Classification of Needle Loss of Individual Scots Pine Trees by Means of Airborne Laser Scanning


 ,
, 
Abstract
:1. Introduction
2. Material and Methods
2.1. Study Area

2.2. Airborne Laser Scanning Data Set
2.3. Reference Data
2.3.1. Field Measurements
2.3.2. Tree Detection and Linking of ALS and Field Data

{kind=link}
{kind=link}
{kind=link}
| (a) | (b) | |||||
|---|---|---|---|---|---|---|
| min | max | mean | sd | Defoliation (%) | Number of trees | |
| dbh (cm) | 53 | 405 | 222 | 13 | 0 | 43 |
| h (m) | 8.6 | 26.2 | 18.8 | 3.1 | 10 | 222 |
| 20 | 266 | |||||
| 30 | 115 | |||||
| 40 | 36 | |||||
| 50–100 | 19 | |||||
| Total | 701 | |||||
2.3.3. Classification Schemes for Defoliation
| Classification | Threshold defoliation levels | Classes (n) |
|---|---|---|
| DEF1 | 20% | 2 |
| DEF2 | 30% | 2 |
| DEF3 | 30%, 60% | 3 |
| DEF4 | 20%, 50% | 3 |
| DEF5 | 20%, 30%, 40% | 4 |
2.4. ALS Feature Extraction
| Feature | Description |
|---|---|
| Hmax | Maximum height of laser returns |
| Hmean | Arithmetic mean of laser heights |
| Hstd | Standard deviation of heights |
| CV | Hstd divided by Hmean |
| h10–h90 | Heights 0th–90th percentile |
| p10–p90 | Percentile of canopy height distribution |
