Spatial Characteristics of Edible Wild Fern Harvesting in Mountainous Villages in Northeastern Japan Using GPS Tracks

Abstract
:
1. Introduction
2. Data and Methods
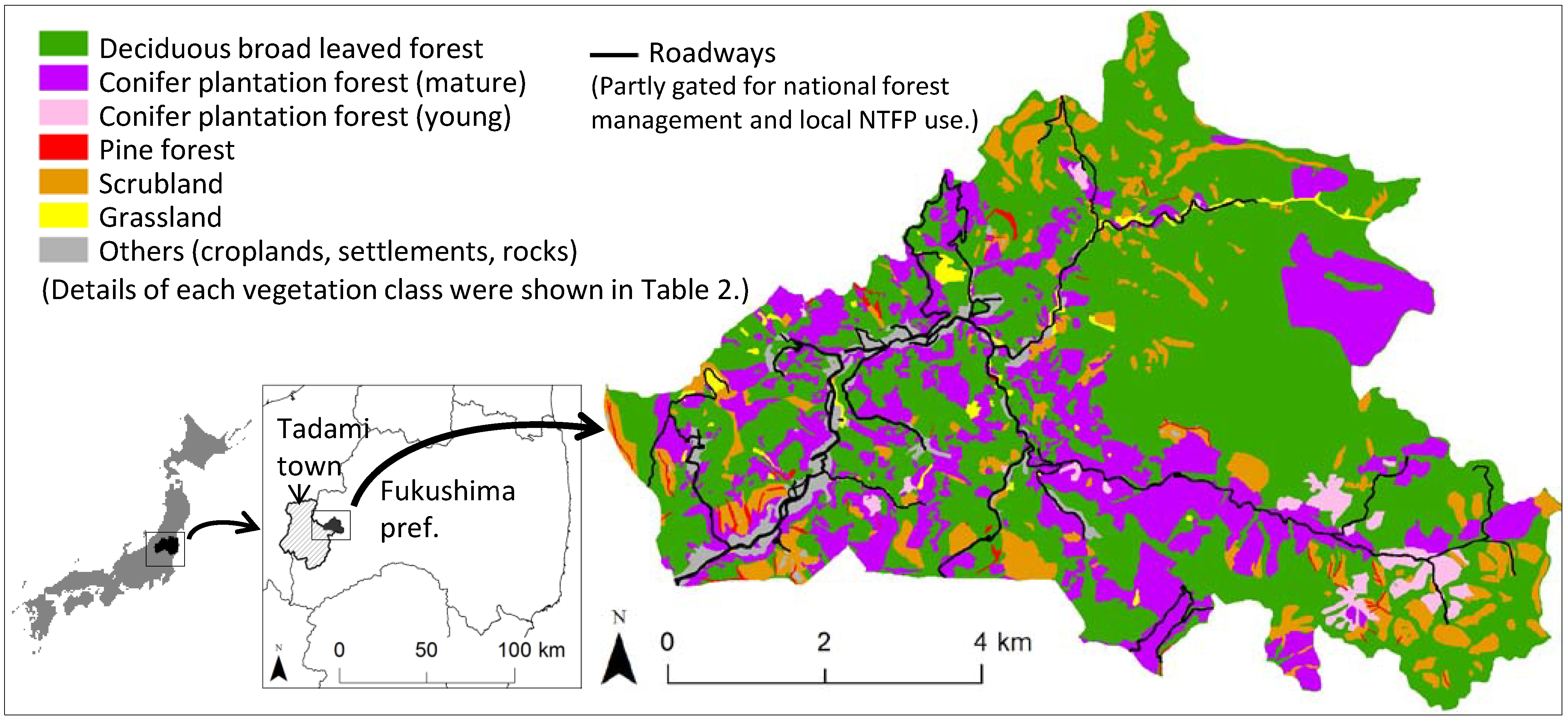
2.1. Study Area
2.2. Target Fern Species
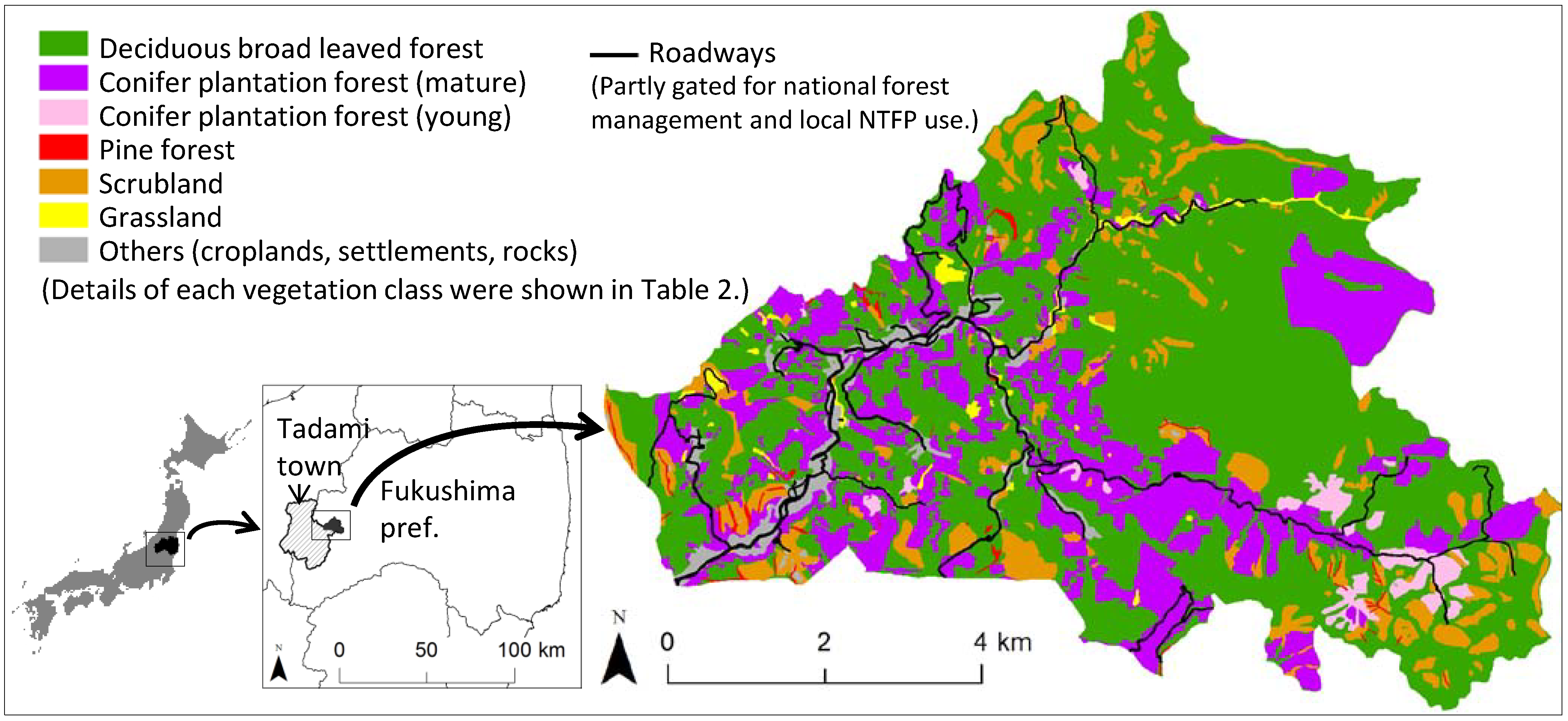
2.3. Harvesting/Non-Harvesting Sites
2.4. GIS Data Set

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Explanatory variables | Description | Min | Max | Mean | Std. Dev. | Unit |
|---|---|---|---|---|---|---|
| Vegetation 1 | Vegetation classes, i.e., deciduous broadleaved forest, young and mature conifer plantations, pine forest, scrubland and grassland. | (binary) | - | |||
| Terrain features derived from a 20-m digital elevation model (DEM) | ||||||
| Gradient 2 | The maximum rate of elevation change between each cell and its neighbors. | 0.06 | 54.68 | 21.41 | 8.63 | degree |
| Aspect 3 | Cosine transformation of aspect (clockwise from north) to 16 directions: cos[(aspect + n × 22.5)/180 × π], n = 0, 1, … 7. | −1 | 1 | −0.050 | 0.714 | - |
| Relative slope position 4 | The relative height from the nearest drainage to each grid point divided by the vertical size of a sideslope. | 0 | 1 | 0.63 | 0.33 | - |
| Potential solar radiation 2 | Potential solar radiation with monthly interval. | 45,0348 | 156,7600 | 126,3764 | 17,4382 | WH/m2 |
| Topographic wetness index (TWI) 5 | TWI = ln(A/tanB), where A is specific catchment area while B is slope gradient. | 9.07 | 27.31 | 11.56 | 1.90 | - |
| Proximity to roads 1 | Path distance to the nearest roads weighted by the slope gradient. | 0 | 2538.6 | 652.3 | 542.4 | m |
| No. | Vegetation class 1 | Subclass | Dominant tree species 2 | Area (%) |
|---|---|---|---|---|
| 1 | Deciduous broadleaved forest | Japanese beech forest | Fagus crenata | 58.4 |
| Coppice forest | Quercus crispula, Quercus serrata | |||
| Riparian forest | Pterocarya rhoifolia, Salix dolichostyla | |||
| 2 | Conifer plantation (mature) | Japanese cedar/larch plantation (age >= 20 years) with closed canopies | Cryptomeria japonica, Larix kaempferi | 24.5 |
| 3 | Conifer plantation (young) | Japanese cedar/larch plantation (age < 20 years) with dominant canopy openings | Cryptomeria japonica, Larix kaempferi | 2.2 |
