Structure and Composition of Old-Growth and Unmanaged Second-Growth Riparian Forests at Redwood National Park, USA

Abstract
:1. Introduction
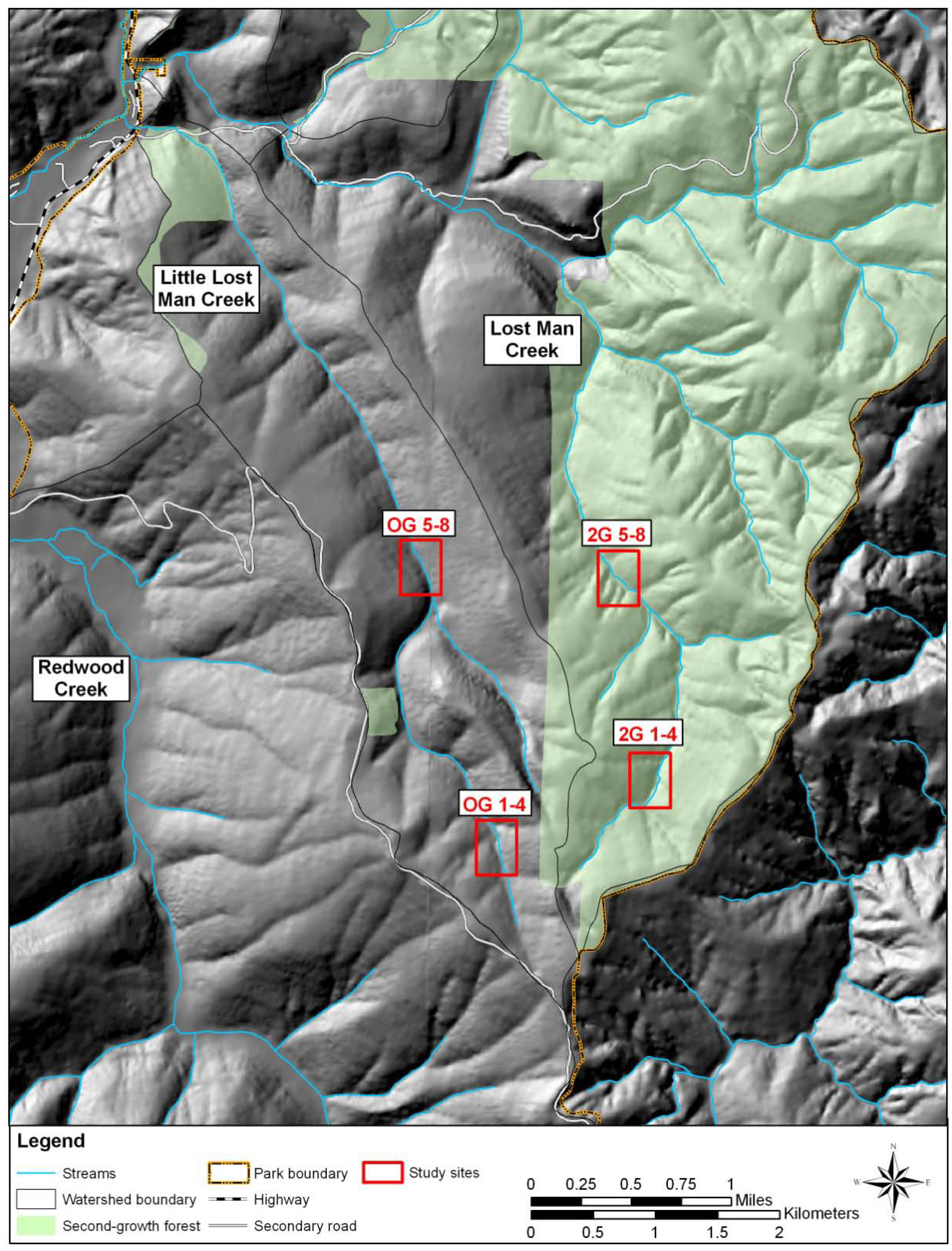
2. Experimental Section
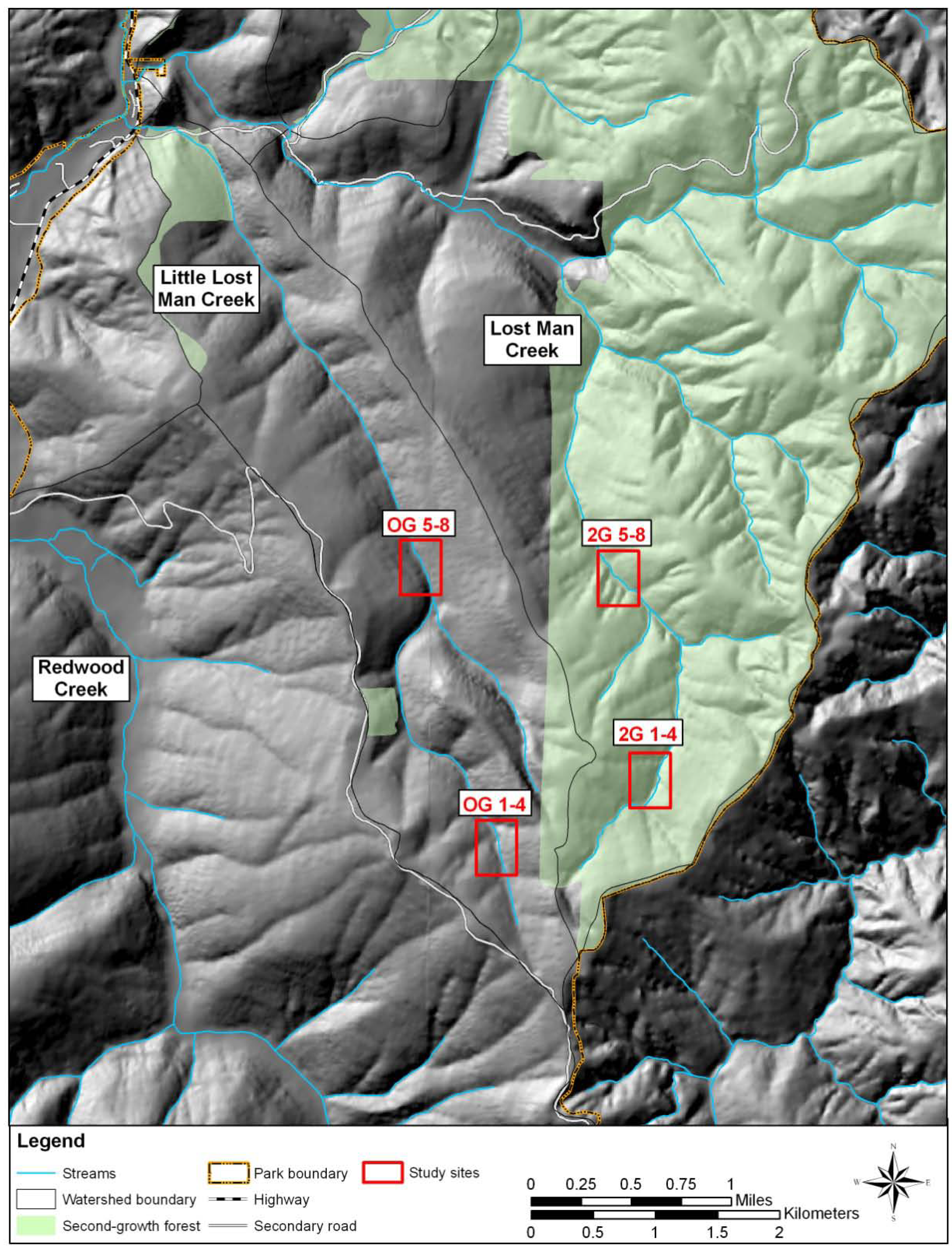
3. Results
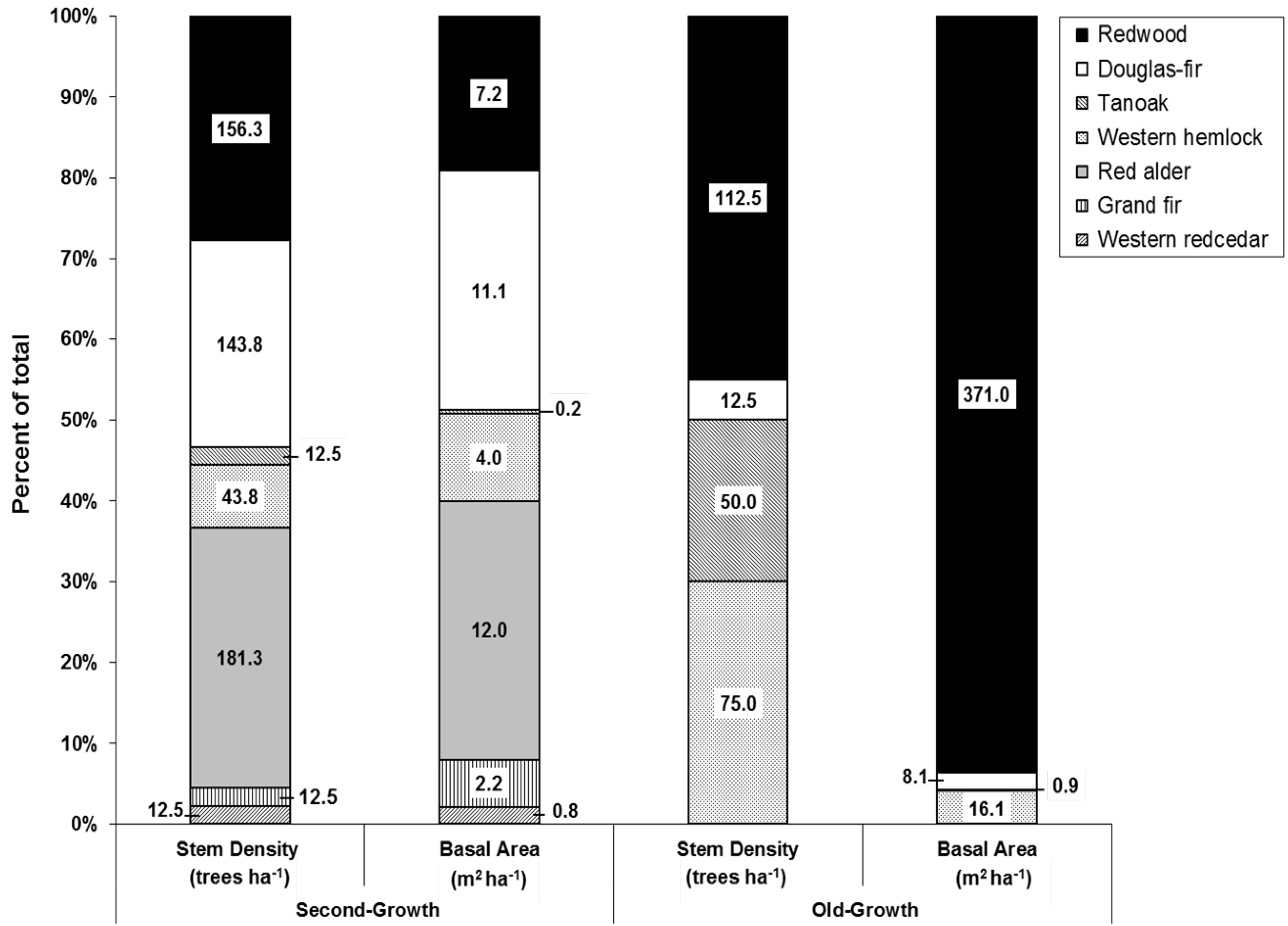
3.1. Overstory Structure and Composition

{kind=link}
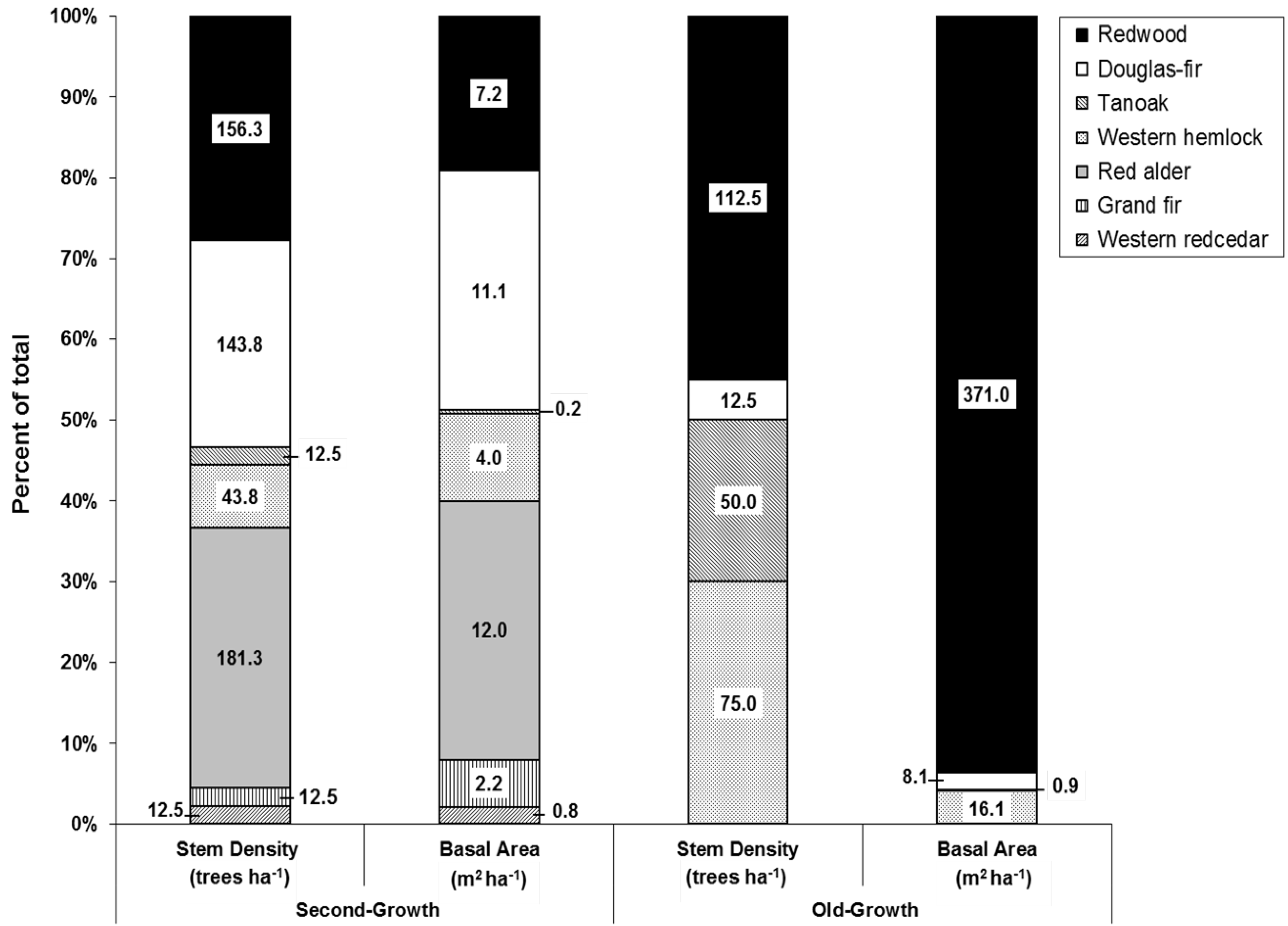{kind=link}
| Species | Frequency (# plots) | Relative Frequency a (%) | Stem Density (trees ha−1) | Relative Density b (%) | Basal Area (m2 ha−1) | Relative BA c (%) | Importance Value d (%) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| OG | 2G | OG | 2G | OG | 2G | OG | 2G | OG | 2G | OG | 2G | OG | 2G | |
| Douglas-fir | 1 | 6 | 12.5 | 75.0 | 12.5 | 143.8 | 5.0 | 25.0 | 8.1 | 11.1 | 2.0 | 18.1 | 6.54 | 39.42 |
| Grand fir | - | 1 | - | 12.5 | - | 12.5 | - | 2.2 | - | 2.2 | - | 3.6 | - | 6.17 |
| Red alder | - | 7 | - | 87.5 | - | 181.3 | - | 31.5 | - | 12.0 | - | 19.5 | - | 46.21 |
| Redwood | 7 | 5 | 87.5 | 62.5 | 112.5 | 156.3 | 45.0 | 27.2 | 371.0 | 7.2 | 93.7 | 11.7 | 75.41 | 33.83 |
| Tanoak | 4 | 2 | 50.0 | 25.0 | 50 | 12.5 | 20.0 | 2.2 | 0.9 | 0.2 | 0.2 | 0.3 | 23.43 | 9.26 |
| W. hemlock | 7 | 2 | 87.5 | 25.0 | 75 | 43.8 | 30.0 | 7.6 | 16.1 | 4.0 | 4.1 | 6.5 | 40.52 | 13.05 |
| W. redcedar | - | 1 | - | 12.5 | - | 12.5 | - | 2.2 | - | 0.8 | - | 1.3 | - | 5.38 |
| Rdwd rsdle | N/A | 2 | N/A | 25.0 | N/A | 12.5 | N/A | 2.2 | N/A | 24.0 | N/A | 39.0 | N/A | 22.14 |
| Totals | N/A | N/A | N/A | N/A | 250 | 575.0 | 100.0 | 100.0 | 396.1 | 61.6 | 100.0 | 100.0 | - | - |
3.2. Understory Vegetation
| Species | Relative Frequency a | Relative Cover b | Importance Value (IV) c | |||
|---|---|---|---|---|---|---|
| OG | 2G | OG | 2G | OG | 2G | |
| Adiantum sp. | 4 | 4 | 0.1 | 0.4 | 2.2 | 2.3 |
| Asplenium sp. | 0 | 4 | 0.0 | 0.0 | 0.0 | 2.1 |
| Athyrium filix-femina | 17 | 21 | 0.4 | 1.8 | 8.67 | 11.37 |
| Atriplex rosea | 13 | 0 | 0.2 | 0.0 | 6.49 | 0.0 |
| Blechnum spicant | 67 | 67 | 15.4 | 10.6 | 41.03 | 38.72 |
| Carex sp. | 4 | 25 | 0.1 | 5.0 | 2.2 | 15.06 |