| pene | Penetration calculated as a proportion of returns below 2 m to total returns |
| Int | Mean intensity |
2.5. Estimation of Defoliation
2.6. Simulation of Pulse Densities
3. Results
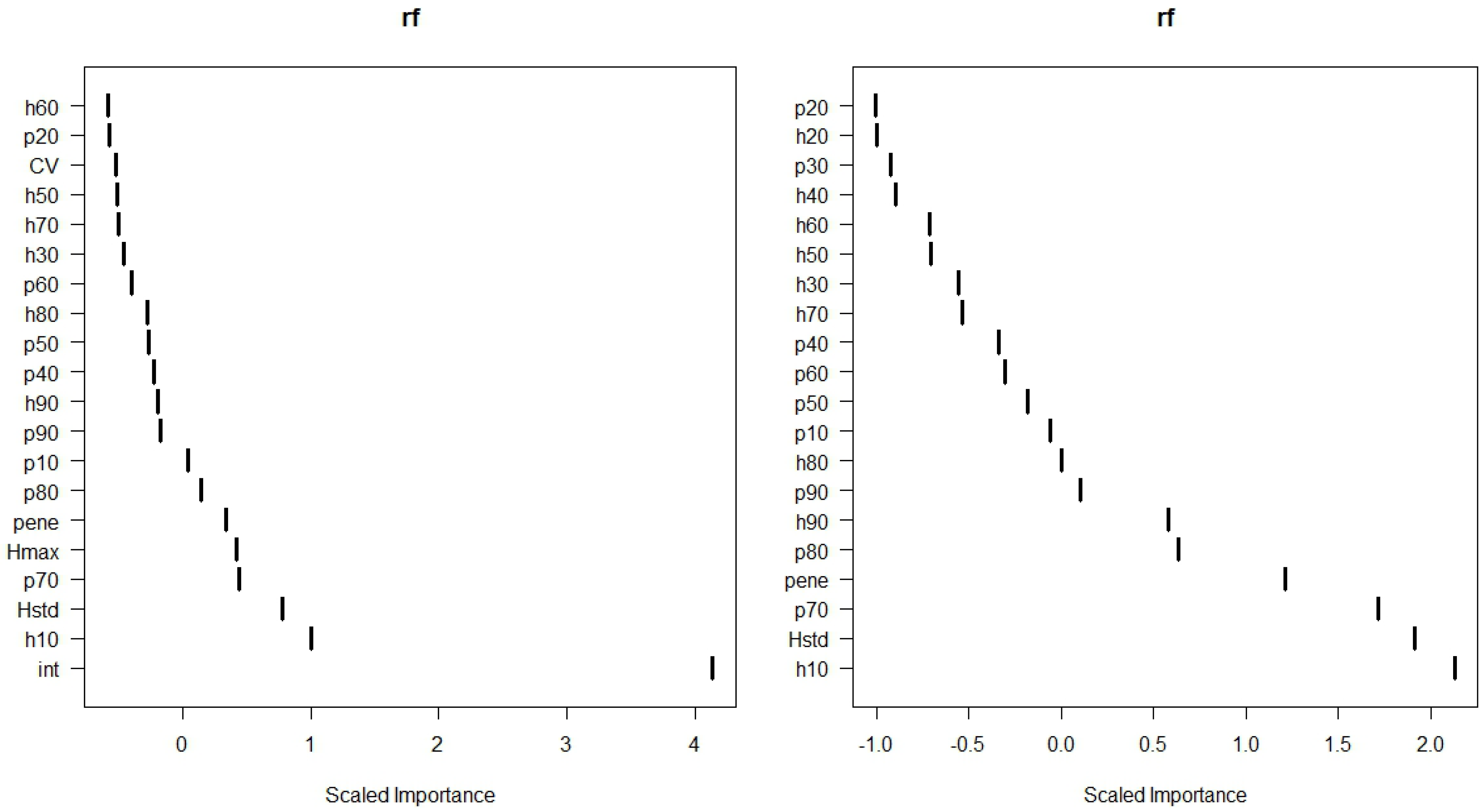
3.1. Classification of Defoliation
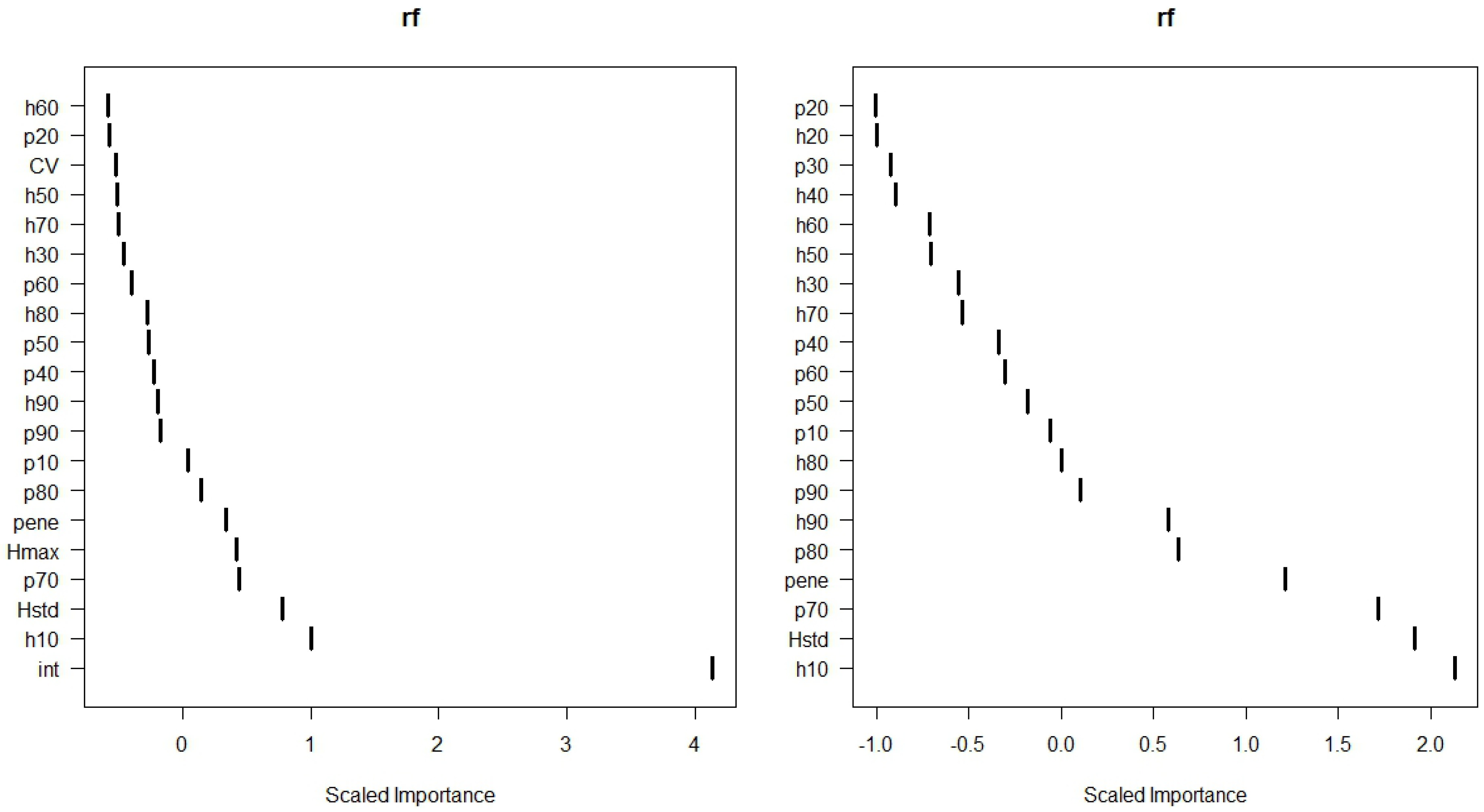
| Correlations | Mean values | t-Test | ||||
|---|---|---|---|---|---|---|
| Feature | h10 | Hstd | p70 | Healthy | Defoliated | p-Value |
| h10 | 1.00 | −0.34 | −0.32 | 0.1802 | 0.9657 | <0.000 |
| Hstd | −0.34 | 1.00 | −0.24 | 6.0684 | 5.0588 | <0.000 |
| p70 | −0.32 | −0.24 | 1.00 | 0.5306 | 0.5153 | 0.19 |
| Classification | Threshold defoliation levels | Classes (n) | CA (%) | Kappa-Value | CAmin (%) | CAmax (%) | CAstd (%) |
|---|---|---|---|---|---|---|---|
| DEF1 | 20% | 2 | 82.9 | 0.63 | 81.1 | 84.8 | 1.4 |
| DEF2 | 30% | 2 | 86.5 | 0.57 | 85.3 | 87.2 | 6.1 |
| DEF3 | 30%, 60% | 3 | 85.4 | 0.53 | 84.9 | 86.3 | 4.6 |
| DEF4 | 20%, 50% | 3 | 81.5 | 0.61 | 79.5 | 81.9 | 7.1 |
| DEF5 | 20%, 30%, 40% | 4 | 71.0 | 0.56 | 68.6 | 72.32 | 10.1 |
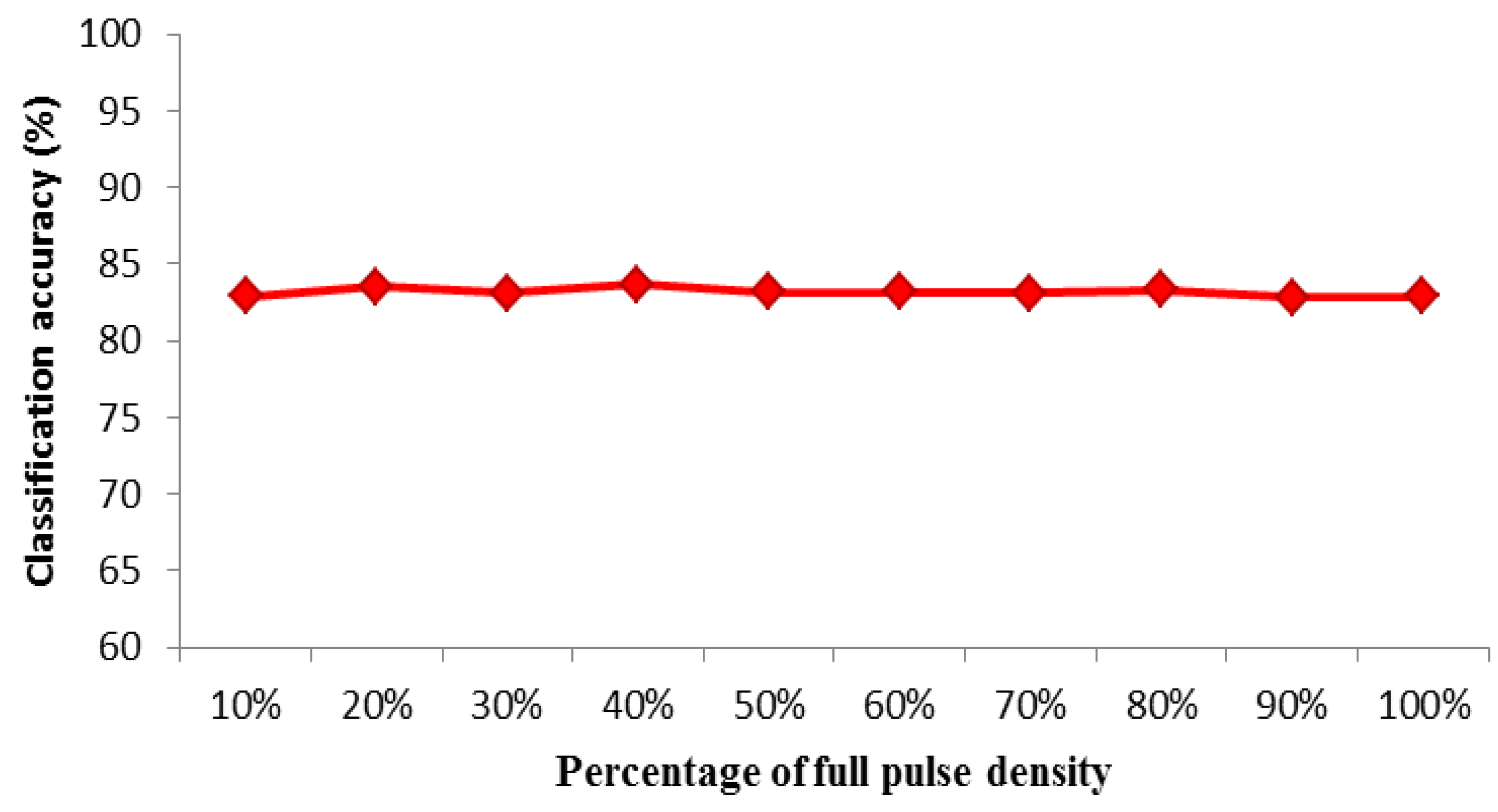
3.2. Effect of Simulated ALS Pulse Density
