Biomass Equations for Tropical Forest Tree Species in Mozambique

Abstract
:
1. Introduction
1.1. Study Background
1.2. Study Justification
1.3. Objectives
2. Materials and Methods
2.1. Study Area
2.2. General Characterization of Species in Stands

| Species type | Stems ha−1 | DBH, cm | Height, m | Basal area, m² ha−1 | ||
|---|---|---|---|---|---|---|
| Locality of Inhaminga, Sofala Province (Lat.18°58’ S.; Long. 34°10’ E, 100–200 m a.s.l) | ||||||
| Chanfuta | 8 ± 4 | 44.4 ± 13.7 | 11.1–79.6 | 16.3 ± 4.1 | 7.0–22.1 | 1.32 ± 0.87 |
| Jambire | 13 ± 12 | 31.2 ± 11.4 | 13.4–65.0 | 15.8 ± 4.4 | 6.0–28,9 | 1.13 ± 0.93 |
| Umbila | 5 ± 0 | 28.8 ± 16.3 | 14.3–46.5 | 10.8 ± 2.0 | 8.5–12.0 | 0.39 ± 0.40 |
| Others | 93 ± 55 | 23.0 ± 14.3 | 7.6–129.6 | 10.1 ± 3.7 | 3.0–25.1 | 5.35 ± 3.98 |
| Locality of Mavume, Inhambane Province (Lat. 23°28’ S.; Long. 34°33’ E, 100–200 m a.s.l) | ||||||
| Chanfuta | 15 ± 14 | 28.6 ± 9.0 | 17.2–42.5 | 10.9 ± 1.9 | 8.8–13.2 | 1.04 ± 1.23 |
| Umbila | 20 ± 14 | 22.8 ± 9.6 | 10.0–44.5 | 8.8 ± 2.9 | 3.8–16.2 | 0.98 ± 0.84 |
| Others | 69 ± 73 | 19.8 ± 8.1 | 10.0–49.0 | 6.6 ± 2.0 | 2.0–14.0 | 2.58 ± 2.06 |
| Locality of Tome, Inhambane Province (Lat. 22°32’ S.; Long. 34°12’ E, 100–200 m a.s.l) | ||||||
| Chanfuta | 17 ± 16 | 25.9 ± 9.6 | 13.5–48.0 | 11.5 ± 2.6 | 7.5–5.5 | 3.42 ± 1.22 |
| Others | 130 ± 83 | 17.4 ± 8.6 | 10.0–54.0 | 7.2 ± 2.8 | 3.0–15.0 | 3.83 ± 1.47 |
2.3. Sampling Design and Sampling Unit
- Biomass measurements in the field
- Biomass estimation
- Moisture content and basic density estimation
2.4. Biomass Measurements in the Field

2.5. Biomass Estimation
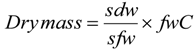
- Where:
- Sdw = dry weight of sub-sample (g)
- Sfw = fresh weight of sub-sample (g)
- fwC = fresh weight of component
2.6. Moisture Content Estimation
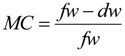
- Where:
- MC = moisture content
- fw = fresh weight (g)
- dw = oven-dry weight (g)
2.7. Biomass Equations
- Where:
- AGB = Kg d.w. tree−1
- D = DBH over bark (ob), mm
- Bd = Basic density, gcm−3
- H = Tree height, m
- β0, β1 and β2 are parameters
2.8. Basic Density Estimation
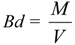
- Where:
- Bd: Basic density (g cm−3).
- M: Dry weight of stem or branch sample (g).
- V: Fresh volume of stem or branch sample (cm3).
2.9. Statistical Analyses
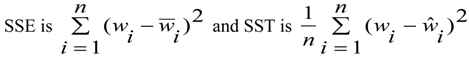
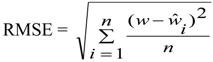
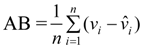
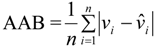
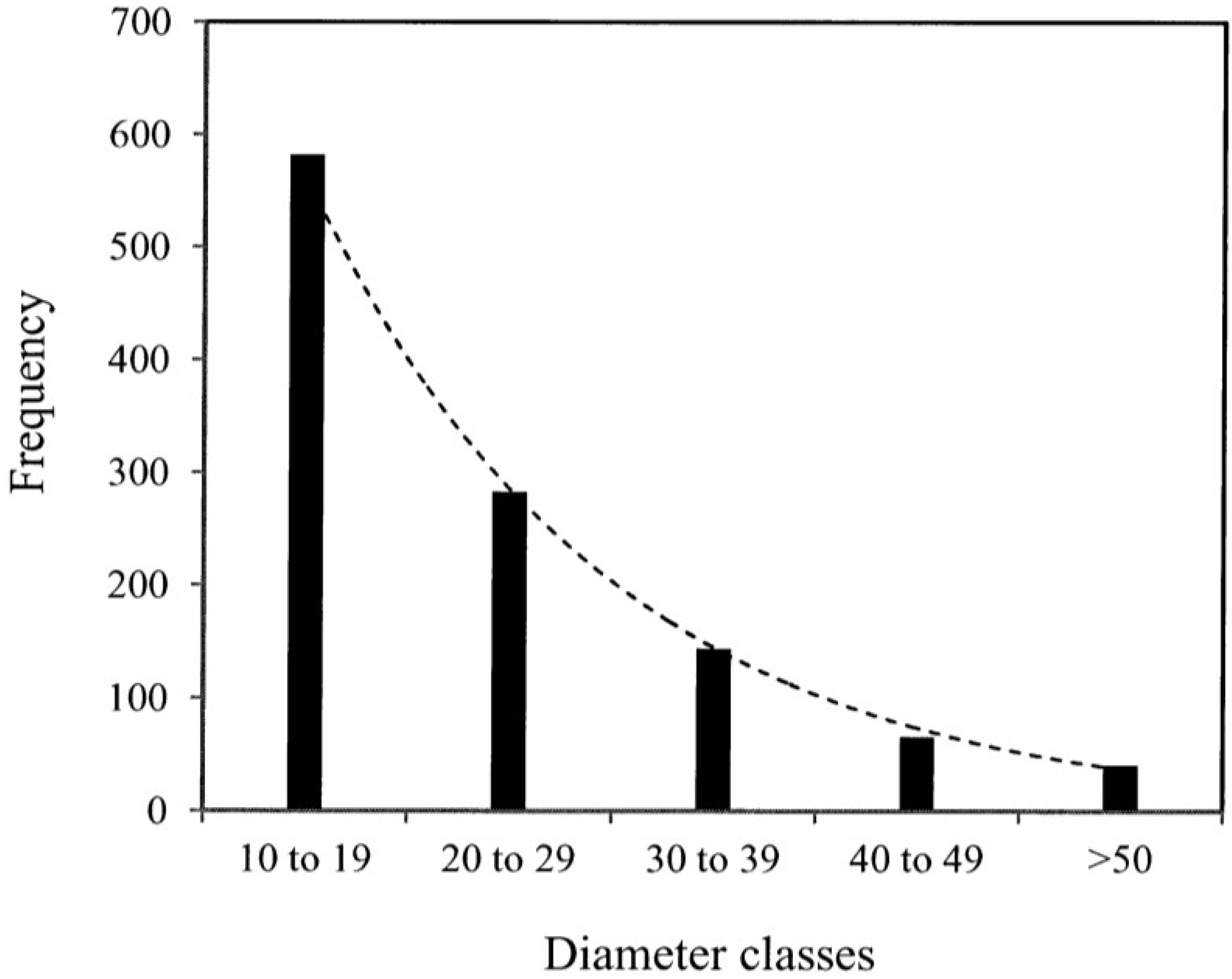
- Where:
- wi = Mean values of dry mass (w)
- vi,
![Forests 05 00535 i008]() , wi and
, wi and ![Forests 05 00535 i009]() = are observed and predicted values of dry mass (w)
= are observed and predicted values of dry mass (w)
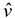
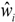
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
