member of the genus
Picea [
22]. David Douglas had identified the species’ ability to grow to large sizes on nutrient poor shallow soils as a valuable feature for British conditions [
23]. The first major imports of seed into Britain were in 1852 and the rapid growth of specimen trees in arboreta led to the establishment of trial plantations in Scotland from 1882 [
24]. The good results from these trials, as well as from those in Ireland [
1], resulted in this non-native spruce becoming the main species used in afforestation in the British Isles from 1950 onwards. Thus in Britain the proportion of Sitka spruce rose from 11% of all conifers planted in the 1920s to nearly 70% in the 1980s [
3]. Forests of Sitka spruce are now the major forest type in both Great Britain and Ireland, covering more than 1 million ha, and there are also substantial areas planted with this species in oceanic regions of mainland Europe such as Brittany, Denmark and Norway [
25].
From the early days of its introduction into British forestry, the practice was to grow Sitka spruce in pure even-aged stands managed on comparatively short rotations (40–60 years) for the production of small roundwood and sawlogs [
24,
25,
26]. The oceanic climate of upland areas of the British Isles can result in serious risks of wind damage to these spruce stands, particularly when located on exposed sites with moist soils resulting in shallow rooting. As a result, many Sitka spruce forests have been managed either on non-thin regimes or with very limited thinning because of the risks of windthrow [
27]. Small proportions of other conifer or broadleaved species may be planted in forests dominated by Sitka spruce, but these would be deployed in discrete stands primarily for aesthetic or biodiversity reasons [
28]. The main exception to the use of spruce in pure stands was where the species was planted in “nursing mixtures” on nutrient poor soils, particularly in the presence of ericaceous vegetation (e.g., heather-
Calluna vulgaris L. Hull) when it was admixed with species such as Scots pine (
Pinus sylvestris L.), lodgepole pine (
Pinus contorta Dougl.) or Japanese larch (
Larix kaempferi Lamb. Carr.) [
29,
30]. At the end of the rotation, the stands are clear-felled with a coupe size varying from 5 to 30 ha depending upon the visibility of the site and the shape of the landform [
28]. Felling is generally followed by planting with a target stocking density at establishment of around 2500 stems ha
−1 [
25]. Thus conventional management of Sitka spruce forests would conform to an international typology of intensive tree plantations [
31].
Natural regeneration of Sitka spruce has been observed in British forests since the early 1950s [
32,
33], and stocking densities in excess of 10,000 naturally regenerated stems ha
−1 have been reported [
34,
35,
36,
37]. Such regeneration has mostly been managed through respacing (pre-commercial thinning) to develop even-aged stands of regular spacing [
37]. However, there was also recognition that the occurrence of natural regeneration offered the possibility to increase the structural and species diversity in planted forests. One of the six experimental areas initiated by Edinburgh University in the 1950s to examine the transformation of even-aged stands [
38] was located on the Corrour estate in northern Scotland in stands where Sitka spruce regeneration had been noticed in windblown gaps [
39]. Penistan [
40] recommended the use of groups of 0.1–0.4 ha for the restocking of spruce forests, accompanied by enrichment planting to increase species diversity. Studies of the effect of different gap sizes on the stability of the surrounding edge trees indicated a greater incidence of windthrow in small (0.04 ha) gaps compared to ones of 0.12 and 0.4 ha [
41]. However, as noted by Paterson [
42], for at least three decades very little research and development was undertaken to develop these observations into a framework which would allow objective consideration of the potential role of CCF in Sitka spruce dominated forests in upland Britain.
Malcolm
et al. [
9] identified three preconditions for success of transforming plantation forests and several constraints. The preconditions were: the availability of adequate seed for natural regeneration; an adequate within stand microclimate for the establishment of natural regeneration or under-planted trees; and an appropriate silvicultural system that delivered both the seed supply and the desired microclimate. Based on their characterisation of Sitka spruce as a species of intermediate shade tolerance, they proposed [
9] (table 3), that an irregular shelterwood system would be most appropriate for transforming forests of this species, using a gap size of 0.1–0.2 ha. The main constraints identified were: the loss of the regeneration niche through vegetation competition; the potential damage to regenerating seedlings from browsing animals, particularly deer; and the risk of wind damage consequent upon opening up the mature stands to develop regeneration.
