
Assessing the Potential of Urban Trees to Accumulate Potentially Toxic Elements: A Network Approach

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling and Analysis
2.3. Network Structure and Centrality Measures Analysis
3. Results and Discussion
3.1. The Potential of Urban Trees for the Accumulation of PTEs
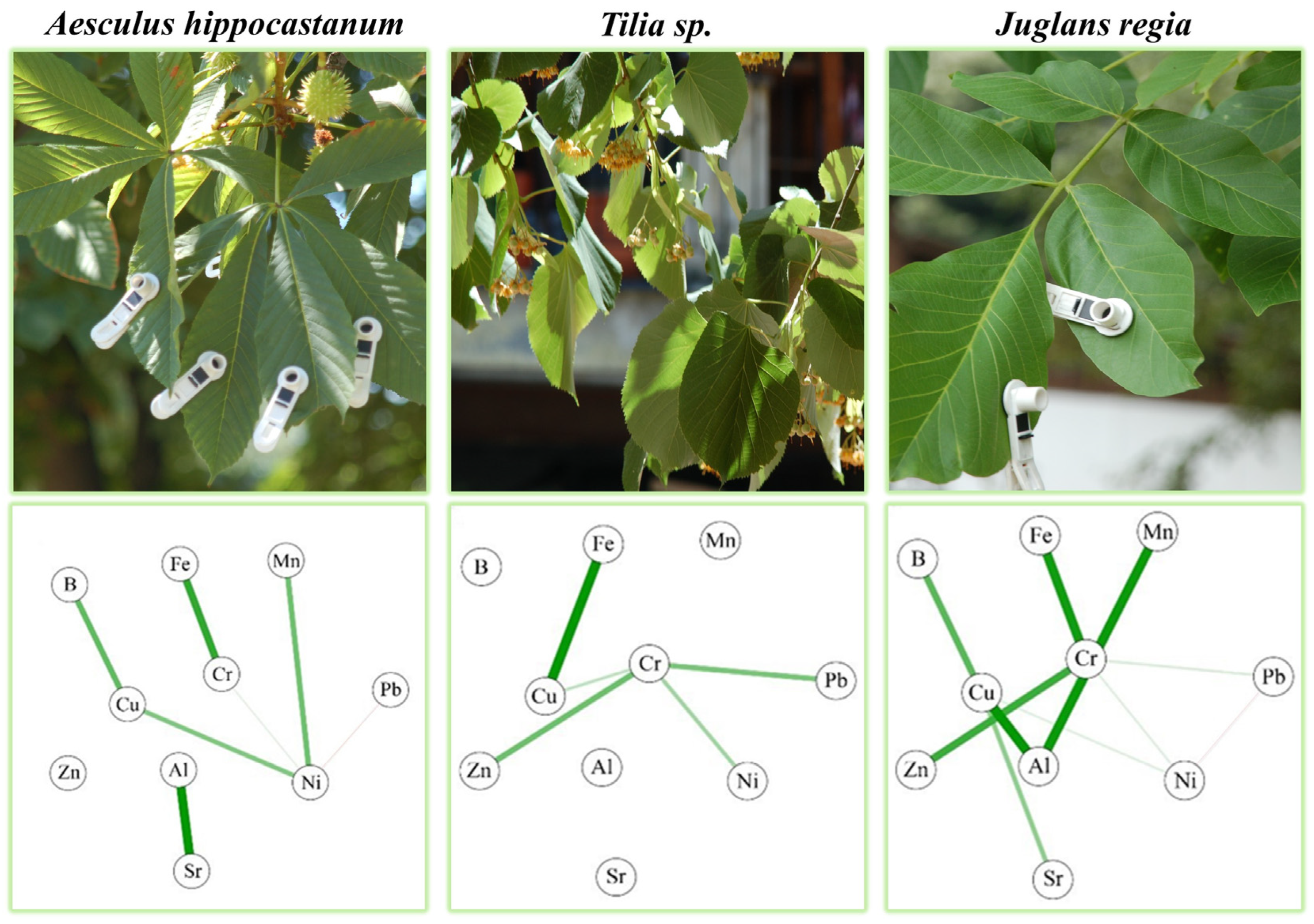
3.2. Associations between PTEs in Leaf Samples Using Network Analysis
3.3. Ecophysiological Response of Plants to PTEs
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Baklanov, A.; Molina, L.T.; Gauss, M. Megacities, air quality and climate. Atmos. Environ. 2016, 126, 235–249. [Google Scholar] [CrossRef]
- Locosselli, G.M.; Buckeridge, M.S. The science of urban trees to promote well-being. Trees 2023, 37, 1–7. [Google Scholar] [CrossRef]
- Arnfield, A.J. Two decades of urban climate research: A review of turbulence, exchanges of energy and water, and the urban heat island. Int. J. Climatol. 2003, 23, 1–26. [Google Scholar] [CrossRef]
- Akbari, H.; Konopacki, S. Calculating energy-saving potentials of heat-island reduction strategies. Energy Policy 2005, 33, 721–756. [Google Scholar] [CrossRef]
- Biasioli, M.; Barberis, R.; Ajmone-Marsan, F. The influence of a large city on some soil properties and metals content. Sci. Total Environ. 2006, 356, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Madlener, R.; Sunak, Y. Impacts of urbanization on urban structures and energy demand: What can we learn for urban energy planning and urbanization management? Sustain. Cities Soc. 2011, 1, 45–53. [Google Scholar] [CrossRef]
- Galitskova, Y.M.; Antonina, I.; Murzayeva, A.I. Urban Soil Contamination. Procedia Eng. 2016, 153, 162–166. [Google Scholar] [CrossRef]
- Bielicka-Giełdoń, A.; Rylko, E.; Zamojc, K. Distribution, bioavailability and fractionation of metallic elements in Allotment garden soils using the BCR sequential extraction procedure. Pol. J. Environ. Stud. 2013, 22, 1013–1021. [Google Scholar]
- Hamad, S.H.; Schauer, J.J.; Shafer, M.M.; Al-Rheem, E.A.; Skaar, P.S.; Heo, J.; Tejedor-Tejedor, I. Risk assessment of total and bioavailable potentially toxic elements (PTEs) in urban soils of Baghdad-Iraq. Sci. Total Environ. 2014, 494–495, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Jiao, X.; Teng, Y.; Zhan, Y.; Wu, J.; Lin, X. Soil heavy metal pollution and risk assessment in Shenyang industrial district, Northeast China. PLoS ONE 2015, 10, e0127736. [Google Scholar] [CrossRef]
- Mitrović, M.; Blanusa, T.; Pavlović, M.; Pavlović, D.; Kostić, O.; Perović, V.; Jarić, S.; Pavlović, P. Using Fractionation Profile of Potentially Toxic Elements in Soils to Investigate Their Accumulation in Tilia sp. Leaves in Urban Areas with Different Pollution Levels. Sustainability 2021, 13, 9784. [Google Scholar] [CrossRef]