| % of full pulse density | Pulse density (appr.) | CA (%) | Kappa-Value | CAmin (%) | CAmax (%) | CAstd (%) |
|---|---|---|---|---|---|---|
| 10% | 20 | 82.9 | 0.63 | 80.5 | 84.7 | 1.4 |
| 20% | 18 | 83.5 | 0.64 | 80.6 | 84.9 | 1.1 |
| 30% | 16 | 83.1 | 0.63 | 81.1 | 85.0 | 0.9 |
| 40% | 14 | 83.7 | 0.64 | 82.8 | 84.7 | 1.4 |
| 50% | 12 | 83.2 | 0.63 | 80.7 | 85.2 | 1.5 |
| 60% | 10 | 83.2 | 0.63 | 81.5 | 85.7 | 1.4 |
| 70% | 8 | 83.1 | 0.63 | 81.5 | 84.8 | 0.6 |
| 80% | 6 | 83.3 | 0.64 | 81.5 | 84.7 | 1.3 |
| 90% | 4 | 82.8 | 0.62 | 81.6 | 84.6 | 1.3 |
| 100% | 2 | 82.9 | 0.63 | 81.1 | 84.8 | 1.5 |
4. Discussion
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Attiwill, P.M. The disturbances of forest ecosystems: The ecological basis for conservative management. For. Ecol. Manag. 1994, 63, 247–300. [Google Scholar] [CrossRef]
- Linke, J.; Betts, M.G.; Lavigne, M.B.; Franklin, S.E. Introduction: Structure, Function, and Change of Forest Landscapes. In Understanding Forest Disturbance and Spatial Pattern; Wulder, M.A., Franklin, S.E., Eds.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2007; pp. 1–29. [Google Scholar]