| Disporum hookeri | 8 | 0 | 0.2 | 0.0 | 4.3 | 0.0 |
| Dryopteris sp. | 0 | 29 | 0.0 | 1.1 | 0.0 | 15.15 |
| Equisetum sp. | 0 | 21 | 0.0 | 0.5 | 0.0 | 10.79 |
| Galium sp. | 17 | 21 | 0.3 | 0.5 | 8.58 | 10.710 |
| Gaultheria shallon | 58 | 50 | 11.4 | 8.6 | 34.94 | 29.33 |
| Goodyera oblongifolia | 8 | 0 | 0.1 | 0.0 | 4.2 | 0.0 |
| Mahonia nervosa | 8 | 8 | 0.6 | 0.4 | 4.5 | 4.4 |
| Oxalis oregana | 96 | 21 | 16.4 | 1.1 | 56.12 | 11.08 |
| Polystichum munitum | 92 | 96 | 50.7 | 52.6 | 71.21 | 74.21 |
| Ranunculus uncinatus | 0 | 8 | 0.0 | 0.6 | 0.0 | 4.4 |
| Rhododendron macrophyllum | 4 | 0 | 0.3 | 0.0 | 2.3 | 0.0 |
| Rhododendron occidentale | 8 | 0 | 1.4 | 0.0 | 4.810 | 0.0 |
| Ribes sp. | 0 | 4 | 0.0 | 0.1 | 0.0 | 2.1 |
| Rubus spectabilis | 0 | 8 | 0.0 | 3.6 | 0.0 | 6.0 |
| Rubus ursinus | 0 | 8 | 0.0 | 0.2 | 0.0 | 4.3 |
| Trientalis borealis | 4 | 4 | 0.0 | 0.1 | 2.1 | 2.1 |
| Trillium ovatum | 46 | 8 | 0.9 | 0.1 | 23.45 | 4.2 |
| Vaccinium ovatum | 4 | 4 | 0.2 | 0.2 | 2.2 | 2.2 |
| Vaccinium parvifolium | 21 | 38 | 1.1 | 12.0 | 11.06 | 24.74 |
| Viola sempervirens | 4 | 13 | 0.1 | 0.4 | 2.1 | 6.5 |
| s | N | H’ | J’ | Percent cover | ||
|---|---|---|---|---|---|---|
| Herbs | Old-growth | 14 | 14.91 (9.62) | 0.41 (0.14) | 0.32 (0.11) | 9.1 (7.7) |
| Second-growth | 16 | 16.38 (18.58) | 0.29 (0.19) | 0.23 (0.15) | 52.3 (26.0) | |
| Shrubs | Old-growth | 5 | 7.29 (14.60) | 0.11 (0.17) | 0.14 (0.21) | 39.0 (20.1) |
| Second-growth | 5 | 7.34 (8.96) | 0.23 (0.13) | 0.29 (0.16) | 13.3 (13.5) | |
| All | Old-growth | 19 | 14.32 (9.60) | 0.45 (0.17) | 0.33 (0.13) | 62.0 (22.7) |
| Second-growth | 21 | 11.34 (7.24) | 0.45 (0.16) | 0.32 (0.12) | 52.9 (27.3) |
4. Discussion
Acknowledgments
Conflicts of Interest
References and Notes
- Porter, D.; Gizinski, V.; Hartley, R.; Kramer, S.H. Restoring Complexity to Industrially Managed Timberlands: The Mill Creek Interim Management Recommendations and Early Restoration Thinning Treatments. In Proceedings of the Redwood Science Symposium: What Does the Future Hold; General Technical Report PSW-GTR-194; USDA Forest Service Pacific Southwest Research Station: Albany, CA, USA, 2007; pp. 283–294. [Google Scholar]