| 4 | Pine forest | Japanese white/red pine forest mainly located on sharp ridges | Pinus parviflora var. pentaphylla, Pinus densiflora | 0.7 |
| 5 | Scrubland | Snow avalanche scrubland | Quercus crispula, Weigela hortensis, Clethra barbinervis | 9.8 |
| Scrubland after clear cut | Fagus crenata, Quercus crispula, Quercus serrata | |||
| 6 | Grassland | Seminatural grassland, Riparian grassland and marsh | - | 1.3 |
| 7 | Others | Settlement, bareland, rock, rice paddy, cropland, or waterbody | - | 3.2 |
2.5. Generalized Linear Model
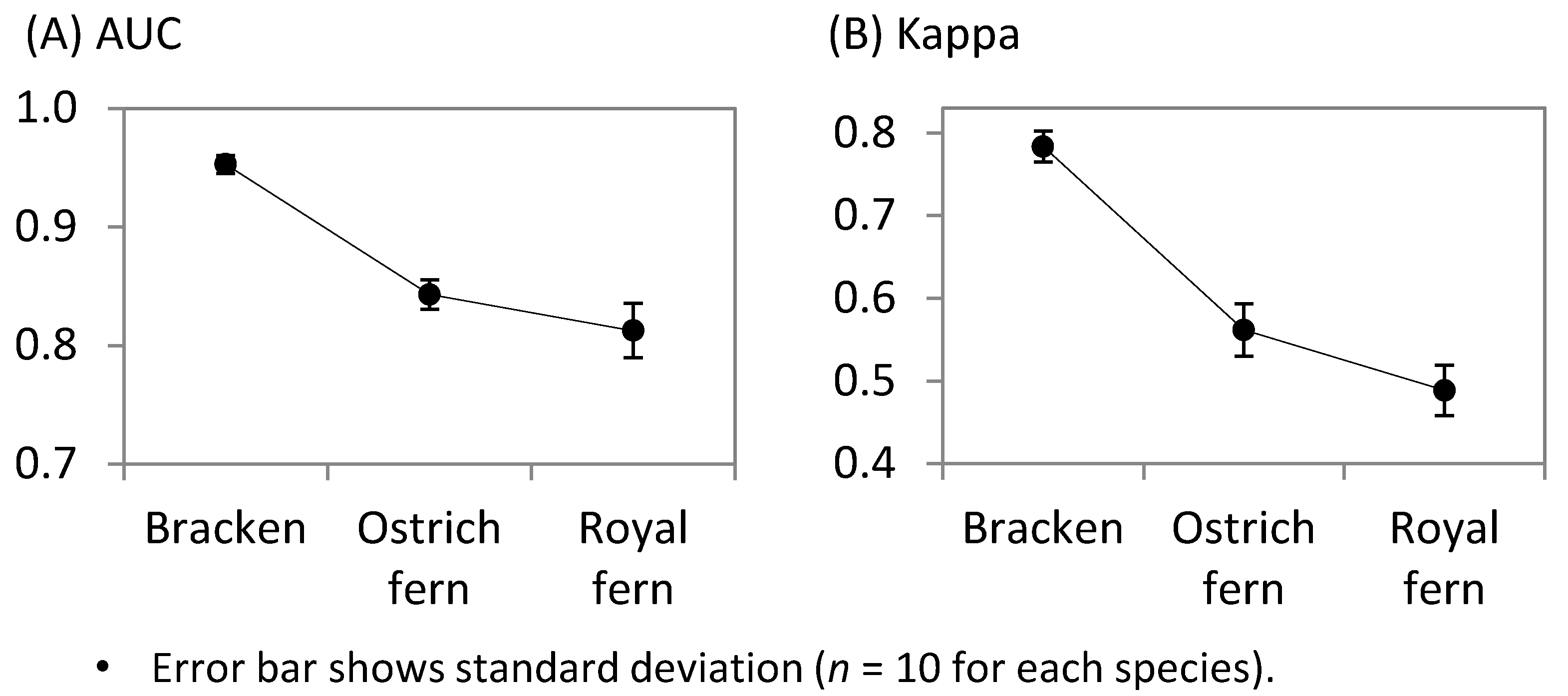
2.6. Model Verification
2.7. Comparison of the Potential Harvesting Sites among the Three Fern Species
3. Results
3.1. Comparison of the Potential Harvesting Sites among the Three Fern Species
| Selected variable 1 | Coefficient | z value | Odds ratio 2 | |
|---|---|---|---|---|
| Mean | Std. Err. | |||
| Ostrich fern | ||||
| Intercept | 2.0149 | 0.0645 | 31.224 | - |
| Grassland | 2.5130 | 0.2091 | 12.016 | 12.34 |
| Scrubland | −1.4180 | 0.1495 | −9.484 | 0.24 |
| Slope position | −2.2410 | 0.0992 | −22.589 | 0.11 |
| cos(aspectE) 3 | 0.6971 | 0.0478 | 14.597 | 2.01 |
| Proximity to roads | −0.0025 | 0.0001 | −22.807 | 0.08 |
| Bracken | ||||
| Intercept | −11.8500 | 0.5745 | −20.62 | - |
| Grassland | 4.9820 | 0.2402 | 20.74 | 145.77 |
| Scrubland | 1.7040 | 0.1448 | 11.77 | 5.50 |
| Mature conifer plantation | 1.1840 | 0.1028 | 11.52 | 3.27 |
| Young conifer plantation | 3.5000 | 0.1905 | 18.37 | 33.12 |
| Potential solar radiation | 8.65 × 10−6 | 4.32 × 10−7 | 20.03 | 2.37 |
| cos(aspectNE) 3 | 2.8490 | 0.1028 | 27.71 | 17.27 |
| Proximity to roads | −0.0041 | 0.0002 | −21.2 | 0.02 |
| Royal fern | ||||
| Intercept | 8.6190 | 0.2796 | 30.83 | - |
| Scrubland | 1.2400 | 0.0834 | 14.86 | 3.46 |
| Potential solar radiation | −6.72 × 10−6 | 2.23 × 10−7 | −30.2 | 0.51 |
| Slope position | −1.4970 | 0.0887 | −16.87 | 0.22 |
| cos(aspectSE) 3 | 1.3150 | 0.0571 | 23.05 | 3.72 |
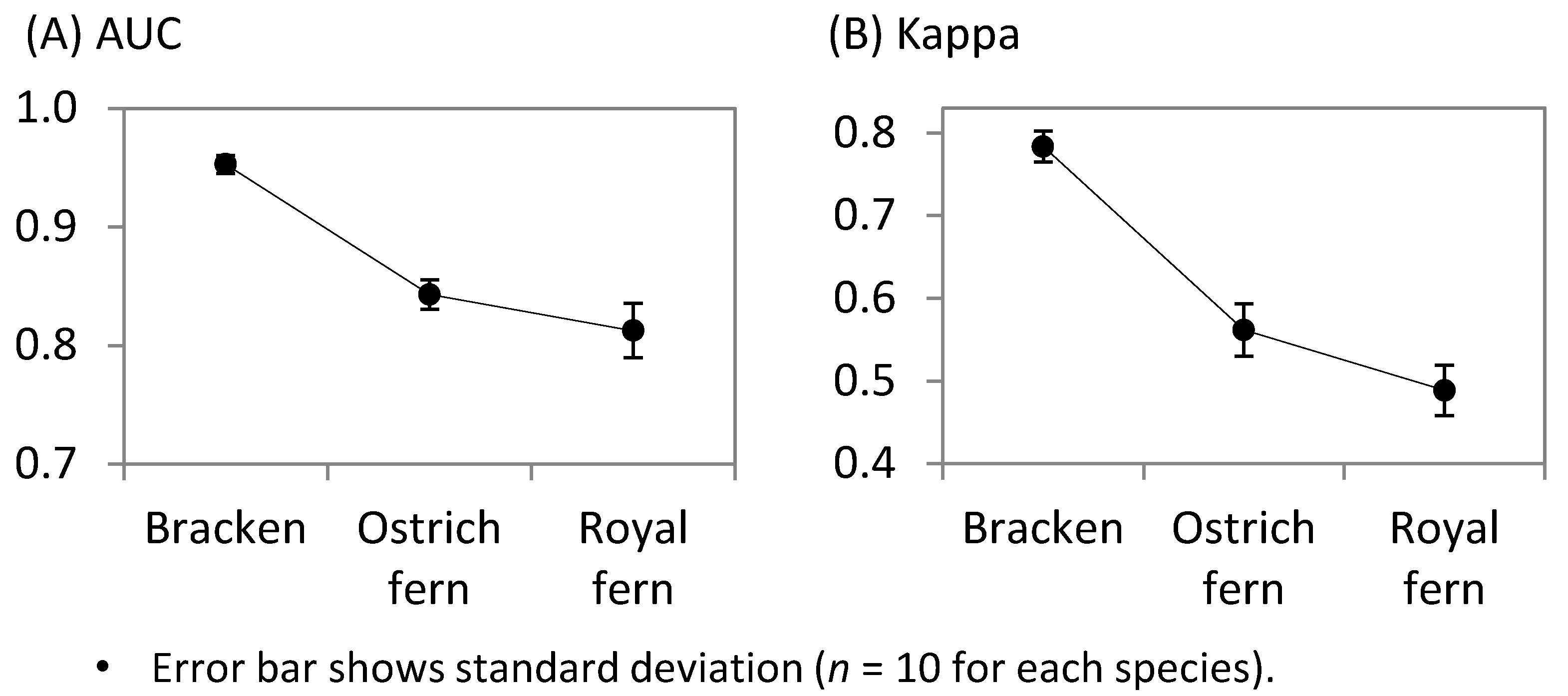
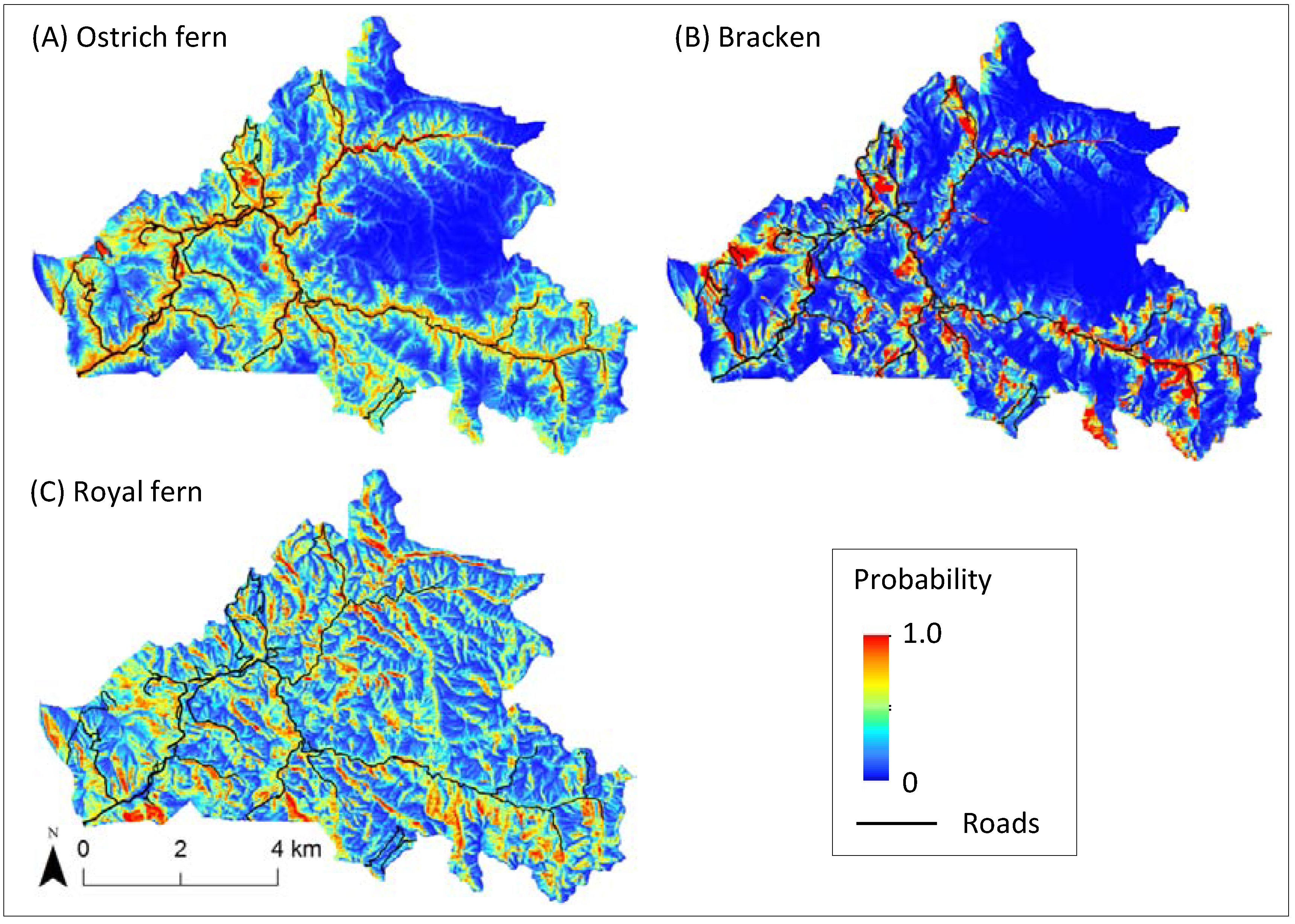
3.2. Estimated Potential Harvest Sites and their Differences among Ferns
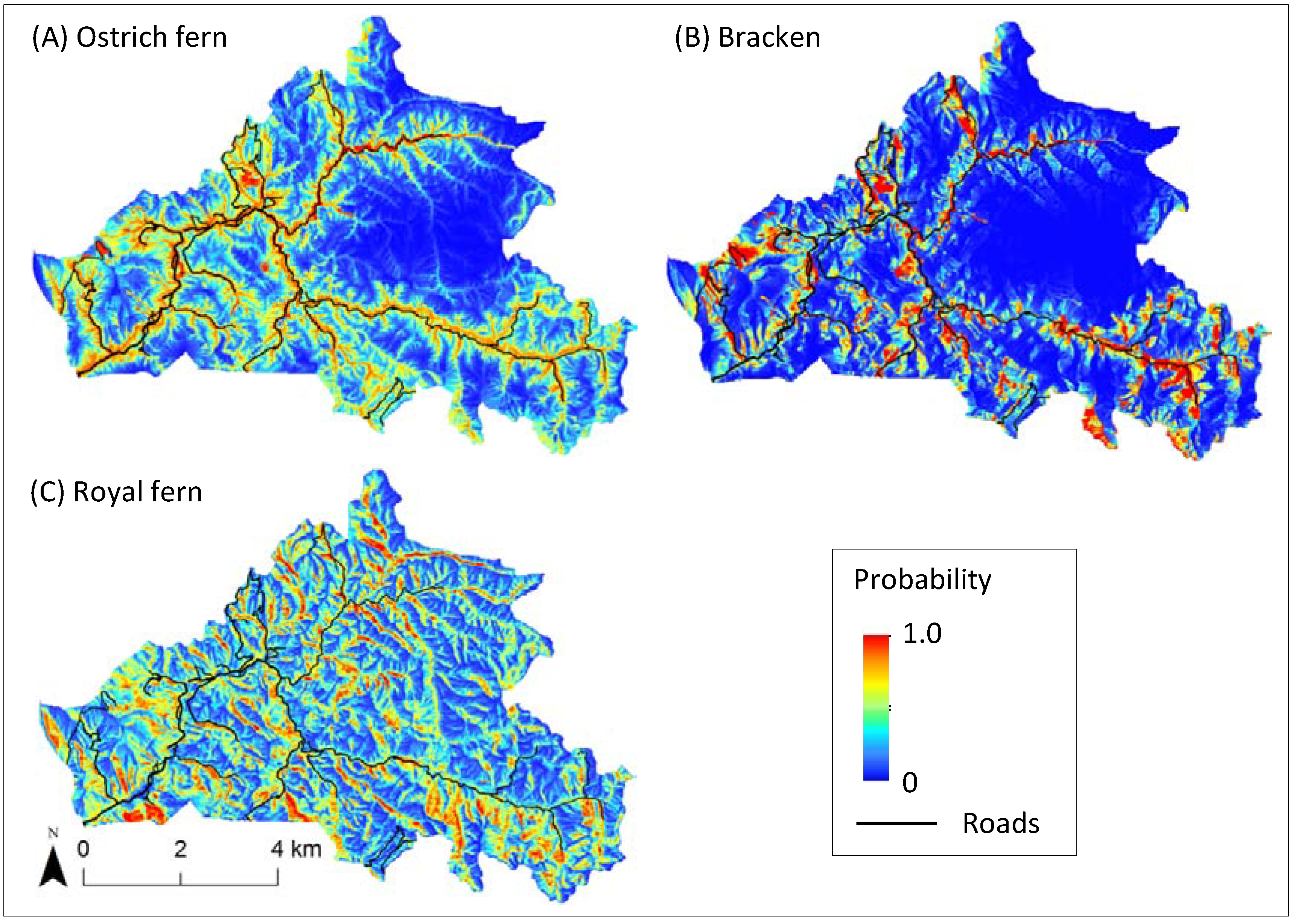
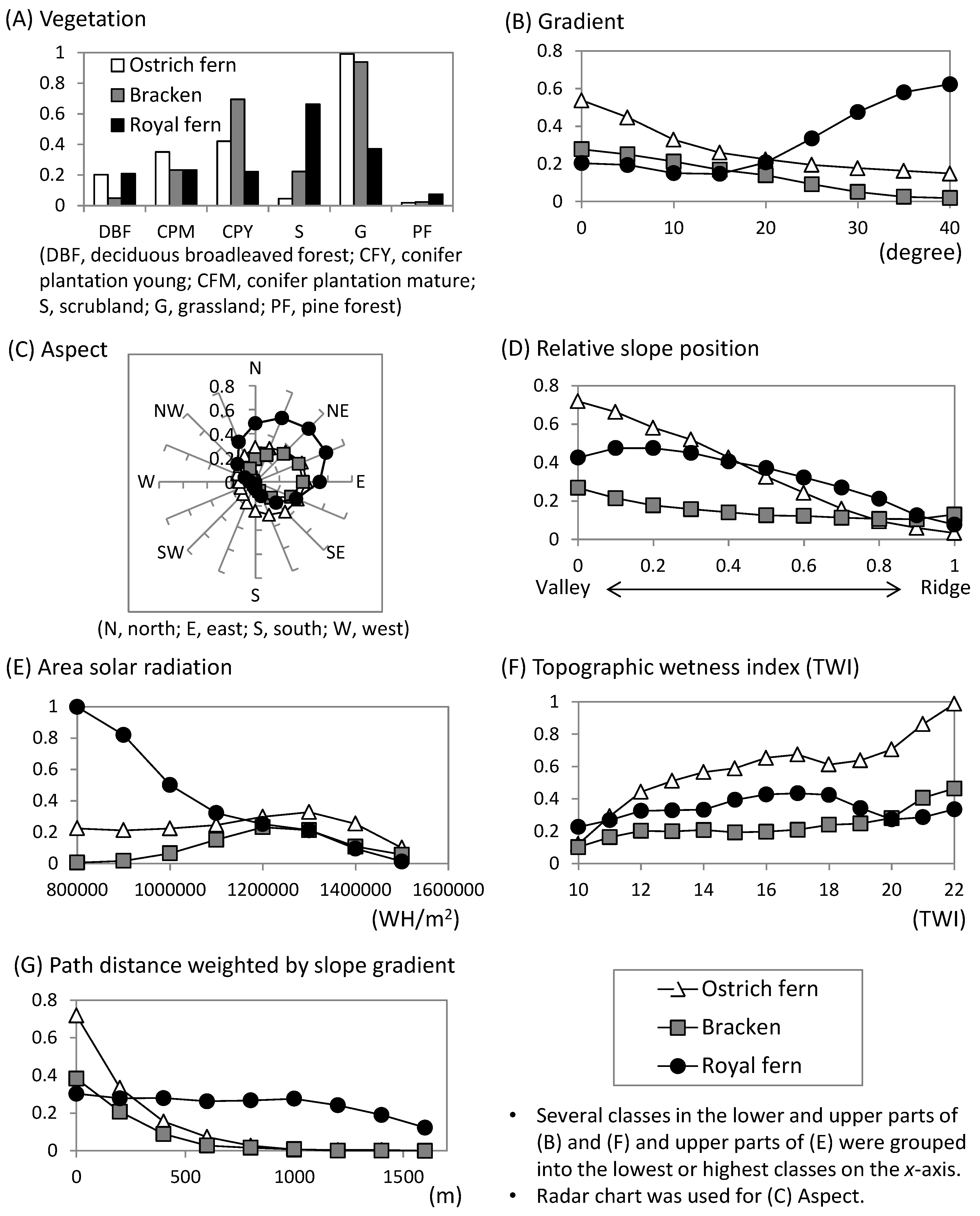
