
The Effect of Chromium on Photosynthesis and Lipid Accumulation in Two Chlorophyte Microalgae
by
 , , and
, , and
Khawaja Muhammad Imran Bashir
1 ,
,
Hyeon-Jun Lee
2,
Sana Mansoor
3,
Alexander Jahn
1 and
Man-Gi Cho
4,* 1
German Engineering Research and Development Center, LSTME-Busan, Busan 46742, Korea
2
Department of Chemical and Biological Engineering, Institute of Fluid Mechanics, Friedrich-Alexander-Universität Erlangen-Nürnberg Busan Campus, Busan 46742, Korea
3
Department of Environmental Engineering, College of Engineering, Dong-A University, Busan 49315, Korea
4
Department of Biotechnology, Division of Energy and Bioengineering, Dongseo University, Busan 47011, Korea
*
Author to whom correspondence should be addressed.
Energies 2021, 14(8), 2260; https://doi.org/10.3390/en14082260
Submission received: 4 March 2021
/
Revised: 9 April 2021
/
Accepted: 16 April 2021
/
Published: 17 April 2021
Abstract
:Heavy metals have adverse effects on microalgae metabolism and growth. Photosynthesis and lipid profile are quite sensitive to heavy metal toxicity. The impact of hexavalent chromium—Cr(VI) on photosynthesis and lipid accumulation in Mucidosphaerium pulchellum and Micractinium pusillum exposed to different concentrations (0–500 μg L−1) was investigated for 11 days. A significant (p < 0.05) increase in lipid content was observed with increasing Cr(VI) concentration. However, growth was suppressed at higher concentrations exceeding 100 μg L−1. Addition of Cr(VI) in the cell culture medium showed a negative effect on quantum yield (Fv/Fm), and a photosynthetic inhibition of >65% was noted in both species at 500 μg L−1. However, the lipid gravimetric analysis presented inner cell lipid content up to 36% and 30% of dry weight biomass for M. pulchellum and M. pusillum, respectively. The fatty acids profiles of both microalgae species showed higher levels of hexadecenoic acid as well as ω3, ω6, and ω7 fatty acids. The effect of Cr(VI) on photosynthesis and lipid accumulation in both microalgae species was concentration and exposure time dependent. This shows that an appropriate concentration of Cr(VI) in culture medium could be beneficial for higher lipid accumulation in freshwater eukaryotic microalgae species.
1. Introduction
The use of microalgae in the pharmaceutical, medical and food industries is increasing due to their no reported side effects on human health. Thus, it is necessary to investigate microalgae growth parameters and potential growth inhibitors [1]. Heavy metals, antibiotics and herbicides are toxic to microalgae even at low concentrations; however, to a certain extent, microalgae could adapt to a contaminated environment [2,3,4,5]. Microalgae, one of the primary producers in the marine ecosystem, change their composition such as fatty acids, lipids, pigments and exopolymers when exposed to heavy metal contamination [6,7]. Therefore, investigating the adaptation and survival of microalgae species in a contaminated environment is imperative [8,9,10,11]. Investigating the effect of heavy metal toxicity on growth and photosynthetic activity of microalgae could substantially influence overall ecological risk evaluation of heavy metals.
Rapid industrialization and urbanization have increased the environmental presence of heavy metals, which is causing water pollution, and it is becoming a serious issue worldwide [12]. Several methods have been developed to reduce the concentration of heavy metals from wastewater [13]; however, non-biodegradable heavy metals resist bioremediation and show long environmental persistence times [14,15], which is posing a serious threat for the biotic life. Heavy metals are abundantly available in nature. Unlike other heavy metals such as zinc (Zn) and copper (Cu), chromium (Cr) is not required for plant respiration or growth. However, due to rapid industrialization, an abundant quantity of Cr is being dumped into the water, causing unfavorable effects on animals and plants [12]. Furthermore, Cr toxicity is form-dependent, with hexavalent chromium—Cr(VI)—being significantly more toxic to humans than trivalent chromium—Cr(III) [16]. Trace amounts of Cr(III) are needed daily for adult humans [17]. On the other hand, Cr(VI) has carcinogenic and mutagenic effects on humans [17]. Severe acute effects including gastrointestinal disorders, hemorrhagic diathesis and convulsions could occur when ingesting 1–5 g of chromate [17,18].