Over a decade has elapsed since that review and an appreciable amount of further research has been undertaken in the British Isles to investigate some of these preconditions and constraints such as seedling growth in the understorey and the wind stability of less regular stands. In addition, a number of operational trials have been undertaken to explore the feasibility of transforming conifer planted forests, including those dominated by Sitka spruce, to more diverse and irregular structures [
19,
43,
44,
45,
46]. This paper summarises the major findings from these studies and trials, considers the knowledge gaps that still remain, and evaluates the practical implications for forest management.
3. Stand Microclimate
Given that other studies (see above) had identified that Sitka spruce was a species of intermediate shade tolerance, an area of interest was to determine how the understorey light climate was influenced by stand and canopy structure. Preliminary studies [
59,
60] indicated that, in regular Sitka spruce stands, a basal area of around 30 m
2 ha
−1 was required to provide a satisfactory light environment for the growth of regenerating seedlings. This suggestion was followed up in several studies examining the light transmittance through canopies of different stocking densities. Hale
et al. [
61] summarised results from 36 plots in six different Sitka spruce forests in upland Britain. These confirmed that, while a basal area of 30 m
2 ha
−1 or less was associated with a favourable light environment for Sitka spruce natural regeneration, this basal area figure on its own was an insufficient predictor of adequate light transmittance and the interaction with stocking density should be considered. At a basal area of 30 m
2 ha
−1 there needed to be a stocking density of <450 stems ha
−1 to provide the minimum 20% transmittance necessary for adequate seedling growth. This was because the high leaf area characteristic of Sitka spruce stands could reduce transmittance below critical levels unless a lower stocking density had provided sufficient tree clumping and irregularity in the canopy to increase light penetration to the forest floor. The combined contribution of basal area and stocking density as predictors of an adequate light environment for natural regeneration is also supported by Bertin
et al. [
51] where poorer growth of regeneration was evident in more densely stocked stands with lower transmittance values. These critical parameters may also need to be adjusted for different climates or regions since studies in Ireland suggested 5–10% lower transmittance for a given basal area [
57] (figure 6), than was found in equivalent British studies [
61]. Time since thinning will also be important because the canopy gaps and more heterogeneous canopy resulting from a recent thinning will lead to a higher RLI: this decreases with time as the canopy closes [
62].
By contrast, there have been few investigations of other aspects of the understorey microclimate of Sitka spruce stands. Sellars [
63] used the site of the seedling growth study of Mason
et al. [
46] to examine the effect of different stand densities upon air and soil temperatures as well below canopy wind speed. She found that increased canopy openness resulted in higher irradiance, greater wind speed and more pronounced fluctuations in air temperature, especially near the ground (
Table 3). A retained canopy was able to buffer the effects of heat loss to the sky during an autumn frost event. In the open area, night temperatures during this event fell to −11 °C, sufficient to cause serious damage to shoots if preceded by a period of warmer weather [
64]. By contrast, in the stands with 156 and 278 stems ha
−1, the minima recorded were −4.3 and −2.9 °C respectively, temperatures which would likely cause little damage to regenerating seedlings. In one Irish study site with three stocking densities varying from 24 to 34 m
2 ha
−1 [
57], the highest soil temperature recorded was in the stand with the lowest basal area, while soil water content also tended to be higher in this treatment.
Table 3.
Selected characteristics of the microclimate underneath a Sitka spruce stand in southern Scotland thinned to different densities (adapted from Sellars, [
63]).
Table 3.
Selected characteristics of the microclimate underneath a Sitka spruce stand in southern Scotland thinned to different densities (adapted from Sellars, [63]).