- Dale, V.H.; Joyce, L.A.; McNulty, S.; Neilson, R.P.; Ayres, M.P.; Flannican, M.D.; Hanson, P.J.; Irland, L.C.; Lugo, A.E.; Peterson, C.J.; et al. Climate change and forest disturbances. Bioscience 2001, 51, 723–734. [Google Scholar] [CrossRef]
- Fleming, R.A.; Candau, J.; McAlpine, R. Landscape scale analysis of interactions between insect defoliation and forest fire in central Canada. Clim. Chang 2002, 55, 251–272. [Google Scholar] [CrossRef]
- Lyytikäinen-Saarenmaa, P.; Tomppo, E. Impact of sawfly defoliation on growth of Scots pine Pinus sylvestris (Pinaceae) and associated economic losses. Bull. Entomol. Res. 2002, 92, 137–140. [Google Scholar]
- Lyytikäinen-Saarenmaa, P.; Niemelä, P.; Annila, E. Growth Responses and Mortality of Scots Pine (Pinus sylvestris L.) after a Pine Sawfly Outbreak. In Proceedings of Forest Insect Population Dynamics and Host Influences, International Symposium of IUFRO, Kanazawa, Japan, 14–19 September 2003; Liebhold, A.L., Quiring, D.T., Clancy, K.M., Eds.; 2006; pp. 81–85. [Google Scholar]
- IPCC, Summary for Policymakers. In Climate Change 2007: The Physical Science basis; Solomon, S.; Qin, D.; Manning, M.; Chen, Z.; Marquis, M.; Avyret, K.B.; Tignor, M.; Miller, H.L. (Eds.) Cambridge University Press: Cambridge, UK, 2007.
