Batch Growth of Chlorella Vulgaris CCALA 896 versus Semi-Continuous Regimen for Enhancing Oil-Rich Biomass Productivity

Abstract
:1. Introduction
2. Materials and Methods
2.1. Organism and Culture Conditions
2.2. Photobioreactor Shapes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Macroelements | Culture broth compositions | ||
|---|---|---|---|
| Modified Kolkwitz medium (g/L) | Modified BG-11 medium (g/L) | Modified basal medium (g/L) | |
| NaNO3 | 0.5 | ||
| KNO3 | 0.59 | 0.59 | |
| K2HPO4 | 0.14 | 0.04 | 0.038 |
| MgSO4 · 7H2O | 0.09 | 0.075 | 0.02 |
| CaCl2 · 2H2O | - | 0.036 | - |
| Na2CO3 | - | 0.2 | - |
| Citric acid | - | 0.006 | - |
| FeEDTA | 1 (mL) | 1 (mL) | - |
| Microelements | (mg/L) | (mg/L) | (mg/L) |
| H3BO3 | 2.86 | 2.86 | 0.05 |
| MnCl2·4H2O | 1.81 | 1.81 | 0.10 |
| ZnSO4·7H2O | 0.22 | 0.22 | 0.01 |
| Co(NO3)2 · 6H2O | 0.05 | 0.05 | 0.01 |
| CuSO4 · 5H2O | 0.08 | 0.08 | 2.50 × 10−6 |
| Na2MoO4 · 2H2O | 0.39 | 0.39 | 0.01 |
| FeSO4 · 7H2O | - | - | 3.50 |
| EDTA | - | - | 4.00 |
2.3. Culture Operations
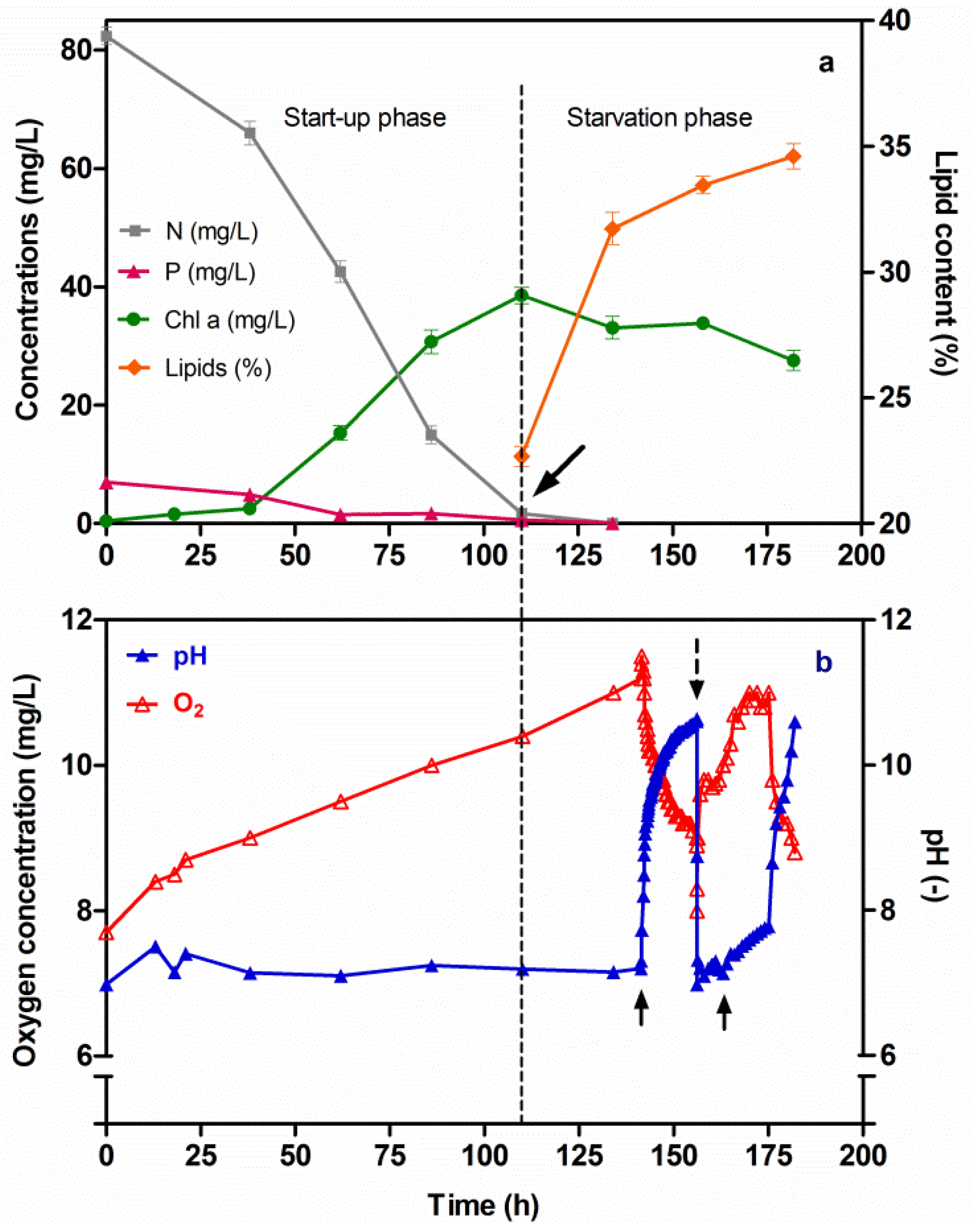
2.3.1. Nitrogen and Phosphorus Starvation Conditions
2.4. Analytical Methods
2.5. Statistical Analyses
3. Results and Discussion
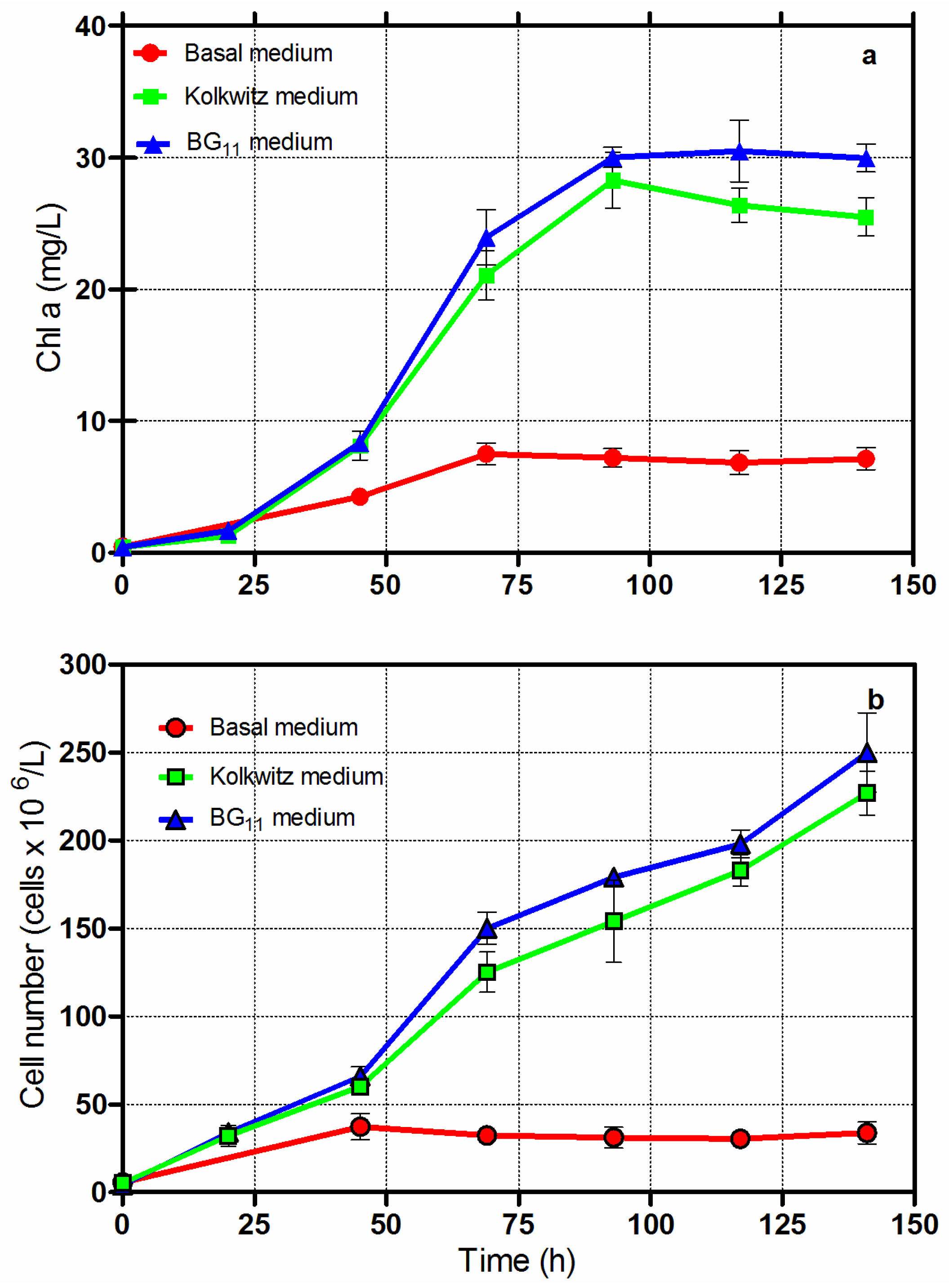
3.1. Cultivation of Chlorella vulgaris CCALA 896 Using Three Different Culture Broths

| Culture broths | Proteins (%) | Carbohydrates (%) | Lipids (%) |
|---|---|---|---|
| Basal | nd 1 | nd | 27.9 ± 0.2 |
| BG-11 | 39.2 ± 1.1 | 24.5 ± 0.3 | 25.3 ± 0.1 |
| Kolkwitz | 37.2 ± 0.8 | 27.7 ± 0.2 | 26.8 ± 0.2 |
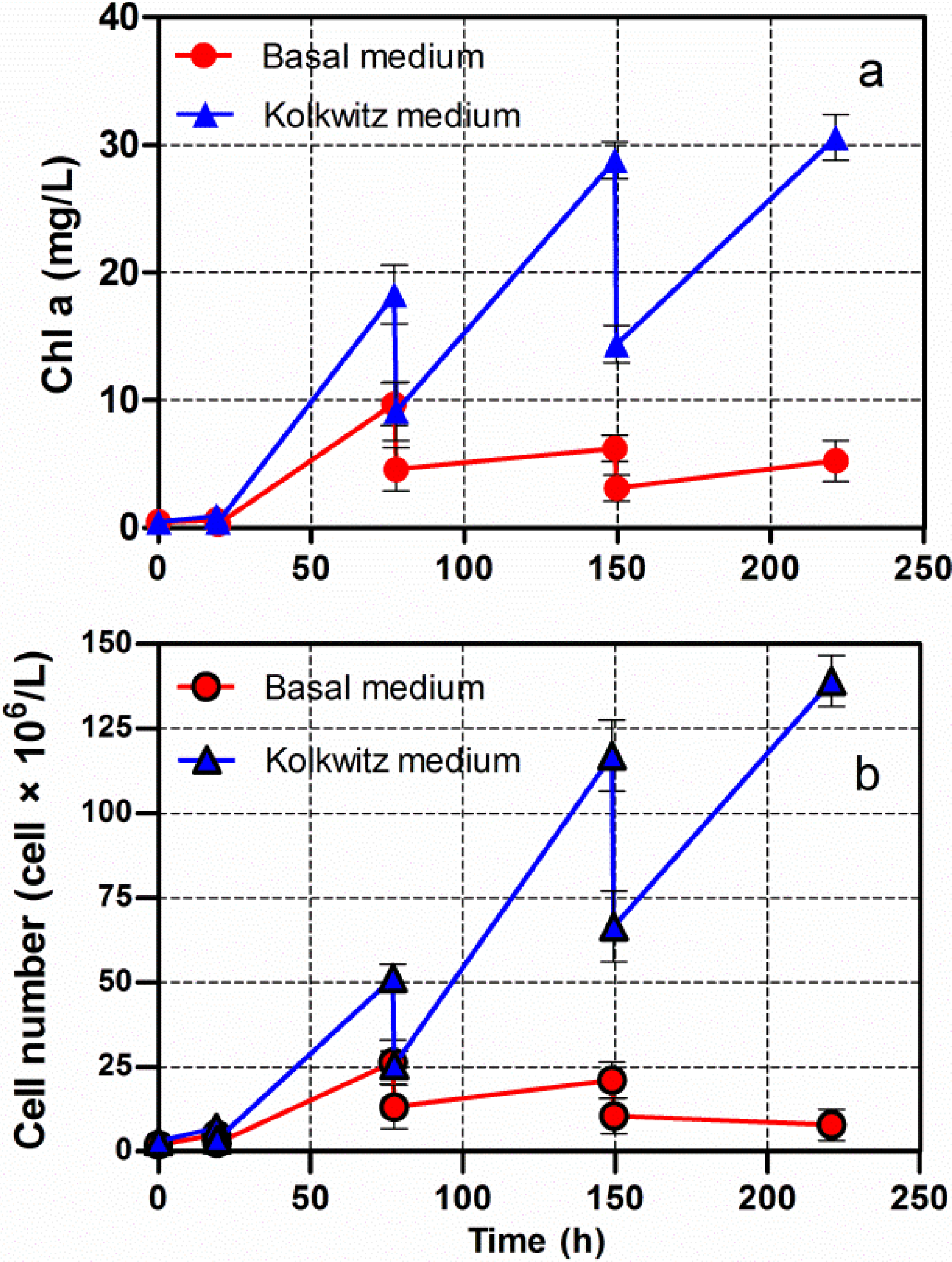
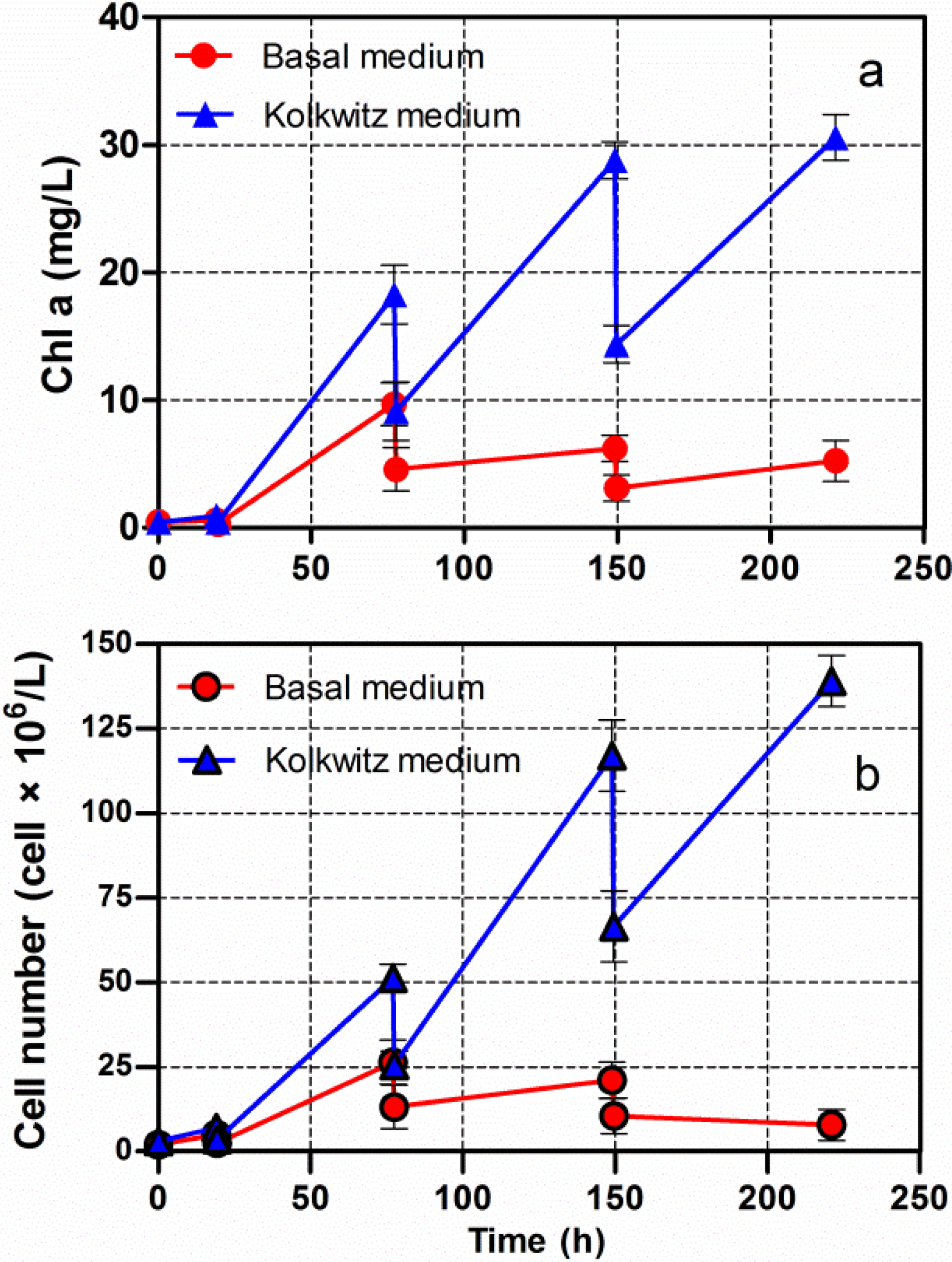
3.2. Cumulative Lipids in Chlorella vulgaris CCALA 896 Grown under Nutritional Starvation Conditions

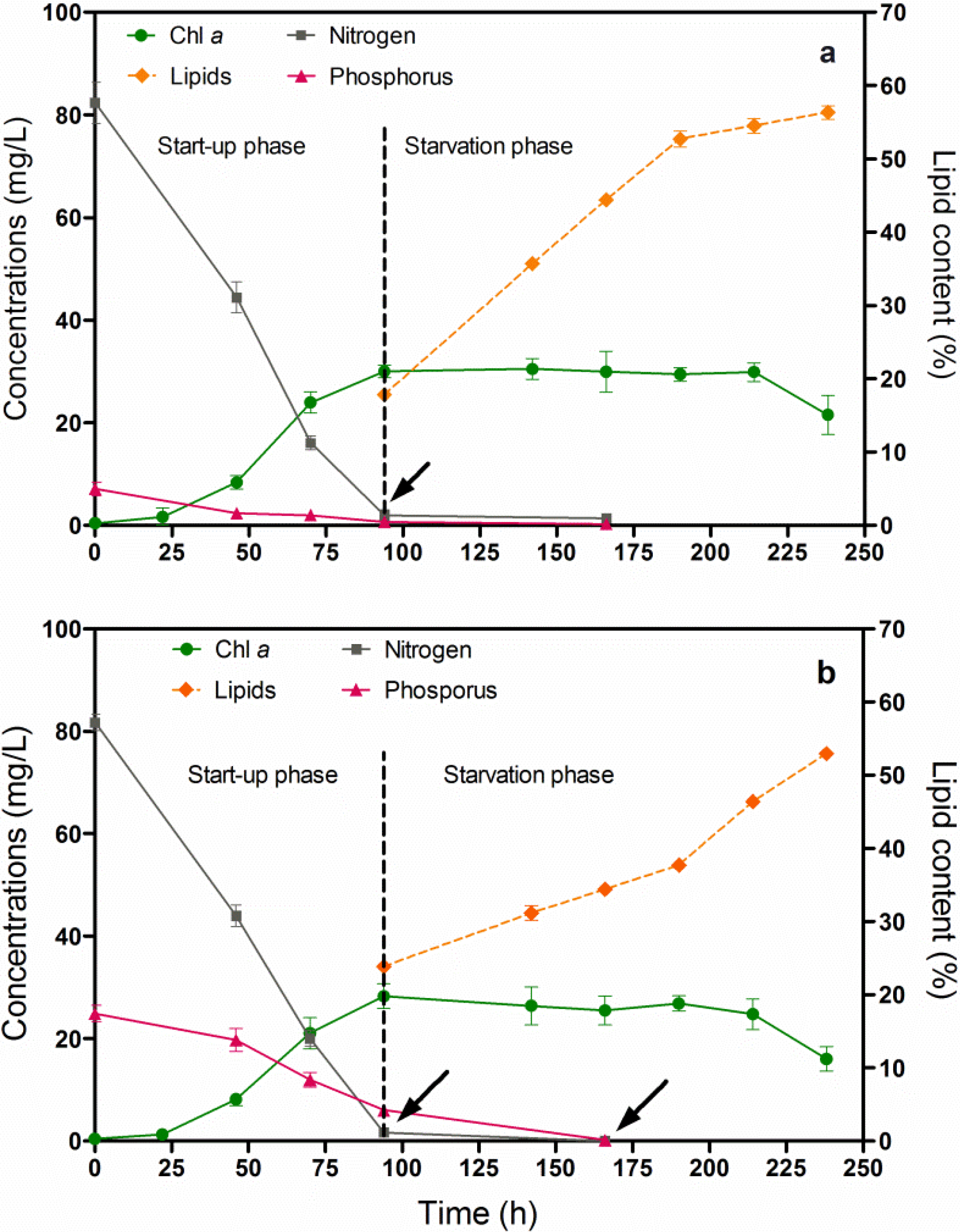
