Volatile Compound Chemistry and Insect Herbivory: Pinus edulis Engelm. (Pinaceae) Seed Cone Resin


1
D. Gary Young Research Institute, Lehi, UT 84043, USA
2
Department of Biology, Utah Valley University, Orem, UT 84058, USA
*
Author to whom correspondence should be addressed.
Forests 2023, 14(9), 1862; https://doi.org/10.3390/f14091862
Submission received: 1 August 2023
/
Revised: 29 August 2023
/
Accepted: 11 September 2023
/
Published: 13 September 2023
(This article belongs to the Section Forest Ecophysiology and Biology)
Abstract
:Pinus edulis Engelm. (pinyon pine) is a tree in the Pinaceae family with wide geographic distribution throughout dry forests of western North America. Pinyon pine seed cones, which mature over a 2-year period before shedding seed, are often resinous. Conifer resin, which is a liquid-soluble mixture of volatile and non-volatile secondary metabolites, typically demonstrates significant ecological functions. In the current study, seed cones (n = 240) were collected monthly for a 1-year period from pinyon pine trees (n = 20) and separated equally into two groups, resinous and non-resinous cones, for research on the volatile compound chemistry and insect herbivory. Upon distillation, resinous cones yielded 41× more volatile oil and contained more viable seeds, compared to non-resinous cones. Chemical profiles of volatile oils were seemingly consistent between seasons but differed between cone groups. In resinous cone samples, volatile oils were largely composed of α-pinene (avg. 75.6%) and δ-3-carene (avg. 7.4%). In contrast, in non-resinous cone samples, the volatile oil profiles were not dominated by any single prominent compound. Cone inquiline insect communities varied with regard to the resinous status of the cones, the month of collection, and with overall volatile oil yield. Typically, with larger oil yields we saw more diverse and more abundant inquiline communities. Findings from this study suggest that the volatile oils, in addition to other components of cone resin and physical structures, play a significant ecological role in pinyon pine seed preservation from insect herbivory.
1. Introduction
Pinus edulis Engelm. (pinyon pine) is a monoecious, aromatic tree in the Pinaceae family. Pinyon pine trees typically grow to 15 m in height, with the crown often as wide as the tree is tall [1]. This species is native to dry, mountain slopes in North America (AZ, CA, CO, NM, OK, TX, UT, WY, Chihuahua Mexico) [2]. Pinus edulis, along with Juniperus osteosperma, form the pinyon–juniper woodlands in Utah, with J. osteosperma acting as a nurse plant for pinyon pine [3,4]. The tree is slow growing and reaches maturity by 200 years, may live to 1000 years, and produces seeds for several centuries [2,5]. The resinous seed cones (also known as ovulate or female cones) mature over a 2-year period before shedding seed [4,5].
Langenheim [6] defines a plant resin as a “liquid-soluble mixture of volatile and nonvolatile terpenoid and/or phenolic secondary compounds” that meet two requirements: the resin is secreted by specialized plant structures, and has a significant ecological interaction. The same author also states that the volatile fraction of resins produced by trees in the Pinaceae family often acts as a defense against insects and pathogens. Previous research has shown that the volatile fraction (composed of terpenoids and sesquiterpenoids) extracted from pinyon pine cone resin has the highest yield (w/w) of any other aromatic portion of the plant and is largely composed of the volatile monoterpene α-pinene [7].
Pinyon pine gum (resin) was used medicinally by Native peoples [8,9] and the edible seeds were historically consumed by humans, and still are in modern times [3,4,10,11,12,13]. Additionally, insects (Dioryctria albovittella) and animals (Aphelocoma sp., Gymnorhinus cyanocephalus, Nucifraga columbiana, Otospermophilus variegatus, Tamias dorsalis) compete for pinyon pine seeds as a food source [1,14,15]. Pinyon pines face considerable levels of seed predation, which has largely been studied in relation to avian [16,17] and mammalian predators [14,18,19].
It was found that during peak herbivory on pinyon pine by the southwestern tiger moth (Lophocampa ingens), monoterpene emissions (including α-pinene) rates were higher (2–6×) from undamaged needles on damaged trees compared to the rest of the season [20]. It was also found that chronic herbivory by the stem-boring moth (Dioryctria albovittella) caused the tree to reduce seed cone production and even caused immature cone abortion on intact branches of damaged trees [15]. In addition to the stem boring moth, pinyon pine also hosts a wide variety of other insects that primarily inhabit and/or attack cones and seeds [21]. These insects have seen considerably less attention from researchers, apart from their application to forest timber health. The insects’ interactions as a community and how they respond to changes in cone status throughout the year are relatively unknown. Insect seed predators can have significant impacts on pinyon seed success as well as on other seed predators, often dictating the outcomes of intra-specific seed predators’ competition [14].
Given that seed cones mature over a 2-year period before shedding, and are a highly sought out food source, further investigation is needed to explore evolutionary mechanisms for seed preservation. The current study investigates the relationship between pinyon pine cone resin volatiles and insect herbivory over a 1-year period.
2. Materials and Methods
Pinus edulis Engelm. seed cones were collected monthly (January 2021–December 2021) from a native population located on private land in Duchesne County, Utah, USA. The tree stand was composed of uneven age class trees co-mingled with juniper (Juniperus spp.) and an extensive shrubby understory. Tree ages varied between 40 and 400 years old with an average age of 114 ± 20 years (Rotter unpublished 2022). During the first week of each month, 12 cones were collected from each of the 20 previously selected trees (Figure 1) (n = 240 cones collected monthly or 2880 cones collected over the duration of the 12-month study) and separated equally into two groups for research on the volatile compound chemistry and insect herbivory. Prior to collecting seed cones from each tree, a visual assessment was performed to determine the ratio of resinous cones to non-resinous cones (Figure 2), and the 12 seed cones were collected from each tree according to that ratio. Seed cones, defined as the female or ovulate cones that develop and house the edible seeds, were separated/studied as two distinct groups, resinous and non-resinous. Plant material was stored at −20 °C until steam distillation and volatile compound analysis or insect herbivory research. Representative voucher samples of Pinus edulis are held in the Young Living Aromatic Herbarium (YLAH): Pinus edulis Engelm., Wilson 2021-01, -02, -03 (YLAH).
Volatile Compound Chemistry Method. Laboratory scale distillation was carried out as follows: 3 L of water added to the bottom of a 12-L distillation chamber (Albrigi Luigi S.R.L., Grezzana, Italy), plant material accurately weighed and added to the distillation chamber, distillation for 2 h using direct steam, and volatile oil separated with a cooled condenser and Florentine flask. Volatile samples were filtered and stored in a sealed amber glass bottle at −20 °C until analysis.
The percent yield was calculated as the ratio of mass of processed plant material immediately before distillation to the mass of volatile oil produced, multiplied by 100.