| Treatment (Stems ha−1) | Canopy Openness (%) 1 | Mean Air Temperature @ 0.23 m (°C) 2 | Difference between Maximum and Minimum Air Temperatures @ 0.23 m (°C) 3 | Soil Temperature at 0.05 m Below Ground (°C) 2 | Mean Windspeed (%) @ 1.05 m 4 |
|---|
| 625 | 0.6 | 14.7 | 5 | - | 12 |
| 278 | 4.8 | 15 | 6.4 | 13.6 | 6 |
| 156 | 28.2 | 16.1 | 14.7 | 14.6 | 34 |
| Open | 100 | 15.6 | 22.8 | 13.2 | 100 |
4. Wind Stability of Irregular Stands
The extent of adoption of CCF in Sitka spruce forests is heavily influenced by the perceived risk of windthrow, widely recognised as the major constraint upon the silvicultural options in forests in Britain and Ireland [
27,
65]. The initial recommendation was that CCF was best confined to sheltered sites on deep rooting soils where thinning at regular intervals could be used to gradually open up stands and provide a favourable regeneration environment [
9]. A review of evidence about the stability of irregular stands [
66] suggested that this recommendation might be pessimistic, since it overlooked the likelihood that dominant trees in irregular stands would be better adapted to withstand the forces of the wind, evidenced in more favourable (lower) height: diameter ratios, greater root mass, and stronger buttressing. Some preliminary modelling using the wind risk model ForestGALES [
67] supported this suggestion, indicating that irregular structures could be more stable than regularly thinned even-aged stands on sites of intermediate wind exposure and that heavy early thinning might be used to develop stable dominants as part of the transformation process [
66,
68]. A wind tunnel examination of the effect of different stand structures (even-aged, shelterwood, selection) upon the incidence of damaging gusts found similar wind and turbulence characteristics over the different types. However, in this study the dominant trees in the structure representing a selection forest were exposed to more extreme wind loading than occurred in a regular stand, but the smaller trees in irregular structures were sheltered from these extremes [
69]. In addition, the presence of an understorey, as might be found in a group selection forest, reduced the wind loading on the overstorey trees, possibly by limiting the amount of sway [
69].
Some of these factors were examined in a 54 years-old stand on an exposed site in Clocaenog forest in Wales undergoing transformation to CCF [
70] where the trees had proved considerably more wind stable than predicted [
71]. Investigation of the wind forces and critical turning moment (the force required to overturn or break a tree) for nine individually instrumented trees of different size and canopy status (
i.e., dominant, co-dominant, sub-dominant, suppressed) [
72] (
Table 2) showed a wider range of wind-tree interactions than in regular stands. Adaptive growth of individual trees had provided a balance between tree exposure to wind and the resistance to this force. The wind environment of individual trees was formalised using competition indices to characterise their status in the stand. Analysis of over 300 h of measurements for each tree showed a negative and broadly linear relationship between the turning moment calculated for an individual tree and its social status as estimated by a range of competition indices [