- Lorimer, C.G.; Porter, D.J.; Madej, M.A.; Stuart, J.D.; Viers, S.D., Jr.; Norman, S.P.; O’Hara, K.L.; Libby, W.J. Presettlement and modern disturbance regimes in coast redwood forests: Implications for the conservation of old-growth stands. For. Ecol. Manag. 2009, 258, 1038–1054. [Google Scholar] [CrossRef]
- Keyes, C.R.; Perry, T.E.; Plummer, J.F. Variable-Density Thinning for Parks and Reserves: An Experimental Case Study at Humboldt Redwoods State Park, California. In Proceedings of the 2009 National Silviculture Workshop; Proceedings RMRS-P-61. USDA Forest Service Rocky Mountain Research Station: Fort Collins, CO, USA, 2010; pp. 227–237. [Google Scholar]
- O’Hara, K.L.; Nesmith, J.C.B.; Leonard, L.; Porter, D.J. Restoration of old forest features in coast redwood forests using early-stage variable-density thinning. Restor. Ecol. 2010, 18, 125–135. [Google Scholar] [CrossRef]
- Chittick, A.J.; Keyes, C.R. Holter Ridge Thinning Study, Redwood National Park: Preliminary Results of a 25-Year Retrospective. In Proceedings of the Redwood Science Symposium: What Does the Future Hold; General Technical Report PSW-GTR-194; USDA Forest Service Pacific Southwest Research Station: Albany, CA, USA, 2007; pp. 271–280. [Google Scholar]
- Teraoka, J.R.; Keyes, C.R. Low thinning as a forest restoration tool at Redwood National Park. West. J. Appl. For. 2011, 26, 91–93. [Google Scholar]
- Plummer, J.F.; Keyes, C.R.; Varner, J.M. Early-Stage thinning for the restoration of young redwood—Douglas-fir forests in northern coastal California, USA. ISRN Ecol. 2012, 2012. [Google Scholar] [CrossRef]
- Keller, E.A.; MacDonald, A.; Tally, T.; Merritt, N.J. Effects of large organic debris on channel morphology and sediment storage in selected tributaries of Redwood Creek, northwestern California. In Geomorphic Processes and Aquatic Habitat in the Redwood Creek Basin, Northwestern California; Nolan, K.M., Kelsey, H.M., Marron, D.C., Eds.; United States Geological Survey: Denver, CO, USA, 1995; pp. P1–P29. [Google Scholar]