3.3. Changes in the Dry-Biomass Composition of Chlorella vulgaris CCALA 896 Cultured by Using Two Phases: A Cultivation Strategy (A Nutrient-Sufficient Phase Followed by a N- and P- Deprived Phase)
| Time (h) | Biomass composition | |||||
|---|---|---|---|---|---|---|
| Lipids (%) | Proteins (%) | Carbohydrates (%) | ||||
| BG-11 | Kolkwitz | BG-11 | Kolkwitz | BG-11 | Kolkwitz | |
| T0 | 20.9 ± 0.07 | nd 1 | 45.1 ± 0.03 | nd | 27.6 ± 0.02 | nd |
| T94 | 17.9 ± 0.01 | 23.9 ± 0.01 | 42.6 ± 1.03 | 37.5 ± 0.17 | 28.8 ± 0.20 | 28.7 ± 0.15 |
| T142 | 35.7 ± 0.06 | 31.1 ± 0.06 | 30.3 ± 0.40 | nd | 30.7 ± 0.15 | nd |
| T166 | 44.4 ± 0.05 | 34.4 ± 0.03 | nd | 27.2 ± 0.01 | nd | 34.7 ± 0.18 |
| T190 | 52.7 ± 0.11 | 46.4 ± 0.03 | 25.9 ± 0.11 | nd | 18.6 ± 0.08 | nd |
| T214 | 54.5 ± 0.10 | 49.7 ± 0.02 | nd | nd | nd | 24.5 ± 0.01 |
| T238 | 56.3 ± 0.09 | 53.0 ± 0.02 | 24.1 ± 0.10 | 25.6 ± 0.12 | 15.2 ± 0.13 | nd |
| Medium | Specific growth rate (h−1) | Biomass productivity (g (dw)/L/d) | Final lipid content (%) | Lipid productivity (g/L/d) |
|---|---|---|---|---|
| Modified BG-11 | 0.0892 ± 0.0061 | 0.200 ± 0.047 | 56.3 ± 0.9 | 0.113 ± 0.050 |
| Modified Kolkwitz | 0.0773 ± 0.0080 | 0.190 ± 0.028 | 53.0 ± 0.2 | 0.101 ± 0.030 |
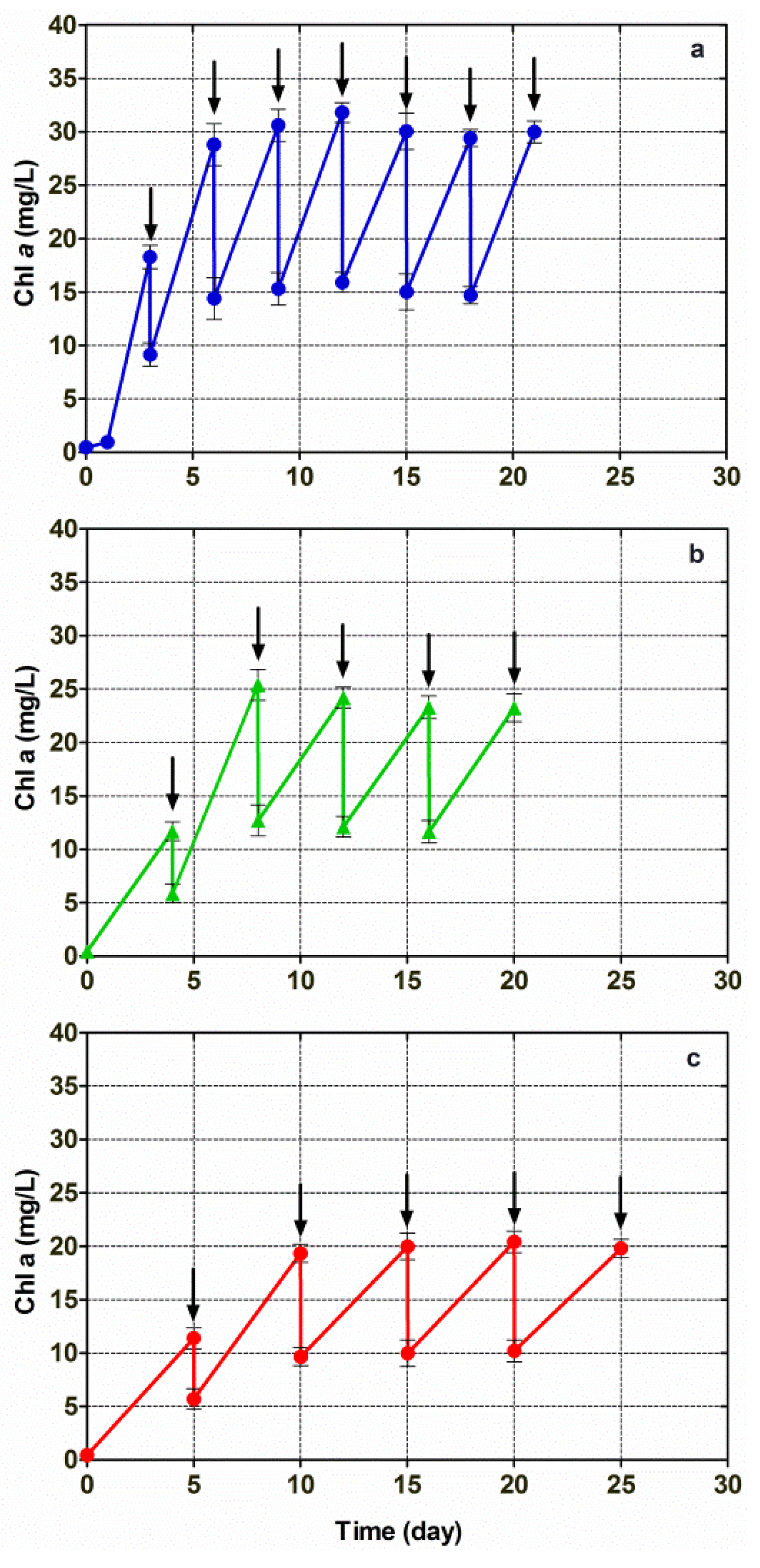
3.4. Cultivation of Chlorella vulgaris CCALA 896 in A Modified Kolkwitz Medium under the Conditions of Three Different Repetitive Batch-Growth Regimens
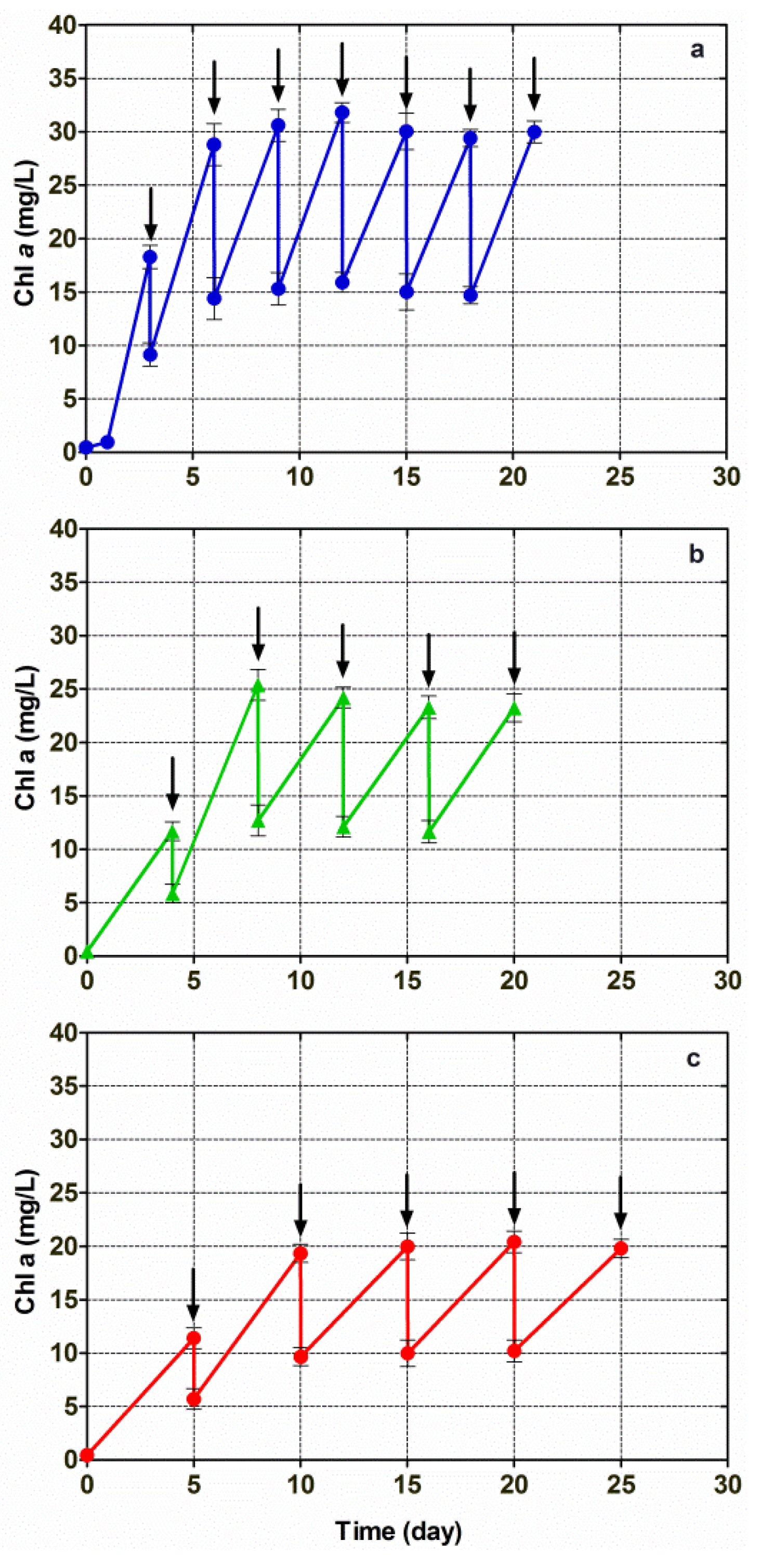

| Chlorella (strains) | PBRs 1 | Growth mode | Light intensity (µE/m2/s) | Media | T (°C) | Biomass productivity (g/L/d) | Lipids | References | |
|---|---|---|---|---|---|---|---|---|---|
| Content (%) | Productivity (g/L/d) | ||||||||
| C. vulgaris CCALA 896 | Cylindrical | B 2 | 150 | BG-11 | 30 | 0.200 | 56.3 | 0.113 | Our results |