4. Discussion
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Delang, C.O. Not just minor forest products: The economic rationale for the consumption of wild food plants by subsistance farmers. Ecol. Econ. 2006, 59, 64–73. [Google Scholar] [CrossRef]
- Cocks, M.; López, C.; Dold, T. Cultural Importance of Non-Timber Forest Products: Opportunities They Pose for Bio-Cultural Diversity in Dynamic Society. In Non-Timber Forest Products in the Global Context, Tropical Forestry 7; Shackleton, S., Shackleton, C., Shanley, P., Eds.; Springer-Verlag: Heidelberg, Germany, 2011; pp. 107–128. [Google Scholar]
- Shackleton, C.M.; Pandey, A.K. Positioning non-timber forest products on the development agenda. For. Policy Econ. 2013, 38, 1–7. [Google Scholar] [CrossRef]
- Ticktin, T.; Shackleton, C. Harvesting Non-Timber Forest Products Sustainably: Opportunities and Challenges. In Non-Timber Forest Products in the Global Context, Tropical Forestry 7; Shackleton, S., Shackleton, C., Shanley, P., Eds.; Springer-Verlag: Heidelberg, Germany, 2011; pp. 149–169. [Google Scholar]
- Schreckenberg, K. Products of a managed landscape: Non-timber forest products in the parklands of the Bassila region, Benin. Glob. Ecol. Biogeogr. 2013, 8, 279–289. [Google Scholar] [CrossRef]
- Matsuura, T.; Sugimura, K.; Miyamoto, A.; Tanaka, N. Knowledge-based estimation of edible fern harvesting sites in mountainous communities of northeastern Japan. Sustainability 2014, 6, 175–192. [Google Scholar]
- Austin, M.P. Spatial prediction of species distribution: An interface between ecological theory and statistical modelling. Ecol. Model. 2002, 157, 101–118. [Google Scholar] [CrossRef]
- Boyce, M.S.; Vernier, P.R.; Nielsen, S.E.; Schmiegelow, F.K.A. Evaluating resource selection functions. Ecol. Model. 2002, 157, 281–300. [Google Scholar] [CrossRef]
- Guisan, A.; Zimmermann, N.E. Predictive habitat distribution models in ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Nishimura, Y.; OKamoto, K.; Somkhit, B. Time-geographic analysis on natural resource use in a village of the Vientiane Plain. Southeast Asian Stud. 2010, 47, 426–450. [Google Scholar]