- Lisle, T.E.; Napolitano, M.B. Effects of Recent Logging on the Main Channel of North Fork Caspar Creek. In Proceedings of the Conference on Coastal Watersheds: The Caspar Creek Story; General Technical Report PSW-GTR-168. USDA Forest Service Pacific Southwest Research Station: Albany, CA, USA, 1988; pp. 81–85. [Google Scholar]
- Andrus, C.W. Woody Debris from the Streamside Forest and its influence on Fish Habitat. In Hydrological and Biological Responses to Forest Practices; Stednick, J.D., Ed.; Springer Science and Business Media LLC: New York, NY, USA, 2008; pp. 211–235. [Google Scholar]
- Spies, T.A.; Franklin, J.F. The Structure of Natural Young, Mature, and Old-Growth Douglas-fir Forests in Oregon and Washington. In Wildlife and Vegetation of Unmanaged Douglas-Fir Forests; General Technical Report PNW-GTR-285; Ruggiero, L.F., Aubry, K.B., Carey, A.B., Huff, M.H., Eds.; USDA Forest Service Pacific Northwest Research Station: Portland, OR, USA, 1991; pp. 91–109. [Google Scholar]
- Naiman, R.J.; Fetherston, K.L.; McKay, S.J.; Chen, J. Riparian Forests. In River Ecology and Management: Lessons from the Pacific Coastal Ecoregion; Naiman, R.J., Bilby, R.E., Eds.; Springer-Verlag: New York, NY, USA, 1998; pp. 289–323. [Google Scholar]
- Busing, R.T.; Fujimori, T. Dynamics of composition and structure in an old Sequoia sempervirens forest. J. Veg. Sci. 2002, 13, 785–792. [Google Scholar]
- Zenner, E.K. Does old-growth condition imply high live-tree structural complexity? For. Ecol. Manag. 2004, 195, 243–258. [Google Scholar] [CrossRef]
- Urner, S.; Madej, M. Changes in Riparian Composition and Density Following Timber Harvest and Floods along Redwood Creek, California. In Ecosystem Restoration: Turning the Tide; Society for Ecological Restoration Northwest Chapter Annual Meeting, Society for Ecological Restoration Northwest Chapter: Tacoma, WA, USA, 1998. [Google Scholar]
- Russell, W. The influence of timber harvest on the structure and composition of riparian forests in the Coastal Redwood region. For. Ecol. Manag. 2009, 257, 1427–1433. [Google Scholar] [CrossRef]
