3.1. Old-Growth Reference Conditions
The old-growth reference sites had much larger trees, lower densities, and different species composition than young even-aged second-growth stands that arose after clearcutting at HFR (
Table 1,
Table 2). Douglas-fir was the most abundant species in young stands, even after PCT to release and favor redwood. Among the three old-growth reference sites, redwood was the most abundant tree species at Governor’s and Elkhead Springs, and nearly rivaled tanoak density at Upper Salmon. Douglas-fir and tanoak were present in all three plots. Grand fir only occurred at Governor’s, and two California bay trees were found in the Upper Salmon plot. Redwood contributed the most towards SDI in all three plots, and was the only species abundant in both the understory and overstory. Mean dbh of conifers in the young stands was approximately one tenth of overstory tree dbh in old-growth reference plots. Young conifer stem densities were approximately 10 times the density of conifers in the old-growth overstory. At the three old-growth reference sites, density of all live stems ≥15 cm dbh ranged from 171 to 216 trees ha
−1. Basal area ranged between 146.9 m
2 ha
−1 at Elkhead Springs and 199.8 m
2 ha
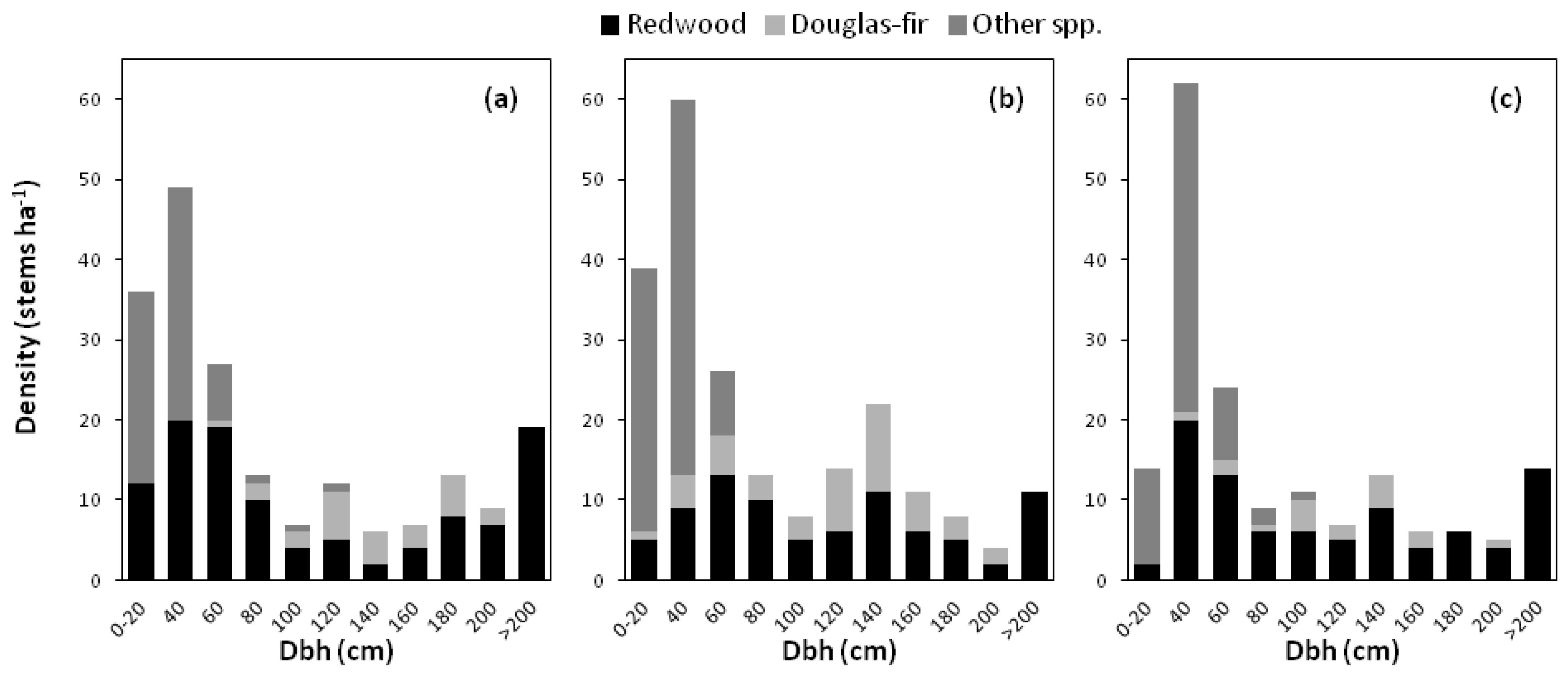
−1 at Governor’s. Total stand SDI values ranged from 1450 at Elkhead Springs, the driest inland location sampled, to 1907 at Governor’s. The maximum tree height did not exceed 84 m, and the tallest trees in each plot included both redwood and Douglas-fir, approximately 80 m tall in all cases. Diameter distributions for old-growth reference plots indicated that redwoods were present in all size classes (
Figure 2). At Elkhead Springs, the density of overstory Douglas-fir was lower than at the other two sites. Only redwood and Douglas-fir trees grew larger than 150 cm dbh. Trees labeled “other species” in the diameter distribution graphs were almost exclusively tanoak at Upper Salmon and Elkhead Springs, and a mix of tanoak and grand fir at Governor’s. These two species were the most abundant of all trees in the smallest size classes. Douglas-fir was poorly represented or nonexistent in the smaller size classes, likely due to high SDI. The thick litter layer and dense understory at HFR may also be impeding Douglas-fir regeneration. Douglas-fir is known to establish well on a seedbed of bare mineral soil and is less tolerant of shade than redwood, grand fir, and tanoak [
19].