72]. In other words, trees in the suppressed or subdominant classes had lower turning moments than dominants.
The relationship between turning moment and competition index calculated for an individual tree was used to predict the critical wind speed for tree failure for all trees in a 0.5 ha subplot of the Clocaenog stand both without thinning and when subjected to four different simulated thinning regimes (
i.e., where trees might have a different competition index value as a result of neighbouring tree removal). All regimes were constrained to remove no more than 20% of the standing basal area and there was a proviso that no trees should be harvested adjacent to ones already selected for thinning to avoid the creation of gaps. The regimes were:
from above–removal of the tallest trees in the stand;
from below–removal of the smallest trees in the stand;
neutral–removal of the same proportion of trees in each 3 cm dbh class;
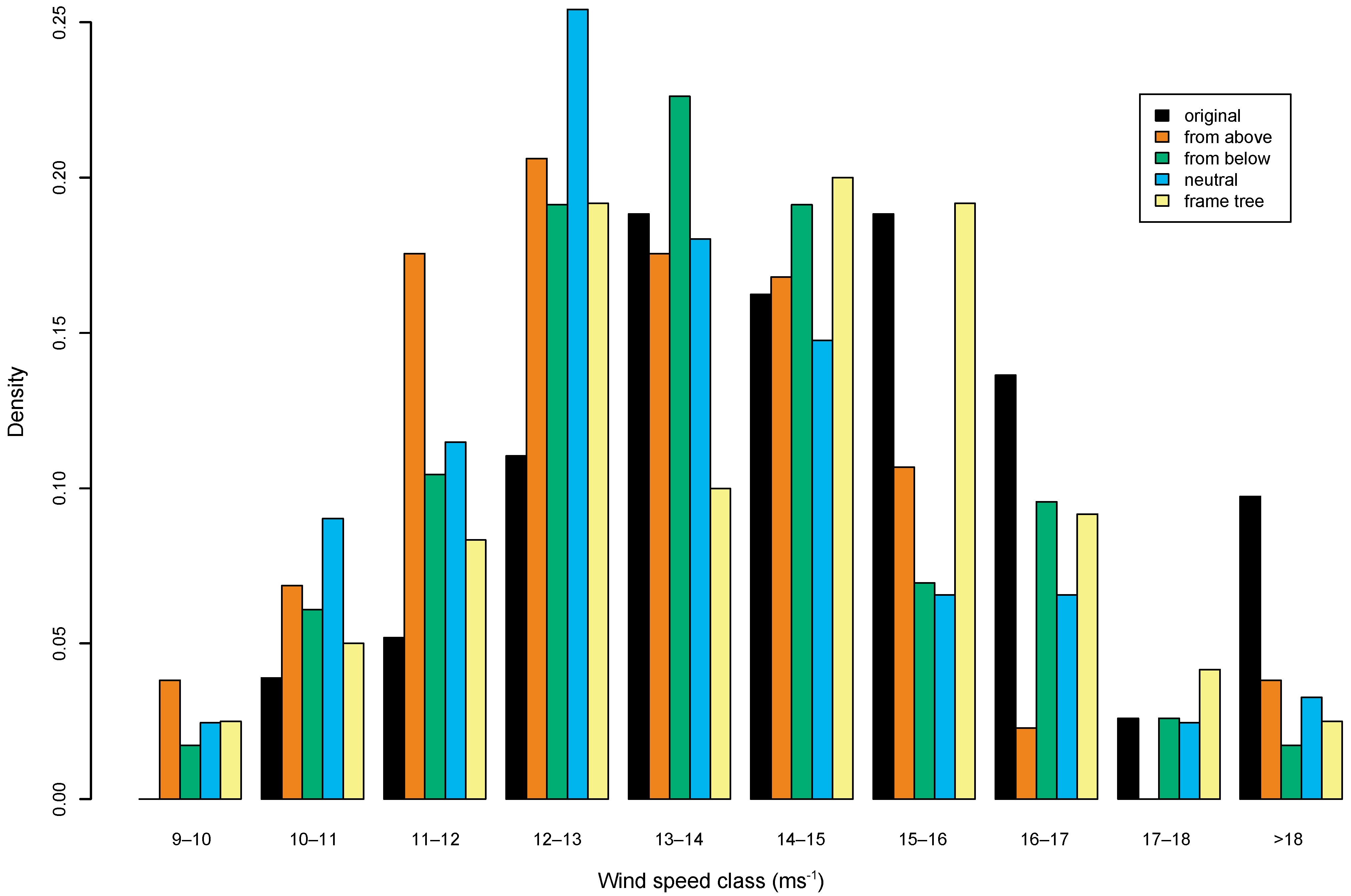
frame-tree–divide the stand into 64 blocks of equal size, identify the most stable tree based on height: diameter ratio and remove its immediate competitor. The simulated thinnings removed between 24 and 40 trees from the subplot representing around 19–23% of standing basal area. The effect of the different regimes was explored by showing the frequency distribution of all trees in the stand against critical wind speed (
Figure 1). All thinning regimes caused some decrease in the overall stability of the stand compared to the unthinned state, which is shown by there being proportionately more trees in the lower wind speed classes after thinning and fewer in the higher classes. However, there were also some interesting differences between the thinning regimes, where the frame-tree thinning tended to provide a higher proportion of trees than other regimes in the higher wind speed classes between 14 and 18 metres s
−1. By contrast, thinning from above or neutral thinning had a higher proportion of trees in the lower wind speed classes [
71].
The implication of these findings is that, while all thinning will increase the vulnerability of a stand to wind until the remaining trees have adapted to their changed wind loading, the type, intensity and frequency of thinning can affect the increase in risk. Parallel observations from the wider transformation trial area in Clocaenog forest supported these conclusions [
73], since there was clear difference between treatments in the incidence of windthrow in both 2005 and 2007 with the highest incidence of windthrow recorded in a treatment which had inadvertently caused the removal of the most stable trees (frame-trees) within the stand (
Table 4). A modelling study in the Aberfoyle Research Forest also showed that thinning from above which removed stable dominants could have a detrimental effect on stand stability [
74].