- Villarin, L.A.; Chapin, D.M.; Jones, J.E. Riparian forest structure and succession in second-growth stands of the central Cascade Mountains, Washington, USA. For. Ecol. Manag. 2009, 257, 1375–1385. [Google Scholar] [CrossRef]
- Magnuson, J.J.; Allendorf, F.W.; Beschta, R.L.; Bisson, P.A.; Carson, H.L.; Chapman, D.W.; Hanna, S.S.; Kapuscinski, A.R.; Lee, K.N.; Lettenmaier, D.P.; et al. Upstream: Salmon and Society in the Pacific Northwest; National Academy Press: Washington, DC, USA, 1996. [Google Scholar]
- Vannote, R.L.; Minshall, G.W.; Cummins, K.W.; Sedell, J.R.; Cushing, C.E. The river continuum concept. Can. J. Fish. Aquat. Sci. 1980, 37, 130–137. [Google Scholar] [CrossRef]
- Gregory, S.V.; Swanson, F.J.; McKee, W.A.; Cummins, K.W. An ecosystem perspective of riparian zones. Bioscience 1991, 41, 540–551. [Google Scholar] [CrossRef]
- Hibbs, D.E.; Bower, A.L. Riparian forests in the Oregon Coast Range. For. Ecol. Manag. 2001, 154, 201–213. [Google Scholar] [CrossRef]
- Bilby, R.E.; Bisson, P.A. Allochthonous versus autochthonous organic matter contributions to the trophic support of fish populations in clear-cut and old-growth forested streams. Can. J. Fish. Aquat. Sci. 1992, 49, 540–551. [Google Scholar] [CrossRef]
- Redwood National and State Parks. Unpublished GIS Stream Layer for the Lost Man Creek and Little Lost Man Creek Watersheds; Data stored at Redwood National and State Parks; South Operations Center: Orick, CA, USA, 2010. [Google Scholar]
- Redwood National and State Parks. Redwood National and State Parks’ Strategic Plan: October 1, 2005–September 30, 2008; USDI National Park Service, Redwood National Park: Crescent City, CA, USA, 2004. [Google Scholar]
- Western Regional Climate Center. Data archives, Station 046498 in Orick, California; period of record: May 1937 to December 2009. Available online: http://www.wrcc.dri.edu/cgi-bin/cliMAIN.pl?ca6498 (accessed on 5 May 2010).
- United States Department of Agriculture. Soil Survey of Redwood National and State Parks, California, 2008. USDA Natural Resources Conservation Service. Available online: http://soils.usda.gov/survey/printed_surveys/ (accessed on 4 March 2010).