Table 2.
Summary data for trees ≥15 cm dbh in three 1-ha plots in old-growth stands at Headwaters Forest Reserve, California. Data aggregated by crown class into understory (u, suppressed and intermediate) and overstory (o, dominant and codominant) groups. Height recorded for conifers only.
Table 2.
Summary data for trees ≥15 cm dbh in three 1-ha plots in old-growth stands at Headwaters Forest Reserve, California. Data aggregated by crown class into understory (u, suppressed and intermediate) and overstory (o, dominant and codominant) groups. Height recorded for conifers only.
| Site and species | Density (stems ha−1) | SDI | Mean Dbh (cm) | Max. Dbh (cm) | Crown Ratio | Max. Ht. (m) |
|---|
| u | o | u | o | u | o | | u | o | |
|---|
| Governor’s Grove | | | | | | | | | | |
| Redwood | 74 | 36 | 336 | 1108 | 55.4 | 207.9 | 305.3 | 0.52 | 0.59 | 79.7 |
| Douglas-fir | 4 | 21 | 34 | 336 | 88.9 | 138.8 | 192.0 | 0.39 | 0.47 | 80.0 |
| Grand fir | 44 | 1 | 67 | 9 | 29.9 | 101.0 | 101.0 | 0.39 | 0.53 | 63.7 |
| Tanoak | 18 | 0 | 16 | 0 | 22.8 | - | 36.0 | | | |
| Total | 140 | 58 | 453 | 1454 | 44.2 | 181.0 | | | | |
| Salmon Pass | | | | | | | | | | |
| Redwood | 51 | 32 | 345 | 750 | 71.3 | 172.4 | 350.2 | 0.51 | 0.54 | 77.1 |
| Douglas-fir | 16 | 29 | 68 | 438 | 57.2 | 134.2 | 191.3 | 0.50 | 0.56 | 81.0 |
| Tanoak | 86 | 0 | 86 | 0 | 24.1 | - | 58.6 | | | |
| California bay | 2 | 0 | 7 | 0 | 52.4 | - | 55.3 | | | |
| Total | 155 | 61 | 504 | 1188 | 43.4 | 154.2 | | | | |
| Elkhead Springs | | | | | | | | | | |
| Redwood | 55 | 34 | 248 | 920 | 58.7 | 190.6 | 347.5 | 0.47 | 0.62 | 84.1 |
| Douglas-fir | 3 | 14 | 9 | 171 | 49.2 | 116.4 | 192.1 | 0.36 | 0.52 | 74.8 |
| Tanoak | 65 | 0 | 102 | 0 | 31.3 | - | 84.6 | | | |
| Total | 123 | 48 | 359 | 1091 | 44.0 | 169.0 | | | | |
Figure 2.
Diameter distributions for all trees ≥15 cm dbh in (a) Governor’s (b) Upper Salmon and (c) Elkhead Springs old-growth reference plots at Headwaters Forest Reserve, California.
Figure 2.
Diameter distributions for all trees ≥15 cm dbh in (a) Governor’s (b) Upper Salmon and (c) Elkhead Springs old-growth reference plots at Headwaters Forest Reserve, California.
