Arbuscular Mycorrhizal Fungi Mitigate Nitrogen Leaching under Poplar Seedlings


1
College of Forestry, Northwest A&F University, Yangling 712100, China
2
State Key Laboratory of Conservation and Utilization of Subtropical Agro-bioresources, Lingnan Guangdong Laboratory of Modern Agriculture, Guangdong Key Laboratory for Innovative Development and Utilization of Forest Plant Germplasm, College of Forestry and Landscape Architecture South China Agricultural University, Guangzhou 510642, China
3
Key Laboratory of State Forestry and Grassland Administration on Forest Ecosystem Protection and Restoration of Poyang Lake Watershed, College of Forestry, Jiangxi Agricultural University, Nanchang 330045, China
4
Department of Microbiology and Plant Biology, Center for Spatial Analysis, University of Oklahoma, Norman, OK 73019, USA
*
Author to whom correspondence should be addressed.
†
Fengru Fang and Chunyan Wang contributed equally to this work.
Forests 2020, 11(3), 325; https://doi.org/10.3390/f11030325
Submission received: 23 February 2020
/
Revised: 12 March 2020
/
Accepted: 13 March 2020
/
Published: 14 March 2020
(This article belongs to the Special Issue Role of Soil Organisms in Soil Formation and Nutrient Circulation in Forest Ecosystems)
Abstract
:The leaching of soil nitrogen (N) has become one of the most concerning environmental threats to ecosystems. Arbuscular mycorrhizal (AM) fungi have important ecological functions, however, their influence on soil N leaching and the mechanism of action remain unclear. We conducted a two-factor (N application level × AM inoculation) experiment on poplar, and for the first time, comprehensively analyzed the mechanism by which AM fungi influence soil N leaching. The results showed that, under optimum (7.5 mM) and high (20 mM) N levels, the nitrate (NO3−) and ammonium (NH4+) concentrations of leachate in the AM inoculated treatment (+AM) were lower than in the non-inoculated treatment (−AM), with significant reductions of 20.0% and 67.5%, respectively, under high N level, indicating that AM inoculation can reduce soil N leaching and that it is more effective for NH4+. The arbuscular and total colonization rates gradually increased, and the morphology of spores and vesicles changed as the N level increased. Under optimum and high N levels, +AM treatment increased the root N concentration by 11.7% and 50.7%, respectively; the increase was significant (p < 0.05) at the high N level, which was associated with slightly increased transpiration and root activity despite reductions in root surface area and root length. Additionally, the +AM treatment increased soil cation exchange capacity (CEC), soil organic carbon (SOC), and significantly (p < 0.05) increased the proportions of macroaggregates (but without significant change in microaggregates), causing soil total nitrogen (TN) to increase by 7.2% and 4.7% under optimum and high N levels, respectively. As the N levels increased, the relative contributions of AM inoculation on N leaching increased, however, the contributions of plant physiological and soil variables decreased. Among all of the variables, SOC had important contributions to NH4+ and total N in the leachate, while root N concentration had a higher contribution to NO3−. In conclusion, AM fungi can mitigate soil N leaching and lower the risk of environmental pollution via enhancing N interception by the inoculated fungi, increasing N sequestration in plant roots, and by improving soil N retention.
1. Introduction
The leaching of nutrients from ecosystems has become an important issue that threatens the global environment. Among the leached soil nutrients, nitrogen (N) is the most serious. It has been reported that only 55% of the 148.14 Tg N output from croplands is taken up by plants, and up to 16% of which is lost via leaching [1]. The leached nitrogen not only decreases soil fertility and crop yield, but it also adversely affects the environment [2,3]. Therefore, preventing excess N leaching is imperative for maintaining ecosystem productivity and sustainable development, and for reducing environmental pollution risk. It is urgent to find an effective method and to employ it scientifically in combination with other soil N leaching mitigation techniques [4,5,6,7,8] to alleviate N leaching.
Plants play an important role in mitigating soil N leaching. In addition to directly absorbing nutrients through their root system, most terrestrial plants can obtain nutrients by forming associations with microorganisms [9]. Arbuscular mycorrhiza (AM) fungi are present in various ecosystems [10,11], and can establish mutual symbiosis with about 80% of land plants [12]. They supply their hosts with large amounts of nutrients (such as N) to obtain plant assimilated carbon (C) [13], while promoting plant growth. Moreover, it has been reported that AM fungi are able to improve soil structure due to their extensive mycelia that affect soil aggregation [14] and absorb nutrients directly from the soil [15]. Finally, AM fungi may alter microbial abundance and diversity via competing with other microorganisms [16] and via the C exuded by their extraradical hyphae [17], further affecting the soil N cycle process (including mineralization, nitrification and denitrification) [16,18]. In summary, AM fungi may have important functions in alleviating soil N leaching.
There have been several studies concerning the influences of AM fungi on soil N leaching, but the results of the limited studies vary widely [9]. Van der Heijden revealed that AM fungi decreased the leaching of grassland soil NH4+ but did not reduce NO3− leaching [19]. Prior to this study, the role of AM fungi in soil N leaching had not been confirmed, but the cause of the difference in the effects of AM fungi on NH4+ and NO3− leaching was not further examined. In contrast, some research has indicated that AM inoculation decreased NO3− loss through leaching while having no impact on NH4+ leaching [20,21,22]. Moreover, Asghari and Cavagnaro found that the inoculation of AM fungi in Phalaris aquatica L. significantly reduced the leaching of both NO3− and NH4+ [23]. It has also been reported that less mineral N [24], dissolved organic N [25], and total dissolved N [26] are leached under AM fungi treatment. The above results indicate that the impacts of AM fungi on N leaching are variable and may depend on the host plant species [27], AM fungal species [28], N application rate [29], the form of N fertilizer [25], the water regime [21], and the soil type [25]. However, the relevant studies (less than 20) that could be retrieved were mainly focused on grassland and farmland ecosystems; to the best of our knowledge, no data have been published regarding the effects of AM fungi on N leaching in arbor ecosystems.
Although it is known that AM fungi can reduce N leaching loss from soil, the underlying mechanisms are still incompletely understood. de Vries et al. reported that N leaching decreased with increasing biomass of AM fungi in a large-scale field study [30]. This may be because AM fungi can enhance N interception from soil, and store substantial amounts of N in their spores and extraradical hyphae [31]. Most of the related articles have reported that the higher capacity of mycorrhizal root systems to reduce NO3−-N leaching could be partly attributed to the larger size and the higher plant N content of mycorrhizal plants [20,21,23], demonstrating that AM enhanced the capacity of plants to acquire N. Furthermore, AM fungi indirectly influenced N leaching through shifting the plant community structure. For example, Martínez-García observed that with increased abundance of forbs and legumes, and decreased abundance of grasses in the presence of AM fungi, N leaching was reduced, especially when rainfall intensity increased [24]. AM fungi might promote the soil microbial communities that immobilized N more efficiently, thus enhancing microbial biomass N content and thereby increasing NH4+ and dissolved organic N retention [25]. Moreover, mineralisation and nitrification could lead to large N leaching losses [32,33], while AM fungi are able to compete with the soil microbes involved in the two processes [9,34], subsequently impacting the abundance of those microbes. For example, Duffková et al. found that net nitrification and total aerobic N mineralization were both affected by inoculation treatment [35]. Finally, AM fungi reduced the leaching volume by increasing water uptake of plants and improving soil moisture retention [21].
Soil aggregates can be divided into microaggregates (<0.25 mm) and macroaggregates (>0.25 mm) [36] that not only influence microbial community structure but also improve soil water and nutrient retention [37]. Cation exchange capacity (CEC) reflects the ability of soil to hold exchangeable cations (such as NH4+) [38]. However, the changes in soil microaggregates and CEC after AM fungi inoculation, as well as the subsequent effects on N leaching, have not been reported. Soil N leaching in the ecosystem is determined by a combination of environmental conditions, vegetations, and soils (including soil microbes). Therefore, it is necessary to analyse this issue from a holistic perspective, and to determine the relative contribution of AM inoculation, plant physiological and morphological indicators, and soil physicochemical indicators to soil N leaching, although to date there has been no relevant research.
Poplar 107 (Populus × canadensis ‘Neva’) has been widely planted because it is easy to propagate, has strong resistance to stress, is economically efficient and plays an important ecological role [39]. N is an indispensable nutrient in the fast growth of poplars, but the frequent application of a large amount of fertilizer in forestry production has caused N leaching and environmental pollution. In addition, some studies have shown that AM fungi can improve plant N uptake and utilization efficiency [40,41,42]. In this study, we conducted a two-factor (N application level and AM fungi inoculation) pot experiment to investigate the influence of AM fungi on soil N leaching and the related mechanisms under different levels of N application. The main objectives of this study were to: (1) screen the low, optimal, and high N levels based on poplar growth indicators; (2) clarify the various effects of AM fungi on N leaching from the poplar microcosm at different N levels; (3) determine the impacts of AM inoculation on selected indicators closely related to nutrient leaching; and (4) quantify relative contributions of the AM inoculation, plant physiological and morphological indicators, and soil physicochemical indicators to N leaching under different levels of N application in order to elucidate the relevant mechanism. The findings of the present study will advance our understanding of the ecological functioning of AM fungi in N cycling.