- Edmondson, J.L.; Stott, I.; Davies, Z.G.; Gaston, K.J.; Leake, J.R. Soil surface temperatures reveal moderation of the urban heat island effect by trees and shrubs. Sci. Rep. 2016, 6, 33708. [Google Scholar] [CrossRef] [PubMed]
- Miyahara, A.A.L.; Wild, T.; Sandre, A.A.; Pellegrino, P.R.M.; Filho, C.A.S.; Buckeridge, M.S.; Locosselli, G.M. Developing and classifying urban biomes as a basis for nature-based solutions. Urban Clim. 2022, 45, 101251. [Google Scholar] [CrossRef]
- Kandziora-Ciupa, M.; Ciepał, R.; Nadgórska-Socha, A.; Barczik, G. Accumulation of heavy metals and antioxidant responses in Pinus sylvestris L. needles in polluted and non-polluted sites. Ecotoxicology 2016, 25, 970–981. [Google Scholar] [CrossRef] [PubMed]
- Gajić, G.; Đurđević, L.; Kostić, O.; Jarić, S.; Mitrović, M.; Stevanović, B.; Pavlović, P. Assessment of the phytoremediation potential and an adaptive response of Festuca rubra L. sown on fly ash deposits: Native grass has a pivotal role in ecorestoration management. Ecol. Eng. 2016, 93, 250–261. [Google Scholar] [CrossRef]
- Sen, A.; Khan, I.; Kundu, D.; Das, K.; Datta, J.A. Ecophysiological evaluation of tree species for biomonitoring of air quality and identification of air pollution-tolerant species. Environ. Monit. Assess. 2017, 189, 262. [Google Scholar] [CrossRef] [PubMed]
- Shi, T.; He, L.; Wang, R.; Li, Z.; Hu, Z.; Wu, G. Digital mapping of heavy metals in urban soils: A review and research challenges. Catena 2023, 228, 107183. [Google Scholar] [CrossRef]
- Čakmak, D.; Pavlović, P.; Mrvić, V.; Saljnikov, E.; Perović, V.; Jaramaz, D.; Sikirić, B. Using different receptor models to determine the sources of available forms of potentially toxic elements in Rasina District—A case study. Catena 2023, 222, 106865. [Google Scholar] [CrossRef]
- Wadoux, A.M.J.-C.; Minasny, B.; McBratney, A.B. Machine learning for digital soil mapping: Applications, challenges and suggested solutions. Earth-Sci. Rev. 2020, 210, 103359. [Google Scholar] [CrossRef]
- Paes, É.C.; Veloso, G.V.; Silva, D.L.d.A.; Fernandes-Filho, E.I.; Fontes, M.P.F.; Soares, E.M.B. Use of modeling to map potentially toxic elements and assess the risk to human health in soils affected by mining activity. Catena 2023, 220, 106662. [Google Scholar] [CrossRef]
- Barberán, A.; Bates, S.; Casamayor, E.; Fierer, N. Using network analysis to explore co-occurrence patterns in soil microbial communities. ISME J. 2012, 6, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Berry, D.; Widder, S. Deciphering microbial interactions and detecting keystone species with co-occurrence networks. Front. Microbiol. 2014, 5, 219. [Google Scholar] [CrossRef] [PubMed]