| C. sp. | Rectangular | B | 600 | ASW 5 | 30 | - | 52.2 | 0.124 | [40] |
| C. sp. | Rectangular | SCR | 600 | ASW | 30 | - | 43.65 | 0.139 | [40] |
| C. vulgaris CCAP 211/11B | Helical | B | 130 | -- | 25 | 0.024 | 58.0 | - | [23] |
| C. vulgaris AG10032 | Column | B/FB 4 | 200 | BG-11 | 18/25 | 0.145 | 53.0 | 0.077 | [41] |
| C. vulgaris 2714 | Flasks | B | - | OCM 6 | 26 | - | 27.38 | - | [42] |
| C. vulgaris ESP-31 | ns | B | 60 | Basal | 25 | - | 22.0 | 0.056 | [24] |
| C. vulgaris CCAP 211 | Erlenmeyer flasks | B | 70 | BBM 7 | 30 | - | - | 0.008 | [43] |
| C. vulgaris | ns 8 | B | 800 lux | - | 22 | - | 62.9 | - | [22] |
| C. vulgaris KCTCAG10032 | ns | B | 150 | BG-11 | 25 | 0.105 | - | 0.007 | [44] |
3.5. Cultivation of Chlorella vulgaris CCALA 896 in a Modified BG-11 Medium Using a Flat-Glass Photobioreactor (FGPBR)


4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Demirbas, A.; Demirbas, M.F. Green Energy and Technology, Algae Energy, Algae as a New Source of Biodiesel, 1st ed.; Springer: London, UK, 2010; pp. 139–157. [Google Scholar]
- Campbell, M.N. Biodiesel: Algae as a renewable source for liquid fuel. Guelph Eng. J. 2008, 1, 2–7. [Google Scholar]
- Demirbas, A. Fuel properties and calculation of higher heating values of vegetable oils. Fuel 1998, 77, 1117–1120. [Google Scholar] [CrossRef]
- Rakopoulos, C.D.; Antonopoulos, K.A.; Rakopoulos, D.C.; Hountalas, D.T.; Giakoumis, E.G. Comparative performance and emissions study of a direct injection diesel engine using blends of diesel fuel with vegetable oils or bio-diesels of various origins. Energy Convers. Manag. 2006, 47, 3272–3287. [Google Scholar]
- Xu, H.; Miao, X.; Wu, Q. High quality biodiesel production from a microalga Chlorella protothecoides by heterotrophic growth in fermenters. J. Biotechnol. 2006, 126, 499–507. [Google Scholar] [CrossRef]