2. Materials and Methods
2.1. Experimental Design
We carried out the pot experiment from April to August 2016, in the greenhouse at the Northwest A&F University, in Yangling (Shaanxi, China). The two-factor experimental design consisted of two factors and a total of 14 treatment combinations as follows: inoculation (inoculated with Rhizophagus irregularis, +AM or inoculated with sterile inoculum, −AM) and different levels of N application (with 0, 1.0, 5.0, 7.5, 10.0, 15.0 and 20.0 mM NH4NO3 being the concentrations in the irrigation solution). Each treatment had 30 pots of poplar tree, for a total of 420 pots. The experiment comprised two parts: in the first part, the low, optimum and high N concentrations were chosen based on the stem length and biomass; in the second part, we kept experimental materials of the low, optimum and high N treatments for the study of N leaching.
2.2. Experimental Materials
The test plant was Poplar 107 (Populus canadensis × ‘Neva’), a hybrid between Populus nigra and Populus deltoides. We collected cuttings (6 cm in length and 4 mm in diameter) from one-year-old poplar shoots with uniform thickness. The surface was sterilized for 15 s with 70% (v/v) ethanol and rinsed five times with distilled water.
The AM fungus, R. irregularis (B109) was provided by the Institute of Plant Nutrition and Resources, Beijing Academy of Agriculture and Forestry Sciences (Beijing, China). The inoculum contained spores (approximately 50 spores per gram of inoculum), mycelia, root segments and propagation substrate.
The potting soil was the 0–20 cm layer Lou soil (Eum-Orthic Anthrosols) collected from the nursery of the Northwest A&F University (Yangling, China). The soil pH was 8.3, available N content was 35.78 mg·kg−1, available phosphorus (P) content was 11.32 mg·kg−1, and available potassium (K) content was 158.56 mg·kg−1. Available N was determined by alkaline hydrolysis diffusion method [43]; available P was measured using 0·5 M NaHCO3 extraction-molybdenum antimony colorimetric method [44]; and available K was determined using 1 M NH4OAc extraction followed by flame spectrometry [44]. The soil and washed river sand were air-dried and sieved with a 2-mm sieve and mixed in a ratio of 2:1 (v/v) to serve as the potting matrix, which was then autoclaved (121 °C and 0.11 MPa) for 2 h and again air-dried. The N nutrient solutions with different concentrations of NH4NO3 were prepared according to Knoth at al. [45].
2.3. Poplar Planting and Treatment
Poplar cuttings were planted in plastic pots (9.0 cm in diameter on the top, 5.4 cm in diameter at the bottom, 13.5 cm in height) as a microcosm with 500 g of the prepared potting matrix. The pots were divided into +AM and −AM treatments. For the +AM treatment, when the cuttings were planted, 20 g of AM inoculum was inoculated into each pot; whereas, for the −AM treatment, 20 g of autoclaved AM inoculum was inoculated into each pot, and the microbial eluent (filtered with 1 µm nylon mesh) with a microbial population free of AM propagules was also included [42]. After inoculation, pot-planted poplar seedlings were cultivated in a greenhouse and were watered regularly.
After two months of growth, we examined the seedlings to make sure the roots formed infection structures. Seven N sub-treatments were established for each of the +AM and −AM treatments; 120 mL of NH4NO3 nutrient solution was applied once every two days (thoroughly watered and the nutrient solution just exuded from the bottom of the pot) for a total of 16 applications in 31 days. After fertilization, the seedlings continued growth under normal conditions and were harvested one month later for measurements.
2.4. Biochemical Measurements
Ten plants from each treatment combination were randomly selected to analyze stem length and biomass. And three pots from each treatment combination were randomly selected to analyze N contents of leachate, physiological indicators of poplar, AM colonization rate, and physicochemical properties of soil.
2.4.1. Leached Fluid Collection and N Contents Determination
Several days before the poplar seedlings were harvested, the leached fluid was collected and analysed. Based on the results of the preliminary experiment, the following method was adopted: the seedlings were treated with the N nutrient solution at 5:00 pm, and 200 mL of distilled water was added to each pot at 9:30 am the next day. A plate was placed under the pot to collect the leached fluid. After most of the distilled water was collected in the plate (after approximately 1.5 h), we filtered the leached fluid with filter paper, determined the filtrate volume, and analysed the nitrate and ammonium N concentration of the filtrate using a continuous-flow autoanalyzer (SA-40, Skalar Analytical B.V., Breda, The Netherlands). We calculated the total amounts of nitrate and ammonium N based on these values.
2.4.2. Seedling Growth and Physiological Indicators
The seedling height was measured using a tape, and stem biomass were determined by drying for 0.5 h at 105 °C and then at 65 °C to a constant weight. The transpiration rate of poplar leaves was measured using a portable photosynthesis analyser (Li-6400, LI-COR, Inc., Lincoln, NE, USA) between 8:30 am to 11:30 am under a light intensity of 1000 µmol·m−2·s−1 [46]. Seedling root morphology was scanned using a root scanner (STD1600 Epson, Long Beach, CA, USA) with a resolution of 300 dpi. The scanned images were analysed using the WinRHIZO root system analysis software (Regent Instrument Inc., Sainte-Foy, QC, Canada), and parameters of root length, root surface area, and mean root diameter were determined and analysed [47]. Root activity was the dehydrogenase activity in fine-roots, which was determined by the triphenyl tetrazolium chloride (TTC) reduction assay [48]. Oven-dried root samples (0.3 g) were ground to pass through a 0.25-mm sieve, and digested 2–3 times with 5 mL sulfuric acid, each time for 0.5 h at 370 °C; and the root N concentration was determined using a Kjeltec 8400 analyser (Foss, Höganäs, Sweden) [49].
2.4.3. AM Colonization Rate
The staining of poplar roots was conducted as described by Phillips and Hayman (1970) [50]. The root length colonization was determined according to the magnifying cross method [51] with a slight modification. Specifically, we randomly chose well-decoloured root segments to make glass slides. For each sample, ten slides were prepared and examined under a compound microscope (Olympus BX51, Olympus Corporation, Tokyo, Japan) with a crosshair (8 μm in line thickness) in one of the eyepieces at a magnification of 20 × 10. We randomly chose visual fields and examined 200 intersections for each slide to determine the arbuscular colonization rate and the total colonization rate.
2.4.4. Soil Physicochemical Properties
After harvest, the soil samples were air-dried in the shade and ground to pass through a 2-mm sieve for microaggregates, a 1-mm sieve for pH, a 0.25-mm sieve for CEC and total nitrogen (TN), and a 0.149-mm sieve for SOC determination. CEC was determined using the sodium acetate–flame photometry method [52]. pH was determined using a pH meter (Mettler-Toledo Instruments, Shanghai Co., Ltd., Shanghai, China) at a soil-to-water ratio of 1:2.5 [53]. SOC was determined using the K2Cr2O7-H2SO4 oxidation method [54]. The particle size distribution of the aggregates (including microaggregates and macroaggregates) was determined using a Malvern laser particle size analyser (Mastersizer 2000, Malvern Instruments, Worcestershire, UK) [55]. TN was determined using the semi-micro Kjeldahl method [49].
2.5. Data Processing
Statistical analyses were performed using the SPSS 17.0 software (SPSS Inc., Chicago, IL, USA), in which two-way analysis of variance (ANOVA) was used to determine the significance of the effects of N treatment, AM treatment and their interaction on experimental parameters. One-way ANOVA followed by Duncan’s multiple range test were used to compare the differences in means among different N levels, and independent sample T tests were used to compare the difference between +AM and −AM treatments. Redundancy analysis (RDA) was conducted using the CANOCO 5.0 to quantify the relative contribution of influencing factors to the changes of NO3− and NH4+ concentrations and their total amount. The figures were drawn using the SigmaPlot 12.5 software (Systat Software, San Jose, CA, USA).
3. Results
3.1. Effects of AM Fungi on Poplar Seedling Height and Stem Biomass Under Different N Levels
As the N application increased, the height and stem biomass of the poplar seedlings initially increased and then decreased (Figure 1). Height and stem biomass peaked at the 7.5 mM N level and were significantly different from the 1.0 mM and 20.0 mM N levels, indicating that the 7.5 mM N level was optimal for poplar tree growth and that exceeding this level leads to excessive fertilization. Accordingly, we chose the N levels of 1.0, 7.5 and 20.0 mM to respectively represent low, optimum and high N application levels in subsequent analyses, and to determine the effects of AM fungi on soil N leaching.
AM fungi inoculation had little effect on poplar seedling height or stem biomass at low N application levels but significantly increased poplar seedling height and stem biomass under optimum and high N levels (Figure 1).
3.2. Effects of AM Fungi on Soil N Leaching
The volumes of leached fluid were smallest at optimum N level, while the NO3− and NH4+ concentrations and the total amounts of NO3− and NH4+ in the leached fluid yielded significantly higher values at the high N level than at low and optimum N levels (Table 1). Regardless of AM fungi inoculation, the NO3− concentration of the leached fluid was higher than the NH4+ concentration, and the differences were highly significant for the optimum and high N levels.
Under optimum and high N levels, the NO3− and NH4+ concentrations and the total amounts of NO3− and NH4+ in the +AM treatment were lower than those in the −AM treatment, but the differences were significant only for the high N level (decreased by 20.0%, 67.5% and 22.5%, respectively) (Table 1). Although AM fungi inoculation had a nonsignificant effect on the leached fluid volume, it significantly reduced the NO3− concentration, NH4+ concentration and the total amounts of NO3− and NH4+ in the leachate (Table 1).
3.3. AM Colonization Rates under Different N Levels
AM colonization in poplar roots differed with varying N levels. As the N application level increased, the vesicle and spore sizes of the +AM fungi increased from the low N level, and then decreased in the high N level, peaking at the optimum N level (Figure 2a–c). While the arbuscular colonization rate and total colonization rate increased gradually and peaked at the high N level. At the optimum and high N levels, the arbuscular colonization rate and the total colonization rate were not significantly different, but were significantly higher than those at the low N level (Figure 2d).