- Saito, H. Wild edible plant gathering activities practiced at ecotones. Bull. Natl. Mus. Jpn. Hist. 2005, 123, 325–353, (in Japanese with English abstract). [Google Scholar]
- Ikeya, K. Spatial structure of Zenmai gathering: A case study of the mountain village in the heavy snow zone. Sci. Rep. Tohoku Univ. Ser. Geogr. 1988, 38, 104–118. [Google Scholar]
- Ikeya, K. Cultural ecology of Zenmai gathering in the northeastern Japan. Sci. Rep. Tohoku Univ. Ser. Geogr. 2004, 53, 1–28. [Google Scholar]
- Kawarasaki, H.S.; Sugimura, K. Estimation of frequency and locality of collection of wild mushroom and wild vegetables by internet search. J. Jpn. For. Soc. 2012, 94, 95–99, (in Japanese with English abstract). [Google Scholar] [CrossRef]
- Matsuura, T.; Hayashi, M.; Sugimura, K.; Tanaka, N.; Miyamoto, A. Ecosystem services valuation of gathering edible wild plants/mushrooms—A case study in Tadami town, Fukushima prefecture. Jpn. J. For. Plan. 2013, 47, 55–81, (in Japanese with English abstract). [Google Scholar]
- Higa, M.; Nakao, K.; Tsuyama, I.; Nakazono, E.; Yasuda, M.; Matsui, T.; Tanaka, N. Indicator plant species selection for monitoring the impact of climate change based on prediction uncertainty. Ecol. Indic. 2013, 29, 307–315. [Google Scholar] [CrossRef]
- Kashimura, T. Ecological study of the natural forest vegetation in the snowy region along the lower Tadami valley. Ecol. Rev. 1968, 17, 153–170. [Google Scholar]
- Kawano, S.; Nikkuni, I. Forest Vegetation, Flora and Fauna, and Biodiversity in Tadami-machi, Fukushima Prefecture, Honshu, Japan -Primaeval Beech and Riparian Forests, Sustainable Rich Flora and Fauna; Tadami-machi Board of Education: Tadami-machi, Japan, 2004; p. 23. [Google Scholar]
- Iwatsuki, K. Ferns and Fern Allies of Japan; (in Japanese with English Abstract); Heibonsha Ltd.: Tokyo, Japan, 1992; p. 311. [Google Scholar]
- Sato, T. Phenology and wintering capacity of sporophytes and gametophytes of ferns native to northern Japan. Oecologia 1982, 55, 53–61. [Google Scholar] [CrossRef]
- Odland, A.; Naujalis, J.R.; Stapulionytė, A. Variation in the structure of Matteuccia struthiopteris populations in Lithuania. Biologija 2006, 1, 83–90. [Google Scholar]
- Von Aderkas, P. Economic history of ostrich fern, Matteuccia struthiopteris, the edible fiddlehead. Econ. Bot. 1984, 38, 14–23. [Google Scholar] [CrossRef]