- Demirbas, A. Importance of biodiesel as transportation fuel. Energy Policy 2007, 35, 4661–4670. [Google Scholar] [CrossRef]
- Moser, B.R. Biodiesel production, properties, and feedstocks. In Biofuels: Global Impact on Renewable Energy, Production Agriculture, and Technological Advancements; Tomas, D., Lakshmanan, P., Songstad, D., Eds.; Springer New York: New York, NY, USA, 2011; pp. 285–347. [Google Scholar]
- Chisti, Y. Fuels from microalgae. Biofuels 2010, 1, 233–235. [Google Scholar] [CrossRef]
- Milne, T.A.; Evans, R.J.; Nagle, N. Catalytic conversion of microalgae and vegetable oils to premium gasoline, with shape-selective zeolites. Biomass 1990, 21, 219–232. [Google Scholar] [CrossRef]
- Dote, Y.; Sawayama, S.; Inoue, S.; Minowa, T.; Yokoyama, S. Recovery of liquid fuel from hydrocarbon-rich microalgae by thermochemical liquefaction. Fuel 1994, 73, 1855–1857. [Google Scholar] [CrossRef]
- Milnowa, T.; Yokoyama, S.; Kishimoto, M.; Okakura, T. Oil production from algae cells of Dunaliella tereiolata by direct thermochemical liquefaction. Fuel 1995, 74, 1735–1738. [Google Scholar] [CrossRef]
- Chisti, Y. Biodiesel from microalgae. Biotechnol. Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef]
- Mata, T.M.; Martins, A.A.; Caetano, N.S. Microalgae for biodiesel production and other applications: A review. Renew. Sustain. Energy Rev. 2010, 14, 217–232. [Google Scholar] [CrossRef]
- Sharma, K.K.; Schuhmann, H.; Schenk, P.M. High lipid induction in microalgae for biodiesel production. Energies 2012, 5, 1532–1553. [Google Scholar] [CrossRef]
- Rodolfi, L.; Zittelli, G.C.; Bassi, N.; Padovani, G.; Biondi, N.; Bonini, G.; Tredici, M.R. Microalgae for oil: Strain selection, induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactor. Biotechnol. Bioeng. 2009, 102, 100–112. [Google Scholar]