3.4. Effects of AM Fungi on Physiological and Root Morphological Variables of Poplar Seedlings
The effects of AM fungi on the leaf transpiration rate, root morphology, root activity and root N content of the poplar seedlings varied with different N application levels (Figure 3). Regardless of AM inoculation treatment, the transpiration rate of poplar was highest at the optimum N level. Under the optimum and high N levels, the transpiration rates of the +AM treatment were 22.1% and 9.2% higher than those of the −AM treatment, respectively. The AM fungi inoculation and the N application level exerted a significant effect on the transpiration rate (Figure 3a).
The root surface area and length in the +AM treatment were lower than those of the −AM treatment (Figure 3b,c), while no significant difference in root surface area was observed between AM inoculation treatments, and the root length differences between the two treatments were significant only at the optimum and high N levels (27.4% and 31.6%, respectively). In contrast, the root diameter of the +AM treatment was higher than that in the −AM treatment, especially at the high N level, which was 28.4% higher (Figure 3d). The AM fungi inoculation and N level exerted significant influences on the root length, root diameter and surface area (Figure 3b–d).
AM fungi inoculation improved the root activity of the poplar seedlings, especially at the optimum N levels, under which the root activity increased by 34.6%, compared with the −AM treatment. The root N concentration also increased by AM inoculation, but this increase was only significant at the high N level (by 50.7%) (Table 1). Different N levels and AM inoculation had significant effect on the root activity and root N content.
3.5. Response of Soil Properties to AM Inoculation
3.5.1. Soil CEC and pH
The soil CEC of the +AM treatment was significantly lower than in the −AM treatment at the low N level (11.3%), while it was higher at the optimum and high N levels, especially for optimum N, under which CEC increased by 9.7% (Figure 4a). The effect of AM fungi inoculation on CEC was not statistically significant, whereas the effects of the N application level and the interaction between the two on CEC were statistically significant.
Soil pH at the optimum and high N levels was lower than that at the low N level in the +AM treatment, and both were 1.6% lower than that the −AM treatment (Figure 4b). Inoculation reduced soil pH, which reached significant levels at high N levels. The N application level and AM fungi inoculation had significant effects on soil pH.
3.5.2. SOC and Soil Aggregates
Regardless of AM inoculation treatment, the highest SOC was observed at the optimum N level (Figure 4c). At the optimum and high N levels, soil SOC contents of the −AM treatment were 6.44, 7.30, and 6.50 g kg−1, respectively, and the corresponding values for the +AM treatment were 6.31, 8.25, and 7.15 g kg−1, which were 13.0% and 10.0% larger than −AM treatment under optimum and high N levels. And the difference was significant for the optimum and high N level. The N application and AM fungi inoculation significantly affected the soil SOC.
Table 2 shows the compositions of soil microaggregates (<0.25 mm) and macroaggregates (≥0.25 mm) of the +AM and −AM treatments under different N levels. The proportion of the 0.01–0.25 mm microaggregates, which reflect soil fertility characteristics, was highest in all the soil aggregate size fractions, varying in a range of 66.9–70.4%. Under the optimum and high N levels, compared to the −AM treatment, the 0.01–0.25 mm micro-aggregate content of the +AM treatment was slightly lower, and the proportions of the 0.25–0.5 mm and the 0.5–1 mm macroaggregates were significantly higher. The 0.25–0.5 mm and the 0.5–1 mm macroaggregates content of +AM treatment were 1.5 times and 1.4 times of the −AM treatment at the optimum N level, and 1.6 times and 2.2 times of the −AM treatment at the high N level, respectively (Table 2). AM fungi inoculation significantly affected the 0.25–0.5 mm and the 0.5–1 mm macroaggregates.
3.5.3. Soil TN
Under different N application levels, the AM fungi inoculation had different effects on soil TN content (Figure 4d). Soil TN in the −AM treatment was slightly higher than that of the +AM treatment under the low N level but was lower under the optimum and high N levels. And under the optimum and high N levels, soil TN values in the +AM treatment were 7.2% and 4.7% higher than those in the −AM treatment, which indicated that AM fungi inoculation can increase the soil TN concentration at optimal and high N levels (Figure 4d).
3.6. The Relative Contributions of the Variables Affecting Soil N Leaching
In general, the relative contribution of the variables on NH4+, NO3− concentrations and the total amounts of NO3− and NH4+ in the leaching solution tended to be uniform as the N levels increased (Figure 5). Their relative contributions to the colonization rate increased, while those to plant physiological and soil variables decreased.
Among all of the variables, the root N concentration had the highest relative contribution rate to NO3− concentration at the optimal N level at 21.8%, and it also had high relative contribution rates at low and high N levels, 16.4% and 10.5%, respectively (Figure 5). The SOC had the highest relative contribution rate to NH4+ concentration at the optimal N level at 22.3%, and the relative contribution rates were 26.3% and 6.7%, respectively, at low and high N levels. The main variables affecting the leaching solution volume also changed with the increase of N level; these were SOC, root N concentration and root activity, respectively, at low, optimal, and high N levels. Meanwhile, SOC was also the main variable affecting the total N content in the leaching solution at low and high N levels, with relative contribution rates of 28.9% and 12.9%, respectively; and this relative contribution rate was 14.9% at the optimal N level, which was only lower than the transpiration rate (15.2%) (Figure 5). In summary, the root N concentration was the main variables related to the NO3− concentration in the leachate, and the increased SOC was linked to lower NH4+ concentration and the total N amount of both forms in the leachate.
4. Discussion
The results of our study showed that N application significantly increased N loss via leaching, and AM inoculation differed in its effectiveness at different N application levels. AM fungi could reduce soil NO3− and NH4+ concentrations and their total leaching amounts at optimum and high N levels, and the reductions were significant for the high N level, thereby lowering the risk of environmental pollution. Compared with NO3−, AM fungi reduced NH4+ leaching more significantly. Meanwhile, under optimum and high N levels, AM inoculation increased the colonization rate of poplar roots, and improved plant physiological conditions and soil physical and chemical characteristics to varying degrees. The results also indicated that SOC content might closely related to NH4+ and the total amounts of NO3− and NH4+, while root N concentration might be correlated with NO3− in the leachate.
4.1. The Influence of Am Fungi on Soil N Leaching
The present study demonstrated that under low, optimum and high N application levels, AM inoculation had no, slight, and significant effects on the reduction of N leaching, respectively, indicating that AM fungi were more effective in reducing N leaching at higher N levels. C and N are two of the most important resources for morphogenesis of the AM fungi. Under low N application, AM fungi cannot obtain enough N from the soil to synthesize their spore wall component (chitin) [56]. However, plant photosynthate production is strongly limited by N availability [57]; therefore, the C flow from the host plant to AM fungi may be reduced under N limited conditions. Thus, under low N levels, the proliferation of AM fungi can be inhibited due to the lack of C and N, which may limit their corresponding ecological functions. AM fungi significantly reduced soil NO3− and NH4+ concentrations and their total leaching amounts at the high N level (Table 1). These results are consistent with the studies by Asghari and Cavagnaro (2011) [23], where inoculation of AM fungi reduced the leaching of both NO3− and NH4+. Thus, AM fungi can reduce N leaching, and their intercepting effects are related to the amount of nitrogen applied, and they are more profound in nutrient leaching caused by excessive fertilization.
We found that AM fungi inoculation significantly decreased soil NO3− and NH4+ leaching by 20.0% and 67.5% at the high N level, respectively, indicating that AM fungi inoculation is more effective in reducing NH4+ leaching. This result is consistent with the results of van der Heijden (2010) [19]. Here, we summarize the reasons as follows: First, AM fungi prefer NH4+ and can thus absorb more NH4+ [58]. Johansen et al. (1996) demonstrated that AM fungi are capable of absorbing exogenous NH4+ [59], and Toussaint et al. (2004) used 15 N-labelled NH4+ and NO3− to show that in the presence of both NH4+ and NO3−, the AM fungi prefer to utilize the former [60]. Second, under the optimum and high N levels, CEC and SOC increased after AM fungi inoculation, enabling the soil to absorb more NH4+. The negative charge density of the soil SOM contributes significantly to the soil CEC [61]. Gai et al. (2014) showed that biocarbon soil made from corn stalks had the highest CEC and the highest ability to adsorb NH4+, thereby playing an important role in reducing NH4+ pollution [62]. Third, although poplar trees can utilize more NO3− relative to NH4+ [63], we found that AM fungi inoculation at optimum and high N levels reduced root length and root surface area; these effects are not conducive to absorbing NO3− in a larger range and in turn can lead to a relatively higher level of NO3− leaching.
4.2. Mechanism of AM Fungi on N Leaching in Poplar Microsystem
In this study, the inoculated exogenous AM fungi caused higher arbuscular colonization and total colonization rates with high concentration of applied N. Previous studies have shown that the AM colonization rate is affected by the amount of N applied [57], but this effect varies. Parniske (2008) reported that plant and AM fungal genes facilitated individual colonization steps [64], indicating that the effects of N application level on root colonization rate may be plant and/or fungal species dependent. For example, Zhen et al. (2014) found that the root colonization rate of Leymus chinensis was slightly increased, while that of Artemisia frigida was decreased by N addition [65]. In this study, we showed that N addition increased the mycorrhizal root infection rate of poplars inoculated with R. irregularis. The results of this study are consistent with those of Wu et al. (2017), indicating that N application can provide a nitrogen source for growth and chitin formation in AM fungi [42], thereby increasing the colonization rate. Govindarajulu et al. detected high N uptake in cultured mycorrhizae, but they did not observe N transfer to the plant [58]. Leigh et al. monitored the process through which N transfers from AM fungi to the whole plant using the grid compartment system and 15N labelling; they found that more than one-third of the N was absorbed by AM fungi and then transported to the plant, thereby improving the plant’s N uptake [16]. Therefore, the increase in AM infection rate under optimal N and high N conditions indicates that, more nitrogen is fixed for its own growth and more nitrogen also is transferred to the plant.