The exposure of microalgae to heavy metals could cause inhibited growth, suppressed cell division, reduced photosynthesis, and restrained enzymatic activity [19,20,21]. Advancement in chlorophyll fluorescence technology has made easier to analyze the photosynthesis processes, which has enabled researchers in finding new factors affecting photosynthesis. Even though the impact of heavy metals on microalgae has gained considerable attention and has been extensively reported, the effects of Cr(VI) toxicity on Chlorophyte microalgae species Mucidosphaerium pulchellum (formerly Dictyosphaerium pulchellum) and Micractinium pusillum have not been yet reported.
In the present study, the toxic effects of heavy metal hexavalent chromium on growth and modulated fluorescence of freshwater eukaryotic non-model microalgae species M. pulchellum and M. pusillum were analyzed and compared. Furthermore, the effect of chromium on lipid accumulation and lipid composition was also investigated.
2. Materials and Methods
2.1. Microalgae Culture and Treatment
The eukaryotic freshwater microalgae species Mucidosphaerium pulchellum and Micractinium pusillum (phylum Chlorophyta, class Trebouxiophyceae) were obtained from the Korea Marine Microalgae Culture Center (KMMCC), Busan, Korea. Stocks were maintained on modified-AF6 agar slants [22]. The microalgae species were cultured in 250 mL flask bioreactors containing modified-AF6 medium [23], without citrate and ETDA, at a constant light intensity of 50 ± 2 µmol photons m−2 s−1, 25 ± 2 °C temperature and 50% humidity. The effects of potassium dichromate (K2Cr2O7, Yakuri Pure Chemicals, Osaka, Japan), a hexavalent form of chromium—Cr(VI), on photosynthesis and lipid accumulation at different concentrations (0 to 500 µg L−1) were investigated. All experiments were repeated at least three times.
2.2. Determination of Microalgae Growth Performance
Growth performance was determined by measuring the cell densities, optical densities and growth rates. Growth rates were calculated using the cell densities. Briefly, on each sampling day, 5 mL sample was collected from each culture flask after a thorough hand mixing and cell densities were determined using hemocytometer (Paul Marienfeld GmbH & Co. KG, Lauda-Königshofen, Germany) under a light microscope at a magnification of 400×. Furthermore, optical density at 750 nm (OD750) was recorded using UV–VIS spectrophotometer (WPA Biowave II, Biochrom Ltd., Cambridge, UK) on every alternating day [5].
2.3. Measurement of Toxicity, Modulated Fluorescence and Photosynthetic Inhibition
The sensitivity of M. pulchellum and M. pusillum in media supplemented with different concentrations of Cr(VI) was evaluated by toxy-PAM dual channel yield analyzer (Heinz Walz GmbH, Effeltrich, Germany) [5]. This toxicity analyzer is extremely sensitive to chlorophyll fluorescence and uses saturation pulse method to determine the effective fluorescence yield of photosystem II (PSII) [24,25]. To induce an equilibrium state for the photosynthetic electron transport, microalgae samples were dark adopted, by placing in complete dark for 30 min before analysis, and fluorescence intensity was measured using low intensity modulated light to avoid the reduction of the PSII primary electron acceptor (QA) [24].
Fluorescence intensity of microalgae cells excited by toxy-PAM blue light was measured at 650 nm. The minimal fluorescence level (F0; fluorescence measured shortly before the application of a saturation pulse) and the maximal fluorescence level (Fm; fluorescence measured during a saturation pulse) were recorded, and the effective overall quantum yield (Y) of PSII was calculated using the following equation [24]:
Quantum Yield (Y) = (Fm − F0)/Fm = Fv/Fm
Relative photosynthetic inhibition was calculated using the following equations [5].
Relative Photosynthetic Inhibition% = 100 (Y2 − Y1)/Y2 (Y1 < Y2)
Relative Photosynthetic Inhibition% = 100 (Y2 − Y1)/Y2 (Y1 > Y2)
2.4. Lipid Extraction
The tested microalgae species were grown in modified-AF6 medium supplemented with different concentrations of Cr(VI) until pre-stationary phase, and total lipids were extracted following the Bligh and Dyer method with slight modifications [26]. Briefly, microalgae cells were harvested by centrifugation at 4 °C and 5000× g for 20 min. Pellets were rinsed with distilled water and freeze-dried at −85 °C. Dried pellets were weighed, dissolved in 10 mL of methanol: chloroform (2:1) solution and homogenized using ULTRA TURRAX Homogenizer (IKA-Werke GmbH & Co. KG, Staufen, Germany at 16,000 rpm for 2 min. Samples were incubated overnight in the dark at room temperature. The following day, 6 mL distilled water and 3 mL chloroform was added in each tube, inverted carefully and centrifuged at 5000× g for 10 min. The lower organic phase was transferred to a new 15 mL falcon tube, freeze-dried overnight at −85 °C and dried for another 4 h at 50 °C in a dry-oven until all the solvent was evaporated. The tubes were weighed, and total lipid content in each sample was calculated.