Figure 1.
Distribution (density) of calculated wind speeds causing tree failure in the range 10–18 m s−1 for the experimental stand in Clocaenog Forest for the unthinned (original) stand and after the application of four different simulated thinning scenarios (see text for details).
Figure 1.
Distribution (density) of calculated wind speeds causing tree failure in the range 10–18 m s−1 for the experimental stand in Clocaenog Forest for the unthinned (original) stand and after the application of four different simulated thinning scenarios (see text for details).
Table 4.
Effect of thinning to implement different silvicultural approaches upon subsequent wind damage in the Clocaenog research area (after Haufe, [
73]).
Table 4.
Effect of thinning to implement different silvicultural approaches upon subsequent wind damage in the Clocaenog research area (after Haufe, [73]).
| Treatment | Basal Area (m2 ha−1) after Thinning in 2004 | Cumulative Per Cent of Wind Blown Trees in 2005–2007 |
|---|
| Control-unthinned | 30.3 | 0.4 |
| Uniform shelterwood | 27.0 | 0.6 |
| Group shelterwood | 25.3 | 0.6 |
| Irregular shelterwood | 25.4 | 1.3 |
| Frame trees felled | <27.0 | 10.1 |
6. Discussion
Recent decades have seen increasing interest in the potential role of CCF as an approach to forest management in many parts of the world [
18,
89]. However, there are few reports of CCF being actively deployed in the management of planted forests of non-native species since these are normally intensively managed for wood production using even-aged management. Although various policy documents were supporting wider use of CCF in planted forests in Britain by the late 1990s [
9], recent analysis indicates that uptake of this approach is still limited and is mainly confined to forests where aspects such as recreation, amenity and biodiversity are high on the list of objectives [
19,
20]. Nevertheless, recent concerns about the potential vulnerability of single species plantations to the impacts of climate change [
13] or to the spread of pests and diseases [
90] raises the question as to whether CCF can be considered as a more resilient approach for management of planted forests. For the purposes of this discussion, it is assumed that the decision to use an alternative approach such as CCF depends as much upon its technical feasibility as its theoretical ability to deliver a range of ecosystem services.
The results reviewed in this paper illustrate the suite of information requirements and research challenges that need to be overcome if an alternative approach is to be successfully introduced into the management of planted forests, such as those of Sitka spruce in the British Isles. There are at least 15 different aspects of stand management where information has been required to elucidate the potential role of CCF in the management of planted forests of Sitka spruce (
Table 6). These can be broadly grouped into three categories covering: the establishment of natural regeneration; the tending of the stands to develop irregular structures; and various operational aspects. Examination of the quality of the evidence (
Table 6) suggests that much useful information has been gained about the requirements for successful natural regeneration of Sitka spruce so that managers can begin to predict with some confidence those sites and stands where regeneration can be anticipated, and the measures that can be taken to foster it when it occurs. Similarly, “good” to “moderate” evidence has been obtained on some of the key operational aspects that will influence the uptake of CCF, such as the ability to use conventional harvesting systems, the potential impacts on timber quality, and the cost and revenue implications of using an alternative silvicultural approach to patch clear-felling.
By contrast, in the stand tending group there are several stages where the quality of evidence is classed either as “poor” or “poor-moderate” and thus could limit the uptake of CCF. A couple of these stages involve the wind stability of irregular stands and the interaction with thinning where concern over the potential wind risks associated with the introduction of CCF may limit forest managers’ willingness to adopt this alternative silvicultural approach [
91]. The rating of this evidence is, at least in part, a reflection of improved knowledge. Thus, when guidance on CCF was first provided, there were strong caveats against using this approach on more exposed sites where windthrow was expected to be a problem [
9,
17]. This reflected a deterministic approach to wind risk management which believed that there was relatively little that silviculture could do to mitigate wind damage which would be determined by a combination of site and climate [
27]. The pessimism about the potential role of CCF on more exposed sites would also have been reinforced by the tendency of early versions of ForestGALES and previous methods of evaluating wind risk to over-predict the occurrence of wind damage [
92]. However, the results reported here [
71,
72] indicate not only that it is possible to develop and maintain irregular structures on more exposed sites than previously thought, but also that the use of crown thinning regimes which develop and favour the most stable trees in the stand may confer enhanced resistance to strong winds, provided that the retained trees have adapted to an increased wind loading following thinning. The retention of these dominant “frame” trees should also limit any risk that removal of other vigorous trees in crown thinning might degrade the genetic potential of the stand. Similar stability benefits from wind-adapted thinning regimes have been reported in Norway spruce stands in the Czech republic where heavy thinning at top heights of 10–15 m improved the sturdiness (h:d) ratio of retained trees compared to unthinned controls [
93]. The effect of thinning type was also found in study of storm damage in southern Germany where increased windthrow risk was incurred by removal of stable dominant trees in thinning [
94].