3. Results
3.1. Stand Characteristics
| Plot no. | Locality | Lat. S | Long. E | Stems ha−1 | DBH, cm | Height, m | Basal area, m2 ha−1 | Biomass, tons ha−1 | ||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean ± SD | Range | Mean ± SD | Range | Total | Total | |||||
| Chanfuta | ||||||||||
| 6 | Inhaminga | 18°15’ | 35°15’ | 10 | 60.5 ± 27.0 | 41.4–79.6 | 14.4 ± 6.9 | 9.5–19.2 | 3.16 | 21.7 |
| 7 | Inhaminga | 18°01’ | 35°17’ | 5 | 43.6 ± 0 | 43.6 | 18.6 ± 0 | 18.6 | 0.75 | 6.2 |
| 9 | Inhaminga | 17° 99’ | 35°19’ | 5 | 61.1 ± 0 | 61.1 | 20.6 ± 0 | 20.6 | 1.47 | 10.6 |
| 10 | Inhaminga | 18°74’ | 35°86’ | 10 | 60.0 ± 9.0 | 44.6–57.3 | 17.5 ± 2.1 | 16.0–18.9 | 2.07 | 15.9 |
| 15 | Inhaminga | 17°99’ | 35°15’ | 5 | 35.4 ± 0 | 35.4 | 15.0 ± 0 | 15 | 0.49 | 4.4 |
| 22 | Inhaminga | 18°08’ | 35°11’ | 10 | 22.3 ± 15.8 | 11.1–33.4 | 9.6 ± 3.7 | 7.0–12.3 | 0.49 | 4.8 |
| 39 | Inhaminga | 18°23’ | 35°13’ | 5 | 48.4 ± 0 | 48.4 | 15.8 ± 0 | 15.8 | 0.92 | 7.3 |
| 43 | Inhaminga | 18°09’ | 35°25’ | 5 | 38.5 ± 0 | 38.5 | 20.9 ± 0 | 20.9 | 0.58 | 5.1 |
| 202 | Inhaminga | 18°40’ | 35°14’ | 5 | 41.4 ± 0 | 41.4 | 19.6 ± 0 | 19.6 | 0.67 | 5.8 |
| 501 | Inhaminga | 17°99’ | 35°15’ | 5 | 51.0 ± 0 | 51.0 | 22.1 ± 0 | 22.1 | 1.02 | 7.8 |
| 502 | Inhaminga | 18°10’ | 35°08’ | 15 | 44.6 ± 1.0 | 43.6–45.5 | 17.0 ± 1.7 | 15.4–18.8 | 2.34 | 19.2 |
| 503 | Inhaminga | 18°17’ | 35°05’ | 10 | 39.6 ± 5.6 | 35.7–43.6 | 13.2 ± 1.6 | 12–14.3 | 1.25 | 10.7 |
| 2 | Mavume | 22°34’ | 34°11’ | 5 | 21.0 ± 0 | 21.0 | 9.0 ± 0 | 9.0 | 0.17 | 1.9 |
| 22 | Mavume | 23°27’ | 34°31’ | 25 | 30.1 ± 9.1 | 17.2–42.5 | 11.4 ± 1.8 | 8.8–13.2 | 1.91 | 17.7 |
| 3 | Tome | 22°34’ | 34°11’ | 5 | 21.0 ± 0 | 21.0 | 12.0 ± 0 | 12.0 | 0.17 | 1.9 |
| 4 | Tome | 22°34’ | 34°11’ | 5 | 18.5 ± 0 | 18.5 | 7.5 ± 0 | 7.5 | 0.13 | 1.6 |
| 5 | Tome | 22°34’ | 34°11’ | 10 | 20.8 ± 10.5 | 13.5–28.0 | 7.5 ± 0 | 7.5 | 0.38 | 4.0 |
| 6 | Tome | 22°35’ | 34°11’ | 5 | 21.0 ± 0 | 21.0 | 9.5 ± 0 | 9.5 | 0.17 | 1.9 |
| 7 | Tome | 22°35’ | 34°12’ | 15 | 29.0 ± 11.5 | 20.0–42.0 | 11.5 ± 1.8 | 9.6–13.0 | 1.09 | 10.2 |
| 8 | Tome | 22°35’ | 34°11’ | 45 | 21.6 ± 6.5 | 13.5–35.0 | 11 ± 1.9 | 8.0–14.5 | 1.79 | 18.9 |
| 9 | Tome | 22°33’ | 34°11’ | 35 | 34.1 ± 10.0 | 22.5–48.0 | 14.2 ± 1.1 | 13.0–15.5 | 3.42 | 30.2 |
| Jambire | ||||||||||
| 1 | Inhaminga | 18°50’ | 35°08’ | 5 | 25.2 ± 0 | 25.2 | 14.0 ± 0 | 14.0 | 0.25 | 3.1 |
| 4 | Inhaminga | 18°06’ | 35°15’ | 10 | 28.8 ± 2.0 | 27.4–30.3 | 16.3 ± 1.8 | 15.0–17.6 | 0.65 | 7.7 |
| 6 | Inhaminga | 18°09’ | 35°09’ | 15 | 41.2 ± 5.5 | 35.0–45.5 | 21.8 ± 6.2 | 18.1–28.9 | 2.02 | 19.4 |
| 8 | Inhaminga | 18°05’ | 35°09’ | 20 | 25.7 ± 6.8 | 16.6–33.1 | 15.5 ± 4.5 | 9.0–19.0 | 1.09 | 13.2 |
| 10 | Inhaminga | 18°07’ | 35°09’ | 5 | 35.0 ± 0 | 35.0 | 16.0 ± 0 | 16.0 | 0.48 | 5.1 |
| 11 | Inhaminga | 18°05’ | 35°16’ | 5 | 31.8 ± 0 | 31.8 | 18.8 ± 0 | 18.8 | 0.40 | 4.4 |