Basal stem cavities, reiterations, and large snags are features indicative of old-growth forests [
20]. The incidence of these features was greatest at Governor’s and least at Elkhead Springs (
Table 3). The majority of basal stem cavities and all reiterations were found on redwood trees. Most snags at Governor’s were grand fir. Grand fir was not found in the overstory, suggesting that under the current stand conditions it was able to grow beneath the canopy but did not survive long enough to attain upper canopy status, giving rise to numerous small snags. The ratio of redwood to Douglas-fir snags was ≥1:1, and the ratio of small to large snags (excluding grand fir) was approximately 2:1. Restoration of large snags first requires large trees be grown. In old-growth stands on alluvial flats, larger redwood trees were most likely to have basal stem cavities and reiterations [
5]. However, to restore these features, restoration treatments accelerating tree-size development will need to be accompanied by disturbances such as fire and storms or surrogate treatments that damage stems and tree tops.
Table 3.
Number of large snags (≥60 cm dbh, ≥10 m tall), small snags, and trees with basal stem cavities and reiterations in three 1-ha old-growth reference plots at Headwaters Forest Reserve, California. Percent redwood indicates how much of each tally was represented by redwood trees.
Table 3.
Number of large snags (≥60 cm dbh, ≥10 m tall), small snags, and trees with basal stem cavities and reiterations in three 1-ha old-growth reference plots at Headwaters Forest Reserve, California. Percent redwood indicates how much of each tally was represented by redwood trees.
| Governor’s | Upper Salmon | Elkhead Springs |
|---|
| Count | % Redwood | Count | % Redwood | Count | % Redwood |
|---|
| Large snags | 3 | 100 | 2 | 50 | 7 | 57 |
| Small snags | 18 | 17 | 6 | 50 | 9 | 56 |
| Basal stem cavities | 14 | 93 | 11 | 100 | 3 | 100 |
| Reiterations | 5 | 100 | 4 | 100 | 2 | 100 |
Comparing our reference conditions for redwood forests on upland terrain at HFR against alluvial flat redwood forest data from a similar study [
4,
5] revealed that SDI on alluvial flats was approximately 26% higher on average than SDI at HFR, and alluvial flat stand basal area was approximately 48% higher on average. The tallest redwoods were 26% taller in alluvial flat reference plots. Our upland redwood plots had many more snags, likely due to the presence of species other than redwood which are not as long lived. Reiterations were five times more frequent at the alluvial flat sites. Total stem density was higher on upland sites while the two forest types had similar overstory densities. At HFR, the overstory trees were smaller in dbh and height than alluvial flat overstory trees, resulting in a more open forest canopy that intercepted less light and presumably allowed for a higher density of understory trees.
3.2. Tree Growth in Harvested Stands
In total, 330 second-growth and remnant old-growth trees were sampled for dbh growth: 174 redwood and 156 Douglas-fir, encompassing a range of tree sizes (
Table 4). On average, the two species had similar growth rates. However, Douglas-fir had lower mean values for SDI and cSDI than redwood. The spread of SDI and cSDI data indicated that our sampling covered a wide range of stand densities, including areas with widely-spaced trees and dense, crowded areas.
Table 4.
Summary data for trees sampled for recent growth at Headwaters Forest Reserve, California. Growth was average annual diameter increment (DBHI) and basal area increment (BAI) over the most recent five years. Redwood ratio was ratio of number of redwood trees to other trees in plot. Competitive stand density index (cSDI) was sum of per-hectare stand density index (SDI) for trees with crown class above or the same as sampled tree, including SDI of sample tree.
Table 4.
Summary data for trees sampled for recent growth at Headwaters Forest Reserve, California. Growth was average annual diameter increment (DBHI) and basal area increment (BAI) over the most recent five years. Redwood ratio was ratio of number of redwood trees to other trees in plot. Competitive stand density index (cSDI) was sum of per-hectare stand density index (SDI) for trees with crown class above or the same as sampled tree, including SDI of sample tree.