- Ofaim, S.; Ofek-Lalzar, M.; Sela, N.; Jinag, J.; Kashi, Y.; Minz, D.; Freilich, S. Analysis of microbial functions in the rhizosphere using a metabolic-network based framework for metagenomics interpretation. Front. Microbiol. 2017, 8, 1606. [Google Scholar] [CrossRef] [PubMed]
- Hewavitharana, S.S.; Klarer, E.; Reed, A.J.; Leisso, R.; Poirier, B.; Honaas, L.; Rudell, D.R.; Mazzola, M. Temporal dynamics of the soil metabolome and microbiome during simulated anaerobic soil disinfestation. Front. Microbiol. 2019, 10, 2365. [Google Scholar] [CrossRef] [PubMed]
- Raiesi, F.; Beheshti, A. Evaluating forest soil quality after deforestation and loss of ecosystem services using network analysis and factor analysis techniques. Catena 2022, 208, 105778. [Google Scholar] [CrossRef]
- Martín-Sanz, J.P.; de Santiago-Martín, A.; Valverde-Asenjo, I.; Quintana-Nieto, J.R.; González-Huecas, C.; López-Lafuente, A.L. Comparison of soil quality indexes calculated by network and principal component analysis for carbonated soils under different uses. Ecol. Ind. 2022, 143, 109374. [Google Scholar] [CrossRef]
- Zahedifar, M. Assessing alteration of soil quality, degradation, and resistance indices under different land uses through network and factor analysis. Catena 2023, 222, 106807. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, J.; Li, B. Determining the influence factors of soil organic carbon stock in opencast coal-mine dumps based on complex network theory. Catena 2019, 173, 433–444. [Google Scholar] [CrossRef]
- Pavlović, D.; Pavlović, M.; Perović, V.; Mataruga, Z.; Čakmak, D.; Mitrović, M.; Pavlović, P. Chemical Fractionation, Environmental, and Human Health Risk Assessment of Potentially Toxic Elements in Soil of Industrialised Urban Areas in Serbia. Int. J. Environ. Res. Public Health 2021, 18, 9412. [Google Scholar] [CrossRef] [PubMed]
- Secretariat for Environmental Protection. Environmental Protection Programme for the City of Belgrade; Secretariat for Environmental Protection, Kirilo Savić Institute: Belgrade, Serbia, 2013; pp. 1–241. (In Serbian) [Google Scholar]
- Secretariat for Environmental Protection. The Quality of the Environment in the City of Belgrade in 2012; The City Institute for Public Health: Belgrade, Serbia, 2013; pp. 1–372. (In Serbian) [Google Scholar]
- Smailagić, J.; Savović, A.; Nešić, D.; Milenković, M.; Zdravković, S. Climatological Analysis of Summer 2012 for Serbia, Seasonal Bulletin; Republic Hydrometeorological Service of Serbia: Belgrade, Serbia, 2012; pp. 1–22. (In Serbian)
- Piczak, K.; Lesniewicz, A.; Zyrnicki, W. Metal concentrations in deciduous tree leaves from urban areas in Poland. Environ. Monit. Assess. 2003, 86, 273–287. [Google Scholar] [CrossRef]