- The Jepson Manual: Higher Plants of California; Hickman, J.C. (Ed.) University of California Press: Berkeley, CA, USA, 1993.
- Mueller-Duombois, D.; Ellenburg, H. Aims and Methods of Vegetation Ecology; John Wiley and Sons, Inc.: New York, NY, USA, 1974. [Google Scholar]
- Curtis, J.T.; McIntosh, R.P. An upland forest continuum in the prairie-forest border region of Wisconsin. Ecol. 1951, 32, 476–496. [Google Scholar] [CrossRef]
- Goebel, P.C.; Hix, D.M. Development of mixed-oak forests in southeastern Ohio: A comparison of second-growth and old-growth forests. For. Ecol. Manag. 1996, 84, 1–21. [Google Scholar] [CrossRef]
- Barker, J.R.; Ringold, P.L.; Bollman, M. Patterns of tree dominance in coniferous riparian forests. For. Ecol. Manag. 2002, 166, 311–329. [Google Scholar] [CrossRef]
- Zar, J.H. Biostatistical Analysis, 4th ed.; Prentice Hall, Inc.: Upper Saddle River, NJ, USA, 1999. [Google Scholar]
- Brower, J.E.; Zar, J.H.; von Ende, C.N. Field and Laboratory Methods for General Ecology, 3rd ed.; Wm. C. Brown Publishers: Dubuque, IA, USA, 1990. [Google Scholar]
- Muldavin, E.H.; Lenihan, J.M.; Lennox, W.S.; Veirs, S.D., Jr. Vegetation Succession in the First Ten Years Following Logging of Coast Redwood Forests; Technical Report No. 6; USDI National Park Service, Redwood National Park: Arcata, CA, USA, 1981. [Google Scholar]
- Lenihan, J.M. The Forest Associations of the Little Lost Man Creek Research Natural Area, Redwood National Park, CA. Master Thesis, Humboldt State University, Arcata, CA, USA, June 1986. [Google Scholar]
- Teraoka, J.R. Stand Response to Restoration Silviculture in a Second-Growth Redwood Stand, Redwood National and State Parks. Master Thesis, Humboldt State University, Arcata, CA, USA, December 2004. [Google Scholar]
- Chittick, A.J. Stand Structure and Development Following Thinning in a Second-Growth Forest, Redwood National Park. Master Thesis, Humboldt State University, Arcata, CA, USA, December 2005. [Google Scholar]