| 15 | Inhaminga | 17°99’ | 35°15’ | 5 | 38.2 ± 0 | 38.2 | 13 ± 0 | 13.0 | 0.57 | 5.8 |
| 27 | Inhaminga | 18°05’ | 35°06’ | 5 | 16.2 ± 0 | 16.2 | 7.0 ± 0 | 7.0 | 0.10 | 1.7 |
| 29 | Inhaminga | 18°09’ | 35°11’ | 55 | 26.9 ± 12.7 | 13.4–44.6 | 17.6 ± 2.8 | 11.0–20.2 | 3.75 | 40.6 |
| 34 | Inhaminga | 18°14’ | 35°08’ | 10 | 34.1 ± 1.4 | 33.1–35.0 | 13 ± 1.4 | 12.0–14.0 | 0.91 | 9.9 |
| 36 | Inhaminga | 18°20’ | 35°12’ | 5 | 32.5 ± 0 | 32.5 | 12.9 ± 0 | 12.9 | 0.41 | 4.7 |
| 37 | Inhaminga | 18°24’ | 35°13’ | 5 | 42.4 ± 0 | 42.4 | 16.3 ± 0 | 16.3 | 0.70 | 6.7 |
| 38 | Inhaminga | 18°21’ | 35°10’ | 10 | 21.5 ± 2 | 20.1–22.9 | 14.9 ± 1.3 | 13.9–158 | 0.36 | 5.0 |
| 39 | Inhaminga | 18°23’ | 35°13’ | 20 | 23.7 ± 9.9 | 13.7–33.4 | 10.1 ± 4.8 | 6.0–14.6 | 1.00 | 12.1 |
| 41 | Inhaminga | 18°13’ | 35°09’ | 5 | 65.0 ± 0 | 65.0 | 13 ± 0 | 13.0 | 1.66 | 12.5 |
| 42 | Inhaminga | 18°21’ | 35°11’ | 10 | 25.6 ± 6.5 | 21.0–30.3 | 18.2 ± 0.7 | 17.7–18.7 | 0.53 | 6.5 |
| 43 | Inhaminga | 18°09’ | 35°25’ | 5 | 55.1 ± 0 | 55.1 | 19.5 ± 0 | 19.5 | 1.19 | 9.9 |
| 101 | Inhaminga | 18°01’ | 35°08’ | 10 | 33.4 ± 27.0 | 14.3–52.5 | 15.5 ± 6.4 | 10.9–20.0 | 1.16 | 10.6 |
| 501 | Inhaminga | 17°59’ | 35°09’ | 30 | 34.3 ± 8.9 | 24.5–46.2 | 17.3 ± 2.6 | 14.0–21.3 | 2.92 | 30.2 |
| 502 | Inhaminga | 18°10’ | 35°08’ | 20 | 36.5 ± 5.1 | 31.2–43.3 | 16.9 ± 6.8 | 10.3–25.8 | 2.12 | 21.7 |
| Umbila | ||||||||||
| 1 | Mavume | 23°37’ | 34°29’ | 25 | 20.6 ± 7.3 | 11.0–28.5 | 10.3 ± 3.1 | 6.8–13.4 | 0.92 | 4.2 |
| 10 | Mavume | 23°37’ | 34°30’ | 15 | 24.3 ± 7.6 | 17.5–32.5 | 9.8 ± 3.3 | 7.2–13.5 | 0.74 | 3.5 |
| 11 | Mavume | 23°37’ | 34°30’ | 40 | 24.6 ± 10.4 | 14.5–42.0 | 11.2 ± 2.5 | 8.5–16.2 | 2.20 | 10.6 |
| 12 | Mavume | 23°37’ | 34°30’ | 35 | 28.0 ± 13.2 | 10.0–44.5 | 8 ± 3.1 | 3.8–11.0 | 2.56 | 12.6 |
| 13 | Mavume | 23°37’ | 34°30’ | 10 | 22.8 ± 18.0 | 10.0–35.5 | 6.9 ± 1.6 | 5.8–8.0 | 0.53 | 2.6 |
| 14 | Mavume | 23°37’ | 34°37’ | 10 | 22.5 ± 7.8 | 17.0–28.0 | 8.5 ± 2.8 | 6.5–10.4 | 0.42 | 2.0 |
| 15 | Mavume | 23°37’ | 34°30’ | 10 | 25.0 ± 12.7 | 16.0–34.0 | 10.5 ± 5.7 | 6.5–14.5 | 0.55 | 2.7 |
| 16 | Mavume | 23°37’ | 34°30’ | 10 | 27.5 ± 3.5 | 25.0–30.0 | 7.9 ± 4.7 | 4.6–11.2 | 0.60 | 2.8 |
| 17 | Mavume | 23°37’ | 34°30’ | 50 | 21.6 ± 9.9 | 13.5–42.0 | 8.2 ± 2.3 | 4.5–11.6 | 2.18 | 10.4 |
| 18 | Mavume | 23°37’ | 34°30’ | 20 | 12.6 ± 1.3 | 11.0–14.0 | 6.3 ± 0.6 | 5.5–7.0 | 0.25 | 1.0 |
| 20 | Mavume | 23°37’ | 34°30’ | 5 | 16.0 ± 0.0 | 16.0 | 6.5 ± 0 | 6.5 | 0.10 | 0.4 |
| 21 | Mavume | 23°37’ | 35°30’ | 15 | 22.8 ± 5.5 | 17.0–28.0 | 7.7 ± 1.2 | 6.5–8.5 | 0.64 | 3.0 |
| 35 | Inhaminga | 18°25’ | 35°13’ | 5 | 25.5 ± 0.0 | 25.5 | 12.0 ± 0 | 12.0 | 0.25 | 1.2 |
| 36 | Inhaminga | 18°20’ | 35°12’ | 5 | 14.3 ± 0.0 | 14.3 | 8.5 ± 0 | 8.5 | 0.08 | 0.3 |
| 40 | Inhaminga | 18°14’ | 35°07’ | 5 | 46.5 ± 0.0 | 46.5 | 12.0 ± 0 | 12.0 | 0.85 | 4.4 |
3.2. Sample Tree Characterization