Root morphology is important for plant N uptake. Here, AM inoculation decreased root length and root surface area, and this could decrease N uptake. However, the root active absorbing area of poplar seedlings was increased after AM inoculation [46], and this could partially offset the negative effects of the smaller sizes of roots. Increased root activity and plant transpiration can promote plant N uptake, thus reducing the amount of leached N [66]. In this study, under optimal and high N levels, we found that the height and stem biomass of poplar seedlings in the +AM treatment increased, while the leaf transpiration rate and root activity increased, ultimately leading to increased root N concentration (Figure 2a,e). Consistent with our results, Li et al. [67] and Bitterlich et al. [68] also reported that AM fungi significantly increased the transpiration rate in barley and tomato, respectively. Aronsson and Bergström found that N leaching in willow significantly decreased after the second year due to the increased transpiration rate caused by biomass accumulation, which increased the plant’s uptake of NO3− [69]. Previous studies showed that plant root activity can be increased by AM fungi [70], whereas it is a comprehensive indicator of root nutrient absorption. Guo et al. demonstrated that increased root activity can increase N uptake of a root system through temporarily fixing Nin organic form and in turn mitigating N leaching [71].
In addition to the increased N uptake by the AM fungi and the plant, the improved ability of the soil to intercept N also plays a key role in reducing N leaching. Various processes such as soil ion exchange and SOC accumulation can change soil structure and thus increase the interception of N. Soil CEC represents the total amount of cations that can be adsorbed by soil colloids. As CEC increases, the ability of the soil to adsorb NH4+ increases accordingly [62]. In this study, under optimal and high N levels, CEC and SOC increased after AM inoculation, which is consistent with the results of Wei and Shao [72] and Curtin et al. [73]. Three factors, SOC, plant roots and AM fungi, play important role in stabilizing the soil, and thereby decreasing the amount of N leaching. Daynes et al. reported that in the presence of adequate SOM, the plant root system is the key factor for soil structure development, and AM fungi can further stabilize soil [74]. Soil aggregates can not only influence microbial community structure but also improve nutrient retention [37]. AM fungi can affect the SOC by secreting various secondary metabolites such as glomalin [75], then facilitating the soil microaggregates to form macroaggregate particles and changing the soil aggregate structure through hyphae [76]. Therefore, AM fungi can increase the soil TN retention and reduce N leaching by changing the CEC, SOC and soil structure.
Analysis of the relative contribution of the variables showed that the root N concentration mainly affected the NO3− concentration in the leaching solution, while SOC was closely related to NH4+ concentration and the total N amount. To our knowledge, no relevant analysis has been performed under AM inoculation conditions. Bergeron et al. reported that tree roots decreased subsoil NO3− leaching by increasing root N uptake, and inferred the role of root microorganisms [77]. Kanthle et al. observed lower NO3− leaching at higher SOC content in a laboratory column experiment [78]. Abdalla et al. also demonstrated that cover plants reduced N leaching associated with increased SOC [79]. The above results indicate that the increase of root N concentration and soil SOC caused by inoculation with AM fungi is of great significance for reducing soil N leaching.
We conclude that AM fungi reduce soil N loss by leaching mainly through the following three aspects: first, increasing the AM fungal biomass to absorb nitrogen and to transfer a large amount of N to plants; second, improving plant physiological conditions to enhance N interception by plants; and third, improving the physical and chemical characteristics of soil to sequester more nitrogen.
5. Conclusions
We confirmed that AM fungi can reduce soil N leaching in a poplar microcosm at optimum and high N application levels. Moreover, AM fungi inoculation may be more effective in reducing NH4+-N leaching. Although the poplar root system became smaller (root length and surface area) and the soil microaggregate content did not change after inoculation, AM fungi might reduce the risk of N leaching by increasing the inoculation rate (arbuscular and total) of poplar roots, by improving plant physiological conditions (transpiration rate, root activity and root N concentration), and by altering soil physical and chemical characteristics (pH, CEC, SOC and TN).
At high N levels, the relative contributions of AM inoculation on N leaching increased, with larger contribution to NH4+ than NO3− leaching. The increased SOC was linked to lower NH4+ and total N in the leachate, while enhanced root N concentration was linked to lower NO3−. This could mean that the lower NH4+ and total N leaching of AM inoculation treatment were due to the improvement of soil properties, while the lower NO3− leaching is due to the improved plant physiological conditions by AM fungi, under optimum and high N application levels. The above results indicate that the mixed application of AM fungi and organic fertilizers to the soil may have important applications for alleviating soil nutrient leaching. More investigations on the basic processes are needed, to fully understand the mechanisms by which AM fungi influence nutrient cycling; such knowledge will guide us as to how to make use of the fungi’s potential to successfully promote the sustainable development of the plant-soil continuum.
Author Contributions
Conceptualization, F.F., C.W. and M.T.; Formal analysis, F.F.; Funding acquisition, C.W. and M.T.; Methodology, F.F. and C.W; Supervision, M.T.; Writing–Original Draft Preparation F.F.; Writing–Review and Editing, C.W., F.W., and M.T. Grammar revision, R.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (41671268), National Key Research and Development Program of China (2018YFD0600203, 2017YFD0600604), and the Youth Science Foundation of Jiangxi Provincial Department of Education, China (GJJ180224).
Acknowledgments
We acknowledge the Light of West China Program of Chinese Academy of Sciences (XAB2018B08), and the Youth Talent Plan Foundation of Northwest A&F University (2452018087).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Liu, J.; You, L.; Amini, M.; Obersteiner, M.; Herrero, M.; Zehnder, A.J.B.; Yang, H. A high-resolution assessment on global nitrogen flows in cropland. Proc. Natl. Acad. Sci. USA 2010, 107, 8035–8040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carpenter, S.R.; Caraco, N.F.; Correll, D.L.; Howarth, R.W.; Sharpley, A.N.; Smith, V.H. Nonpoint pollution of surface waters with phosphorus and nitrogen. Ecol. Appl. 1998, 8, 559–568. [Google Scholar] [CrossRef]
- Matson, P.A.; Parton, W.J.; Power, A.G.; Swift, M.J. Agricultural intensification and ecosystem properties. Science 1997, 277, 504–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohm, C.; Landgraf, D.; Makeschin, F. Effects of two contrasting agricultural land-use practices on nitrogen leaching in a sandy soil of Middle Germany. J. Plant Nutr. Soil Sci. 2009, 172, 408–417. [Google Scholar] [CrossRef]
- Farneselli, M.; Tosti, G.; Onofri, A.; Benincasa, P.; Guiducci, M.; Pannacci, E.; Tei, F. Effects of N sources and management strategies on crop growth, yield and potential N leaching in processing tomato. Eur. J. Agron. 2018, 98, 46–54. [Google Scholar] [CrossRef]
- Peng, S.Z.; He, Y.P.; Yang, S.H.; Xu, J.Z. Effect of controlled irrigation and drainage on nitrogen leaching losses from paddy fields. Paddy Water Environ. 2015, 13, 303–312. [Google Scholar] [CrossRef]
- Shaddox, T.W.; Kruse, J.K.; Miller, G.L.; Nkedi-Kizza, P.; Sartain, J.B. Surfactant-modified soil amendments reduce nitrogen and phosphorus leaching in a sand-based rootzone. J. Environ. Qual. 2016, 45, 1549–1556. [Google Scholar] [CrossRef]
- Yu, Q.G.; Chen, Y.X.; Ye, X.Z.; Zhang, Q.L.; Zhang, Z.J.; Tian, P. Evaluation of nitrification inhibitor 3,4-dimethyl pyrazole phosphate on nitrogen leaching in undisturbed soil columns. Chemosphere 2007, 67, 872–878. [Google Scholar] [CrossRef]
- Cavagnaro, T.R.; Bender, S.F.; Asghari, H.R.; van der Heijden, M.G.A. The role of arbuscular mycorrhizas in reducing soil nutrient loss. Trends Plant Sci. 2015, 20, 283–290. [Google Scholar] [CrossRef] [Green Version]