- Means, J.E.; Harris, R.R.; Sabin, T.E.; McCain, C.N. Spatial variation in productivity of Douglas-fir stands on a valley floor in the western Cascades range, Oregon. Northwest Sci. 1996, 70, 201–212. [Google Scholar]
- Pabst, R.J.; Spies, T.A. Structure and composition of unmanaged riparian forests in the coastal mountains of Oregon, U.S.A. Can. J. For. Res. 1999, 29, 1557–1573. [Google Scholar] [CrossRef]
- Nierenberg, T.R.; Hibbs, D.E. A characterization of unmanaged riparian areas in the central Coast Range of western Oregon. For. Ecol. Manag. 2000, 129, 195–206. [Google Scholar] [CrossRef]
- Stubblefield, G.; Oliver, C.D. Silvicultural Implications of the Reconstruction of Mixed Alder/Conifer Stands. In Utilization and Management of Alder: Proceedings of a Symposium; Briggs, D.G., DeBell, D.S., Atkinson, W.A., Eds.; General Technical Report PNW-GTR-70. USDA Forest Service Pacific Northwest Research Station: Portland, OR, USA, 1978; pp. 307–320. [Google Scholar]
- Franklin, J.F.; Spies, T.A.; Van Pelt, R.; Carey, A.B.; Thornburgh, D.A.; Berg, D.R.; Lindenmayer, D.B.; Harmon, M.E.; Keeton, W.S.; Shaw, D.C.; et al. Disturbances and structural development of natural forest ecosystems with silvicultural implications, using Douglas-fir forests as an example. For. Ecol. Manag. 2002, 155, 399–423. [Google Scholar] [CrossRef]
- Oliver, C.D. Forest development in North America following major disturbances. For. Ecol. Manag. 1981, 3, 153–168. [Google Scholar] [CrossRef]
- Klinka, K.; Chen, H.Y.H.; Wang, Q.; de Montigny, L. Forest canopies and their influence on understory vegetation in early-seral stands on West Vancouver Island. Northwest Sci. 1996, 70, 193–200. [Google Scholar]
- Mahoney, T.M.; Stuart, J.D. Old-Growth forest associations of the northern range of coastal redwood. Madroño 2000, 47, 53–60. [Google Scholar]
- Mahoney, T.M.; Stuart, J.D. Status of Vegetation Classification in Redwood Ecosystems; General Technical Report PSW-GTR-194; USDA Forest Service Pacific Southwest Research Station: Albany, CA, USA, 2007. [Google Scholar]