| Fresh weight (kg tree−1) | Percentage of total fresh weight. % | |||||||
|---|---|---|---|---|---|---|---|---|
| DBH,cm | Total | Stem | Branches | Leaves | Stem | Branches | Leaves | |
| Chanfuta (n = 24) | ||||||||
| Mean ± SD | 33.8 ± 12.6 | 1563 ± 928 | 970 ± 838 | 552 ± 339 | 42 ± 43 | 51 ± 26 | 45 ± 25 | 4 ± 3 |
| Range | 13.5–61.1 | 221–3018 | 28–2882 | 111–1187 | 0–47 | 6–96 | 4–90 | 0–10 |
| Jambire (n = 15) | ||||||||
| Mean ± SD | 34.8 ± 8.2 | 1840 ± 651 | 1389 ± 473 | 423 ± 250 | 28 ± 21 | 76 ± 9 | 22 ± 9 | 2 ± 1 |
| Range | 21.0–52.2 | 750–3504 | 536–2489 | 94–981 | 5–78 | 64–94 | 5–36 | 0–4 |
| Umbila (n = 19) | ||||||||
| Mean ± SD | 27.0 ± 9.5 | 630 ± 400 | 276 ± 231 | 339 ± 221 | 14 ± 10 | 42 ± 18 | 54 ± 17 | 3 ± 4 |
| Range | 14.0–46.5 | 140–1785 | 34–946 | 57–820 | 3–38 | 14–87 | 12–84 | 1–18 |
3.3. Biomass Equation for Individual Trees
| Dry weight (kg tree−1) | Percentage of total dry weight. % | Percentage dry weightby fresh weight | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| DBH, cm | Total | Stem | Branches | Leaves | Stem | Branches | Leaves | Stem | Branches | Leaves | |
| Chanfuta (n = 24) | |||||||||||
| Mean ± SD | 33.8 ± 12.6 | 864 ± 548 | 569 ± 524 | 280 ± 187 | 15 ± 19 | 54 ± 27 | 43 ± 26 | 3 ± 4 | 56 ± 7 | 50 ± 9 | 29 ± 19 |
| Range | 13.5–61.1 | 107–2018 | 14–1956 | 57–667 | 0–77 | 6–97 | 3–92 | 0–16 | 36–75 | 29–65 | 0–70 |
| Jambire (n = 15) | |||||||||||
| Mean ± SD | 34.8 ± 8.2 | 1016 ± 438 | 782 ± 341 | 222 ± 161 | 11 ± 7 | 77 ± 11 | 22 ± 10 | 1 ± 1 | 56±13 | 51 ± 8 | 43 ± 14 |
| Range | 21.0–52.2 | 411–2086 | 296–1412 | 42–659 | 1–27 | 53–96 | 3–44 | 0–3 | 21–77 | 35–67 | 18–2 |
| Umbila (n = 19) | |||||||||||
| Mean ± SD | 27.0 ± 9.5 | 321 ± 240 | 152 ± 140 | 162 ± 133 | 7 ± 5 | 46 ± 22 | 51 ± 21 | 3 ± 4 | 52 ± 5 | 46 ± 13 | 50 ± 4 |
| Range | 14.0–46.5 | 52–1121 | 16–96 | 16–516 | 1–17 | 13–92 | 6–84 | 1–17 | 45–66 | 5–63 | 42–60 |
| Components | Parameter estimates | AB | AAB | R2 | RMSE |
|---|---|---|---|---|---|
| Chanfuta | |||||
| Total | 3.1256 × D1.5833 | −10.6 | 159.8 | 0.97 | 194.37 |
| Stem | 0.4369 × D2.0033 | −20.0 | 171.6 | 0.91 | 227.90 |
| Branches | 22.7577 × D0.7335 | −0.1 | 15.0 | 0.79 | 168.19 |
| Leaves | 19.9625 × D−0.0836 | 2.1 | 13.2 | 0.40 | 19.14 |
| Jambire | |||||
| Total | 5.7332 × D1.4567 | 49.5 | 250.0 | 0.95 | 256.83 |
| Stem | 4.8782 × D1.4266 | 43.5 | 217.6 | 0.94 | 220.25 |
| Branches | 0.3587 × D1.8091 | 10.3 | 90.7 | 0.78 | 142.48 |
| Leaves | 77.0114 × D−0.5511 | −0.7 | 6.3 | 0.72 | 4.09 |
| Umbila | |||||
| Total | 0.2201 × D2.1574 | 9.6 | 103.8 | 0.89 | 140.69 |
| Stem | 0.0083 × D2.8923 | −1.6 | 23.1 | 0.95 | 51.43 |
| Branches | 2.3596 × D1.2690 | 3.7 | 96.0 | 0.70 | 120.68 |
| Leaves | 4.0400 × D0.1680 | 0.0 | 3.3 | 0.71 | 4.71 |
 ), stem (
), stem (  ) branches (---- ▲) and leaves (– · – ∆) for samples of Chanfuta (a), Jambire (b), and Umbila (c).