Volatile oil samples were analyzed, and volatile compounds identified, with GC/MS using an Agilent 7890B GC/5977B MSD (Agilent Technologies, Santa Clara, CA, USA) and Agilent J&W DB-5, 0.25 mm × 60 m, 0.25 μm film thickness, and fused silica capillary column. Operating conditions: 0.1 μL of sample (20% soln. for volatile oils in ethanol), 100:1 split ratio, initial oven temp. of 40 °C with an initial hold time of 5 min., oven ramp rate of 4.5 °C per min. to 310 °C with a hold time of 5 min. The electron ionization energy was 70 eV, scan range 35–650 amu, scan rate 2.4 scans per s, source temp. 230 °C, and quadrupole temp. 150 °C. Volatile compounds were identified using the Adams volatile oil library [22] using Chemstation library search in conjunction with retention indices. When identifications could not be made using Adam’s library, the NIST Mass Spectral Library [23] was used, and KI calculated using C7-C30 alkane standards. Note that limonene/β-phellandrene co-elutes and their amounts were determined via the ratio of masses 68 and 79 (limonene), and 77 and 93 (β-phellandrene). Volatile compounds were quantified and are reported as a relative area percentage by GC/FID using an Agilent 7890B GC and Agilent J&W DB-5, 0.25 mm × 60 m, 0.25 μm film thickness, fused silica capillary column. Operating conditions: 0.1 μL of sample (20% soln. for volatile oils in ethanol, 1% for reference compounds in ethanol, 0.1% soln. for C7–C30 alkanes in hexane), 25:1 split ratio, initial oven temp. of 40 °C with an initial hold time of 2 min., oven ramp rate of 3.0 °C per min. to 250 °C with a hold time of 3 min. For quantification, compounds were identified using retention indices coupled with retention time data of reference compounds (MilliporeSigma, Sigma-Aldrich, St. Louis, MS, USA).
Insect Herbivory Method. Cones collected for insect sampling were stored at −20 °C until being dissected for insects living inside the cones. All insects were identified to the lowest possible taxonomic level (typically to genus, with a few groups to family). We then counted how many taxonomic groups were represented in each cone. Cones for each month were divided into either resinous or non-resinous groups (as in the chemistry methods). In addition, we also measured the thickness of a random central scale on each cone (as a potential measurement of a physical cone defense) and counted the number of viable and non-viable seeds in the cones.
Statistical Analyses. To understand the relationships influencing insect community composition and abundance, we used Canonical Correspondence Analysis (CCA). The use of CCA in this case is ideal as it allows us to look at the insect community as it relates to our measured variables. A 999-run iteration of the Monte Carlo randomization tests was used to determine the significance of the CCA ordination. The analysis of the insect cone community was carried out in PC ORD ver. 7.10 [24].
We also used relative insect abundance, Shannon’s diversity index and Simpson’s diversity index to track the monthly average changes in insect communities over the course of the year. This was also used to look for monthly differences between resinous and non-resinous cones and tracked with monthly changes to percentage yield of total volatile oils.
3. Results and Discussion
The monthly volatile oil yield for resinous and non-resinous cones is detailed in Table 1. The resinous cone yield was, on average, 41× higher than the non-resinous cone yield. The resinous cone yield ranged from 0.26% (w/w) in November to 0.59% (w/w) in January. The non-resinous cone yield ranged from <0.01% (w/w) in February to 0.02% (w/w) in October and December. The yield data from resinous cones were similar to previous findings from the same population of Pinus edulis Engelm. trees (plant material collected during summer), where average yields were 0.6% (w/w) [7]. In the current study, resinous seed cones weighed 42% more (on average) than non-resinous cones (5.52 g vs. 3.89 g).
The complete volatile compound profiles for the winter-dormant season (January) and the summer-growing season (July) are detailed below in Table 2 for resinous and non-resinous seed cones. The months of January and July were selected since, on average, those are the coldest and warmest months in Utah, respectively (www.weather.gov, accessed on 29 August 2023). While chemical profiles were seemingly consistent between seasons, they differed greatly between cone source (resinous vs. non-resinous). For January and July, the average values for α-pinene in resinous cones and non-resinous cones were 75.7% and 3.2%, respectively. In resinous seed cones, a total of 64 volatile compounds were detected and the majority (63) were confidently identified. By contrast, in non-resinous seed cones, a total of 88 compounds were detected with only 71 confidently identified. Sixteen of the seventeen unidentified volatile compounds in non-resinous seed cones were late eluting sesquiterpenoids, with relative area percentages ranging from 0.2% to 2.6%. Since these same compounds were not detected in any of the resinous seed cone samples, they are possibly either degradation products or stable compounds that are present in all samples but below the level of detection in some samples. Future updates of the commercially available mass spectral libraries used in this study [22,23] may allow for a confident identification of these compounds in question. To identify these compounds, future investigations could include the isolation of the unknown compounds using preparative chromatography followed by analytical techniques suitable for structure elucidation, such as nuclear magnetic resonance spectroscopy (NMR), crystallography, and exact mass mass spectrometry (ex.: GC/TOF/MS).
Summarized profiles for the most abundant volatile compounds present in resinous and non-resinous cone samples are provided in Table 3 and Table 4, respectively. In resinous cone samples, the average sum of α-pinene (avg. 75.6%), δ-3-carene (avg. 7.4%), and eight others of the most prominent compounds comprised 92.9% of each volatile oil sample. In contrast, in non-resinous cone samples, the profile was not dominated by any single prominent compound. On average, α-pinene (avg. 14.6%), δ-3-carene (avg. 4.5%), β-bourbonene (5.1%), longifolene (8.8%), and thirteen others of the most prominent compounds comprised only 59.9% of each volatile oil sample from non-resinous cones.
The insecticidal properties of terpenes have been a promising topic of recent research [25,26]. Given the consistent average relative abundance of α-pinene (avg. 75.6%, σ 1.4) and δ-3-carene (avg. 7.4%, σ 0.9) in resinous cone samples, these are likely important volatile compounds, either directly or indirectly, in ecological interactions. Both compounds have been the topic of insecticidal activity research on maize weevil (Sitophilus zeamais), where they were found to repel, inhibit developmental stages, and reduce the progeny of weevils [27]. Other researchers associated α-pinene with indirectly reducing the development and reproduction of pinewood nematodes (Bursaphelenchus xylophilous) [28]. Antifeedant effects and growth inhibition have been observed by using α-pinene as an insecticide on flour beetles (Tribolium castaneum) [29]. In ecological settings, terpenes have been shown to attract specialized insect herbivores when they exist in greater concentration in branches [30]. Another consideration is the indirect use of terpenes in pinyon pine. Previous researchers have found that certain terpenes are catabolized into usable primary metabolites [31,32]. However, the catabolism of α-pinene and δ-3-carene needs to be studied to investigate this further.
In non-resinous cones, the α-pinene content is much lower than in resinous cones, and values are inconsistent (avg. 14.6%, σ 19.2). Samples from October and December are potentially outliers (volatile oil yield 2× the mean and mode—see Table 1; highest values for α-pinene—see Table 4). Since human judgment balances on the verge of error, this is likely due to human error in distinguishing the cutoff between resinous and non-resinous cones. When considering the larger picture, it is unlikely that the volatile compound profile is of significance in non-resinous cones since the volatile oil profile differs from that of resinous cones and the yield is 41× lower.
Overall, we recorded six groups of inquiline invertebrates in collected cones (Table 5). Invertebrates represented several different trophic guilds, feeding strategies, and orders. The recorded groups were all typical of the cones of Pinus edulis [21]. The seed predator groups represent specialized feeding groups on cones of various Pinus species, and the other groups represent predators of invertebrates that are associated with pinyon pine communities [33]. In addition to identifiable taxa, we had six specimens that we could not identify. These likely represent remains of prey items or other insect artifacts. One specimen was a hyperparasitoid (Ichneumonidae Family Hymenoptera). Since it was recovered only once, it was noted but not included in our analysis.