| Variable | Mean | Std. Dev. | Min. | Max. |
|---|
| Redwood (n = 174) | | | | |
| Dbh (cm) | 47.17 | 44.23 | 10.20 | 283.10 |
| Height (m) | 21.96 | 14.29 | 4.39 | 72.36 |
| Crown ratio | 0.59 | 0.14 | 0.15 | 0.88 |
| DBHI (cm year−1) | 0.78 | 0.40 | 0.08 | 1.64 |
| BAI (cm2 year−1) | 53.66 | 40.63 | 1.47 | 159.91 |
| Redwood ratio (stems per plot) | 0.76 | 0.20 | 0.25 | 1.00 |
| SDI | 940.52 | 463.09 | 175.19 | 2161.29 |
| cSDI | 683.57 | 415.32 | 119.35 | 2123.17 |
| Douglas-fir (n = 156) | | | | |
| Dbh (cm) | 47.26 | 34.21 | 11.20 | 177.00 |
| Height (m) | 26.18 | 14.62 | 7.07 | 75.11 |
| Crown ratio | 0.58 | 0.20 | 0.12 | 0.92 |
| DBHI (cm year−1) | 0.77 | 0.40 | 0.08 | 1.66 |
| BAI (cm2 year−1) | 54.10 | 36.30 | 1.45 | 134.70 |
| Redwood ratio (stems per plot) | 0.44 | 0.27 | 0.00 | 0.89 |
| SDI | 800.00 | 368.72 | 196.36 | 1951.47 |
| cSDI | 568.67 | 323.21 | 123.11 | 1951.47 |
Regression analysis indicated that cSDI was a stronger predictor of tree dbh growth than SDI, and model fit improved further when the sine-transformed cSDI index replaced cSDI (
i.e., AIC values improved from 83.7 to 20.4 to 19.9 for redwood, and from 56.0 to 19.1 to 13.5 for Douglas-fir). This suggested that growth of a sampled tree was less affected by trees in subordinate crown classes, consistent with higher growth efficiencies reported for redwood trees of superior crown class and canopy strata [
21]. Due to convergence problems, we fitted the DBHI models while iteratively varying the “d” coefficient until AIC was minimized. This gave “d” parameter estimates of 52 for redwood and 200 for Douglas-fir. All other DBHI model coefficients and fit statistics are listed in
Table 5.
Table 5.
Inverse polynomial diameter increment model coefficients and fit statistics for redwood and Douglas-fir trees sampled for growth at Headwaters Forest Reserve, California.
Table 5.
Inverse polynomial diameter increment model coefficients and fit statistics for redwood and Douglas-fir trees sampled for growth at Headwaters Forest Reserve, California.
| Parameter | Estimate | Std. Error | t value | Pr (>|t|) |
|---|
| Redwood (n = 174) | a | 110.223 | 27.931 | 3.946 | 0.000 |
| b | −1.665 | 0.579 | −2.874 | 0.005 |