), stem ( ) branches (---- ▲) and leaves (– · – ∆) for samples of Chanfuta (a), Jambire (b), and Umbila (c).
) branches (---- ▲) and leaves (– · – ∆) for samples of Chanfuta (a), Jambire (b), and Umbila (c).
), stem ( ) branches (---- ▲) and leaves (– · – ∆) for samples of Chanfuta (a), Jambire (b), and Umbila (c). 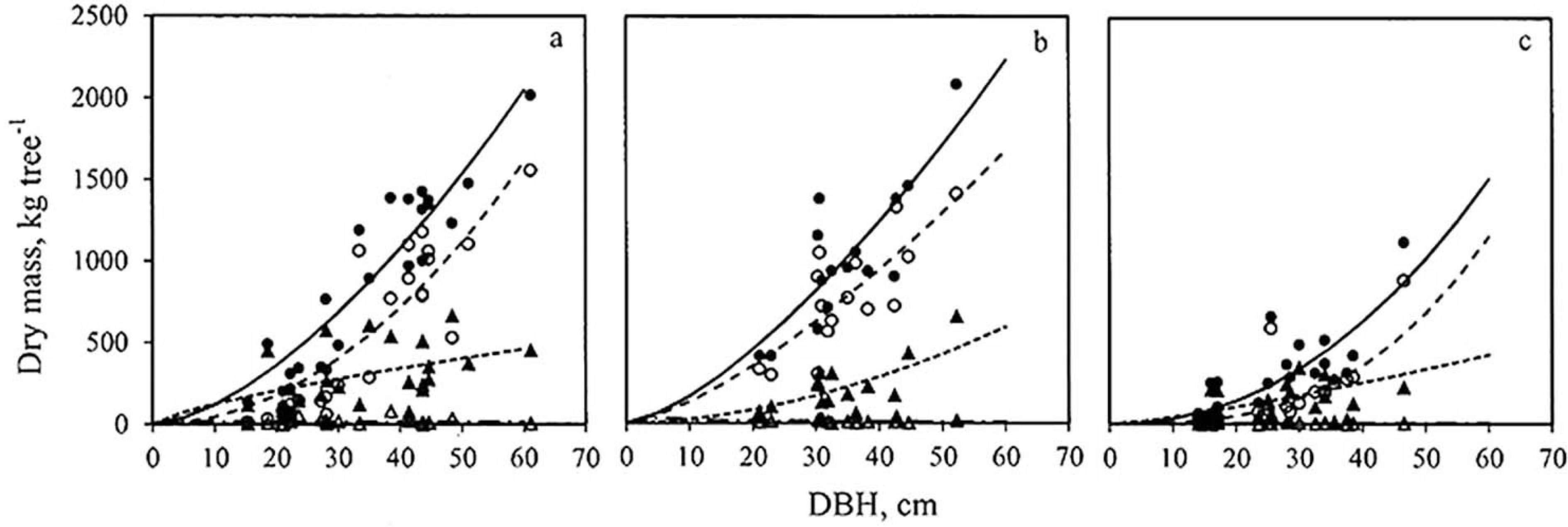
3.4. Biomass Estimates at Plot and Stand Level
3.5. Basic Density

4. Discussion
4.1. Biomass Equations
4.2. Biomass Distribution
4.3. Basic Density
5. Conclusions
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Global Forest Resources Assessment; 2010 Main Report; FAO Forestry Paper 163; FAO: Rome, Italy, 2012; p. 60.
- Marzoli, A. National Forest Inventory. In Integrated Forest Assessment of Mozambique (AIFM), Maputo; National Directorate of Land and Forest (DNTF)-Ministry of Agriculture: Maputo, Mozambique, 2007; p. 92. [Google Scholar]
- Brouwer, R.; Falcão, M.P. Wood fuel consumption in Maputo, Mozambique. Biomass Bioenergy 2004, 27, 233–245. [Google Scholar] [CrossRef]
- National Directorate of Forest and Wildlife (DNFFB). Regulation for the forestry and wildlife policy. Govern. Bull. 2002, 1, 25. [Google Scholar]
- Fath, H. Commercial Timber harvesting in the natural forest of Mozambique. In FAO Forest Harvesting: Case Study; Food and Agricultural Organization of the United Nations: Rome, Italy, 2001; p. 50. [Google Scholar]
- Annual Statistic Report; National Directorate of Land and Forestry (DNTF): Maputo, Mozambique, 2010; p. 21.