Cone inquiline communities varied with regard to the resinous status of the cones (Figure 3A), the month of the collection (Figure 3B), and with overall oil yield (Figure 4). Non-resinous cone communities were very different than the resinous cone communities. These differences were heavily influenced by the presence of seeds and an increase in volatile oils. Non-resinous cones typically lack seeds as they are usually dry remains and have long stopped producing any volatile oils. These cones attract species of insects and arthropods that are likely “interlopers” that only take advantage of the three-dimensional habitats of the cone. These arthropod communities are dominated by spiders and other non-seed predator insects. The resinous cones are characterized by the presence of seeds and a corresponding increase in volatile oils. In this situation, the seeds are likely driving this change in the inquiline community, as species here are specialized seed predators of pinyon cones (such as Conophthorus edulis and Conotrachelus spp.). Further, these specialized seed predators likely attract their own predators, such as in the presence of several parasitic wasp genera.
Changes in resinous and non-resinous cone abundance likely result in the influence of temporal changes on insect communities. November and December had only a minimal collection of resinous cones. As the cone develops, it will produce volatile oils that coat the cone in a resinous layer to prevent insects from consuming the developing embryo. However, as the cone matures and the seed ripens, oil production decreases (Table 1). The stronger dichotomy in our study was the difference between resinous vs. non-resinous cones and not the month-to-month changes. There is a risk that our own collection could have contributed to this, as we selected the cones. Early in our collection, we may have had a slight sample bias for easier to reach or larger cones.
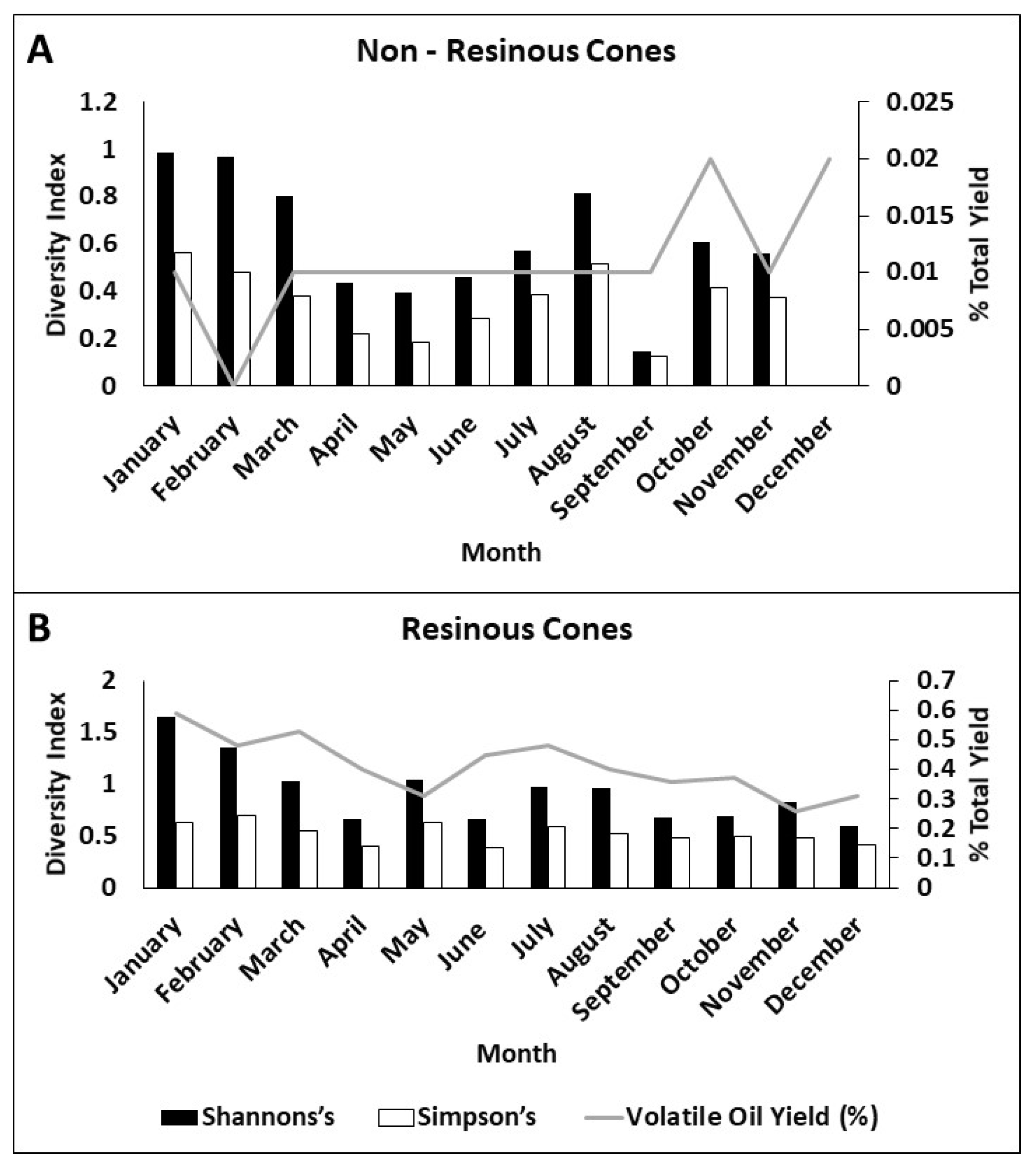
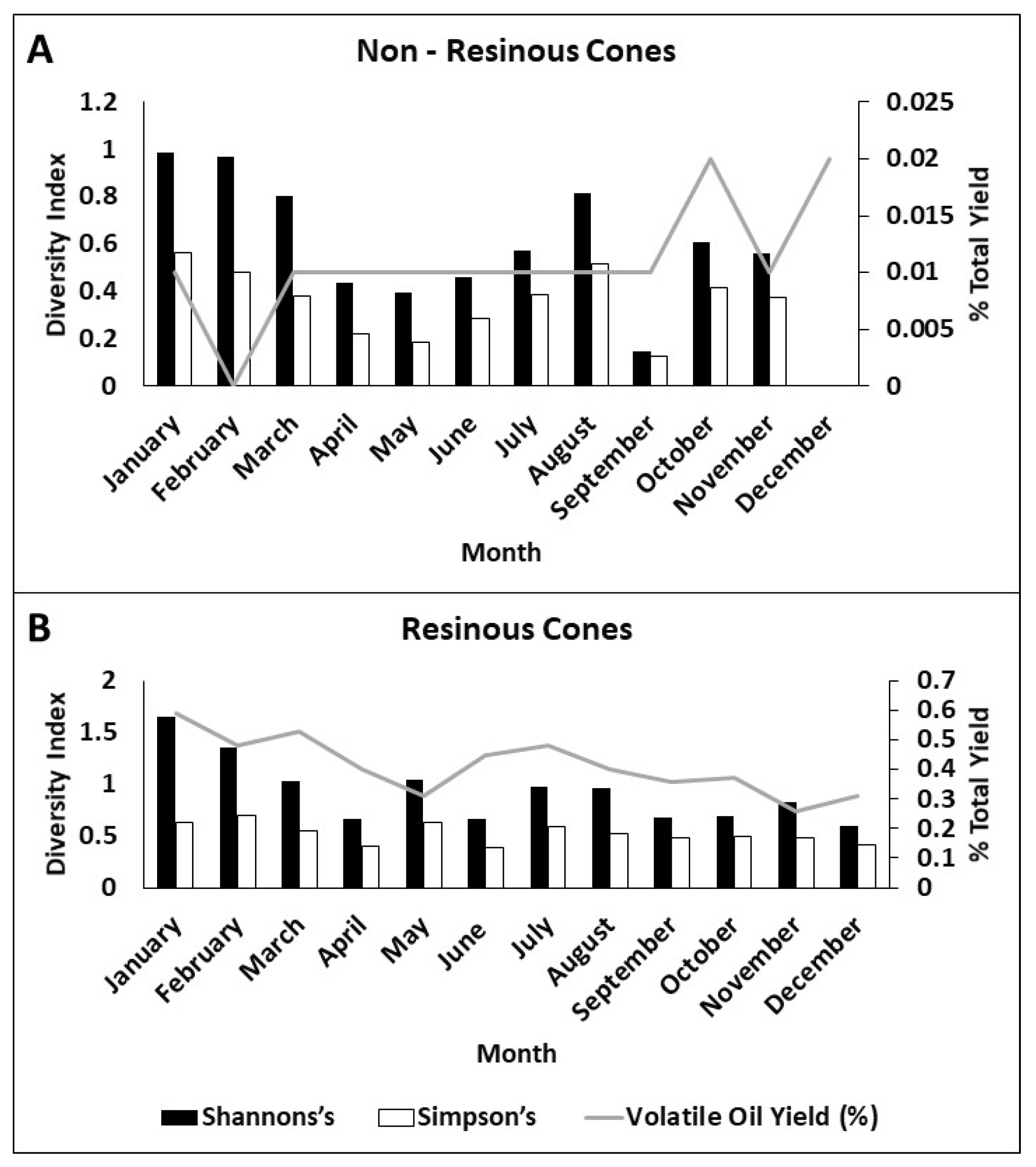
Within the cones, insect diversity changed throughout the year (Figure 4). Non-resinous cones exhibited spikes in diversity, particularly in colder winter months. This, again, is likely due to non-specialized inquiline insects moving into more available dry cones at these times of year. This included most of our findings of spiders in these dry cones. This was likely because these habitats are more available and because they allow them to exploit better shelter and foraging areas [34]. It is also important to note that there seems to be no relationship between diversity in these non-resinous cones and oil yield. Resinous cones had a higher overall diversity that fluctuated with the total volatile oil yield. As these insect communities are composed of specialist insects, it is likely that they are influenced by the development of the seeds and the corresponding fluctuations in volatile oil. Inquiline relative abundance was predicted by total volatile oil yield in resinous cones (R2 = 0.41, p = 0.025), and this relationship was un-informative in non-resinous cones due to the overall low abundances and low volatile oil yield (R2 = 0.005, p = 0.988).