- Illman, A.M.; Scragg, A.H.; Shales, S.W. Increase in Chlorella strains calorific values when grown in low nitrogen medium. Enzyme Microb. Technol. 2000, 27, 631–635. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Yeh, K.-L.; Aisyah, R.; Lee, D.-J.; Chang, J.-S. Cultivation, photobioreactor design and harvesting of microalgae for biodiesel production: A critical review. Bioresour. Technol. 2011, 102, 71–81. [Google Scholar] [CrossRef]
- Hu, Q. Environmental effects on cell composition. In Handbook of Microalgal Culture: Biotechnology and Applied Phycology; Richmond, A., Ed.; Blackwell Science Ltd.: Oxford, UK, 2004; Volume 1, pp. 83–93. [Google Scholar]
- Jiang, Y.; Hebly, M.; Kleerebezem, R.; Muyzer, G.; van Loosdrecht, M.C.M. Metabolic modeling of mixed substrate uptake for polyhydroxyalkanoate (PHA) production. Water Res. 2011, 45, 1309–1321. [Google Scholar]
- Guest, J.S.; van Loosdrecht, M.C.M.; Skerlos, S.J.; Love, N.G. Lumped pathway metabolic model of organic carbon accumulation and mobilization by the alga Chlamydomonas reinhardtii. Environ. Sci. Technol. 2013, 47, 3258–3267. [Google Scholar]
- Pulz, O. Photobioreactors: Production systems for phototrophic microorganisms. Appl. Microbiol. Biotechnol. 2001, 57, 287–293. [Google Scholar] [CrossRef]
- Piorreck, M.; Baasch, K.H.; Pohl, P. Biomass production, total protein, chlorophylls, lipids and fatty acids of freshwater green and blue-green algae under different nitrogen regimes. Phytochemistry 1984, 23, 207–216. [Google Scholar] [CrossRef]
- Scragg, A.H.; Illman, A.M.; Carden, A.; Shales, S.W. Growth of microalgae with increased calorific values in a tubular bioreactor. Biomass Bioenergy 2002, 23, 67–73. [Google Scholar] [CrossRef]