- Hoogwijk, M.; Faaij, A.; van den Broek, R.; Berndes, G.; Gielen, D.; Turkenberg, W. Exploration of the ranges of the global potential of biomass for energy. Biomass Bioenergy 2003, 25, 119–133. [Google Scholar] [CrossRef]
- Richardson, J.; Bjorheden, R.; Hakkila, P.; Lowe, A.T.; Smith, C.T. Bioenergy from Sustainable Forestry: Guiding Principles and Practice-Kluwer; Academic Publishers: Dordrecht, The Netherlands, 2002; p. 344. [Google Scholar]
- Wyk, B.V.; Wyk, P.V. Field Guide to Trees of Southern Africa; Struik Publisher: Cape Town, South Africa, 1997; p. 525. [Google Scholar]
- Danida Forest Seed Center (DFSC). Afzelia Quanzensis Welw. Seed Leafl. 2000, 31, 2. [Google Scholar]
- Gomes e Sousa, A. Dendrology of Mozambique: A General Study. In Memories; Institute of Agrarian Research of Mozambique: Maputo, Mozambique, 1966; pp. 1–822. [Google Scholar]
- Hines, D.A.; Eckman, K. Indigenous Multipurpose Trees of Tanzania: Uses and Economic Benefits For People; Cultural Survival Canada and Development Services Foundation of Tanzania & Tree Roots Canada: Ottawa, Canada, 1993. [Google Scholar]
- Bunster, J.H. 52 Woods of Mozambique “52 Madeiras de Moçambique”: Technologic Cathalogue; UEM/FAEF/Forestry Department: Maputo, Mozambique, 1995; pp. 1–80. [Google Scholar]
- Edinburgh Centre for Carbon Management (ECCM). Technical Specification; Mozambique Technical Specification Boundary Planting: Edinburgh, UK, 2005; p. 8. [Google Scholar]
- Woodcrafters Association of Durban (WAD). Wood of the month. Newsl. Woodcrafters Assoc. Durb. Issue 2005, 15, 9. [Google Scholar]
- Lemmens, R.H.M.J. Millettia stuhlmannii Taub. In Timbers/Bois d’œuvre 1; [CD-Rom]; Louppe, D., Oteng-Amoako, A.A., Brink, M., Eds.; Plant Resource of Tropical Africa (PROTA): Wageningen, The Netherlands, 2008. [Google Scholar]
- Therrell, M.D.; Stahle, D.W.; Mukelabai, M.M.; Shugart, H.H. Age and radial growth dynamics of Pterocarpus angolensis in Southern Africa. For. Ecol. Manag. 2007, 244, 24–31. [Google Scholar] [CrossRef]
- Fichtler, E.; Trouet, V.; Beeckman, H.; Coppin, P.; Worbes, M. Climatic Signals in Tree Rings of Burkea Africana and Pterocarpus Angolensis from semiarid forests in Namibia. Trees 2004, 18, 442–451. [Google Scholar]
- Vermeulen, W.J. A Monograph on Pterocarpus angolensis; SARCCUS Standing Committee for Forestry: Pretoria, South Africa, 1990; p. 133. [Google Scholar]
- Van Daalen, J.C.; Vogel, J.C.; Malan, F.S.; Fuls, A. Dating of Pterocarpus angolensis trees. S. Afr. For. J. 1992, 162, 1–7. [Google Scholar]
- Shackleton, C.M. Growth patterns of Pterocarpus angolensis in Savannahs of the SOUTH African Lowveld. For. Ecol. Manag. 2002, 166, 85–97. [Google Scholar] [CrossRef]
- Mozambique Biomass National Strategy; Final Report; Ministry of Energy: Maputo, Mozambique, 2012; p. 171.
- Sitoe, A.; Mirira, R.; Tchaúque, F. Assessment of Consumption levels of biomass energy in Tete, Nampula, Zambézia, Sofala, Gaza and Maputo Provinces. Ministry of Energy/Faculty of Agronomy and Forestry Engineering; Ministry of Energy: Maputo, Mozambique, 2007; p. 50.
- Cuvilas, C.A.; Llate, I.; Jirjis, R.; Terziev, N. The characterization of wood species from Mozambique as a fuel. Energy Sources. Part A 2014, 36, 851–857. [Google Scholar] [CrossRef]
- Gillespie, A.J.; Brown, S.; Lugo, A.E. Tropical forest biomass estimation from truncated stand tables. For. Ecol. Manag. 1992, 48, 69–87. [Google Scholar] [CrossRef]
- Návar-Cháidez, J.J. Allometric equations and expansion factors for tropical dry forest trees of eastern Sinalo, Mexico. Trop. Subtrop. Agroecossystems 2009, 10, 45–52. [Google Scholar]
- Brown, S.; Gillespie, A.J.; Lugo, A.E. Biomass estimation methods for tropical forests with applications to forest inventory data. For. Sci. 1989, 35, 881–902. [Google Scholar]
- Chambers, J.Q.; dos Santos, J.; Ribeiro, R.J.; Higuchi, N. Tree damage, allometric relationships, and above-ground net primary production in central Amazon forest. For. Ecol. Manag. 2001, 152, 73–84. [Google Scholar] [CrossRef]
- Chavé, J.; Andalo, C.; Brown, S.; Cairns, M.A.; Chambers, J.Q.; Eamus, D.; Fölster, H.; Fromard, F.; Higuchi, N.; Kira, T.; et al. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 2005, 145, 87–99. [Google Scholar] [CrossRef]
- Návar, J. Measurement and assessment methods of forest aboveground biomass: a literature review and challenges ahead. In Biomass; Momba, M., Bux, F., Eds.; Sciyo: Rijeka, Croatia, 2010; pp. 27–64. [Google Scholar]
- Frost, P. The ecology of miombo woodlands. In The Miombo in Transition: Woodlands and Welfare in Africa; Campbell, B.M., Ed.; Center for International Forestry Research: Bogor, Indonesia, 1996; pp. 11–55. [Google Scholar]
- Lawton, R.M. Natural resources of miombo woodland and recent changes in agricultural and land-use practices. For. Ecol. Manag. 1982, 4, 287–297. [Google Scholar] [CrossRef]
- Chidumayo, E.N. Above-ground woody biomass structure and productivity in Zambezian woodlands. For. Ecol. Manag. 1990, 36, 33–46. [Google Scholar] [CrossRef]
- Mozambique National Institute of Statistics. Available online: http//www.ine.gov.mz/pt/Data analysis (assessed on 15 May 2013).