4. Conclusions
The current study investigates the ecological role that Pinus edulis Engelm. (pinyon pine) cone volatile oils play in seed preservation, in conjunction with non-volatile fractions of resin and the physical structures of the cone. Resinous and non-resinous seed cones displayed seasonal variation in their volatile oil yield and profile, number and viability of seeds, and inquiline insect communities. Specialist seed predators were likely able to use these biotic and abiotic changes as cues to find food sources.
In pinyon pine, cones are produced in a somewhat random pattern, which is influenced by general environmental conditions and conditions presented to the individual tree itself. This was observed in the current study, with a sharp dichotomy between dry and resinous cones. Resinous cones, which were primarily composed of α-pinene (avg. 75.6%, σ 1.4) and δ-3-carene (avg. 7.4%, σ 0.9), typically bore viable seeds and the insect inquiline community in these cones was distinct, with more abundant and diverse insect speciation. As cone growth began in the spring with warmer temperatures, increases in volatile oils and resins encompassing the growth areas of cones were observed, which continued throughout fertilization. The data indicate that this first period tends to be the most active for the insect inquiline community, where specialist insects are attracted to the cones to lay their eggs. As the seeds began to set within the cone, volatile oil yields tended to stabilize, or even trended slightly downward, while the inquiline diversity and relative abundance stabilized as well. This long “calm” period may be punctuated by changes in the inquiline community, likely as insects are preyed upon or other species turnover occurs. As the seed set (or failed), volatile oil yields plummeted within the cones. With almost no volatile oils being produced, these dry cones remained on the tree and became home to overwintering insects or spiders.
Findings from this study demonstrate the significant ecological role that volatile oils, as well as the non-volatile fractions of the resin and the cone structure, play in seed preservation from insect herbivory. While these volatile oils play a significant role in repelling more generalized insect herbivores during seed maturation, other important seed predators, including birds and mammals, were not considered in the current study. Future investigations should research how these same factors impact predation from larger animals, and also investigate the role that non-volatile fractions of cone resin play in seed preservation. Additionally, insecticidal activity of the most prevalent volatile compounds in resinous cones, α-pinene and δ-3-carene, should be investigated as isolated compounds with isolated insect species.
Author Contributions
Conceptualization, T.M.W. and M.C.R.; methodology, T.M.W. and M.C.R.; software, T.M.W. and M.C.R.; validation, R.E.C.; formal analysis, T.M.W., E.A.Z. and M.C.R.; investigation, T.M.W., E.A.Z. and M.C.R.; resources, R.E.C.; data curation, T.M.W. and M.C.R.; writing—original draft preparation, T.M.W. and M.C.R.; writing—review and editing, E.A.Z. and R.E.C.; funding acquisition, R.E.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Young Living Essential Oils.
Data Availability Statement
The data presented in this study are available upon request from the corresponding author.
Acknowledgments
The authors would like to thank the following individuals and organizations for their assistance with the study: the late Robert P. Adams for originally proposing the topic of study, Ariel Poulson (YLEO), John Whetten (YLEO), Mackenzie G. Dey (USU), Sheldon Giles (Skyrider Ranch), Skyler Olson (Skyrider Ranch), and the D. Gary Young Research Institute for providing support for this project. We also thank Melissa Fuhriman, Mason Hoffman, Josh Leon, and Samantha Ingram for their help in the field and lab.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Van Buren, R.; Cooper, J.G.; Shultz, L.M.; Harper, K.T. Woody Plants of Utah: A Field Guide with Identification Keys to Native and Naturalized Trees, Shrubs, Cacti, and Vines; University Press of Colorado: Denver, CO, USA, 2012; pp. 30, 118, 119. [Google Scholar]
- Flora of North American Editorial Committee. Flora of North America: Pteridophytes and Gymnosperms; Oxford University Press: New York, NY, USA, 1993; Volume 2, pp. 382–383. [Google Scholar]
- Cronquist, A.; Holmgren, A.H.; Holmgren, N.H.; Reveal, J.L. Intermountain Flora; Vascular Plants of the Intermountain West, USA; Hafner Publishing Company, Inc.: New York, NY, USA, 1972; Volume 1, p. 232. [Google Scholar]
- Welsh, S.L.; Atwood, N.D.; Goodrich, S.; Higgins, L.C. A Utah Flora, 5th ed.; Brigham Young University: Provo, UT, USA, 2016; p. 20. [Google Scholar]
- Ronco, F.P. Pinus edulis Engelm., pinyon. In Silvics of North America; United States Department of Agriculture: Washington, DC, USA, 1990; Volume 1, pp. 327–337. [Google Scholar]
- Langenheim, J.H. Plant Resins: Chemistry, Evolution, Ecology, Ethnobotany; Timber Press, Inc.: Portland, OR, USA, 2003; pp. 24, 33–36. [Google Scholar]
- Poulson, A.; Wilson, T.M.; Packer, C.; Carlson, R.E.; Buch, R.M. Essential oils of trunk, limbs, needles, and seed cones of Pinus edulis (Pinaceae) from Utah. Phytologia 2020, 102, 200–207. [Google Scholar]
- Kindscher, K. Medicinal Wild Plants of the Prairie; University Press of Kansas: Lawrence, KS, USA, 1992; p. 62. [Google Scholar]
- Murphy, E.V.A. Indian Uses of Native Plants; Desert Printers Inc.: Palm Desert, CA, USA, 1959; p. 44. [Google Scholar]
- Bentancourt, J.L.; Schuster, W.S.; Mitton, J.B.; Anderson, R.S. Fossil and genetic history of a pinyon pine (Pinus edulis) isolate. Ecology 1991, 72, 1685–1697. [Google Scholar] [CrossRef]
- Rhode, D.; Madsen, D.B. Pine nut use in the early Holocene and beyond: The Danger Cave archaeobotanical record. J. Archaeol. Sci. 1998, 25, 1199–1210. [Google Scholar] [CrossRef]
- Grand Canyon Nature Notes. Available online: http://npshistory.com/nature_notes/grca/vol8-9c.htm (accessed on 1 August 2023).