- Netherer, S.; Schopf, A. Potential effects of climate change on insect herbivores in European forests—General aspects and the pine processionary moth as specific example. For. Ecol. Manag. 2010, 259, 831–838. [Google Scholar] [CrossRef]
- Seidl, R.; Schelaas, M.; Lexar, M. Unraveling the drivers of intensifying forest disturbance regimes in Europe. Glob. Chang Biol. 2011, 17, 2842–2852. [Google Scholar] [CrossRef]
- Volney, W.J.A.; Fleming, R.A. Climate change and impacts of boreal forest insects. Agr. Ecosyst. Environ. 2000, 82, 283–294. [Google Scholar] [CrossRef]
- Walther, G.R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.C.; Fromentin, J.-M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar]
- Björkman, C.; Bylund, H.; Klapwijk, M.J.; Kollberg, I.; Schroeder, M. Insect pests in future forests: More severe problems? Forests 2011, 2, 474–485. [Google Scholar] [CrossRef]
- Bale, J.S.; Masters, G.J.; Hodkinson, I.D.; Awmack, C.; Bezemer, T.M.; Brown, V.K.; Butterfield, J.; Buse, A.; Coulson, J.C.; Farrar, J.; et al. Herbivory in global climate change research: Direct effects of rising temperatures on insect herbivores. Glob. Chang Biol. 2002, 8, 1–16. [Google Scholar] [CrossRef]
- Logan, J.A.; Regniere, J.; Powell, J.A. Assessing the impacts of global warming on forest pest dynamics. Front. Ecol. Environ. 2003, 1, 130–137. [Google Scholar] [CrossRef]
- Régniére, J. Predicting insect continental distributions from species physiology. Unasylva 2009, 60, 37–42. [Google Scholar]
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Garcia-Gonzalo, J.; Seidl, R.; Delzon, S.; Corona, P.; Kolström, M.; et al. Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Battisti, A.; Stastny, M.; Buffo, E.; Larsson, S. A rapid altitudinal range expansion in the pine processionary moth produced by the 2003 climatic anomaly. Glob. Chang Biol. 2006, 12, 662–671. [Google Scholar]
- Vanhanen, H.; Veteli, T.O.; Päivinen, S.; Kellomäki, S.; Niemelä, P. Climate change and range shifts in two insect defoliators: Gypsy moth and nun moth—A model study. Silva Fenn. 2007, 41, 621–638. [Google Scholar]
- Hlásny, T.; Zajičková,, L.; Turčáni, M.; Holuša, J.; Sitková, Z. Gepgraphical variability of Spruce bark beetle development under climate change in the Czech Republic. J. For. Sci. 2011, 57, 242–249. [Google Scholar]
- De Somviele, B.; Lyytikäinen-Saarenmaa, P.; Niemelä, P. Stand edge effects on distribution and condition of Diprionid sawflies. Agr. For. Entomol. 2007, 9, 17–30. [Google Scholar]
- Lyytikäinen-Saarenmaa, P.; Holopainen, M.; Ilvesniemi, S.; Haapanen, R. Detecting pine sawfly defoliation by means of remote sensing and GIS. Forstsch. Aktuell. 2008, 44, 14–15. [Google Scholar]
- Juutinen, P.; Varama, M. Ruskean mäntypistiäisen (Neodiprion sertifer) esiintyminen Suomessa vuosina 1966–83. Folia For. 1986, 662, 1–39. [Google Scholar]
- Tomppo, E. The Finnish National Forest Inventory. In Forest Inventory; Kangas, A., Maltamo, M., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 179–194. [Google Scholar]
- Hall, R.J.; Skakun, R.S.; Arsenault, E.J. Remotely Sensed Data in the Mapping of Insect Defoliation. In Understanding Forest Disturbance and Spatial Pattern. Remote Sensing and GIS Approaches; Wulder, M.A., Franklin, S.E., Eds.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2007; pp. 85–111. [Google Scholar]
- Magnussen, S.; Eggermont, P.; LaRiccia, V.N. Recovering tree heights from airborne laser scanner data. For. Sci. 1999, 45, 407–422. [Google Scholar]
- Maltamo, M.; Mustonen, K.; Hyyppä, J.; Pitkänen, J.; Yu, X. The accuracy of estimating individual tree variables with airborne laser scanning in boreal nature reserves. Can. J. For. Res. 2004, 34, 1791–1801. [Google Scholar] [CrossRef]
- Falkowski, M.J.; Smith, A.M.S.; Hudak, A.T.; Gessler, P.E.; Vierling, L.A.; Crookston, N.L. Automated estimation of individual conifer tree height and crown diameter via two-dimensional spatial wavelet analysis of lidar data. Can. J. Remote Sens. 2006, 32, 153–161. [Google Scholar] [CrossRef]
- Bortolot, Z.; Wynne, R.H. Estimating forest biomass using small footprint LiDAR data: An individual tree-based approach that incorporates training data. ISPRS J. Photogramm. Remote Sens. 2005, 59, 342–360. [Google Scholar] [CrossRef]