| c | 0.012 | 0.003 | 4.039 | 0.000 |
| e | 0.605 | 0.124 | 4.879 | 0.000 |
| Douglas-fir (n = 156) | a | 1850.000 | 585.600 | 3.159 | 0.002 |
| b | −14.740 | 4.758 | −3.098 | 0.002 |
| c | 0.033 | 0.097 | 3.349 | 0.001 |
| e | 0.304 | 0.107 | 2.851 | 0.005 |
| f | 0.650 | 0.106 | 6.123 | 0.000 |
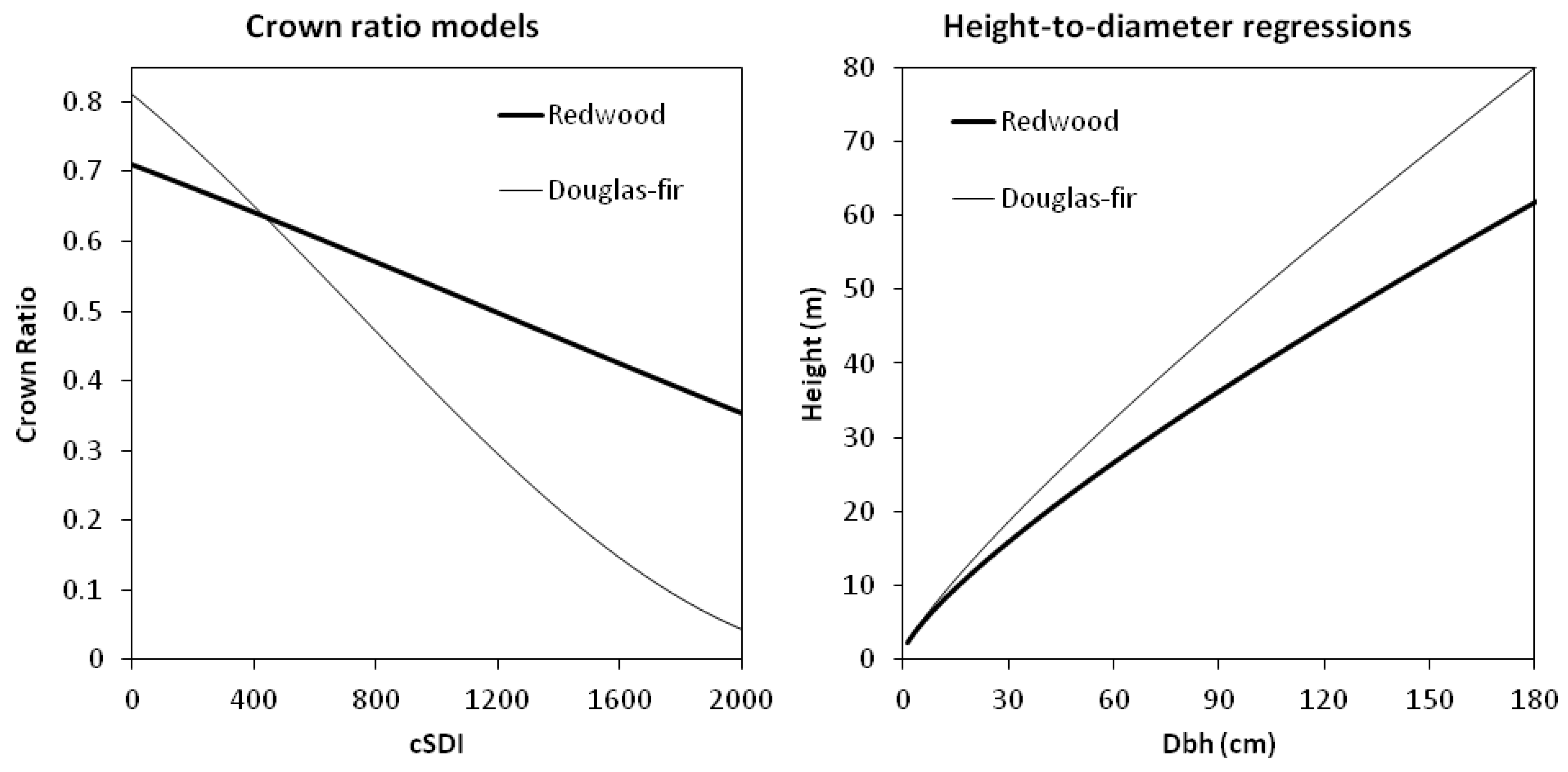
Model predictions indicated that mean dbh increment peaked between 35 and 45 cm dbh for redwood and Douglas-fir (
Figure 3). Rate of growth was generally higher for redwood, except for small Douglas-fir trees at low stand densities (cSDI). Douglas-fir dbh growth was more sensitive to competition (in terms of cSDI) than redwood.
Figure 3.
Relationship between expected (average) annual dbh increment (cm year−1) and dbh (cm) in redwood and Douglas-fir at Headwaters Forest Reserve, California, as a function of competitive SDI (cSDI) and crown ratio predicted from cSDI.
Figure 3.
Relationship between expected (average) annual dbh increment (cm year−1) and dbh (cm) in redwood and Douglas-fir at Headwaters Forest Reserve, California, as a function of competitive SDI (cSDI) and crown ratio predicted from cSDI.
On average, redwood trees maintained longer live crowns than Douglas-fir at higher cSDI (
Figure 4). Presumably this result reflected the greater shade tolerance of redwood. Height-to-diameter relationships indicated that Douglas-fir trees were taller than redwoods of similar dbh.
Figure 4.
Relationships between crown ratio and competitive SDI (cSDI) for redwood: CRrw = sin(1.002 − 0.0001823cSDI)2; and Douglas-fir: CRdf = sin(1.121 − 0.0004553cSDI)2 (left panel); and height-dbh relationship for redwood: HT = 1.37 + e(0.7899lnDBH); and Douglas-fir: HT = 1.37 + e(0.8407lnDBH) (right panel); for n = 330 trees sampled for growth at Headwaters Forest Reserve, California.
Figure 4.
Relationships between crown ratio and competitive SDI (cSDI) for redwood: CRrw = sin(1.002 − 0.0001823cSDI)2; and Douglas-fir: CRdf = sin(1.121 − 0.0004553cSDI)2 (left panel); and height-dbh relationship for redwood: HT = 1.37 + e(0.7899lnDBH); and Douglas-fir: HT = 1.37 + e(0.8407lnDBH) (right panel); for n = 330 trees sampled for growth at Headwaters Forest Reserve, California.