- Brown, S.; Lugo, A.E. Aboveground biomass estimates for tropical moist forests of the Brazilian Amazon. Interciencia 1992, 1, 8–18. [Google Scholar]
- Parresol, B.R. Assessing tree and stand biomass: A review with examples and critical comparisons. For. Sci. 1999, 45, 575–593. [Google Scholar]
- Pearson, T.; Walker, S.; Brown, S. Sourcebook for Land Use, Land-Use Change and Forestry Project; BioCarbon Fund WinRock International: Bryssels, Belgium, 2005. [Google Scholar]
- Johansson, T.; Karacic, A. Increment and biomass in hybrid poplar and some practical implications. Biomass Bioenergy 2011, 35, 1925–1934. [Google Scholar]
- Grundy, I.M. Wood biomass estimation in dry miombo in Zimbabwe. For. Eco. Manag. 1995, 72, 109–117. [Google Scholar] [CrossRef]
- Kittredge, J. Estimation of the amount of foliage of trees and stands. J. For. 1944, 42, 905–912. [Google Scholar]
- Payandeh, B. Choosing regression models for biomass prediction models. For. Chron. 1981, 57, 229–232. [Google Scholar] [CrossRef]
- Satoo, T.; Madgewick, H.A.I. Forest Biomass; Martinus Nijhoff/DR W. Junk. Publishers: London, UK, 1985; p. 23. [Google Scholar]
- Bolstad, P.V.; Gower, S.T. Estimation of leaf area index in fourteen southern wisconsin forests stands using a portable radiometer. Tree Physiol. 1990, 7, 115–124. [Google Scholar] [CrossRef]
- Johansson, T. Biomass equations for determining fractions of pendula and pubescent birches growing on abandoned farmland and some practical implications. Biomass Bioenergy 1999, 16, 223–238. [Google Scholar] [CrossRef]
- Návar-Cháidez, J.; Rodríguez-Flores, F.J.; Domínguez-Calleros, P.A. Allometric equations for tropical trees: Application of forest inventory of Sianloa, México. Agron. Mesoam. 2013, 24, 347–356. [Google Scholar]
- Návar, J. Allometric equations for tree species and carbon stocks for forests of north-western Mexico. For. Ecol. Manag. 2009, 257, 427–434. [Google Scholar] [CrossRef]
- Andersson, E.; Tuimala, A. Definitioner, mm, vid vedundersökningar (Definitions for Wood Technology); (in Swedish); The Nordic Joint Group in Wood Technology Research Note, No. 4. Joint Group for Wood Technology and Producers: Ås, Norway, 1980; pp. 53–66. [Google Scholar]
- SAS, Version 9.1; SAS Institute Inc.: Cary, NC, USA, 2006.
- Zar, J.H. Biostatistical Analysis; Prentice-Hall: Englewood Cliffs, NJ, USA, 1999. [Google Scholar]
- Parresol, B.R.; Hotvedt, J.E.; Cao, Q.V. A Volume and taper prediction system for bald cypress. Can. J. For. Res. Res. 1987, 17, 250–259. [Google Scholar] [CrossRef]
- Verwijst, T.; Telenius, B. Biomass estimation procedures in short rotation forestry. For. Ecol. Manag. 1999, 121, 137–146. [Google Scholar] [CrossRef]
- Karacic, A.; Verwijst, T.; Weih, M. Above-ground woody biomass production of short-rotation Populus plantations on agricultural land in Sweden. Scand. J. For. Res. 2003, 18, 427–437. [Google Scholar] [CrossRef]
- Zewdie, M.; Olsson, M.; Verwijst, T. Above-ground biomass production and allometric relations of Eucalyptus Globulus Labill. coppice plantations along a consequence in the central highlands Of Ethiopia. Biomass Bioenergy 2009, 33, 421–428. [Google Scholar] [CrossRef]
- Tomo, F.A. Estimates of Carbon Stock in Miombo Forest in Gondola District; Faculty of Agronomy and Forestry Engineering: Maputo, Mozambique, 2012; p. 63. [Google Scholar]
- Chave, J.; Andalo, C.; Brown, S.; Cairns, M.A.; Chambers, J.Q.; Eamus, D.; Fölster, H.; Fromard, F.; Higuchi, N.; Kira, T.; et al. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 2005, 145, 87–99. [Google Scholar] [CrossRef]
- Litton, C.M.; Kauffman, J.B. Allometric models for predicting aboveground biomass in two widespread woody plants in Hawaii. Biotropica 2008, 40, 313–320. [Google Scholar] [CrossRef]
- Henry, M.; Picard, N.; Trotta, C.; Manlay, R.J.; Valentini,, R.; Bernoux, M; Saint-André, L. Estimating tree biomass of Sub-Saharan African forests: A review of available allometric equations. Silva Fenni. 2011, 45, 477–569. [Google Scholar]
- Ketterings, Q.M.; Coe, R.; van Noordwijk, M.; Ambagau, Y.; Palm, C.A. Reducing uncertainty in the use of allometric biomass equations for predicting above-ground tree biomass in mixed secondary forests. For. Eco. Manag. 2001, 146, 199–209. [Google Scholar] [CrossRef]
- Tchaúque, F.D.D.L.J. Assessment of Aboveground Biomass in Beira Corridor; Faculty of Agronomy and Forestry Engineering: Maputo, Moçambique, 2004; p. 49. [Google Scholar]
- Geldenhuys, C.J.; Golding, J.S. Resource use activities, conservation and management of natural resources of African Savannas. In Savannas: Challenges and Strategies for Equilibrium between Society and Agribusiness and Natural Resources; Faleiro, F.G., Lopez, A., Neto, D., Eds.; Embrapa Cerrados: Brasilia, Brazil, 2008; pp. 225–260. [Google Scholar]