2.5. Fatty Acid Analysis
Total fatty acids extracted from M. pulchellum and M. pusillum were analyzed by gas chromatography (GC) of fatty acid methyl esters (FAMEs) prepared according to Ichihara and Fukubayashi [27], with slight modifications. Briefly, total fatty acids were trans-methylated with 3 mL of methanolic-hydrochloric acid solution at 90 °C for 1 h. Samples were cooled at room temperature. A total of 1 mL hexane and 1 mL distilled water was added to the extracts and mixed well by vortex. The upper fatty acid layer was separated by centrifugation at 3000× g for 6 min and collected in GC vials.
GC analysis of FAMEs was conducted using PerkinElmer Clarus 680 Gas Chromatograph (PerkinElmer, Inc., Waltham, MA, USA) equipped with 30 m × 20 μm × 25 mm SP−2380 capillary column (Sigma-Aldrich, Co., St. Louis, MO, USA). The initial oven temperature was set to 140 °C and increased at a rate of 4 to 240 °C. Helium (He) was used as a carrier gas at a flow rate of 1 mL/min. Injection temperature and volume were set at 230 °C and 1 µL, respectively. Fatty acids were identified by comparison with retention times of 37-component FAME mix standards (Sigma-Aldrich) and were expressed as mg/L. The corresponding fatty acids were further cross checked with the instrument database containing the NIST® library [28].