3.3. Simulating Restoration Prescriptions
Based on the range of values found in our old-growth reference plots, we specified “restoration goals” of attaining:
- (i)
Overstory tree size and density: 32–36 redwood trees ha−1 averaging 170 cm dbh minimum, and 14–29 Douglas-fir trees ha−1 averaging 120 cm dbh minimum;
- (ii)
Understory tree size and density: 51–74 redwood trees ha−1 with a mean of 56 cm dbh, and 3–16 Douglas-fir trees ha−1 with a mean dbh of 50 cm;
- (iii)
Crown ratio: 54%–62% for redwood and 47%–56% for Douglas-fir overstory trees (
Table 2); and;
- (iv)
Snag densities: 2–7 large snags ha
−1 (
Table 3).
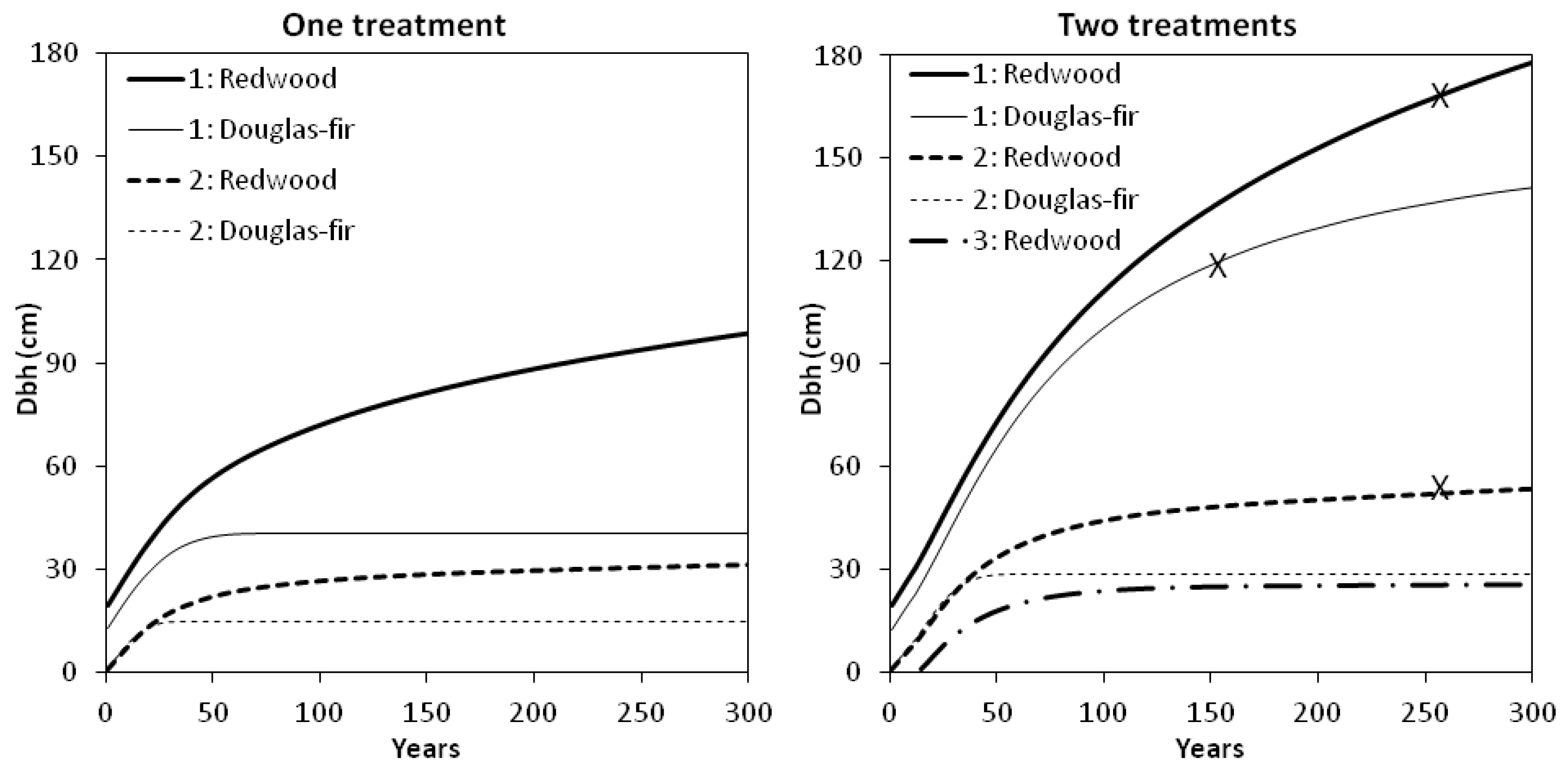
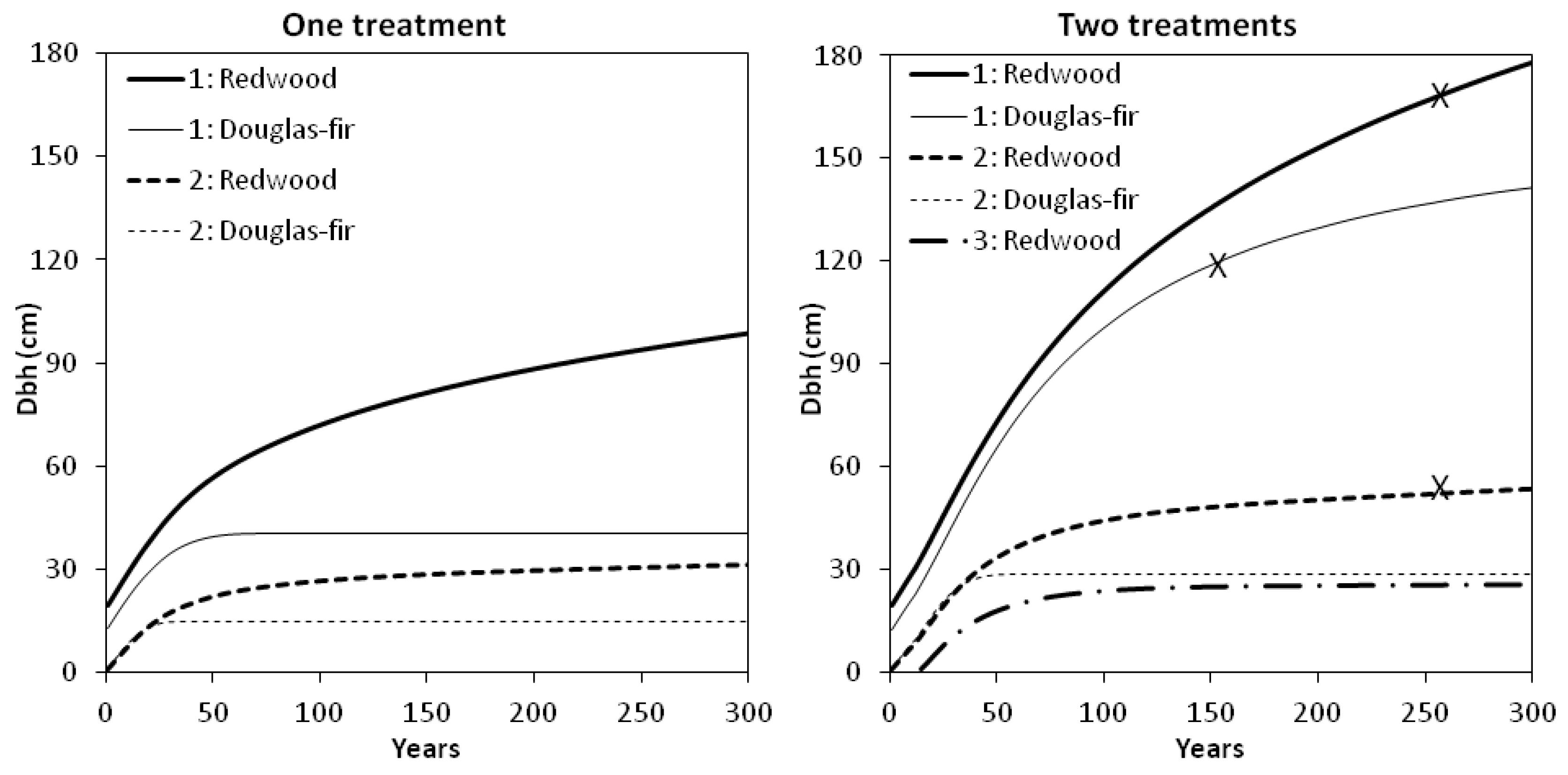
Our first simulation was a “one treatment” restoration prescription (PCT only), without any subsequent intervention or disturbance. Model predictions indicated that tree growth would proceed slowly (
Figure 5). Overstory redwood had not attained a mean dbh of 100 cm after 300 years. Stand density exceeded 1480 (the maximum SDI for Douglas-fir [
12]) by year 70, at which time the average overstory Douglas-fir had reached 41 cm dbh. Our models did not predict mortality explicitly, but we speculated that density-dependent mortality among understory and smaller overstory Douglas-fir could result in a gradual increase in average tree dbh among fewer, larger surviving Douglas-fir. Under this scenario, crown ratio fell below the reference condition and minimum tree-size and density thresholds were not attained within 300 years.