- Yeh, K.-L.; Chang, J.-S. Effects of cultivation conditions and media composition on cell growth and lipid productivity of indigenous microalga Chlorella vulgaris ESP-31. Bioresour. Technol. 2012, 105, 120–127. [Google Scholar] [CrossRef]
- Olivieri, G.; Salatino, P.; Marzocchella, A. Advances in photobioreactors for intensive microalgal production: Configurations, operating strategies and applications. J. Chem. Technol. Biotechnol. 2014, 89, 178–195. [Google Scholar] [CrossRef]
- Carlozzi, P. Hydrogen photoproduction by Rhodopseudomonas palustris 42OL cultured at high irradiance under a semicontinuous regime. J. Biomed. Biotechnol. 2012, 2012. [Google Scholar] [CrossRef]
- Carlozzi, P.; Pinzani, E. Growth characteristics of Arthrospira platensis cultured inside a new close-coil photobioreactor incorporating a mandrel to control culture temperature. Biotechnol. Bioeng. 2005, 90, 675–684. [Google Scholar] [CrossRef]
- Strickland, J.D.H.; Parsons, T.R. A Practical Hand Book of Seawater Analysis, 2nd ed.; Fisheries Research Board of Canada: Woods Hole, MA, USA, 1972; pp. 185–199. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method for total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Marsh, J.B.; Weinstein, D.B. Simple charring methods for determination of lipids. J. Lipid Res. 1966, 7, 574–576. [Google Scholar]
- Carlozzi, P. The effect of irradiance growing on hydrogen photoevolution and on the kinetic growth in Rhodopseudomonas palustris, strain 420L. Int. J. Hydrog. Energy 2009, 34, 7949–7958. [Google Scholar] [CrossRef]
- Takagi, M.; Watanabe, K.; Yamaberi, K.; Yoshida, T. Limited feeding of potassium nitrate for intracellular lipid and triglyceride accumulation of Nannochloris sp. UTEX LB1999. Appl. Microbiol. Biotechnol. 2000, 54, 112–117. [Google Scholar]