- Uphof, T.C.T. Dictionary of Economic Plants; J. Cramer Publisher: Lehre, Germany, 1968; p. 408. [Google Scholar]
- Christensen, K.M.; Whitham, T.G. Impact of insect herbivores on competition between birds and mammals for pinyon pine seeds. Ecology 1993, 74, 2270–2278. [Google Scholar] [CrossRef]
- Cobb, N.S.; Trotter, R.T.; Whitham, T.G. Long-term sexual allocation in herbivore resistant and susceptible pinyon pine (Pinus edulis). Oecologia 2002, 130, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Christensen, K.M.; Whitham, T.G. Indirect herbivore mediation of avian seed dispersal in pinyon pine. Ecology 1991, 72, 534–542. [Google Scholar] [CrossRef]
- Siepielski, A.M.; Benkman, C.W. Selection by a predispersal seed predator constrains the evolution of avian seed dispersal in pines. Funct. Ecol. 2007, 21, 611–618. [Google Scholar] [CrossRef]
- Hollander, J.L.; Vander Wall, S.B. Effectiveness of six species of rodents as dispersers of singleleaf pinon pine (Pinus monophylla). Oecologia 2004, 138, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Vander Wall, S.B. Dispersal of singleleaf pinon pine (Pinus monophylla) by seed-caching rodents. J. Mammal. 1997, 78, 181–191. [Google Scholar] [CrossRef]
- Trowbridge, A.M.; Daly, R.W.; Helmig, D.; Stoy, P.C.; Monson, R.K. Herbivory and climate interact serially to control monoterpene emissions from pinyon pine forests. Ecology 2014, 95, 1591–1603. [Google Scholar] [CrossRef] [PubMed]
- Little, E.L. Common insects on pinyon (Pinus edulis). J. N. Y. Entomol. Soc. 1943, 51, 239–252. [Google Scholar]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4th ed.; Allured Publ.: Carol Stream, IL, USA, 1997. [Google Scholar]
- National Institute of Standards and Technology Mass Spectrometry Data Center. Available online: https://chemdata.nist.gov (accessed on 7 March 2022).
- McCune, B.; Mefford, M.J. PC-ORD. Multivariate Analysis of Ecological Data, Version 7.10; MjM Software Design: Gleneden Beach, OR, USA, 2018. [Google Scholar]
- López, M.D.; Jordán, M.J.; Pascual-Villalobos, M.J. Toxic compounds in essential oils of coriander, caraway and basil active against stored rice pests. J. Stored. Prod. Res. 2008, 44, 273–278. [Google Scholar] [CrossRef]
- Reis, S.L.; Mantello, A.G.; Macedo, J.M.; Gelfuso, E.A.; Da Silva, C.P.; Fachin, A.L.; Cardoso, A.M.; Beleboni, R.O. Typical monoterpenes as insecticides and repellents against stored grain pests. Molecules 2016, 21, 258. [Google Scholar] [CrossRef] [PubMed]
- Langsi, J.D.; Nukenine, E.N.; Oumarou, K.M.; Moktar, H.; Fokunang, C.N.; Mbata, G.N. Evaluation of the insecticidal activities of α-pinene and 3-carene on Sitophilus zeamais Motschulsky (Coleoptera: Curculionidae). Insects 2020, 11, 540. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yu, Y.; Ge, J.; Xie, B.; Zhu, S.; Cheng, X. Effects of α-pinene on the pinewood nematode (Bursaphelenchus xylophilus) and its symbiotic bacteria. PLoS ONE 2019, 14, e0221099. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Hee, S.K.; Ho, S.H. Antifeedant and growth inhibitory effects of α-pinene on the stored-product insects, Tribolium castaneum (Herbst) and Sitophilus zeamais Motsch. Int. Pest. Control. 1998, 40, 18–20. [Google Scholar]
- Ruel, J.; Whitham, T.G. Fast-growing juvenile pinyons suffer greater herbivory when mature. Ecology 2002, 83, 2691–2699. [Google Scholar] [CrossRef]
- Croteau, R.; Martinkus, C. Metabolism of monoterpenes: Demonstration of (+)-neomenthyl-β-d-glucoside as a major metabolite of (−)-menthone in peppermint (Mentha piperita). Plant Physiol. 1979, 64, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Croteau, R.; Sood, V.K. Metabolism of monoterpenes: Evidence for the function of monoterpene catabolism in peppermint (Mentha piperita) rhizomes. Plant Physiol. 1985, 77, 801–806. [Google Scholar] [CrossRef] [PubMed]
- Fairweather, M.L. Field Guide to Insects and Diseases of Arizona and New Mexico Forests; USDA Forest Service, Southwestern Region: Washington, DC, USA, 2006. [Google Scholar]
- Ramseyer, L.J.; Crawford, R.L. A survey of spiders found in fallen pine cones in eastern Washington State. West. N. Am. Nat. 2014, 74, 405–415. [Google Scholar] [CrossRef]
Figure 1.
Map (Google Earth) showing location of Pinus edulis Engelm. trees #1–20. The area is approximately 200 m by 50 m. Coordinates range from north (40°19′55″ N, 100°42′31″ W), east (40°19′54″ N, 100°42′29″ W), south (40°19′48″ N, 100°42′41″ W), and west (40°19′49″ N, 100°42′42″ W). Elevations range from 2034 to 2066 m.
Figure 1.
Map (Google Earth) showing location of Pinus edulis Engelm. trees #1–20. The area is approximately 200 m by 50 m. Coordinates range from north (40°19′55″ N, 100°42′31″ W), east (40°19′54″ N, 100°42′29″ W), south (40°19′48″ N, 100°42′41″ W), and west (40°19′49″ N, 100°42′42″ W). Elevations range from 2034 to 2066 m.

Figure 2.
Photos of resinous (A–F) and non-resinous (G–L) Pinus edulis Engelm. seed cones (top-to-bottom, left-to-right).
Figure 2.
Photos of resinous (A–F) and non-resinous (G–L) Pinus edulis Engelm. seed cones (top-to-bottom, left-to-right).

Figure 3.
Canonical correspondence analysis of cones displayed for insect communities inhabiting non-resinous and resinous cones. Biplots show correlations of R2 value greater than 0.1. Eigenvalues for axis 1 = 0.176 (32.5%) and axis 2 = 0.006 (1.1%). For the 999 iteration Monte Carlo randomization test, p = 0.029. (A) CCA displaying community differences between resinous and non-resinous cones. (B) CCA displaying community differences between months with community average displayed by the plus symbol.
Figure 3.
Canonical correspondence analysis of cones displayed for insect communities inhabiting non-resinous and resinous cones. Biplots show correlations of R2 value greater than 0.1. Eigenvalues for axis 1 = 0.176 (32.5%) and axis 2 = 0.006 (1.1%). For the 999 iteration Monte Carlo randomization test, p = 0.029. (A) CCA displaying community differences between resinous and non-resinous cones. (B) CCA displaying community differences between months with community average displayed by the plus symbol.