- Van Aardt, J.A.N.; Wynne, R.H.; Scrivani, J.A. Lidar-based mapping of forest volume and biomass by taxonomic group using structurally homogenous segments. Photogramm. Eng. Remote Sens. 2008, 74, 1033–1044. [Google Scholar]
- Korpela, I.; Ørka, H.O.; Maltamo, M.; Tokola, T.; Hyyppä, J. Tree species classification using airborne LiDAR—Effects of stand and tree parameters, downsizing of training set, intensity normalization, and sensor type. Silva Fenn. 2010, 44, 319–339. [Google Scholar]
- Hyyppä, J.; Kelle, O.; Lehikoinen, M.; Inkinen, M. A segmentation-based method to retrieve stem volume estimates from 3-D tree height models produced by laser scanners. Geosci. Remote Sens. 2001, 39, 969–975. [Google Scholar] [CrossRef]
- Wallerman, J.; Holmgren, J. Estimating field-plot data of forest stands using airborne laser scanning and SPOT HRG data. Remote Sens. Environ. 2007, 110, 501–508. [Google Scholar]
- Means, J.E.; Acker, S.A.; Fitt, B.J.; Renslow, M.; Emerson, L.; Hendrix, C.J. Predicting forest stand characteristics with airborne scanning ALS. Photogramm. Eng. Remote Sens. 2000, 66, 1367–1371. [Google Scholar]
- Næsset, E. Predicting forest stand characteristics with airborne scanning laser using a practical two-stage procedure and field data. Remote Sens. Environ. 2002, 80, 88–99. [Google Scholar] [CrossRef]
- Holmgren, J.; Persson, A. Identifying species of individual trees using airborne laser scanner. Remote Sens. Environ. 2004, 90, 415–423. [Google Scholar] [CrossRef]
- Brandtberg, T. Classifying individual tree species under leaf-off and leaf-on conditions using airborne lidar. ISPRS J. Photogramm. Remote Sens. 2007, 61, 325–340. [Google Scholar] [CrossRef]
- Holopainen, M.; Vastaranta, M.; Rasinmäki, J.; Kalliovirta, J.; Mäkinen, A.; Haapanen, R.; Melkas, T.; Yu, X.; Hyyppä, J. Uncertainty in timber assortment estimates predicted from forest inventory data. Eur. J. For. Res. 2010, 129, 1131–1142. [Google Scholar] [CrossRef]
- Vastaranta, M.; Holopainen, M.; Yu, X.; Hyyppä, J.; Hyyppä, H. Predicting stand-thinning maturity from airborne laser scanning data. Scand. J. For. Res. 2010, 26, 187–196. [Google Scholar]
- Kantola, T.; Vastaranta, M.; Yu, X.; Lyytikäinen-Saarenmaa, P.; Holopainen, M.; Talvitie, M.; Kaasalainen, S.; Solberg, S.; Hyyppä, J. Classification of defoliated trees using tree-level airborne laser scanning data combined with aerial images. Remote Sens. 2010, 2, 2665–2679. [Google Scholar] [CrossRef]
- Vastaranta, M.; Korpela, I.; Uotila, A.; Hovi, A.; Holopainen, M. Mapping of snow-damaged trees in bi-temporal airborne LiDAR data. Eur. J. For. Res. 2012, 131, 1217–1228. [Google Scholar] [CrossRef]
- Vastaranta, M.; Holopainen, M.; Yu, X.; Haapanen, R.; Melkas, T.; Hyyppä, J.; Hyyppä, H. Individual tree detection and area-based approach in retrieval of forest inventory characteristics from low-pulse airborne laser scanning data. Photogramm. J. Fin. 2011, 22, 1–13. [Google Scholar]
- Sohlberg, S.; Næsset, E.; Hanssen, K.H.; Christiansen, E. Mapping defoliation during a severe insect attack on Scots pine using airborne laser scanning. Remote Sens. Environ. 2006, 102, 364–376. [Google Scholar] [CrossRef]
- Hawbaker, T.J.; Keuler, N.S.; Lesak, A.A.; Gobakken, T.; Contrucci, K.; Radeloff, V.C. Improved estimates of forest vegetation structure and biomass with a ALS-optimized sampling design. J. Geophys. Res. Lett. 2009, 114. [Google Scholar] [CrossRef]
- Zhao, K.; Popescu, S.; Nelson, R. ALS remote sensing of forest biomass: A scale-invariant estimation approach using airborne lasers. Remote Sens. Environ. 2009, 113, 182–196. [Google Scholar] [CrossRef]
- Hanssen, K.; Solberg, S. Assessment of defoliation during a pine sawfly outbreak: Calibration of airborne laser scanning data with hemispherical photography. For. Ecol. Manag. 2007, 250, 9–16. [Google Scholar] [CrossRef]
- Solberg, S. Mapping gap fraction, LAI and defoliation using various ALS penetration variables. Int. J. Remote Sens. 2010, 32, 1227–1244. [Google Scholar] [CrossRef]
- Räty, M.; Kankare, V.; Yu, X.; Holopainen, M.; Vastaranta, M.; Kantola, T.; Hyyppä, J.; Viitala, R. Tree Biomass Estimation Using ALS Features. In Proceedings of Silvilaser, the 11th International Conference on ALS Applications for Assessing Forest Ecosystems, Hobart, Australia, 16–20 November 2011.