- Guether, M.; Balestrini, R.; Hannah, M.; He, J.; Udvardi, M.K.; Bonfante, P. Genome-wide reprogramming of regulatory networks, transport, cell wall and membrane biogenesis during arbuscular mycorrhizal symbiosis in Lotus japonicus. New Phytol. 2009, 182, 200–212. [Google Scholar] [CrossRef]
- Smith, S.E.; Smith, F.A. Roles of arbuscular mycorrhizas in plant nutrition and growth: New paradigms from cellular to ecosystem scales. Annu. Rev. Plant Biol. 2011, 62, 227–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis; Academic Press: New York, NY, USA, 2008. [Google Scholar]
- Bucking, H.; Kafle, A. Role of arbuscular mycorrhizal fungi in the nitrogen uptake of plants: Current knowledge and research gaps. Agronomy (Basel) 2015, 5, 587–612. [Google Scholar] [CrossRef] [Green Version]
- Leifheit, E.F.; Veresoglou, S.D.; Lehmann, A.; Morris, E.K.; Rillig, M.C. Multiple factors influence the role of arbuscular mycorrhizal fungi in soil aggregation-a meta-analysis. Plant Soil 2014, 374, 523–537. [Google Scholar] [CrossRef]
- Li, X.L.; George, E.; Marschner, H. Extension of the phosphorus depletion zone in VA-mycorrhizal white clover in a calcareous soil. Plant Soil 1991, 136, 41–48. [Google Scholar] [CrossRef]
- Leigh, J.; Hodge, A.; Fitter, A.H. Arbuscular mycorrhizal fungi can transfer substantial amounts of nitrogen to their host plant from organic material. New Phytol. 2009, 181, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Hooker, J.E.; Piatti, P.; Cheshire, M.V.; Watson, C.A. Polysaccharides and monosaccharides in the hyphosphere of the arbuscular mycorrhizal fungi Glomus E3 and Glomus tenue. Soil Biol. Biochem. 2007, 39, 680–683. [Google Scholar] [CrossRef]
- Veresoglou, S.D.; Chen, B.D.; Rillig, M.C. Arbuscular mycorrhiza and soil nitrogen cycling. Soil Biol. Biochem. 2012, 46, 53–62. [Google Scholar] [CrossRef]
- Van der Heijden, M.G.A. Mycorrhizal fungi reduce nutrient loss from model grassland ecosystems. Ecology 2010, 91, 1163–1171. [Google Scholar] [CrossRef]
- Asghari, H.R.; Cavagnaro, T.R. Arbuscular mycorrhizas reduce nitrogen loss via leaching. PLoS ONE 2012, 7, e29825. [Google Scholar] [CrossRef] [Green Version]
- Bowles, T.M.; Jackson, L.E.; Cavagnaro, T.R. Mycorrhizal fungi enhance plant nutrient acquisition and modulate nitrogen loss with variable water regimes. Glob. Chang. Biol. 2018, 24, E171–E182. [Google Scholar] [CrossRef]
- Kucova, L.; Zahora, J.; Pokluda, R. Effect of mycorrhizal inoculation of leek Allium porrum L. on mineral nitrogen leaching. Hortic. Sci. 2016, 43, 195–202. [Google Scholar] [CrossRef] [Green Version]
- Asghari, H.R.; Cavagnaro, T.R. Arbuscular mycorrhizas enhance plant interception of leached nutrients. Funct. Plant Biol. 2011, 38, 219–226. [Google Scholar] [CrossRef]
- Martínez-García, L.B.; De Deyn, G.B.; Pugnaire, F.I.; Kothamasi, D.; van der Heijden, M.G.A. Symbiotic soil fungi enhance ecosystem resilience to climate change. Glob. Chang. Biol. 2017, 23, 5228–5236. [Google Scholar] [CrossRef] [PubMed]
- Bender, S.F.; Conen, F.; van der Heijden, M.G.A. Mycorrhizal effects on nutrient cycling, nutrient leaching and N2O production in experimental grassland. Soil Biol. Biochem. 2015, 80, 283–292. [Google Scholar] [CrossRef]
- Bender, S.F.; van der Heijden, M.G.A. Soil biota enhance agricultural sustainability by improving crop yield, nutrient uptake and reducing nitrogen leaching losses. J. Appl. Ecol. 2015, 52, 228–239. [Google Scholar] [CrossRef] [Green Version]
- Kohl, L.; van der Heijden, M.G.A. Arbuscular mycorrhizal fungal species differ in their effect on nutrient leaching. Soil Biol. Biochem. 2016, 94, 191–199. [Google Scholar] [CrossRef] [Green Version]
- Kohl, L.; Oehl, F.; van der Heijden, M.G.A. Agricultural practices indirectly influence plant productivity and ecosystem services through effects on soil biota. Ecol. Appl. 2014, 24, 1842–1853. [Google Scholar] [CrossRef]
- Corkidi, L.; Merhaut, D.J.; Allen, E.B.; Downer, J.; Bohn, J.; Evans, M. Effects of mycorrhizal colonization on nitrogen and phosphorus leaching from nursery containers. HortScience 2011, 46, 1472–1479. [Google Scholar] [CrossRef] [Green Version]
- De Vries, F.T.; Thebault, E.; Liiri, M.; Birkhofer, K.; Tsiafouli, M.A.; Bjornlund, L.; Jorgensen, H.B.; Brady, M.V.; Christensen, S.; de Ruiter, P.C.; et al. Soil food web properties explain ecosystem services across European land use systems. Proc. Natl. Acad. Sci. USA 2013, 110, 14296–14301. [Google Scholar] [CrossRef] [Green Version]
- Hodge, A.; Fitter, A.H. Substantial nitrogen acquisition by arbuscular mycorrhizal fungi from organic material has implications for N cycling. Proc. Natl. Acad. Sci. USA 2010, 107, 13754–13759. [Google Scholar] [CrossRef] [Green Version]
- Cameron, K.C.; Di, H.J.; Moir, J.L. Nitrogen losses from the soil/plant system: A review. Ann. Appl. Biol. 2013, 162, 145–173. [Google Scholar] [CrossRef]
- Francis, G.S.; Haynes, R.J.; Sparling, G.P.; Ross, D.J.; Williams, P.H. Nitrogen mineralization, nitrate leaching and crop growth following cultivation of a temporary leguminous pasture in autumn and winter. Fertil. Res. 1992, 33, 59–70. [Google Scholar] [CrossRef]
- Hodge, A.; Storer, K. Arbuscular mycorrhiza and nitrogen: Implications for individual plants through to ecosystems. Plant Soil 2015, 386, 1–19. [Google Scholar] [CrossRef]
- Duffkova, R.; Fucik, P.; Jurkovska, L.; Janouskova, M. Experimental evaluation of the potential of arbuscular mycorrhiza to modify nutrient leaching in three arable soils located on one slope. Appl. Soil Ecol. 2019, 143, 116–125. [Google Scholar] [CrossRef]
- Rabbi, S.M.F.; Wilson, B.R.; Lockwood, P.V.; Daniel, H.; Young, I.M. Soil organic carbon mineralization rates in aggregates under contrasting land uses. Geoderma 2014, 216, 10–18. [Google Scholar] [CrossRef]
- Six, J.; Bossuyt, H.; Degryze, S.; Denef, K. A history of research on the link between (micro)aggregates, soil biota, and soil organic matter dynamics. Soil Till. Res. 2004, 79, 7–31. [Google Scholar] [CrossRef]
- Benucci, G.M.N.; Lefevre, C.; Bonito, G. Characterizing root-associated fungal communities and soils of Douglas-fir (Pseudotsuga menziesii) stands that naturally produce Oregon white truffles (Tuber oregonense and Tuber gibbosum). Mycorrhiza 2016, 26, 367–376. [Google Scholar] [CrossRef]
- Manzone, M.; Bergante, S.; Facciotto, G. Energy and economic evaluation of a poplar plantation for woodchips production in Italy. Biomass Bioenerg. 2014, 60, 164–170. [Google Scholar] [CrossRef] [Green Version]
- Baum, C.; Makeschin, F. Effects of nitrogen and phosphorus fertilization on mycorrhizal formation of two poplar clones (Populus trichocarpa and P. tremula × tremuloides). J. Plant Nutr. Soil Sci. 2000, 163, 491–497. [Google Scholar] [CrossRef]
- Rooney, D.C.; Prosser, J.I.; Bending, G.D.; Baggs, E.M.; Killham, K.; Hodge, A. Effect of arbuscular mycorrhizal colonisation on the growth and phosphorus nutrition of Populus euramericana c.v. Ghoy. Biomass Bioenerg. 2011, 35, 4605–4612. [Google Scholar] [CrossRef]
- Wu, F.; Zhang, H.Q.; Fang, F.R.; Wu, N.; Zhang, Y.X.; Tang, M. Effects of nitrogen and exogenous rhizophagus irregularis on the nutrient status, photosynthesis and leaf anatomy of Populus × canadensis ‘Neva’. J. Plant Growth Regul. 2017, 36, 824–835. [Google Scholar] [CrossRef]
- He, X.; Li, Y.; Zhao, L. Dynamics of arbuscular mycorrhizal fungi and glomalin in the rhizosphere of Artemisia ordosica Krasch. In Mu Us sandland, China. Soil Biol. Biochem. 2010, 42, 1313–1319. [Google Scholar] [CrossRef]
- Feng, C.; Ma, Y.; Fu, S.; Chen, H.Y.H. Soil carbon and nutrient dynamics following cessation of anthropogenic disturbances in degraded subtropical forests. Land Degrad. Dev. 2017, 28, 2457–2467. [Google Scholar] [CrossRef]
- Knoth, J.L.; Kim, S.H.; Ettl, G.J.; Doty, S.L. Effects of cross host species inoculation of nitrogen-fixing endophytes on growth and leaf physiology of maize. GCB Bioenergy 2013, 5, 408–418. [Google Scholar] [CrossRef]
- Liu, T.; Sheng, M.; Wang, C.Y.; Chen, H.; Li, Z.; Tang, M. Impact of arbuscular mycorrhizal fungi on the growth, water status, and photosynthesis of hybrid poplar under drought stress and recovery. Photosynthetica 2015, 53, 250–258. [Google Scholar] [CrossRef]
- Chu, H.L.; Wang, C.Y.; Li, Z.M.; Wang, H.H.; Xiao, Y.G.; Chen, J.; Tang, M. The dark septate endophytes and ectomycorrhizal fungi effect on Pinus tabulaeformis carr. seedling growth and their potential effects to pine wilt disease resistance. Forests 2019, 10, 16. [Google Scholar] [CrossRef] [Green Version]