Table 6.
The main stages involved in evaluation of the feasibility of introducing CCF into the management of Sitka spruce planted forests in Britain, together with conclusions on the present situation and an assessment of the quality of the supporting evidence.
Table 6.
The main stages involved in evaluation of the feasibility of introducing CCF into the management of Sitka spruce planted forests in Britain, together with conclusions on the present situation and an assessment of the quality of the supporting evidence.
| Stage | Conclusion | Evidence Quality | Main References |
|---|
| NATURAL REGENERATION | | | |
| Seed availability | Mast years recorded in British Sitka spruce stands every 4–5 years | Good-moderate | [9,95] |
| Germination conditions | Favourable conditions involve moist soils with needle litter, light moss cover | Moderate-poor | [9] |
| Light requirements for growth of young seedlings | At least 20% RLI plus a canopy with gaps equivalent to basal area of 30 m2 ha−1 and a reduced stand density | Good | Table 1; [61] |
| Other aspects of understorey microclimate | Retain some canopy cover to limit frost damage but provide adequate light | Moderate | [48,63] |
| Vegetation competition | Avoid fertile sites or competition from ericaceous vegetation | Moderate | [9,96] |
| Browsing pressure | Keep deer population below 5 animals per 100 ha–lower levels necessary for admixed species | Moderate | [9] |
| STAND TENDING | | | |
| Management of established regeneration | Respace to
ca. 2000 stems ha−1 at 3–4 m height | Good | [37] |
| Thinning type and intensity | In young stands (<25 years old) use crown thinning to develop stability of selected dominants. Intervention in older stands depends upon wind risk. | Good-moderate for young stands; poor-moderate for older stands | [71,72] |
| Vulnerability to abiotic disturbance (e.g., wind) | Use of CCF can be extended to sites of intermediate wind exposure if soils allow good rooting and if stand will respond to thinning | Poor-moderate | [66,71] |
| Formation of mixtures | Gap environment required for Sitka spruce natural regeneration should favour regeneration/underplanting of a range of species. Little knowledge of potential long-term species for admixing with Sitka spruce. | Poor | [9] |
| Identification of appropriate silvicultural system | Irregular shelterwood proposed based on characterisation of Sitka spruce as having intermediate shade tolerance | Poor-moderate | [9] |
| Knowledge of natural stand dynamics | Wind disturbance a major driver of stand structures. Possible influence of deer browsing | Poor-moderate | [97,98] |
| OPERATIONAL | | | |
| Use of standard harvesting systems/machinery | The major problem encountered was the need to include motor manual working to deal with large trees. The lack of brash availability may be problematic in later thinnings on wet soils. | Good-moderate | [83,84] |
| Forecasting and monitoring of growth of irregular stands | A range of models evaluated and MOSES GB calibrated with promising results. Monitoring system available for CCF stands. | Moderate | [45,80] |
| Availability of long-term growth and yield data from irregular stands | A series of plots have been established since 2000 in stands being transformed to CCF. However only limited data have been published to date. | Poor-moderate | [44,70] |
| Impacts on timber quality | Modelling studies suggest limited impacts provided CCF stands are carefully thinned. Further validation required. | Moderate | [76] |
| Predicting costs and revenues | Field experiments and modelling suggests CCF can be competitive with standard management provided successful regeneration is obtained. Links to wind risk need exploration. | Moderate | [86–88] |
| Installation of field scale trials | At least three field scale trials of CCF installed in Sitka spruce dominated forests. | Good-moderate | [19,83] |
| Provision of other support mechanisms e.g., training, subsidies. | Training for foresters and operators available. Grant schemes available to private growers more suited to even-aged management than CCF | Moderate | [19] |
Although there is awareness of the possibility of fostering irregular stands on more exposed sites, it remains difficult to predict the potential risks involved. This will require a better understanding of how the wind loading on individual trees is altered by thinning, of how the loading changes subsequently as a result of adaptive growth, not to mention how these factors are affected by tree age and the social position of a tree within the stand. While much of this information can be obtained through field study, one aim should be to formalise the knowledge gained in an integrated modelling environment where the outputs of a wind risk model such as ForestGALES can be combined with those of a single tree growth model such as MOSES GB. Such integration would allow exploration of the type, timing and intensity of thinning to see when and how more stable structures might be developed in Sitka spruce stands on more exposed sites and what the distribution of risk might be to individual trees within the stand [
74]. The results from such simulations, and from the long-term growth and yield plots, could also be used to gain further understanding of the potential effects of transformation to CCF upon financial returns and profitability, including the calculation of annuities [
99]. This may require abandoning some traditional tenets of stand management based on average tree dimensions and stocking densities and moving towards a more spatially explicit form of management which recognises individual variation between and within stands [
100].