2.6. Statistical Analysis
The statistical significance of the results was calculated using Analysis of Variance via SPSS ver. 27 (SPSS, Chicago, IL, USA). The significance among the samples was assessed using Duncan’s multiple-range test and the results were considered statistically significant at p < 0.05.
3. Results
3.1. Influence of Chromium on Microalgal Growth
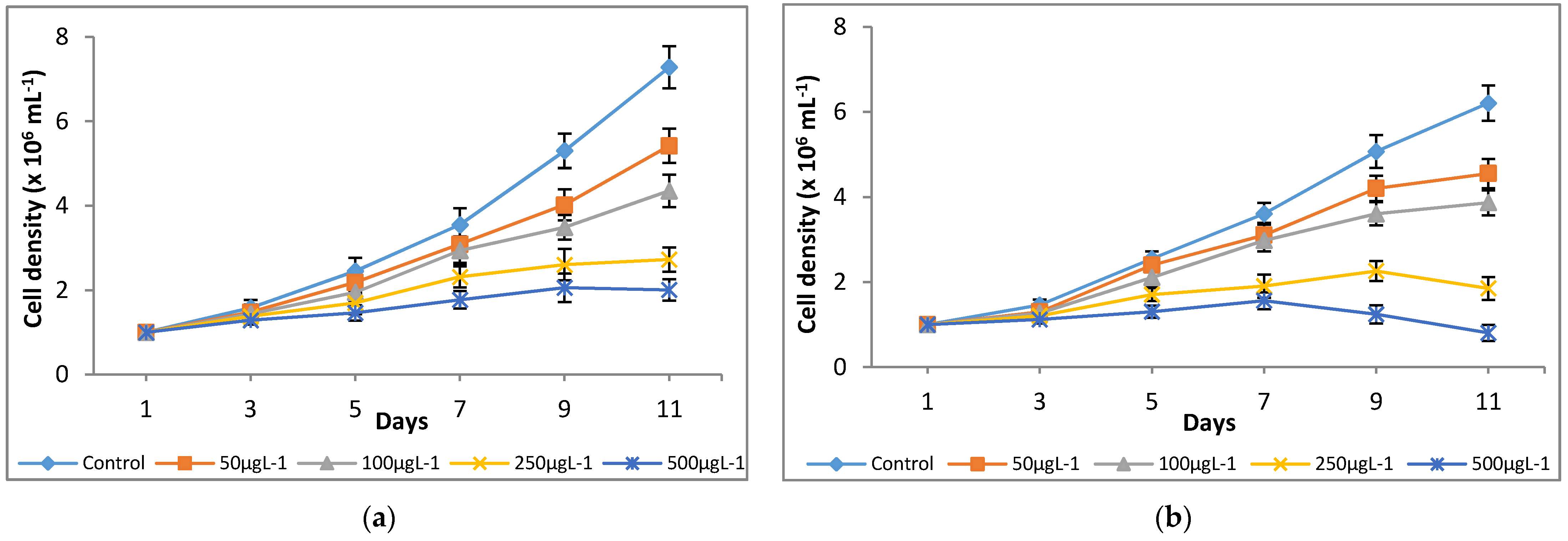
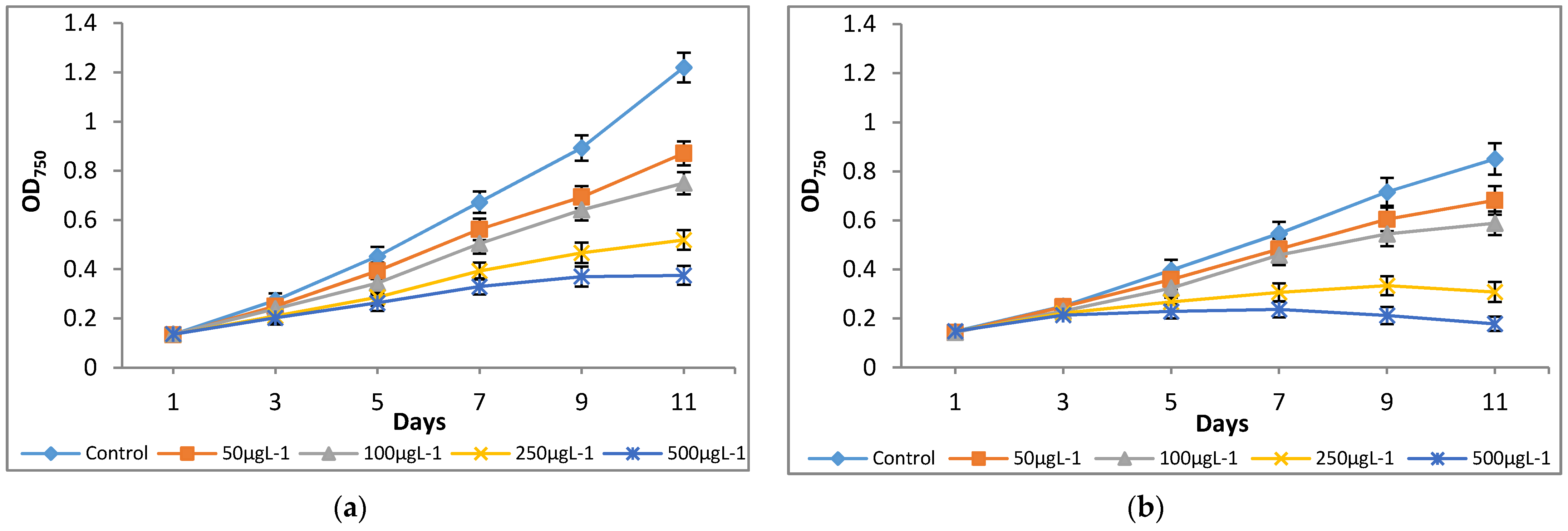
The culture medium, modified-AF6 medium, was supplemented with different concentrations of Cr(VI) ranging from 0 to 500 μg L−1, and their effects on growth parameters were investigated for 11 days. The microalgae species M. pulchellum and M. pusillum showed significant sensitivities to the tested Cr(VI) concentrations as indicated by the optical density and cell density values. As shown in Figure 1, Cr concentrations, 0–100 μg L−1, showed normal cell densities whereas the concentrations exceeding 100 μg L−1 here, 250 μg L−1 and 500 μg L−1, showed a decline in growth after day 7. Both microalgae cultures showed reduced growth as compared to the control; it could be due to the possible toxicity of the Cr(VI). Interestingly, growth measurement study at absorbance of 750 nm (OD750) showed similar results with the cell densities (Figure 2).