- Clemenssonlindell, A. Triphenyltetrazolium chloride as an indicator of fine-root vitality and environmental-stress in coniferous forest stands—Applications and limitations. Plant Soil 1994, 159, 297–300. [Google Scholar] [CrossRef]
- Schuman, G.E.; Stanley, M.A.; Knudsen, D. Automated total nitrogen analysis of soil and plant samples. Soil Sci. Soc. Am. J. 1973, 37, 480–481. [Google Scholar] [CrossRef]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- McGonigle, T.; Miller, M.; Evans, D.; Fairchild, G.; Swan, J. A new method which gives an objective measure of colonization of roots by vesicular—Arbuscular mycorrhizal fungi. New Phytol. 1990, 115, 495–501. [Google Scholar] [CrossRef]
- Qiu, X.C.; Zhu, Y.Q. Rapid analysis of cation-exchange properties in acidic soils. Soil Sci. 1993, 155, 301–308. [Google Scholar] [CrossRef]
- Wei, S.H.; Zhou, Q.X.; Koval, P.V. Flowering stage characteristics of cadmium hyperaccumulator Solanum nigrum L. and their significance to phytoremediation. Sci. Total Environ. 2006, 369, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.; Sommers, L. Total carbon, organic carbon, and organic matter. In Methods of Soil Analysis. Part 2. Chemical and Microbiological Properties; American Society of Agronomy: Madison, WI, USA, 1982; pp. 539–579. [Google Scholar]
- Bieganowski, A.; Ryzak, M.; Witkowska-Walczak, B. Determination of soil aggregate disintegration dynamics using laser diffraction. Clay Min. 2010, 45, 23–34. [Google Scholar] [CrossRef]
- Bago, B.; Cano, C.; Azcon-Aguilar, C.; Samson, J.; Coughlan, A.P.; Piche, Y. Differential morphogenesis of the extraradical mycelium of an arbuscular mycorrhizal fungus grown monoxenically on spatially heterogeneous culture media. Mycologia 2004, 96, 452–462. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, H.L.; Hartley, A.E.; Vogelsang, K.M.; Bever, J.D.; Schultz, P.A. Arbuscular mycorrhizal fungi do not enhance nitrogen acquisition and growth of old-field perennials under low nitrogen supply in glasshouse culture. New Phytol. 2005, 167, 869–880. [Google Scholar] [CrossRef] [PubMed]
- Govindarajulu, M.; Pfeffer, P.E.; Jin, H.; Abubaker, J.; Douds, D.D.; Allen, J.W.; Bücking, H.; Lammers, P.J.; Shachar-Hill, Y. Nitrogen transfer in the arbuscular mycorrhizal symbiosis. Nature 2005, 435, 819. [Google Scholar] [CrossRef]
- Johansen, A.; Finlay, R.D.; Olsson, P.A. Nitrogen metabolism of external hyphae of the arbuscular mycorrhizal fungus Glornus intraradices. New Phytol. 1996, 133, 705–712. [Google Scholar] [CrossRef]
- Toussaint, J.P.; St-Arnaud, M.; Charest, C. Nitrogen transfer and assimilation between the arbuscular mycorrhizal fungus Glomus intraradices Schenck & Smith and Ri T-DNA roots of Daucus carota L. in an in vitro compartmented system. Can. J. Microbiol. 2004, 50, 251–260. [Google Scholar]
- Curtin, D.; Rostad, H.P.W. Cation exchange and buffer potential of Saskatchewan soils estimated from texture, organic matter and pH. Can. J. Soil Sci. 1997, 77, 621–626. [Google Scholar] [CrossRef]
- Gai, X.P.; Wang, H.Y.; Liu, J.; Zhai, L.M.; Liu, S.; Ren, T.Z.; Liu, H.B. Effects of feedstock and pyrolysis temperature on biochar adsorption of ammonium and nitrate. PLoS ONE 2014, 9, e113888. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.X.; Meng, S.; Li, Y.M.; Su, L.; Zhao, Z. Nitrogen uptake and allocation in Populus simonii in different seasons supplied with isotopically labeled ammonium or nitrate. Trees Struct. Funct. 2016, 30, 2011–2018. [Google Scholar] [CrossRef]
- Parniske, M. Arbuscular mycorrhiza: The mother of plant root endosymbioses. Nat. Rev. Microbiol. 2008, 6, 763–775. [Google Scholar] [CrossRef] [PubMed]
- Zhen, L.N.; Yang, G.W.; Yang, H.J.; Chen, Y.L.; Liu, N.; Zhang, Y.J. Arbuscular mycorrhizal fungi affect seedling recruitment: A potential mechanism by which N deposition favors the dominance of grasses over forbs. Plant Soil 2014, 375, 127–136. [Google Scholar] [CrossRef]
- Vogeler, I.; Green, S.R.; Mills, T.; Clothier, B.E. Modelling nitrate and bromide leaching from sewage sludge. Soil Till. Res. 2006, 89, 177–184. [Google Scholar] [CrossRef]
- Li, T.; Lin, G.; Zhang, X.; Chen, Y.L.; Zhang, S.B.; Chen, B.D. Relative importance of an arbuscular mycorrhizal fungus (Rhizophagus intraradices) and root hairs in plant drought tolerance. Mycorrhiza 2014, 24, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Bitterlich, M.; Sandmann, M.; Graefe, J. Arbuscular mycorrhiza alleviates restrictions to substrate water flow and delays transpiration limitation to stronger drought in tomato. Front. Plant Sci. 2018, 9, 15. [Google Scholar] [CrossRef] [Green Version]
- Aronsson, P.G.; Bergström, L.F. Nitrate leaching from lysimeter-grown short-rotation willow coppice in relation to N-application, irrigation and soil type. Biomass Bioenerg. 2001, 21, 155–164. [Google Scholar] [CrossRef]
- Sheng, M.; Tang, M.; Chen, H.; Yang, B.W.; Zhang, F.F.; Huang, Y.H. Influence of arbuscular mycorrhizae on the root system of maize plants under salt stress. Can. J. Microbiol. 2009, 55, 879–886. [Google Scholar] [CrossRef]
- Guo, R.Y.; Qin, W.; Jiang, C.G.; Kang, L.Y.; Nendel, C.; Chen, Q. Sweet corn significantly increases nitrogen retention and reduces nitrogen leaching as summer catch crop in protected vegetable production systems. Soil Till. Res. 2018, 180, 148–153. [Google Scholar] [CrossRef]
- Wei, X.; Shao, M. Distribution characteristics of soil pH, CEC and organic matter in a small watershed of the Loess Plateau. Yingyong Shengtai Xuebao 2009, 20, 2710–2715, (Chinese with English abstract). [Google Scholar]
- Curtin, D.; Fraser, P.M.; Beare, M.H. Loss of soil organic matter following cultivation of long-term pasture: Effects on major exchangeable cations and cation exchange capacity. Soil Res. 2015, 53, 377–385. [Google Scholar] [CrossRef]
- Daynes, C.N.; Field, D.J.; Saleeba, J.A.; Cole, M.A.; McGee, P.A. Development and stabilisation of soil structure via interactions between organic matter, arbuscular mycorrhizal fungi and plant roots. Soil Biol. Biochem. 2013, 57, 683–694. [Google Scholar] [CrossRef]
- Sarapatka, B.; Alvarado-Solano, D.P.; Cizmar, D. Can glomalin content be used as an indicator for erosion damage to soil and related changes in organic matter characteristics and nutrients? Catena 2019, 181, 8. [Google Scholar] [CrossRef]
- Sharifi, Z.; Azadi, N.; Rahimi, S.; Certini, G. The response of glomalin-related soil proteins to fire or tillage. Geoderma 2018, 329, 65–72. [Google Scholar] [CrossRef]
- Bergeron, M.; Lacombe, S.; Bradley, R.L.; Whalen, J.; Cogliastro, A.; Jutras, M.F.; Arp, P. Reduced soil nutrient leaching following the establishment of tree-based intercropping systems in eastern Canada. Agrofor. Syst. 2011, 83, 321–330. [Google Scholar] [CrossRef]
- Kanthle, A.K.; Lenka, N.K.; Lenka, S.; Tedia, K. Biochar impact on nitrate leaching as influenced by native soil organic carbon in an Inceptisol of central India. Soil Tillage Res. 2016, 157, 65–72. [Google Scholar] [CrossRef]
- Abdalla, M.; Hastings, A.; Cheng, K.; Yue, Q.; Chadwick, D.; Espenberg, M.; Truu, J.; Rees, R.M.; Smith, P. A critical review of the impacts of cover crops on nitrogen leaching, net greenhouse gas balance and crop productivity. Glob. Chang. Biol. 2019, 25, 2530–2543. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Effects of AM fungi on stem length (a) and stem biomass(b) of poplar seedlings under different N application levels. * Significant effect at p < 0.05; ** Significant effect at p < 0.01; *** Significant effect at p < 0.001; NS: no significant effect. Values are presented as the means ± SE (n = 10). Different lowercase letters indicate significant difference among different N levels with the same AM treatment and different uppercase letters indicate significant difference between +AM and −AM treatments under the same N level at p < 0.05.
Figure 1.
Effects of AM fungi on stem length (a) and stem biomass(b) of poplar seedlings under different N application levels. * Significant effect at p < 0.05; ** Significant effect at p < 0.01; *** Significant effect at p < 0.001; NS: no significant effect. Values are presented as the means ± SE (n = 10). Different lowercase letters indicate significant difference among different N levels with the same AM treatment and different uppercase letters indicate significant difference between +AM and −AM treatments under the same N level at p < 0.05.