Figure 5.
Simulated diameter development of redwood and Douglas-fir in two or three cohorts (overstory cohort, “1”; second cohort, “2”; third cohort, “3”) under two different scenarios. Left panel: “One treatment” with 465 overstory stems ha−1 remaining after one precommercial thinning (PCT) (before time zero). Right panel: “Two treatments” including one PCT (before time zero) followed by a second stand entry (partial cutting to leave 70 stems ha−1 in the overstory) before the average overstory redwood reached 30 cm dbh. Restoration goals for mean dbh denoted by “x”.
Figure 5.
Simulated diameter development of redwood and Douglas-fir in two or three cohorts (overstory cohort, “1”; second cohort, “2”; third cohort, “3”) under two different scenarios. Left panel: “One treatment” with 465 overstory stems ha−1 remaining after one precommercial thinning (PCT) (before time zero). Right panel: “Two treatments” including one PCT (before time zero) followed by a second stand entry (partial cutting to leave 70 stems ha−1 in the overstory) before the average overstory redwood reached 30 cm dbh. Restoration goals for mean dbh denoted by “x”.
In a second scenario, we simulated a second stand entry after 12 years, before the average redwood reached 30 cm dbh (
Figure 5). Partial cutting left 40 redwood trees and 30 Douglas-fir trees ha
−1 in the overstory. To re-create snag reference conditions, these densities included an allowance for natural mortality or snag creation at the rate of two overstory snags ha
−1 and four understory snags ha
−1 (equal numbers of Douglas-fir and redwood) every 50 years. Under this “two treatment” restoration prescription (PCT + second entry) our models predicted rapid growth of overstory conifers towards our old-growth reference conditions. Overstory redwood reached a mean dbh of 100 cm at year 83, and Douglas-fir attained the same mean diameter at year 99. After 154 years, the remaining overstory Douglas-fir (27 trees ha
−1) reached 120 cm average dbh and had 47% crown ratio, the minimum thresholds based on our reference condition. At year 261, 35 overstory redwood trees reached our tree-size goal of 170 cm mean dbh. Stand density index was approaching the maximum for redwood, higher than the reference condition and indicative of crowding. However, overstory redwood crown ratio was predicted to remain within the range of values sampled in old-growth stands at HFR. At this time in the simulation, redwood trees in the midstory (53 cm mean dbh) had almost attained the 56 cm threshold.
Neither treatment scenario depicted in
Figure 5 predicted restoration of the understory Douglas-fir reference condition. Under both scenarios, predicted dbh increment of understory Douglas-fir ceased once stand density climbed above 1480 SDI. Unknown was how long this cohort would survive, and how density-dependent mortality would alter average dbh over time. In our reference condition plots, understory Douglas-fir were rare but present. Managing for Douglas-fir regeneration and additional recruitment into the overstory will likely require some form of disturbance, such as the reintroduction of fire or partial harvesting to alter forest floor and light conditions and make growing space available. Identifying Douglas-fir trees furthest from overstory redwood trees, and reducing stand density in their immediate vicinity should relieve crowding. Theoretically this modification to the spatial pattern of tree locations would result in allocation of more growing space to Douglas-fir and less to other stand components. Our second simulation appeared to make enough growing space available for overstory Douglas-fir while meeting the 30 cm dbh limit for cutting and slowing the process of (predicted) crown rise. Unknown is whether such heavy early cutting would encourage development of large, persistent lower branches in the oldest cohort. Raising the 30 cm dbh limit for cutting specified under the current management plan [
15] could allow deferral of cutting until crown rise had proceeded and the overstory had lower stems free of live branches.
Our models only provide estimates indicative of tree growth and stand development. They have a wide range of applicability in terms of tree size and stand density. However, predictions should only be regarded as “hypotheses for future testing” until DBHI models are validated with independent data. Incorporating models of tanoak growth, conifer and hardwood mortality, dbh distributions, and height growth would improve predictions of tree growth and stand structure. We recommend field-testing the restoration treatments presented and some alternative treatments (different densities and timing of cutting) and monitoring subsequent tree growth, regeneration, and mortality for future model validation and to inform adaptive management at HFR.


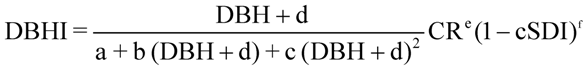

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}