Significant (p < 0.05) differences in growth rates among chromium treatments for both microalgae species were observed. For M. pulchellum, control (0 μg L−1) and 50 μg L−1 chromium treatments showed exponential growth at μ = 0.1912 and μ = 0.1605, respectively, while the Cr(VI) induced inhibition of the PSII resulted in a pseudo substrate limited linear growth at 0.3688 × 106 (100 μg L−1 chromium), 0.2139 × 106 (250 μg L−1 chromium) and 0.1311 × 106 (500 μg L−1 chromium) cell per day. Furthermore, a significant reduction in growth rate was observed at Cr(VI) concentrations, 250 μg L−1 and 500 μg L−1, after day 10. At the given photon flux density, the control cultivation for M. pusillum was substrate limited and showed a linear growth at 0.6007 × 106 cells per day. Increasing concentrations of Cr(VI) decreased growth rates in a similar manner to M. pulchellum, with a growth rate of 0.4706 × 106 (50 μg L−1 chromium), 0.3937 × 106 (100 μg L−1 chromium), 0.168 × 106 (150 μg L−1 chromium) and 0.1110 × 106 (500 μg L−1 chromium) cells per day (Table 1). The 50, 100 and 250 μg L−1 chromium treatments showed reduction in growth after day nine, while 500 μg L−1 chromium treatment showed reduction in growth rate and cell number after day 7.
3.2. Effect of Chromium on Modulated Fluorescence and Photosynthetic Inhibition
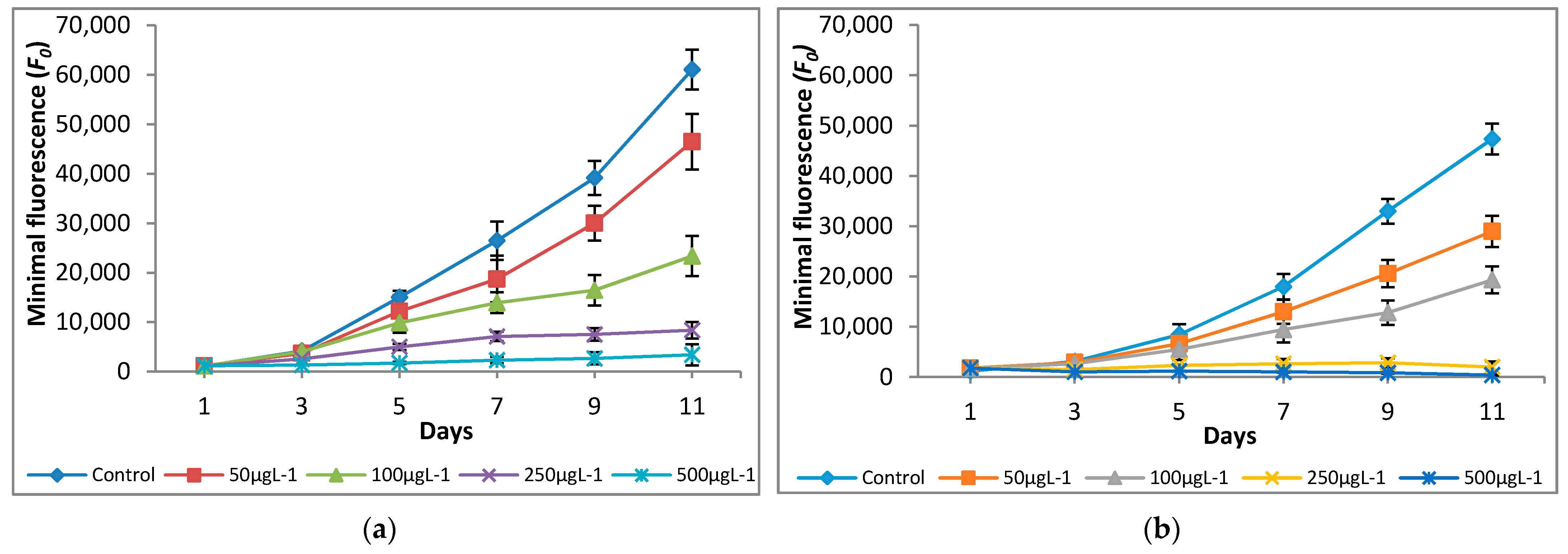
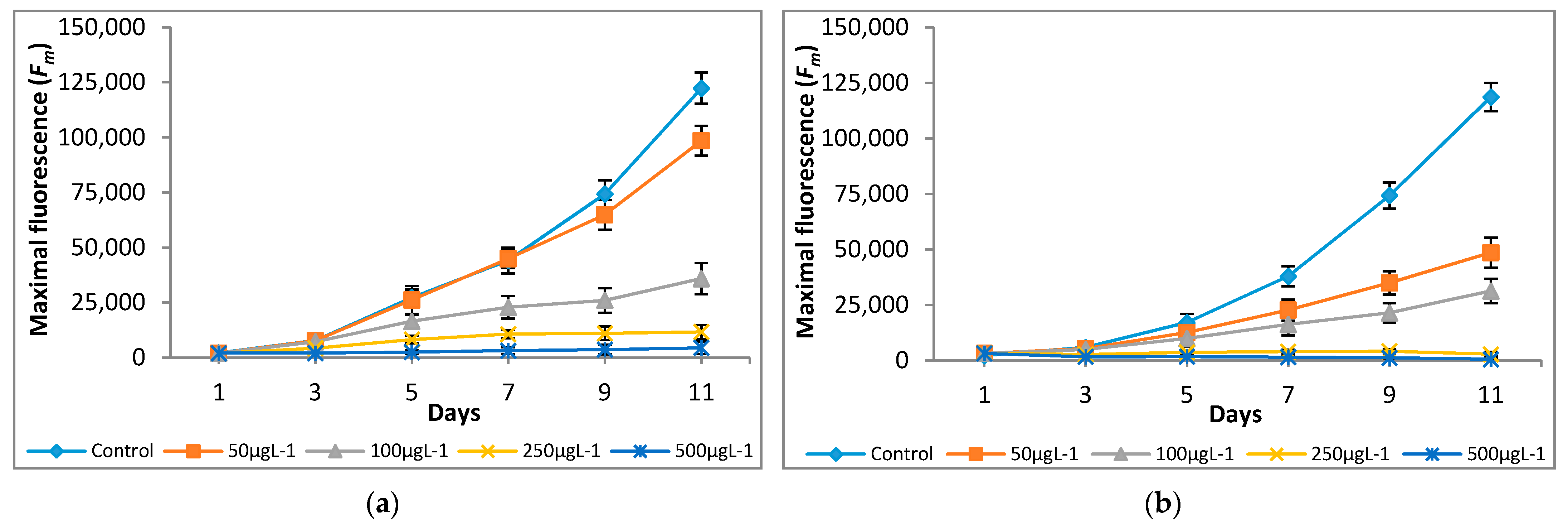
The variations in quantum yield and photosynthetic inhibition of the tested microalgae cultures supplemented with different concentrations of Cr(VI) were studied for 11 days. Different Cr(VI) concentrations showed significant differences in fluorescence yield and photosynthetic inhibition. The