Figure 4.
Changes in insect community diversity over months of cone collections and volatile oil yield in each month. Shannon’s diversity index in black, Simpson’s diversity index in white. Volatile oil yield (in %) as grey line. (A) Non-resinous cones and (B) resinous cones.
Figure 4.
Changes in insect community diversity over months of cone collections and volatile oil yield in each month. Shannon’s diversity index in black, Simpson’s diversity index in white. Volatile oil yield (in %) as grey line. (A) Non-resinous cones and (B) resinous cones.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Yield data, including month of collection, seed cone condition, seed cone weight (g), seed cone number (count), average seed cone weight (g), volatile oil weight (g), and calculated yield % (w/w). During the first week of each month, 12 cones were collected from each of the 20 previously selected trees (n = 240 cones collected monthly or 2880 cones collected the duration of the 12-month study).
Table 1.
Yield data, including month of collection, seed cone condition, seed cone weight (g), seed cone number (count), average seed cone weight (g), volatile oil weight (g), and calculated yield % (w/w). During the first week of each month, 12 cones were collected from each of the 20 previously selected trees (n = 240 cones collected monthly or 2880 cones collected the duration of the 12-month study).
| Date/Condition | Cone Weight (g) | Cone # | Avg. Seed Cone Weight (g) | Volatile Oil Weight (g) | Yield (%) | |
|---|---|---|---|---|---|---|
| January | resinous | 437.49 | 75 | 5.83 | 2.60 | 0.59 |
| non-resinous | 194.98 | 45 | 4.33 | 0.01 | 0.01 | |
| February | resinous | 384.36 | 68 | 5.65 | 1.83 | 0.48 |
| non-resinous | 205.49 | 52 | 3.95 | 0.01 | <0.01 | |
| March | resinous | 360.52 | 69 | 5.22 | 1.91 | 0.53 |
| non-resinous | 191.70 | 51 | 3.76 | 0.01 | 0.01 | |
| April | resinous | 384.39 | 74 | 5.19 | 1.55 | 0.40 |
| non-resinous | 182.82 | 46 | 3.97 | 0.01 | 0.01 | |
| May | resinous | 572.46 | 71 | 8.06 | 1.78 | 0.31 |
| non-resinous | 263.91 | 49 | 5.39 | 0.02 | 0.01 | |
| June | resinous | 380.99 | 77 | 4.95 | 1.71 | 0.45 |
| non-resinous | 138.43 | 43 | 3.22 | 0.01 | 0.01 | |
| July | resinous | 363.85 | 69 | 5.27 | 1.75 | 0.48 |
| non-resinous | 167.46 | 51 | 3.28 | 0.01 | 0.01 | |
| August | resinous | 369.89 | 72 | 5.14 | 1.48 | 0.40 |
| non-resinous | 195.3 | 48 | 4.07 | 0.02 | 0.01 | |
| September | resinous | 366.58 | 75 | 4.89 | 1.32 | 0.36 |
| non-resinous | 164.49 | 45 | 3.66 | 0.02 | 0.01 | |
| October | resinous | 354.99 | 68 | 5.22 | 1.33 | 0.37 |
| non-resinous | 183.21 | 52 | 3.52 | 0.04 | 0.02 | |
| November | resinous | 395.01 | 70 | 5.64 | 1.04 | 0.26 |
| non-resinous | 195.08 | 50 | 3.90 | 0.01 | 0.01 | |
| December | resinous | 318.52 | 62 | 5.14 | 0.98 | 0.31 |
| non-resinous | 213.63 | 58 | 3.68 | 0.05 | 0.02 | |
Table 2.
GC/FID profile (area%) of volatile fraction extracted from Pinus edulis Engelm. seed cone resin, resinous and non-resinous, from winter (January) and summer (July) months. Compounds not detected in a sample are denoted as not detected (nd) and those with values less than 0.1% are denoted as traces (tr). Compounds less than 0.5% that were unidentified are not included. KI is the Kovat’s Index using a linear calculation on the DB-5 column [22], and those in bold font were calculated using alkane standards. Relative area percentage was determined using GC/FID. On average, 95.6% of sample profiles were quantified.
Table 2.
GC/FID profile (area%) of volatile fraction extracted from Pinus edulis Engelm. seed cone resin, resinous and non-resinous, from winter (January) and summer (July) months. Compounds not detected in a sample are denoted as not detected (nd) and those with values less than 0.1% are denoted as traces (tr). Compounds less than 0.5% that were unidentified are not included. KI is the Kovat’s Index using a linear calculation on the DB-5 column [22], and those in bold font were calculated using alkane standards. Relative area percentage was determined using GC/FID. On average, 95.6% of sample profiles were quantified.
| Compound Name | KI | Resinous Cones | Non-Resinous Cones | ||
|---|---|---|---|---|---|
| January | July | January | July | ||
| hexanal | 801 | tr | nd | nd | nd |
| tricyclene | 921 | 0.2 | 0.2 | nd | tr |
| α-thujene | 924 | 0.2 | 0.2 | nd | tr |
| α-pinene | 932 | 75.2 | 76.1 | 3.6 | 2.7 |
| camphene | 946 | 0.9 | 1.0 | tr | tr |
| thuja-2,4(10)diene | 953 | 0.4 | 0.9 | tr | 0.3 |
| 3,7,7-trimethyl-1,3,5-cycloheptatriene | 966 | 0.5 | 0.7 | tr | 0.2 |
| sabinene | 969 | 2.2 | 1.5 | tr | 0.1 |
| β-pinene | 974 | 2.5 | 2.9 | tr | 0.3 |
| myrcene | 988 | 0.6 | 0.5 | nd | 0.5 |
| δ-2-carene | 1001 | tr | tr | nd | nd |
| δ-3-carene | 1008 | 8.8 | 7.4 | 0.6 | 1.7 |
| α-terpinene | 1014 | 0.1 | 0.1 | nd | 0.1 |
| p-cymene | 1020 | 0.1 | 0.1 | tr | tr |
| 1-p-menthene | 1021 | tr | tr | nd | nd |
| o-cymene | 1022 | 0.3 | 0.4 | 0.6 | 1.0 |
| limonene | 1024 | 1.6 | 1.7 | 0.5 | 0.6 |
| β-phellandrene | 1025 | tr | tr | nd | tr |
| 1,8-cineole | 1026 | 0.1 | 0.1 | nd | nd |
| (Z)-β-ocimene | 1032 | 0.2 | 0.1 | nd | 0.1 |
| (E)-β-ocimene | 1044 | tr | tr | nd | nd |
| γ-terpinene | 1054 | 0.1 | 0.1 | tr | 0.4 |
| cis-sabinene hydrate | 1065 | tr | tr | nd | nd |
| m-cymenene | 1082 | nd | tr | nd | nd |
| terpinolene | 1086 | 0.4 | 0.3 | 0.9 | 2.4 |
| linalool | 1095 | tr | tr | nd | nd |
| n-nonanal | 1100 | nd | nd | nd | 0.2 |