- Vastatanta, M.; Kantola, T.; Lyytikäinen-Saarenmaa, P.; Holopainen, M.; Kankare, V.; Wulder, M.; Hyyppä, J.; Hyyppä, H. Area-Based Mapping of Defoliation of Scots Pine Stands Using Airborne Scanning LiDAR. Remote Sens. 2013, 5, 1220–1234. [Google Scholar] [CrossRef]
- Hyyppä, J.; Jaakkola, A.; Hyyppä, H.; Kaartinen, H.; Kukko, A.; Holopainen, M.; Zhu, L.; Vastaranta, M.; Kaasalainen, S.; Krooks, A.; et al. Map Updating and Change Detection Using Vehicle-Based Laser Scanning. In Proceedings of JURSE 2009, Shanghai, China, 20–22 May 2009.
- Cajander, A.K. The theory of forest types. Acta For. Fenn. 1926, 29, 1–108. [Google Scholar]
- Axelsson, P. DEM Generation from Laser Scanner Data Using Adaptive TIN Models. In Proceedings of XIX ISPRS Congress, Commission I–VII, Amsterdam, The Netherlands, 16–23 July 2000; pp. 110–117.
- Talvitie, M.; Kantola, T.; Holopainen, M.; Lyytikainen-Saarenmaa, P. Adaptive cluster sampling in inventorying forest damage by the common pine sawfly (Diprion pini). J. For. Plan. 2011, 16, 141–148. [Google Scholar]
- Thompson, S.K. Adaptive cluster sampling. J. Am. Stat. Assoc. 1990, 85, 1050–1059. [Google Scholar] [CrossRef]
- Roesch, F.A., Jr. Adaptive cluster sampling for forest inventories. For. Sci. 1993, 39, 655–669. [Google Scholar]
- Eichhorn, J. Manual on Methods and Criteria for Harmonized Sampling, Assessment, Monitoring and Analysis of the Effects of Air Pollution on Forests. Part II. Visual Assessment of Crown Condition and Submanual on Visual Assessment of Crown Condition on Intensive Monitoring Plots; United Nations Economic Commission for Europe Convention on Long-Range Transboundary Air Pollution: Hamburg, Germany, 1998. [Google Scholar]
- Hyyppä, J.; Inkinen, M. Detecting and estimating attributes for single trees using laser scanner. Photogramm. J. Fin. 1999, 16, 27–42. [Google Scholar]
- Yu, X.; Hyyppä, J.; Holopainen, M.; Vastaranta, M.; Viitala, R. Predicting individual tree attributes from airborne laser point clouds based on random forest technique. ISPRS J. Photogramm. Remote Sens. 2011, 66, 28–37. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Crookston, N.L.; Finley, A.O. yaImpute: AR package for e_cient nearest neighbor imputation routines, variance estimation, and mapping. Available online: http://cran.r-project.org (accessed on 10 November 2012).
- Falkowski, M.; Hudak, A.; Crookston, N.; Gessler, P.; Smith, A. Landscape-scale parameterization of a tree-level forest growth model: A k-NN imputation approach incorporating LiDAR data. Can. J. For. Res. 2010, 40, 184–199. [Google Scholar] [CrossRef]
- Hudak, A.; Crookston, N.; Evans, J.; Hall, D.; Falkowski, M. Nearest neighbor imputation of species-level, plot-scale forest structure attributes from LiDAR data. Remote Sens. Environ. 2008, 112, 2232–2245. [Google Scholar]
- Latifi, H.; Nothdurft, A.; Koch, B. Non-parametric prediction and mapping of standing timber volume and biomass in temperate forest: Application of multiple optical/ALS-derived predictors. Forestry 2010, 83, 395–407. [Google Scholar] [CrossRef]
- Falkowski, M.J.; Evans, J.S.; Martinuzzi, S.; Gessler, P.E.; Hudak, A.T. Characterizing forest succession with ALS data: An evaluation for the Inland Northwest, USA. Remote Sens. Environ. 2009, 113, 946–956. [Google Scholar] [CrossRef]
- Kaartinen, H.; Hyyppä, J. EuroSDR/ISPRS Project Commission II, Tree Extraction, Final Report; EuroSDR, 2008. Available online: http://bono.hostireland.com/~eurosdr/publications/53.pdf (accessed on 10 November 2012).