significant (p < 0.05) increases in minimal (F0) and maximal (Fm) fluorescence values were observed at 0 μg L−1—100 μg L−1, however, a decline in F0 and Fm values was observed at 250 μg L−1 and 500 μg L−1 (Figure 3 and Figure 4).
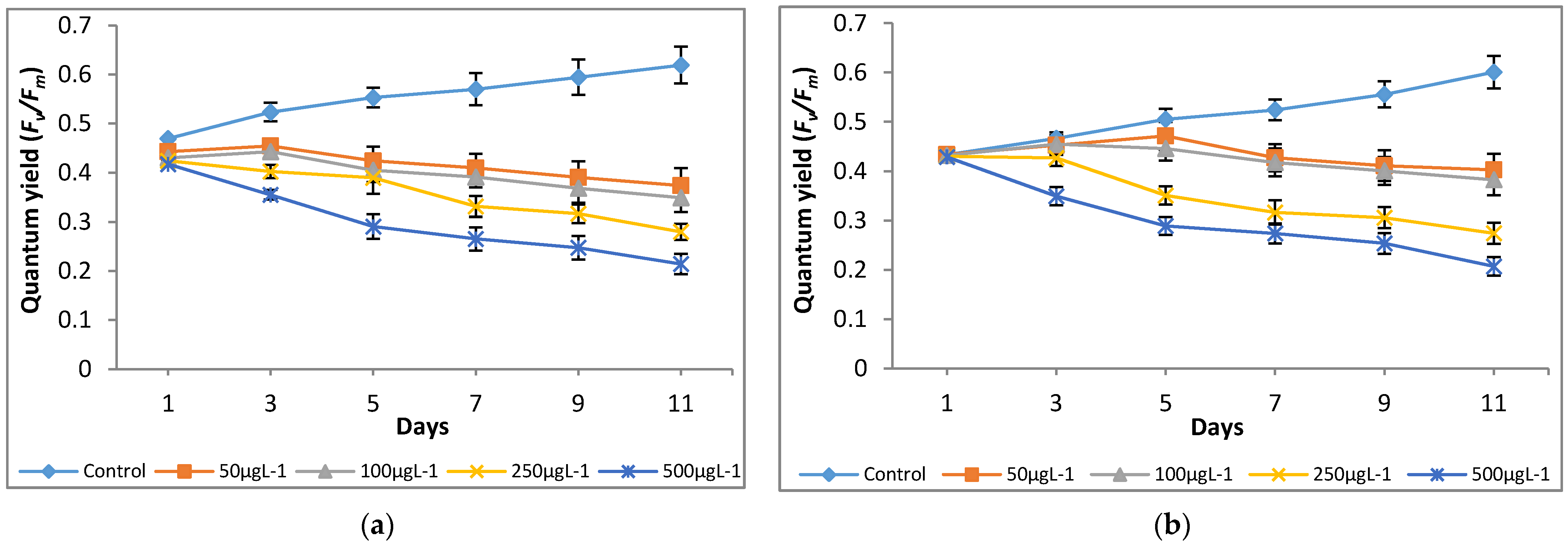
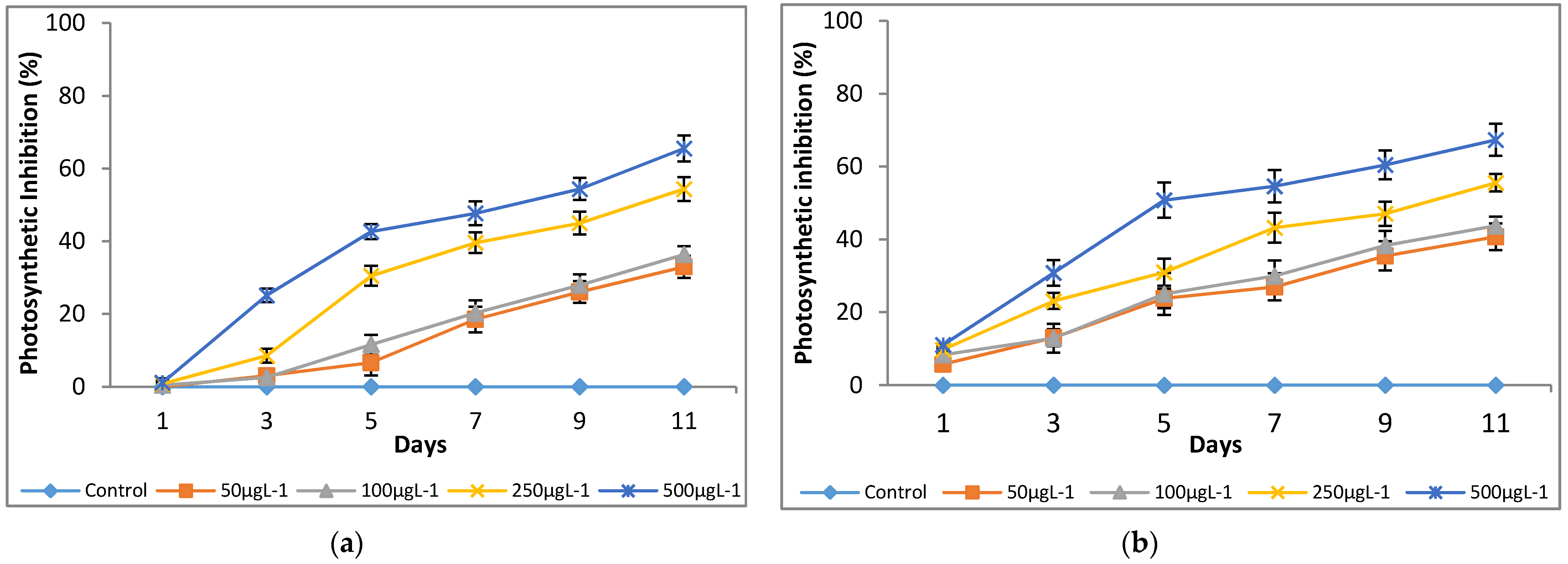
A significant (p < 0.05) reduction in quantum yield (Y) was noted at all the tested Cr(VI) concentrations as compared to the control (0 μg L−1; Figure 5). It shows that chromium had a negative effect on the tested microalgae growth by interrupting photosynthesis. Both microalgae species showed a similar trend of photosynthetic inhibition at the tested Cr(VI) concentrations after day 3 of culturing and a maximum inhibition of up to 67%, and 66% was observed for M. pulchellum and M. pusillum, respectively, at 500 μg L−1 (Figure 6).