- Yanqun, Z.; Yuan, L.; Schvartz, C.; Langlade, L.; Fan, L. Accumulation of Pb, Cd, Cu and Zn in plants and hyperaccumulator choice in Lanping lead-zinc mine area, China. Environ. Int. 2004, 30, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Rossini Oliva, S.; Mingorance, M.D. Study of the impact of industrial emission on the vegetation grown around Huelva (South of Spain) City. J. Atmos. Chem. 2004, 49, 291–302. [Google Scholar] [CrossRef]
- Serbula, S.M.; Radojevic, A.A.; Kalinovic, J.V.; Kalinovic, T.S. Indication of airborne pollution by birch and spruce in the vicinity of copper smelter. Environ. Sci. Pollut. Res. 2014, 21, 11510–11520. [Google Scholar] [CrossRef]
- Kalinovic, T.S.; Serbula, S.M.; Radojevic, A.A.; Kalinovic, J.V.; Steharnik, M.M.; Petrovic, J.V. Elder, linden and pine biomonitoring ability of pollution emitted from the copper smelter and the tailings ponds. Geoderma 2016, 262, 266–275. [Google Scholar] [CrossRef]
- USEPA (United States Enviromnental Protection Agency). Method 3052: Microwave assisted acid digestion of siliceous and organically based matrices. In Test Methods for Evaluating Solid Waste; SW 846; USEPA: Washington, DC, USA, 1996. [Google Scholar]
- Krause, G.; Weis, E. Chlorophyll fluorescence and photosynthesis: The basics. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1991, 42, 313–349. [Google Scholar] [CrossRef]
- Arnon, D. I Copper enzymes in isolated chloroplasts. Polyphenoloxidases in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Robinaugh, D.J.; Millner, A.J.; McNally, R.J. Identifying highly influential nodes in the complicated grief network. J. Abnorm. Psychol. 2016, 125, 747–757. [Google Scholar] [CrossRef] [PubMed]
- Epskamp, S.; Borsboom, D.; Fried, E.I. Estimating psychological networks and their accuracy: A tutorial paper. Behav. Res. 2018, 50, 195–212. [Google Scholar] [CrossRef]
- Kabata-Pendias, A.; Mukherjee, A.B. Trace Elements from Soil to Human; Springer: Berlin, Germany, 2007; pp. 1–550. [Google Scholar] [CrossRef]
- Shacklette, H.T.; Erdman, J.A.; Harms, T.F. Trace elements in plants foodstuffs. In Toxicity of Heavy Metals in the Environment; Oehme, F.W., Ed.; Part I; Marcel Dekker Inc.: New York, NY, USA, 1978; p. 25. [Google Scholar]
- Greksa, A.; Ljevnaić-Mašić, B.; Grabić, J.; Benka, P.; Radonić, V.; Blagojević, B.; Sekulić, M. Potential of urban trees for mitigating heavy metal pollution in the city of Novi Sad, Serbia. Environ. Monit. Assess. 2019, 191, 636. [Google Scholar] [CrossRef] [PubMed]
- Jia, M.; Zhou, D.; Lu, S.; Yu, J. Assessment of foliar dust particle retention and toxic metal accumulation ability of fifteen roadside tree species: Relationship and mechanism. Atmos. Pollut. Res. 2021, 12, 36–45. [Google Scholar] [CrossRef]