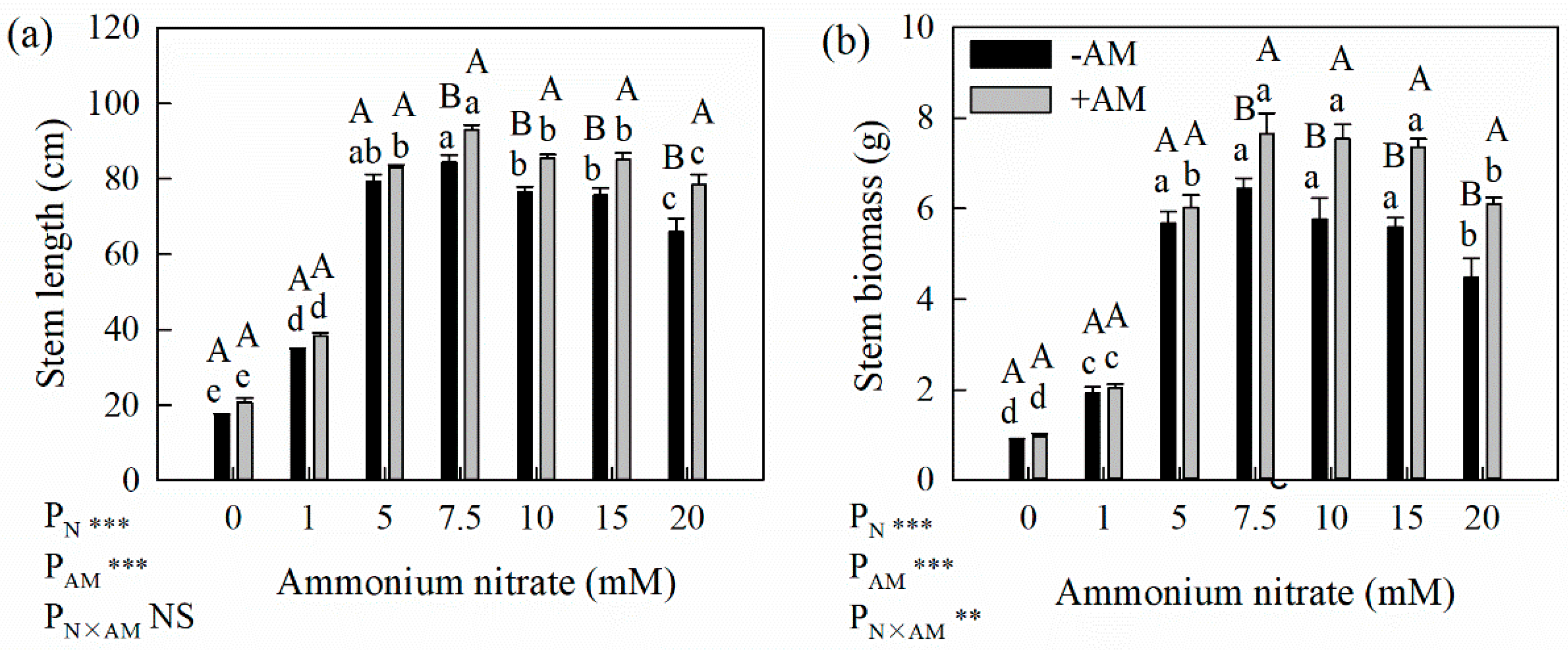
Figure 2.
AM colonization rate in poplar roots under (a) low, (b) optimum, and (c) high N application levels, and the root length colonization (d). Values are presented as the means ± SE (n = 3). Different lowercase letters indicate a significant difference at p < 0.05 according to Duncan’s test.
Figure 2.
AM colonization rate in poplar roots under (a) low, (b) optimum, and (c) high N application levels, and the root length colonization (d). Values are presented as the means ± SE (n = 3). Different lowercase letters indicate a significant difference at p < 0.05 according to Duncan’s test.

Figure 3.
Effects of AM fungi on physiological indicators and root properties of poplar seedlings under different N application levels (a. transpiration rate; b. root surface area; c. root length; d. average root diameter; e. root activity; f. root N concentration). * Significant effect at p < 0.05; ** Significant effect at p < 0.01; *** Significant effect at p < 0.001; NS: no significant effect. Values are presented as the means ± SE (n = 3). Different lowercase letters indicate significant difference among different N levels with the same AM treatment and different uppercase letters indicate significant difference between +AM and −AM treatments under the same N level at p < 0.05.
Figure 3.
Effects of AM fungi on physiological indicators and root properties of poplar seedlings under different N application levels (a. transpiration rate; b. root surface area; c. root length; d. average root diameter; e. root activity; f. root N concentration). * Significant effect at p < 0.05; ** Significant effect at p < 0.01; *** Significant effect at p < 0.001; NS: no significant effect. Values are presented as the means ± SE (n = 3). Different lowercase letters indicate significant difference among different N levels with the same AM treatment and different uppercase letters indicate significant difference between +AM and −AM treatments under the same N level at p < 0.05.
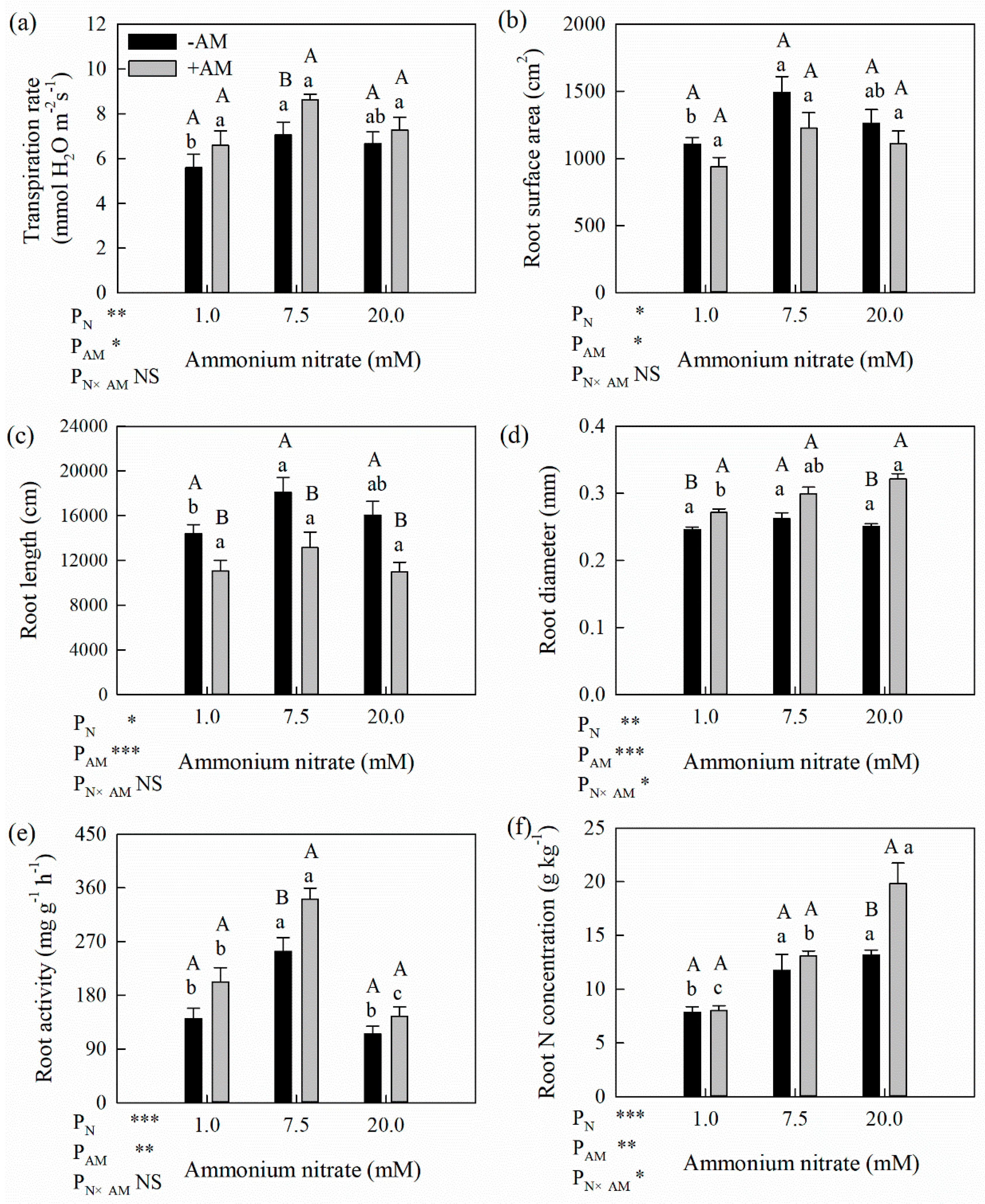
Figure 4.
Effects of AM fungi on soil properties at different N levels (a. cation exchange capacity (CEC); b. pH; c. soil organic carbon (SOC); d. total N (TN)). * Significant effect at p < 0.05; ** Significant effect at p < 0.01; *** Significant effect at p < 0.001; NS: no significant effect. Values are presented as the means ± SE (n = 3). Different lowercase letters indicate significant difference among different N levels with the same AM treatment and different uppercase letters indicate significant difference between +AM and −AM treatments under the same N level at p < 0.05.
Figure 4.
Effects of AM fungi on soil properties at different N levels (a. cation exchange capacity (CEC); b. pH; c. soil organic carbon (SOC); d. total N (TN)). * Significant effect at p < 0.05; ** Significant effect at p < 0.01; *** Significant effect at p < 0.001; NS: no significant effect. Values are presented as the means ± SE (n = 3). Different lowercase letters indicate significant difference among different N levels with the same AM treatment and different uppercase letters indicate significant difference between +AM and −AM treatments under the same N level at p < 0.05.