- Kaartinen, H.; Hyyppä, J.; Yu, X.; Vastaranta, M.; Hyyppä, H.; Kukko, A.; Holopainen, M.; Heipke, C.; Hirschmugl, M.; Morsdorf, F.; et al. An international comparison of individual tree detection and extraction using airborne laser scanning. Remote Sens. 2012, 4, 950–974. [Google Scholar] [CrossRef]
- Vauhkonen, J.; Korpela, I.; Maltamo, M.; Tokola, T. Imputation of single-tree attributes using airborne laser scanning-based height, intensity and alpha shape metrics. Remote Sens. Environ. 2010, 114, 1263–1276. [Google Scholar] [CrossRef]
- Vastaranta, M.; Korpela, I.; Uotila, M.; Hovi, A.; Holopainen, M. Area-Based Snow Damage Classification of Forest Canopies Using Bi-Temporal Lidar Data. In Proceedings of ISPRS Workshop on Laser Scanning 2011, Calgary, AB, Canada, 29–31 August 2011; p. 5.
- Vehmas, M.; Packalén, P.; Maltamo, M. Assessing Deadwood Existence in Canopy Gaps by Using ALS Data. In Proceedings of Silvilaser 2009, College Station, TX, USA, 14–16 October 2009.
- Lyytikäinen, P. Effects of natural and artificial defoliations on sawfly performance and foliar chemistry of Scots pine saplings. Ann. Zool. Fenn. 1994, 31, 307–318. [Google Scholar]
- Vehmas, M.; Eerikäinen, K.; Peuhkurinen, J.; Packalén, P.; Maltamo, M. Airborne laser scanning for the site type identification of mature boreal forest stands. Remote Sens. 2011, 3, 100–116. [Google Scholar] [CrossRef]
- Ilvesniemi, S. Numeeriset Ilmakuvat ja Landsat TM-Satelliittikuvat Männyn Neulaskadon Arvioinnissa (in Finnish); Helsingin yliopisto: Helsinki, Finland, 2009; p. 62. [Google Scholar]
- Haara, A.; Nevalainen, S. Detection of dead or defoliated spruces using digital aerial data. For. Ecol. Manag. 2002, 160, 97–107. [Google Scholar] [CrossRef]
- Karjalainen, M.; Kaasalainen, S.; Hyyppä, J.; Holopainen, M.; Lyytikäinen-Saarenmaa, P.; Krooks, A.; Jaakkola, A. SAR Satellite Images and Terrestrial Laser Scanning in Forest Damages Mapping in Finland. In Proceedings of ESA Living Planet Symposium 2010 ESA Special Publication, Bergen, Norway, 28 June–2 July 2010.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kantola, T.; Vastaranta, M.; Lyytikäinen-Saarenmaa, P.; Holopainen, M.; Kankare, V.; Talvitie, M.; Hyyppä, J. Classification of Needle Loss of Individual Scots Pine Trees by Means of Airborne Laser Scanning. Forests 2013, 4, 386-403. https://doi.org/10.3390/f4020386
Kantola T, Vastaranta M, Lyytikäinen-Saarenmaa P, Holopainen M, Kankare V, Talvitie M, Hyyppä J. Classification of Needle Loss of Individual Scots Pine Trees by Means of Airborne Laser Scanning. Forests. 2013; 4(2):386-403. https://doi.org/10.3390/f4020386
Chicago/Turabian StyleKantola, Tuula, Mikko Vastaranta, Päivi Lyytikäinen-Saarenmaa, Markus Holopainen, Ville Kankare, Mervi Talvitie, and Juha Hyyppä. 2013. "Classification of Needle Loss of Individual Scots Pine Trees by Means of Airborne Laser Scanning" Forests 4, no. 2: 386-403. https://doi.org/10.3390/f4020386
APA StyleKantola, T., Vastaranta, M., Lyytikäinen-Saarenmaa, P., Holopainen, M., Kankare, V., Talvitie, M., & Hyyppä, J. (2013). Classification of Needle Loss of Individual Scots Pine Trees by Means of Airborne Laser Scanning. Forests, 4(2), 386-403. https://doi.org/10.3390/f4020386