- Aničić, M.; Spasić, T.; Tomašević, M.; Rajšić, S.; Tasić, M. Trace elements accumulation and temporal trends in leaves of urban deciduous trees (Aesculus hippocastanum and Tilia spp.). Ecol. Indic. 2011, 11, 824–830. [Google Scholar] [CrossRef]
- Wang, J.; Huang, Y.; Beiyuan, J.; Wei, X.; Qi, J.; Wang, L.; Fang, F.; Liu, J.; Cao, J.; Xiao, T. Thallium and potentially toxic elements distribution in pine needles, tree rings and soils around a pyrite mine and indication for environmental pollution. Sci. Total Environ. 2022, 828, 154346. [Google Scholar] [CrossRef] [PubMed]
- Petrova, S.; Yurukova, L.; Velcheva, I. Possibilities of using deciduous tree species in trace element biomonitoring in an urban area (Plovdiv, Bulgaria). Atmos. Pollut. Res. 2014, 5, 196–202. [Google Scholar] [CrossRef]
- Kosiorek, M.; Modrzewska, B.; Wyszkowski, M. Levels of selected trace elements in Scots pine (Pinus sylvestris L.), silver birch (Betula pendula L.), and Norway maple (Acer platanoides L.) in an urbanized environment. Environ. Monit. Assess. 2016, 188, 598–610. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Kapoor, D.; Wang, J.; Shahzad, B.; Kumar, V.; Bali, A.S.; Jasrotia, S.; Zheng, B.; Yuan, H.; Yan, D. Chromium Bioaccumulation and Its Impacts on Plants: An Overview. Plants 2020, 9, 100. [Google Scholar] [CrossRef] [PubMed]
- Alloway, B.J. Heavy metals in soils. In Trace Metals and Metalloids in Soils and Their Bioavailability, 3rd ed.; Springer: New York, NY, USA, 2013; pp. 1–613. [Google Scholar]
- Marschner, H. Mineral Nutrition of Higher Plants, 2nd ed.; Academic Press: New York, NY, USA, 1995; pp. 1–889. [Google Scholar]
- Ofoe, R.; Thomas, R.H.; Asiedu, S.K.; Wang-Pruski, G.; Fofana, B.; Abbey, L. Aluminum in plant: Benefits, toxicity and tolerance mechanisms. Front. Plant Sci. 2023, 13, 1085998. [Google Scholar] [CrossRef] [PubMed]
- Kabata-Pendias, A.; Pendias, H. Trace Elements in Soils and Plants, 3rd ed.; CRC Press LLC: Boca Raton, FL, USA, 2001; pp. 1–413. [Google Scholar]
- Moyen, C.; Roblin, G. Uptake and translocation of strontium in hydroponically grown maize plants, and subsequent effects on tissue ion content, growth and chlorophyll a/b ration: Comparison with Ca effects. Environ. Exp. Bot. 2010, 68, 247–257. [Google Scholar] [CrossRef]
- Li, L.; Zhang, K.; Gill, R.A.; Islam, F.; Farooq, M.A.; Wang, J.; Zhou, W. Ecotoxicological and Interactive Effects of Copper and Chromium on Physiochemical, Ultrastructural, and Molecular Profiling in Brassica napus L. Biomed Res. Int. 2018, 2018, 9248123. [Google Scholar] [CrossRef]