- Deal, R.L. Understory Plant Diversity in Riparian Alder-Conifer Stands after Logging in Southeast Alaska; Research Note PNW-RN-523; USDA Forest Service Pacific Northwest Research Station: Portland, OR, USA, 1997. [Google Scholar]
- Loya, D.T.; Jules, E.S. Use of species richness estimators improves evaluation of understory plant response to logging: A study of redwood forests. Plant Ecol. 2008, 194, 179–194. [Google Scholar] [CrossRef]
- Sawyer, J.O. Forests of northwestern California. In Terrestrial Vegetation of California, 3rd ed.; Barbour, M.G., Keeler-Wolf, T., Schoenherr, A.A., Eds.; University of California Press: Berkeley, CA, USA, 2007; pp. 253–295. [Google Scholar]
- O’Hara, K.L.; Leonard, L.P.; Keyes, C.R. Variable-Density thinning and a marking paradox: Comparing prescription protocols to attain stand variability in coast redwood. West. J. Appl. For. 2012, 27, 143–149. [Google Scholar] [CrossRef]
- Newton, M.; Cole, E.C. Stand development and successional implications: Pure and mixed stands. In The Biology and Management of Red Alder; Hibbs, D.E., Bell, D.S., Tarrant, R.F., Eds.; Oregon State University Press: Corvallis, OR, USA, 1994; pp. 106–115. [Google Scholar]
- Hibbs, D.E.; DeBell, D.S. Management of young red alder. In The Biology and Management of Red Alder; Hibbs, D.E., Bell, D.S., Tarrant, R.F., Eds.; Oregon State University Press: Corvallis, OR, USA, 1994; pp. 202–215. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Keyes, C.R.; Teraoka, E.K. Structure and Composition of Old-Growth and Unmanaged Second-Growth Riparian Forests at Redwood National Park, USA. Forests 2014, 5, 256-268. https://doi.org/10.3390/f5020256
Keyes CR, Teraoka EK. Structure and Composition of Old-Growth and Unmanaged Second-Growth Riparian Forests at Redwood National Park, USA. Forests. 2014; 5(2):256-268. https://doi.org/10.3390/f5020256
Chicago/Turabian StyleKeyes, Christopher R., and Emily K. Teraoka. 2014. "Structure and Composition of Old-Growth and Unmanaged Second-Growth Riparian Forests at Redwood National Park, USA" Forests 5, no. 2: 256-268. https://doi.org/10.3390/f5020256