- Page, C.N. The taxonomy and phytogeography of bracken. Bot. J. Linn. Soc. 1976, 73, 1–34. [Google Scholar] [CrossRef]
- Pakeman, R.J.; Marrs, R.H. The conservation value of bracken, Pteridium aquilinum (L.) Kuhn, dominated communities in the UK, and an assessment of the ecological impact of bracken expansion or its removal. Biol. Conserv. 1992, 62, 101–114. [Google Scholar] [CrossRef]
- Wilson, D.; Donaldson, L.J.; Sepai, O. Should we be frightened of bracken? A review of the evidence. J. Epidemiol. Community Health 1998, 52, 812–817. [Google Scholar] [CrossRef]
- Gil da Costa, R.M.; Bastos, M.; Oliveira, P.A.; Lopes, C. Bracken-associated human and animal health hazards: Chemical, biological and pathological evidence. J. Hazard. Mater. 2012, 203–204, 1–12. [Google Scholar] [CrossRef]
- Hodge, W.H. Fern foods of Japan and the probrem of toxicity. Am. Fern J. 1973, 63, 77–80. [Google Scholar] [CrossRef]
- May, L.W. The economic uses and associated folklore of ferns and fern allies. Bot. Rev. 1978, 44, 491–528. [Google Scholar] [CrossRef]
- Pemberton, R.; Lee, N. Wild food plants in South Korea; market presence, new crops, and exports to the United States. Econ. Bot. 1996, 50, 57–70. [Google Scholar] [CrossRef]
- Hirono, I. Edible plants containing naturally occurring carcinogens in Japan. Jpn. J. Cancer Res. 1993, 84, 997–1006. [Google Scholar] [CrossRef]
- Kato, M. Distribution of Osmundaceae. Bull. Natl. Mus. Nat. Sci. Ser. B Bot. 2007, 33, 81–90. [Google Scholar]
- Beyer, H.L. Geospatial Modelling Environment. Version: 0.7.2 RC2. 2012, p. 158. Available online: www.spatialecology.com/gme (accessed on 17 February 2014).
- Hirzel, A.H.; Guisan, A. Which is the optimal sampling strategy for habitat suitability modelling. Ecol. Model. 2002, 157, 331–341. [Google Scholar] [CrossRef]
- Mizuno, N. Sansai/Kinomi (Edible Wild Plants in Japan). In Yama-Kei Pocket Guide 4; (in Japanese). Yama-Kei Publishers Co., Ltd: Tokyo, Japan, 1999; p. 281. [Google Scholar]
- Miller, J.; Franklin, J. Modeling the distribution of four vegetation alliances using generalized linear models and classification trees with spatial dependence. Ecol. Model. 2002, 157, 227–247. [Google Scholar] [CrossRef]