- Amari, T.; Ghnaya, T.; Abdelly, C. Nickel, cadmium and lead phytotoxicity and potential of halophytic plants in heavy metal extraction. S. Afr. J. Bot. 2017, 111, 99–110. [Google Scholar] [CrossRef]
- Joshi, N.; Chauhan, A.; Joshi, P.C. Impact of industrial air pollutants on some biochemical parameters and yield in wheat and mustard plants. Environmentalist 2009, 29, 398–404. [Google Scholar] [CrossRef]
- Seyyednejad, S.M.; Niknejad, M.; Koochak, H. A review of some different effects of air pollution on plants. Res. J. Environ. Sci. 2011, 5, 302–309. [Google Scholar] [CrossRef]
- Smirnoff, N. Ascorbate, tocopherol and carotenoids: Metabolism, pathway engineering and functions. In Antioxidants and Reactive Oxygen Species in Plants; Smirnoff, N., Ed.; Blackwell Publishing: Oxford, UK, 2005; pp. 53–87. [Google Scholar]
- Yruela, I. Copper in plants. Braz. J. Plant Physiol. 2005, 17, 145–156. [Google Scholar] [CrossRef]
- Broadley, M.R.; White, P.J.; Hammond, J.P.; Zelko, I.; Lux, A. Zinc in plants. New Phytol. 2007, 173, 677–702. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, N.; Alatar, A.A.; Faisal, M.; Khan, M.I.; Fatima, N.; Anis, M.; Hegazy, A.K. Effect of copper and zinc on the in vitro regeneration of Rauvolfia serpentina. Biol. Plant 2015, 59, 11–17. [Google Scholar] [CrossRef]
- Grotz, N.; Guerinot, M.L. Molecular aspects of Cu, Fe and Zn homeostasis in plants. Biochim. Biophys. Acta—Mol. Cell Res. 2006, 1763, 595–608. [Google Scholar] [CrossRef] [PubMed]
- Briat, J.F.; Duc, C.; Ravet, K.; Gaymard, F. Ferritins and iron storage in plants. Biochim. Biophys. Acta—Mol. Cell Res. 2010, 1800, 806–814. [Google Scholar] [CrossRef] [PubMed]
- Demming-Adams, B.; Adams, W.W. Photoprotection and other responses of plants to high light stress. Annu. Rev. Plant. Physiol. Plant. Mol. Biol. 1992, 43, 99–626. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence-A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Boussac, A.; Rappaport, F.; Carrier, P.; Verbavatz, J.M.; Gobin, R.; Kirilovsky, D.; Rutherford, A.W.; Sugiura, M. Biosynthetic Ca2+/Sr2+ exchange in the photosystem II oxygenevolving enzyme of Thermosynechococcus elongatus. J. Biol. Chem. 2004, 279, 22809–22819. [Google Scholar] [CrossRef] [PubMed]
- Pavlović, M.; Pavlović, D.; Kostić, O.; Jarić, S.; Čakmak, D.; Pavlović, P.; Mitrović, M. Evaluation of urban contamination with trace elements in city parks in Serbia using pine (Pinus nigra Arnold) needles, bark and urban topsoil. Int. J. Environ. Res. 2017, 11, 625–639. [Google Scholar] [CrossRef]