Another stage with “poor” evidence quality involves the desirability of diversifying the species composition of Sitka spruce stands managed by CCF. Introduction of selected complementary species is recommended as a means of increasing the functional diversity of planted forests by enhancing their capacity to respond to unforeseen changes [
31,
101]. A number of under-planting trials with a range of conifer species have been installed recently in Sitka spruce stands and should provide guidance within a few years (pers. comm. Gary Kerr and Victoria Stokes, Forest Research). Evidence from field sites in Britain suggests that natural regeneration within mature stands is almost exclusively of Sitka spruce with only occasional broadleaves or other species [
50,
85,
97]. Therefore any attempt to diversify the species composition of CCF stands will probably need to rely on under-planting to establish a future seed source. However, the fast and sustained height growth of Sitka spruce means that it is likely to suppress most broadleaves or other light demanding conifer species that may regenerate within gaps in a stand. This suggests that the species chosen as part of a diversification strategy should be intermediate or shade-tolerant broadleaves or conifers that can grow in the small gaps that develop in a CCF stand [
9,
102]. For instance, recent reviews have suggested that both western hemlock (
Tsuga heterophylla (Raf.) Sarg.)) and a number of
Abies species could be successfully under-planted in gaps in CCF stands [
103,
104]. The potential invasive effects of species such as western hemlock were thought to be limited, given the faster height growth of Sitka spruce [
103]. One other functional trait that could be usefully explored is rooting habit where admixture with a deeper rooting species might improve stability of stands of Sitka spruce. For example, results from tree-pulling studies showed that grand fir (
Abies grandis (Dougl. ex D. Don) Lindl.) had better anchorage on mineral soils than Sitka spruce [
105].
As yet, there appears to be no consistent view as to the silvicultural system(s) that might be best suited to supporting the implementation of CCF in Sitka spruce planted forests. Malcolm
et al. [
9] recommended an irregular shelterwood as the preferred option based on the view that Sitka spruce was a species of intermediate shade tolerance. However, the operational trial area at Clocaenog contain stands managed using uniform and group/strip as well as irregular shelterwoods [
70] and, as yet, there have been no major differences recorded in their effectiveness. A similar range of shelterwood systems have been reported from other trial areas [
19]. Ideally, information from these trials would be complemented by information on the dynamics of structural development observed in stands of varying ages, as has recently been reported from the species’ natural range in Alaska [
106]. The limited studies that have been carried out revealed a range of wind-created gap sizes in a number of planted Sitka spruce forests [
97,
98]. Larger gaps (>0.03 ha) appeared to be filling with established natural regeneration of Sitka spruce, but presence of other species was limited, possibly because of browsing by deer or because of the lack of a seed source [
97]. These findings suggest that, provided there is an adequate light environment for natural regeneration, Sitka spruce can be managed under CCF using a variety of silvicultural systems and gap sizes depending upon management objectives and other factors such as the incidence of wind risk. This potential flexibility of approach would accord with current thinking of embracing complexity in forest management [
100].



{kind=link}