- Rennó, C.D.; Nobre, A.D.; Cuartas, L.A.; Soares, J.V.; Hodnett, M.G.; Tomasella, J.; Waterloo, M.J. HAND, a new terrain descriptor using SRTM-DEM: Mapping terra-firme rainforest environments in Amazonia. Remote Sens. Environ. 2008, 112, 3469–3481. [Google Scholar] [CrossRef]
- Tarboton, D.G. A new methods for the determination of flow directions and upslope areas in grid digital elevation models. Water Resour. Res. 1997, 33, 309–319. [Google Scholar] [CrossRef]
- Kawabata, Y. Shokusei: Tadami-machi-no-Shinrin-ya-Sougen (Vegetation: Forest and Grassland in Tadami Town). In Aizu Tadami no Shizen: Shokubutu-hen (Nature in Aizu-Tadami: Vegetation); (in Japanese). Editorial Committee of the History of Tadami: Tadami-machi, Japan, 2001; pp. 52–71. [Google Scholar]
- Whittingham, M.J.; Stephens, P.A; Bradbury, R.B.; Freckleton, R.P. Why do we still use stepwise modelling in ecology and behaviour? J. Anim. Ecol. 2006, 75, 1182–1189. [Google Scholar] [CrossRef]
- Murtaugh, P.A. Performance of several variable-selection methods applied to real ecological data. Ecol. Lett. 2009, 12, 1061–1068. [Google Scholar] [CrossRef]
- Fielding, H.A.; Bell, F.J. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Manel, S.; Williams, C.H.; Ormerod, J.S. Evaluating presence-absence models in ecology: The need to account for prevalence. J. Appl. Ecol. 2001, 38, 921–931. [Google Scholar]
- Freeman, E.; Moisen, G. A comparison of the performance of threshold criteria for binary classification in terms of predicted prevalence and kappa. Ecol. Model. 2008, 217, 48–58. [Google Scholar] [CrossRef]
- Pearce, J.; Ferrier, S. Evaluating the predictive performance of habitat models developed using logistic regression. Ecol. Model. 2000, 133, 225–245. [Google Scholar] [CrossRef]
- Sato, T. Comparative life history of aspidiaceous ferns in northern Japan with reference to fertility during sporophyte development in relation to habitats. Bot. Mag. Tokyo 1985, 98, 371–381. [Google Scholar] [CrossRef]
- Landi, M.; Angiolini, G. Ecological responses of Osmunda regalis to forest canopy cover and grazing. Am. Fern J. 2010, 100, 137–147. [Google Scholar] [CrossRef]