- Ribeiro, N.S.; Shugart, H.H.; Washington-Allen, R. The effects of fire and elephants on species composition and structure of the Niassa Reserve, northern Mozambique. For. Ecol. Manag. 2008, 255, 1626–1636. [Google Scholar] [CrossRef]
- Henry, M.; Besnard, A.; Asante, W.A.; Eshun, J.; Adu-Bredu, S.; Valentini, R. Woody density, phytomass variations within and among trees, and allometric equations in a tropical rainforest of Africa. For. Ecol. Manag. 2010, 260, 1375–1386. [Google Scholar] [CrossRef]
- Chamshama, S.A.O.; Mugasha, A.G.; Zahabu, E. Stand biomass and volume estimation for miombo woodlands at Kitulangalo, Morogoro, Tanzania. South. Afr. For. J. 2004, 200, 59–70. [Google Scholar]
- Abbot, P.; Lowero, J.; Werren, M. Models for the estimation of single tree volume in four miombo woodlands types. For. Ecol. Manag. 1997, 97, 25–37. [Google Scholar] [CrossRef]
- Nelson, B.W.; Mesquita, R.; Pereira, J.L.G.; de Souza, S.G.A.; Batisad, G.T.; Coutoe, L.B. Allometric regressions for improved estimate of secondary forest biomass in the central Amazonia, Brazil. For. Ecol. Manag. 1999, 117, 149–167. [Google Scholar] [CrossRef]
- Segura, M.; Kanninen, M. Allometric models for tree volume and total aboveground biomass in a tropical humid forest in Costa Rica. Biotropica 2005, 37, 2–8. [Google Scholar] [CrossRef]
- Murali, K.S.; Bhat, D.M.; Ravindranath, N.H. Biomass estimation equation for tropical deciduous and evergreen forests. Int. J. Agric. Resour. Gov. Ecol. 2005, 1, 81–92. [Google Scholar]
- Chidumayo, E.; Timberlake, J.; Sawadogo, L. Distribution and characteristics of African dry forests and woodlands. In The Dry Forests and Woodlands of Africa: Managing for products and services; Earthscan: London, UK, 2010; pp. 11–41. [Google Scholar]
- Sitoe, A.; Salomão, A.; Wertz-Kanounnikoff, S. The Context of REDD+ in Mozambique: Causes, Actors and Institutions; CIFOR Occasional Paper 76; CIFOR: Bogor, Indonesia, 2012. [Google Scholar]
- Ali, A.M; Chirkova, J.; Terziev, N.; Elowson, T. Physical properties of two tropical species from Mozambique. Wood Mat. Sci. Engin. 2010, 5, 151–161. [Google Scholar]
- Givnish, T.J. Adaptation to sun and shade, a whole plant perspective. J. Plant Phys. 1988, 15, 63–92. [Google Scholar]
- Poorter, L.; Lianes, E.; Moreno-de las Heras, M.; Zavala, M.A. Architecture of iberian canopy tree species in relation to wood density, shade tolerance and climate. Plant Ecol. 2012, 213, 707–722. [Google Scholar] [CrossRef]
- Enquist, B.J.; West, G.B.; Charnov, E.L.; Brown, J.H. Allometric scaling of production and life-history variation in vascular plants. Nature 1999, 401, 907–911. [Google Scholar] [CrossRef]
- Whitmore, T.C. Potential impact of climate change on tropical forest seedling and forest regeneration. Clim. Chang. 1998, 39, 429–438. [Google Scholar] [CrossRef]
- Nock, C.A.; Geihofer, D.; Rabner, M.; Baker, P.J.; Bunyavejchewin, S.; Hietz, P. Wood density and its radial variation in six canopy tree species differing in shade-tolerance in western Thailand. Ann. Bot. 2009, 104, 297–306. [Google Scholar] [CrossRef]
- Chidumayo, E.; Gambiza, J.; Grundy, I. Managing miombo woodlands. In The Miombo In Transition: Woodlands and Welfare in Africa; Campbell, B.M., Ed.; Center for International Forestry Research: Bogor, Indonesia, 1996; pp. 175–193. [Google Scholar]
- Silva-Arredondo, F.M.; Návar-Cháidez, J.J. Estimating bole wood specific gravity in trees of temperate forest communities of northern Durango, Mexico. Madera y Bosques 2012, 18, 77–88. [Google Scholar]
- Silva-Arredondo, F.M.; Návar-Cháidez, J.J. Biomass expansion factors in temperate forest communities of north Durango, Mexico. Rev. Mex. Cien. For. 2011, 1, 55–62. [Google Scholar]
- Higuchi, N.; Dos Santos, J.; Ribeiro, R.F.; Minette, L.; Biot, Y. Aboveground biomass of tropical humid forest vegetation in Terra Firme, Amazonia. Acta Amazonica 1998, 2, 153–166. [Google Scholar]
- Nogueira, E.M.; Nelson, B.W.; Fearnside, P.M. Wood density in dense forest in the central Amazonas, Brazil. For. Ecol. Manag. 2005, 208, 261–286. [Google Scholar] [CrossRef]
- De Miranda, M.C.; Castelo, P.A.R.; De Miranda, D.L.C.; Rondon, E.V. Physical and mechanical properties of the wood from Parkia gigantocarpa Ducke. Braz. J. Wood Sci. 2012, 3, 55–65. [Google Scholar]
- Omonte, M.; Valenzuela, L. Radial and longitudinal basic density variation in 16 years old Eucalyptus regnans trees. Maderas. Ciencia y Tecnología 2011, 13, 211–224. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Mate, R.; Johansson, T.; Sitoe, A. Biomass Equations for Tropical Forest Tree Species in Mozambique. Forests 2014, 5, 535-556. https://doi.org/10.3390/f5030535
Mate R, Johansson T, Sitoe A. Biomass Equations for Tropical Forest Tree Species in Mozambique. Forests. 2014; 5(3):535-556. https://doi.org/10.3390/f5030535
Chicago/Turabian StyleMate, Rosta, Tord Johansson, and Almeida Sitoe. 2014. "Biomass Equations for Tropical Forest Tree Species in Mozambique" Forests 5, no. 3: 535-556. https://doi.org/10.3390/f5030535