- Schmidt, S.B.; Jensen, P.E.; Husted, S. Review: Manganese deficiency in plants: The impact on photosystem II. Trends Plant Sci. 2016, 21, 622–623. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthalter, H.K.; Buschmann, C. Chlorophylls and carotenoids—Measurement and characterisation by UV-VIS Spectroscopy. In Currenent Protocols in Food Analytical Chemistry; Wrolstad, R.E., Acree, T.E., An, H., Decker, E.A., Penner, M.H., Reid, D.S., Schwartz, S.J., Shoemaker, C.F., Sporns, P., Eds.; John Wiley and Sons: New York, NY, USA, 2001; Volume 1, pp. F4.3.1–F4.3.8. [Google Scholar]
- Sumalan, R.L.; Nescu, V.; Berbecea, A.; Sumalan, R.M.; Crisan, M.; Negrea, P.; Ciulca, S. The Impact of Heavy Metal Accumulation on Some Physiological Parameters in Silphium perfoliatum L. Plants Grown in Hydroponic Systems. Plants 2023, 12, 1718. [Google Scholar] [CrossRef] [PubMed]
- Beneragama, C.K.; Goto, K. Chlorophyl a:b ratio increases under low-light in ‘shade tolerant’ Euglena gracilis. Trop. Agric. Res. 2010, 22, 12–25. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PTEs | A. platanoides | A. pseudoplatanus | B. pendula | A. hippocastanum | Tilia sp. | J. regia | P. acerifolia | P. nigra I | P. nigra II |
|---|---|---|---|---|---|---|---|---|---|
| Al | 0.778 | −0.698 | 0.817 | 1.047 | −0.769 | 1.276 | 0.580 | 0.938 | 1.887 |
| B | 0.234 | −1.336 | −0.178 | −0.883 | −1.108 | −0.673 | −0.220 | −0.921 | −1.311 |
| Cr | −0.936 | 1.208 | 2.357 | 1.064 | 2.160 | 1.569 | 0.140 | 0.363 | 0.277 |
| Cu | 0.891 | 2.032 | 0.344 | 0.888 | 0.900 | 1.331 | 1.933 | −0.700 | 1.245 |
| Fe | −0.239 | 0.052 | −0.933 | −0.607 | 0.490 | −0.311 | −0.526 | −0.724 | −0.277 |
| Mn | 1.406 | −0.662 | −0.727 | −0.368 | −0.565 | −0.156 | −0.524 | −0.179 | −0.680 |
| Ni | −0.319 | −0.110 | −0.717 | 0.725 | −0.357 | −0.821 | 1.190 | 0.361 | −0.659 |
| Pb | −1.812 | −0.794 | −0.840 | −1.816 | 0.277 | −1.107 | −0.334 | −0.870 | −0.832 |
| Sr | 0.789 | 0.232 | −0.104 | 0.637 | −0.970 | −0.712 | −0.752 | −0.496 | −0.254 |
| Zn | −0.793 | 0.076 | −0.019 | −0.687 | −0.059 | −0.396 | −1.487 | 2.227 | 0.604 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matić, M.; Pavlović, D.; Perović, V.; Čakmak, D.; Kostić, O.; Mitrović, M.; Pavlović, P. Assessing the Potential of Urban Trees to Accumulate Potentially Toxic Elements: A Network Approach. Forests 2023, 14, 2116. https://doi.org/10.3390/f14112116
Matić M, Pavlović D, Perović V, Čakmak D, Kostić O, Mitrović M, Pavlović P. Assessing the Potential of Urban Trees to Accumulate Potentially Toxic Elements: A Network Approach. Forests. 2023; 14(11):2116. https://doi.org/10.3390/f14112116
Chicago/Turabian StyleMatić, Marija, Dragana Pavlović, Veljko Perović, Dragan Čakmak, Olga Kostić, Miroslava Mitrović, and Pavle Pavlović. 2023. "Assessing the Potential of Urban Trees to Accumulate Potentially Toxic Elements: A Network Approach" Forests 14, no. 11: 2116. https://doi.org/10.3390/f14112116