- Li, Y.; Horsman, M.; Wang, B.; Wu, N.; Lan, C.Q. Effects of nitrogen sources on cell growth and lipid accumulation of green alga Neochloris oleoabundans. Appl. Microbiol. Biotechnol. 2008, 81, 629–636. [Google Scholar] [CrossRef]
- Fogg, G.E.; Collyer, D.M. Accumulation of lipids by algae. In Algal Culture: From Laboratory to Pilot Plant; Burlew, J.S., Ed.; Carnegie Institution of Washington: Washington, DC, USA, 1953; Volume 1, pp. 177–181. [Google Scholar]
- Xin, L.; Hu, H.; Ke, G.; Sun, Y. Effects of different nitrogen and phosphorus concentrations on the growth, nutrient uptake, and lipid accumulation of freshwater microalga Scenedesmus sp. Bioresour. Technol. 2010, 101, 5494–5500. [Google Scholar] [CrossRef]
- Pittman, J.K.; Dean, A.P.; Osundeko, O. The potential of sustainable algal biofuel production using wastewater resources. Bioresour. Technol. 2011, 102, 17–25. [Google Scholar]
- Yeh, K.-L.; Chen, Y.-C.; Chang, J.-S. pH-stat photoheterotrophic cultivation of indigenous Chlorella vulgaris ESP-31 for biomass and lipid production using acetic acid as the carbon source. Biochem. Eng. J. 2012, 64, 1–7. [Google Scholar] [CrossRef]
- Hsieh, C.-H.; Wu, W.-T. Cultivation of microalgae for oil production with a cultivation strategy of urea limitation. Bioresour. Technol. 2009, 100, 3921–3926. [Google Scholar] [CrossRef]
- Mujtaba, G.; Choi, W.; Lee, C.-G.; Lee, K. Lipid production by Chlorella vulgaris after a shift from nutrient-rich to nitrogen starvation conditions. Bioresour. Technol. 2012, 123, 279–283. [Google Scholar] [CrossRef]
- Heredia-Arroyo, T.; Wei, W.; Ruan, R.; Hu, B. Mixotrophic cultivation of Chlorella vulgaris and its potential application for the oil accumulation from non-sugar materials. Biomass Bioenergy 2011, 35, 2245–2253. [Google Scholar]