| α-campholenal | 1122 | 0.2 | 0.3 | 1.5 | 3.3 |
| trans-pinocarveol | 1135 | 0.2 | 0.1 | 1.4 | 0.8 |
| cis-verbenol | 1137 | 0.1 | 0.1 | nd | nd |
| trans-verbenol | 1140 | 0.3 | 0.3 | 0.7 | 0.3 |
| unknown compound #1 | 1154 | 0.1 | 0.1 | 0.6 | 0.6 |
| trans-pinocamphone | 1158 | tr | 0.1 | tr | 0.5 |
| cis-pinocamphone | 1172 | 0.1 | 0.1 | tr | 0.7 |
| terpinen-4-ol | 1174 | tr | tr | tr | 0.1 |
| p-cymen-8-ol | 1179 | tr | tr | tr | tr |
| α-terpineol | 1186 | tr | tr | nd | nd |
| myrtenal | 1195 | 0.1 | 0.2 | 1.6 | 1.4 |
| ethyl octanoate | 1196 | 0.5 | 0.5 | 4.4 | 3.4 |
| verbenone | 1204 | tr | tr | 0.6 | 0.3 |
| trans-carveol | 1215 | nd | tr | nd | nd |
| thymol methyl ether | 1232 | tr | tr | tr | 0.3 |
| cumin aldehyde | 1238 | nd | nd | nd | 0.1 |
| carvone | 1239 | nd | tr | nd | nd |
| (E)-anethole | 1282 | nd | nd | 0.7 | 0.1 |
| bornyl acetate | 1284 | 0.2 | 0.2 | 6.1 | 4.4 |
| ethyl nonanoate | 1286 | nd | nd | nd | tr |
| thymol | 1289 | nd | nd | tr | nd |
| α-terpinyl acetate | 1346 | 0.1 | 0.1 | 1.1 | 0.6 |
| α-cubebene | 1348 | tr | tr | tr | 0.3 |
| α-longipinene | 1350 | tr | tr | tr | 0.9 |
| α-ylangene | 1373 | nd | nd | nd | 0.3 |
| α-copaene | 1374 | 0.4 | 0.3 | 6.5 | 4.5 |
| ethyl(4E)-decenoate | 1380 | tr | tr | 0.5 | 0.7 |
| β-bourbonene | 1387 | 0.7 | 0.6 | 11.3 | 3.3 |
| sativene | 1390 | 0.1 | tr | 1.1 | 0.1 |
| ethyl decenoate | 1395 | tr | tr | tr | tr |
| longifolene | 1407 | 0.4 | 0.4 | 13.3 | 8.9 |
| (E)-caryophyllene | 1417 | tr | tr | 0.3 | 0.5 |
| β-ylangene | 1419 | 0.1 | 0.1 | 0.5 | 0.7 |
| β-copaene | 1430 | 0.1 | 0.1 | 1.4 | 0.6 |
| isogermacrene D | 1445 | tr | tr | 0.9 | 0.3 |
| α-humulene | 1452 | nd | nd | tr | 0.4 |
| γ-muurolene | 1478 | tr | tr | 1.3 | 1.0 |
| germacrene D | 1480 | 0.2 | 0.1 | 0.7 | 9.9 |
| epi-cubebol | 1493 | tr | tr | 0.8 | 0.7 |
| α-muurolene | 1500 | 0.1 | 0.1 | 3.8 | 2.8 |
| γ-cadinene | 1513 | tr | tr | 0.1 | 0.8 |
| cubebol | 1514 | tr | tr | 0.1 | 0.8 |
| δ-cadinene | 1522 | 0.1 | 0.1 | 3.3 | 2.4 |
| α-calacorene | 1544 | nd | nd | 0.3 | 0.3 |
| longicamphenylone | 1562 | nd | nd | tr | 0.3 |
| unknown compound #2 | 1564 | nd | nd | 1.1 | 1.0 |
| caryophyllene oxide | 1582 | nd | nd | 1.0 | 1.2 |
| 6,10-epoxy-7(14)-isodaucene | 1586 | nd | nd | tr | 0.3 |
| β-copaen-4α-ol | 1590 | nd | nd | tr | 0.6 |
| unknown compound #3 | 1594 | nd | nd | 0.6 | 0.5 |
| salvial-4(14)en-1-one | 1594 | nd | nd | 0.7 | 0.8 |
| cedrol | 1600 | nd | nd | 0.6 | 0.4 |
| unknown compound #4 | 1624 | nd | nd | 0.9 | 0.9 |
| unknown compound #5 | 1667 | nd | nd | 0.6 | 0.9 |
| unknown compound #6 | 1691 | nd | nd | nd | 0.5 |
| ent-germacra-4(15),5,10(14)trien-β-ol | 1699 | nd | nd | 1.0 | 0.8 |
| unknown compound #7 | 1901 | nd | nd | 2.6 | 1.0 |
| unknown compound #8 | 1935 | nd | nd | 1.2 | 0.9 |
| manool oxide | 1987 | nd | nd | 1.8 | 0.9 |
| unknown compound #9 | 1998 | nd | nd | 1.2 | 0.6 |
| 18-norabieta-8,11,13-triene | 2036 | tr | tr | 4.9 | 2.8 |
| unknown compound #10 | 2061 | nd | nd | 0.5 | 0.2 |
| unknown compound #11 | 2064 | nd | nd | 1.3 | 0.9 |
| abietadiene | 2087 | nd | nd | 1.1 | 1.4 |
| unknown compound #12 | 2106 | nd | nd | 0.9 | 0.6 |
| unknown compound #13 | 2160 | nd | nd | 0.6 | 0.5 |
| unknown compound #14 | 2201 | nd | nd | 0.9 | 0.9 |
| unknown compound #15 | 2239 | nd | nd | 0.4 | 0.8 |
| unknown compound #16 | 2280 | nd | nd | nd | 0.5 |
| unknown compound #17 | 2291 | nd | nd | 0.4 | 0.5 |
| neoabietal | 2319 | nd | nd | 1.2 | 0.9 |
Table 3.
Summarized GC/FID profile (area%) of volatile fraction extracted from Pinus edulis Engelm. seed cone resin, resinous, from all months (January–December). KI is the Kovat’s Index using a linear calculation on the DB-5 column [22], and those in bold font were calculated using an alkane standard. Relative area percentage is determined with GC/FID. On average, 92.9% of the sample profiles are quantified in the table.
Table 3.
Summarized GC/FID profile (area%) of volatile fraction extracted from Pinus edulis Engelm. seed cone resin, resinous, from all months (January–December). KI is the Kovat’s Index using a linear calculation on the DB-5 column [22], and those in bold font were calculated using an alkane standard. Relative area percentage is determined with GC/FID. On average, 92.9% of the sample profiles are quantified in the table.
| Compound Name | KI | Jan. | Feb. | Mar. | Apr. | May | Jun. | Jul. | Aug. | Sept. | Oct. | Nov. | Dec. |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| α-pinene | 932 | 75.2 | 77.4 | 77.4 | 72.0 | 77.0 | 75.1 | 76.1 | 76.6 | 76.0 | 75.0 | 75.5 | 74.4 |
| camphene | 946 | 0.9 | 0.9 | 0.9 | 0.9 | 0.9 | 0.9 | 1.0 | 1.0 | 1.0 | 1.0 | 1.1 | 1.0 |
| thuja-2,4(10)diene | 953 | 0.4 | 0.5 | 0.5 | 0.8 | 0.5 | 0.8 | 0.9 | 1.1 | 1.4 | 1.2 | 1.4 | 1.4 |
| 3,7,7-trimethyl-1,3,5-cycloheptatriene | 966 | 0.5 | 0.5 | 0.5 | 0.6 | 0.5 | 0.6 | 0.7 | 0.8 | 0.8 | 1.0 | 0.9 | 1.0 |
| sabinene | 969 | 2.2 | 1.8 | 1.4 | 1.9 | 2.0 | 1.7 | 1.5 | 1.5 | 1.4 | 1.3 | 1.3 | 1.3 |
| β-pinene | 974 | 2.5 | 3.2 | 2.9 | 3.2 | 2.8 | 2.9 | 2.9 | 2.7 | 3.0 | 2.5 | 2.7 | 2.6 |
| myrcene | 988 | 0.6 | 0.4 | 0.4 | 0.7 | 0.4 | 0.6 | 0.5 | 0.8 | 0.3 | 0.3 | 0.2 | 0.2 |
| δ-3-carene | 1008 | 8.8 | 7.5 | 8.0 | 9.2 | 7.4 | 7.6 | 7.4 | 6.8 | 6.1 | 7.5 | 5.8 | 7.2 |
| limonene | 1024 | 1.6 | 1.8 | 1.7 | 2.0 | 1.9 | 2.0 | 1.7 | 1.7 | 1.4 | 1.7 | 1.8 | 2.2 |
| β-bourbonene | 1387 | 0.7 | 0.6 | 0.5 | 0.8 | 0.5 | 0.6 | 0.6 | 0.5 | 0.7 | 0.6 | 0.7 | 0.7 |
Table 4.