- Suzuki, W.; Osumi, K.; Masaki, T. Disturbance regimes and community structures of a riparian and an adjacent terrace stand in the Kanumazawa Riparian Research Forest, northern Japan. For. Ecol. Manage. 2002, 157, 285–301. [Google Scholar] [CrossRef]
- Kawanishi, M.; Sakio, H.; Kubo, M.; Shimano, K.; Ohno, K. Effect of micro-landforms on forest vegetation differentiation and life-form diversity in the Chichibu Mountains, Kanto District, Japan. Veg. Sci. 2006, 23, 13–24. [Google Scholar]
- Shimada, Y. Ecology of bracken (Pteridium aquilinum (L.) Kuhn): Dynamics of bracken population in native grassland. Weed Res. Jpn. 1962, 1, 70–77, (in Japanese with English abstract). [Google Scholar]
- Miyama, T.; Goto, Y. The soil conservation function of bracken fern (Pteridium aquilinum Kuhn var. latiuschlum Underw) after the forest fire. Jpn. Soc. Reveg. Technol. 2000, 26, 36–41, (in Japanese with English abstract). [Google Scholar] [CrossRef]
- Takahashi, Y.; Ide, Y.; Kobayashi, H.; Hayasaka, K. Effects of cutting management on the vegetation changes of the Zoysia-Pteridium-type pasture in western Honshu, Japan. Jpn. J. Grassl. Sci. 2008, 54, 230–235, (in Japanese with English abstract). [Google Scholar]
- Yukawa, Y. Henyou Suru Sanson (Changing Mountainous Villages in Japan); (in Japanese). Japan Editors School Publishing Division: Tokyo, Japan, 1991; p. 328. [Google Scholar]
- Shimokawa, K. Geomorphic study of avalanche chute in the upper drainage basin of the Tadami River. Geogr. Rev. Jpn. 1980, 53, 171–188, (in Japanese with English abstract). [Google Scholar] [CrossRef]
- Aiura, H. Erosive phenomenon in heavy snow mountainous regions in Japan. J. For. Res. 2002, 7, 57–66. [Google Scholar] [CrossRef]
- Matsuura, T.; Suzuki, W. Analysis of topography and vegetation distribution using a digital elevation model: Case study of a snowy mountain basin in northeastern Japan. Landsc. Ecol. Eng. 2013, 9, 143–155. [Google Scholar] [CrossRef]
- Miida, K. Zenmai (royal fern) production and its village sustaining function in the Tohoku district of Japan. Geogr. Rev. Jpn. 1974, 47, 370–386, (in Japanese with English abstract). [Google Scholar] [CrossRef]
- Fukamachi, K.; Oku, H.; Nakashizuka, T. The change of a satoyama landscape and its causality in Kamiseya, Kyoto Prefecture, Japan between 1970 and 1995. Landsc. Ecol. 2001, 16, 703–717. [Google Scholar] [CrossRef]