- Converti, A.; Casazza, A.A.; Ortiz, E.Y.; Perego, P.; Borghi, M.D. Effect of temperature and nitrogen concentration on the growth and lipid content of Nannochloropsis oculata and Chlorella vulgaris for biodiesel production. Chem. Eng. Process. Process Intensif. 2009, 48, 1146–1151. [Google Scholar] [CrossRef]
- Yoo, C.; Jun, S.-Y.; Lee, J.-Y.; Ahn, C.-Y.; Oh, H.-M. Selection of microalgae for lipid production under high levels carbon dioxide. Bioresour. Technol. 2010, 101, S71–S74. [Google Scholar] [CrossRef]
- Li, Y.; Hua, D.; Zhang, J.; Gao, M.; Zhao, Y.; Xu, H.; Liang, X.; Jin, F.; Zhang, X. Influence of inoculum to substrate ratios (ISRs) on the performance of anaerobic digestion of algal residues. Ann. Microbiol. 2013. [Google Scholar] [CrossRef]
- Castrillo, M.; Lucas-Salas, L.M.; Rodriguez-Gil, C.; Martinez, D. High pH-induced flocculation-sedimentation and effect of supernatant reuse on growth rate and lipid productivity of Scenedesmus obliquus and Chlorella vulgaris. Bioresour. Technol. 2013, 128, 324–329. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Vaičiulytė, S.; Padovani, G.; Kostkevičienė, J.; Carlozzi, P. Batch Growth of Chlorella Vulgaris CCALA 896 versus Semi-Continuous Regimen for Enhancing Oil-Rich Biomass Productivity. Energies 2014, 7, 3840-3857. https://doi.org/10.3390/en7063840
Vaičiulytė S, Padovani G, Kostkevičienė J, Carlozzi P. Batch Growth of Chlorella Vulgaris CCALA 896 versus Semi-Continuous Regimen for Enhancing Oil-Rich Biomass Productivity. Energies. 2014; 7(6):3840-3857. https://doi.org/10.3390/en7063840
Chicago/Turabian StyleVaičiulytė, Sigita, Giulia Padovani, Jolanta Kostkevičienė, and Pietro Carlozzi. 2014. "Batch Growth of Chlorella Vulgaris CCALA 896 versus Semi-Continuous Regimen for Enhancing Oil-Rich Biomass Productivity" Energies 7, no. 6: 3840-3857. https://doi.org/10.3390/en7063840