Summarized GC/FID profile (area%) of volatile fraction extracted from Pinus edulis Engelm. seed cone resin, non-resinous, from all months (January–December). Compounds not detected in a sample are denoted as not detected (nd). KI is the Kovat’s Index using a linear calculation on the DB-5 column [22], and those in bold font were calculated using an alkane standard. Relative area percentage is determined with GC/FID. On average, 59.9% of the sample profiles are quantified in the table.
Table 4.
Summarized GC/FID profile (area%) of volatile fraction extracted from Pinus edulis Engelm. seed cone resin, non-resinous, from all months (January–December). Compounds not detected in a sample are denoted as not detected (nd). KI is the Kovat’s Index using a linear calculation on the DB-5 column [22], and those in bold font were calculated using an alkane standard. Relative area percentage is determined with GC/FID. On average, 59.9% of the sample profiles are quantified in the table.
| Compound Name | KI | Jan. | Feb. | Mar. | Apr. | May | Jun. | Jul. | Aug. | Sept. | Oct. | Nov. | Dec. |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| α-pinene | 932 | 3.6 | 0.0 | 2.6 | 0.0 | 0.3 | 0.0 | 2.7 | 20.8 | 23.9 | 47.6 | 15.5 | 58.8 |
| thuja-2,4(10)diene | 953 | 0.0 | nd | 0.3 | nd | nd | nd | 0.3 | 2.3 | 2.6 | 2.2 | 2.1 | 3.3 |
| β-pinene | 974 | 0.0 | nd | 0.2 | nd | nd | nd | 0.3 | 1.5 | 1.2 | 2.9 | 0.9 | 2.1 |
| δ-3-carene | 1008 | 0.6 | nd | 0.6 | nd | nd | 0.0 | 1.7 | 4.7 | 4.3 | 11.8 | 9.5 | 7.0 |
| limonene | 1024 | 0.5 | nd | 0.3 | 0.7 | 0.3 | 0.0 | 0.6 | 1.9 | 3.4 | 3.2 | 2.7 | 1.5 |
| terpinolene | 1086 | 0.9 | 0.7 | 0.8 | 0.0 | 0.9 | 0.3 | 2.4 | 3.6 | 3.7 | 2.2 | 3.8 | 1.5 |
| α-campholenal | 1122 | 1.5 | 1.3 | 1.4 | 1.0 | 1.9 | 1.0 | 3.3 | 3.1 | 3.0 | 1.0 | 3.4 | 1.3 |
| ethyl octanoate | 1196 | 4.4 | 1.1 | 6.0 | 0.7 | 1.1 | 0.8 | 3.4 | 3.8 | 4.6 | 1.2 | 4.4 | 0.7 |
| bornyl acetate | 1284 | 6.1 | 4.9 | 10.1 | 2.2 | 4.5 | 3.0 | 4.4 | 3.7 | 3.9 | 1.1 | 3.4 | 1.1 |
| α-copaene | 1374 | 6.5 | 2.9 | 5.8 | 2.7 | 3.2 | 2.3 | 4.5 | 3.1 | 2.8 | 1.6 | 2.9 | 1.0 |
| β-bourbonene | 1387 | 11.3 | 5.5 | 11.5 | 5.8 | 3.8 | 5.0 | 3.3 | 4.6 | 3.8 | 1.8 | 4.4 | 0.9 |
| longifolene | 1407 | 13.3 | 10.4 | 12.4 | 10.5 | 10.1 | 8.4 | 8.9 | 7.9 | 8.3 | 4.4 | 8.1 | 2.9 |
| germacrene D | 1480 | 0.7 | 2.7 | 0.5 | 6.6 | 2.1 | 3.1 | 9.9 | 1.7 | 0.6 | 1.6 | 1.1 | 0.6 |
| α-muurolene | 1500 | 3.8 | 3.8 | 3.4 | 3.7 | 2.5 | 2.6 | 2.8 | 1.3 | 1.1 | 0.5 | 1.3 | 0.4 |
| δ-cadinene | 1522 | 3.3 | 3.8 | 2.2 | 3.8 | 2.5 | 2.4 | 2.4 | 1.1 | 0.9 | 0.4 | 1.1 | 0.4 |
| unknown compound #7 | 1901 | 2.6 | 2.3 | 2.3 | 2.1 | 1.7 | 3.1 | 1.0 | 0.4 | 0.3 | 0.0 | 0.5 | 0.1 |
| 18-norabieta-8,11,13-triene | 2036 | 4.9 | 5.0 | 3.7 | 4.9 | 5.1 | 6.3 | 2.8 | 1.0 | 0.6 | 0.1 | 1.1 | 0.3 |
Table 5.
Groups of insect/arthropod inquiline community found in cones from the 12-month study period. Lowest taxonomic unit, rank of unit, and order are noted.
Table 5.
Groups of insect/arthropod inquiline community found in cones from the 12-month study period. Lowest taxonomic unit, rank of unit, and order are noted.
| Taxa | Taxonomic Resolution | Order | Notes |
|---|---|---|---|
| Conophthorus | Genus | Coleoptera | Often damage whole cones |
| Megastigmus | Genus | Hymenoptera | Develops in seeds |
| Cecidomyiidae | Family | Diptera | Many species reported; develops in seeds |
| Eucosma | Genus | Lepidoptera | Diverse group; larvae damages whole cones |
| Spiders | Order | Araneae | Likely using cones for shelter and foraging |
| Unknown | Unranked | Various | Rare in study (6 individuals); likely prey remains or, in one case, a hyperparasitoid |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wilson, T.M.; Rotter, M.C.; Ziebarth, E.A.; Carlson, R.E. Volatile Compound Chemistry and Insect Herbivory: Pinus edulis Engelm. (Pinaceae) Seed Cone Resin. Forests 2023, 14, 1862. https://doi.org/10.3390/f14091862
AMA Style
Wilson TM, Rotter MC, Ziebarth EA, Carlson RE. Volatile Compound Chemistry and Insect Herbivory: Pinus edulis Engelm. (Pinaceae) Seed Cone Resin. Forests. 2023; 14(9):1862. https://doi.org/10.3390/f14091862
Chicago/Turabian StyleWilson, Tyler M., Michael C. Rotter, Emma A. Ziebarth, and Richard E. Carlson. 2023. "Volatile Compound Chemistry and Insect Herbivory: Pinus edulis Engelm. (Pinaceae) Seed Cone Resin" Forests 14, no. 9: 1862. https://doi.org/10.3390/f14091862
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.