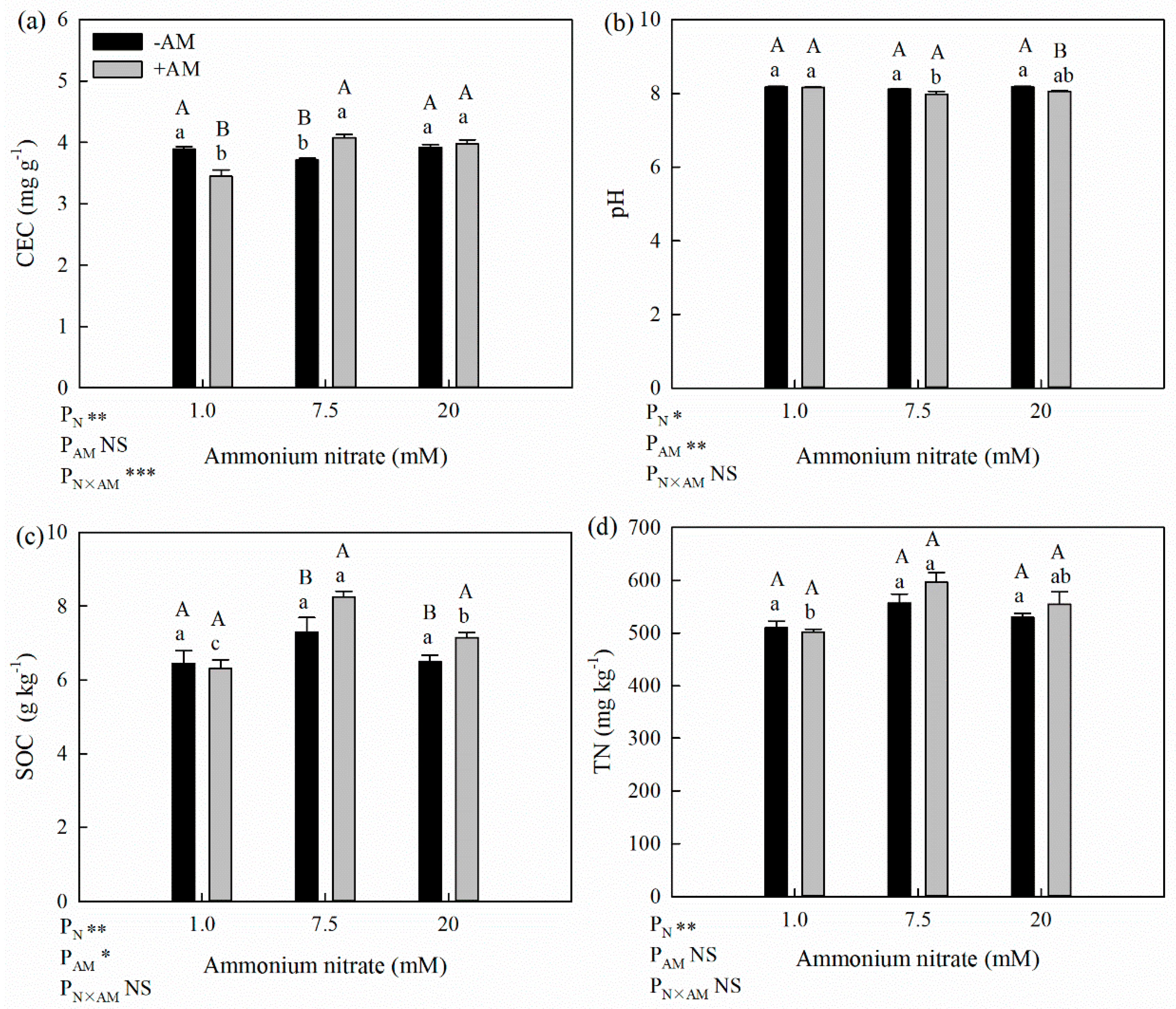
Figure 5.
Relative contributions to data variability explanation of NO3− and NH4+ concentrations, leaching volumes and the total amount of both forms at (a) low, (b) optimum and (c) high N level, by following variables: Ac, arbuscular colonization rate; Tc, total colonization rate; Tr, transpiration rate; Ra, root activity; Rl, root length; Rs, root area; Rd, root diameter; RN, root N concentration; CEC, cation exchange capacity; SOC, soil organic carbon; TN, soil total N concentration, A(0.5–1), 0.5–1 mm aggregates; A(0.25–0.5), 0.25–0.5 mm aggregates. n = 3.
Figure 5.
Relative contributions to data variability explanation of NO3− and NH4+ concentrations, leaching volumes and the total amount of both forms at (a) low, (b) optimum and (c) high N level, by following variables: Ac, arbuscular colonization rate; Tc, total colonization rate; Tr, transpiration rate; Ra, root activity; Rl, root length; Rs, root area; Rd, root diameter; RN, root N concentration; CEC, cation exchange capacity; SOC, soil organic carbon; TN, soil total N concentration, A(0.5–1), 0.5–1 mm aggregates; A(0.25–0.5), 0.25–0.5 mm aggregates. n = 3.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The N leaching in poplar microsystems.
| N Levels (mM) | Inoculation | Volume (mL) | Nitrate N (mg L−1) | Ammonium N (mg L−1) | Nitrate & Ammonium (mg) |
|---|---|---|---|---|---|
| 1 | −AM | 66.8 ± 5.1 | 0.75 ± 0.19 Ab | 0.35 ± 0.04 Ab | 0.07 ± 0.02 Ab |
| +AM | 67.0 ± 3.8 | 0.57 ± 0.13 Ab | 0.36 ± 0.01 Ab | 0.07 ± 0.01 Ab | |
| 7.5 | −AM | 58.8 ± 6.8 | 4.71 ± 0.47 Ab | 0.63 ± 0.22 Ab | 0.31 ± 0.05 Ab |
| +AM | 60.9 ± 6.9 | 3.75 ± 0.67 Ab | 0.45 ± 0.04 Ab | 0.26 ± 0.05 Ab | |
| 20 | −AM | 63.8 ± 2.1 | 31.71 ± 2.38 Aa | 4.56 ± 0.70 Aa | 2.31 ± 0.15 Aa |
| +AM | 68.5 ± 10.2 | 25.36 ± 2.57 Ba | 1.48 ± 0.23 Ba | 1.79 ± 0.12 Ba | |
| Significance | N | NS | *** | *** | *** |
| AM | NS | * | ** | * | |
| N × AM | NS | * | ** | * |
Notes: Data presented are the means ± SE (n = 3). Different lowercase letters indicate a significant difference among different N levels with the same AM treatment and different uppercase letters indicate significant difference between +AM and −AM under the same N level. * Significance level p < 0.05; ** Significance level p<0.01; *** Significance level p< 0.001; NS: No significant effect.
Table 2.
Composition of rhizosphere soil aggregates under different N application levels.
| N Levels (mM) | Inoculation | Aggregates Distribution (%) | ||||
|---|---|---|---|---|---|---|
| <0.002 mm | 0.002–0.01 mm | 0.01–0.25 mm | 0.25–0.5 mm | 0.5–1 mm | ||
| 1 | −AM | 13.37 ± 1.04 | 12.31 ± 0.98 | 70.42 ± 2.09 | 3.12 ± 0.14 Aa | 0.79 ± 0.01 Aa |
| +AM | 14.49 ± 0.13 | 13.24 ± 0.33 | 68.61 ± 0.37 | 3.03 ± 0.06 Ab | 0.63 ± 0.03 Ac | |
| 7.5 | −AM | 14.48 ± 0.40 | 13.23 ± 0.65 | 68.57 ± 0.89 | 2.88 ± 0.02 Bab | 0.83 ± 0.01 Ba |
| +AM | 14.18 ± 0.51 | 13.04 ± 1.09 | 67.29 ± 1.60 | 4.32 ± 0.02 Aa | 1.17 ± 0.01 Ab | |
| 20 | −AM | 14.05 ± 0.10 | 12.86 ± 1.40 | 69.70 ± 1.46 | 2.74 ± 0.09 Bb | 0.65 ± 0.02 Bb |
| +AM | 14.49 ± 1.20 | 12.73 ± 0.44 | 66.92 ± 0.63 | 4.45 ± 0.25 Aa | 1.41 ± 0.01 Aa | |
| Significance | N | NS | NS | NS | ** | *** |
| AM | NS | NS | NS | *** | *** | |
| N × AM | NS | NS | NS | *** | *** | |
Notes: Data presented are the means ± SE (n = 3). Different lowercase letters represent significant difference among different N levels with the same AM treatment and different uppercase letters indicate significant difference between +AM and −AM treatments under the same N level. ** Significance level p < 0.01; *** Significance level p < 0.001; NS: No significant effect.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Fang, F.; Wang, C.; Wu, F.; Tang, M.; Doughty, R. Arbuscular Mycorrhizal Fungi Mitigate Nitrogen Leaching under Poplar Seedlings. Forests 2020, 11, 325. https://doi.org/10.3390/f11030325
AMA Style
Fang F, Wang C, Wu F, Tang M, Doughty R. Arbuscular Mycorrhizal Fungi Mitigate Nitrogen Leaching under Poplar Seedlings. Forests. 2020; 11(3):325. https://doi.org/10.3390/f11030325
Chicago/Turabian StyleFang, Fengru, Chunyan Wang, Fei Wu, Ming Tang, and Russell Doughty. 2020. "Arbuscular Mycorrhizal Fungi Mitigate Nitrogen Leaching under Poplar Seedlings" Forests 11, no. 3: 325. https://doi.org/10.3390/f11030325
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.