- Osumi, K.; Ikeda, S.; Okamoto, T. Vegetation patterns and their dependency on site conditions in the pre-industrial landscape of north-eastern Japan. Ecol. Res. 2003, 18, 753–765. [Google Scholar] [CrossRef]
- Nagaike, T.; Kamitani, T. Factors affecting changes in a landscape structure dominated by both primary and coppice forests in the Fagus crenata forest region of central Japan. J. For. Res. 1997, 2, 193–198. [Google Scholar] [CrossRef]
- Miyamoto, A.; Sano, M. The influence of forest management on landscape structure in the cool-temperate forest region of central Japan. Landsc. Urban Plan. 2008, 86, 248–256. [Google Scholar] [CrossRef]
- Albers, H.; Robinson, E. A review of the spatial economics of non-timber forest product extraction: Implications for policy. Ecol. Econ. 2013, 92, 87–95. [Google Scholar] [CrossRef]
- Nackoney, J.; Rybock, D.; Dupain, J.; Facheux, C. Coupling participatory mapping and GIS to inform village-level agricultural zoning in the Democratic Republic of the Congo. Landsc. Urban Plan. 2013, 110, 164–174. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Matsuura, T.; Sugimura, K.; Miyamoto, A.; Tanaka, H.; Tanaka, N. Spatial Characteristics of Edible Wild Fern Harvesting in Mountainous Villages in Northeastern Japan Using GPS Tracks. Forests 2014, 5, 269-286. https://doi.org/10.3390/f5020269
Matsuura T, Sugimura K, Miyamoto A, Tanaka H, Tanaka N. Spatial Characteristics of Edible Wild Fern Harvesting in Mountainous Villages in Northeastern Japan Using GPS Tracks. Forests. 2014; 5(2):269-286. https://doi.org/10.3390/f5020269
Chicago/Turabian StyleMatsuura, Toshiya, Ken Sugimura, Asako Miyamoto, Hiroshi Tanaka, and Nobuhiko Tanaka. 2014. "Spatial Characteristics of Edible Wild Fern Harvesting in Mountainous Villages in Northeastern Japan Using GPS Tracks" Forests 5, no. 2: 269-286. https://doi.org/10.3390/f5020269
APA StyleMatsuura, T., Sugimura, K., Miyamoto, A., Tanaka, H., & Tanaka, N. (2014). Spatial Characteristics of Edible Wild Fern Harvesting in Mountainous Villages in Northeastern Japan Using GPS Tracks. Forests, 5(2), 269-286. https://doi.org/10.3390/f5020269