3.3. Effect of Chromium on Lipid Accumulation
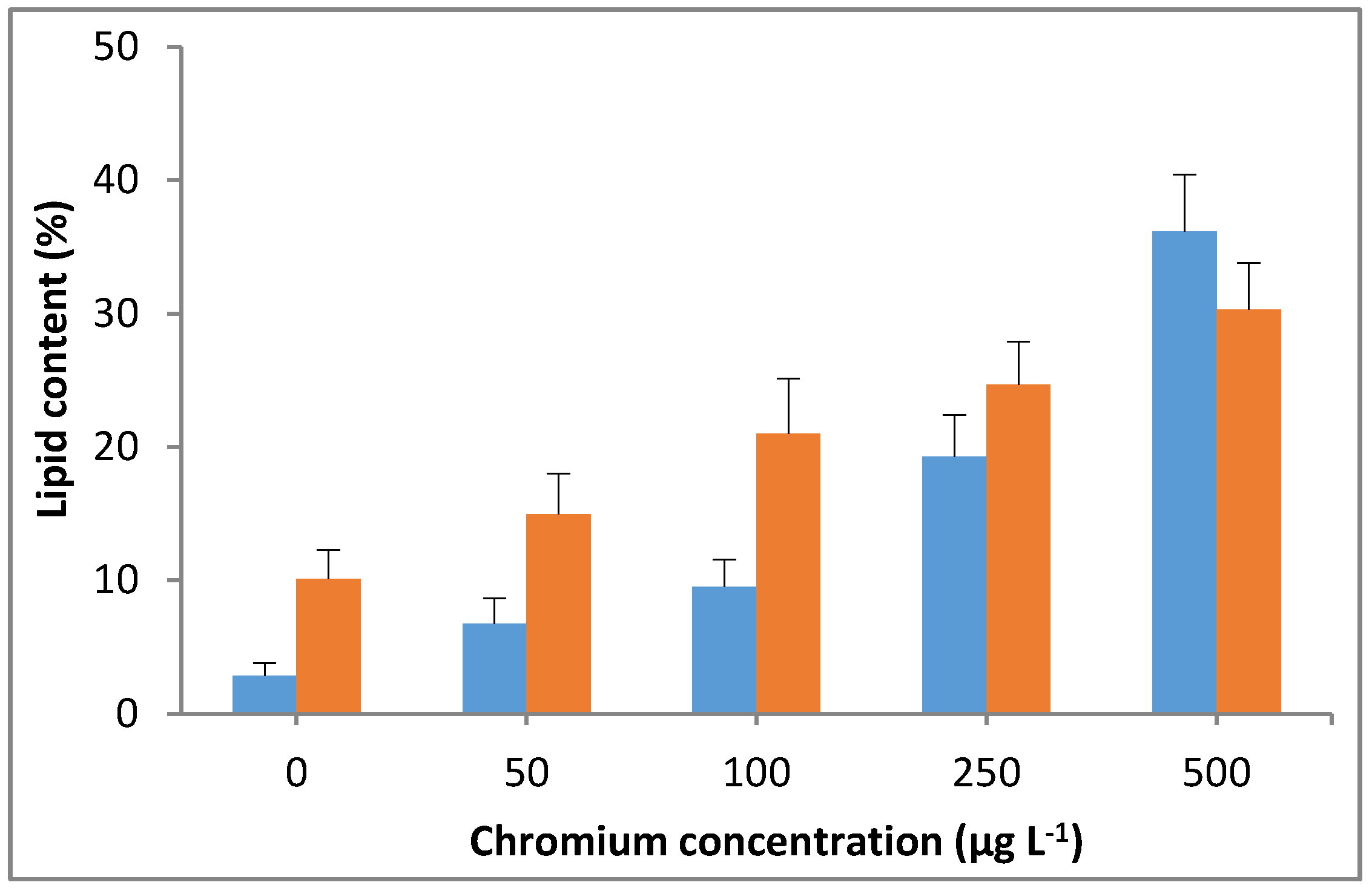
The significant (p < 0.05) increases in lipid content were observed with the increasing Cr(VI) concentration in the culture medium. A lipid content of up to 36% was observed in M. pulchellum cultures at the maximum tested Cr(VI) concentration in this study (500 µg L−1), which was approximately 12 times higher than the control (0 µg L−1). Whereas a maximum lipid content of 30% was observed in M. pusillum cultures, which was approximately 3 times higher than the control. A drastic increase in Cr(VI) concentration-dependent lipid content was observed in M. pulchellum; however, the increase in concentration-dependent lipid content was not consistent in M. pusillum (Figure 7). However, both tested microalgae species showed significantly (p < 0.05) increased lipid content, which shows that the addition of chromium could significantly enhance the lipid accumulation in the tested freshwater microalgae species.
3.4. Fatty Acid Composition
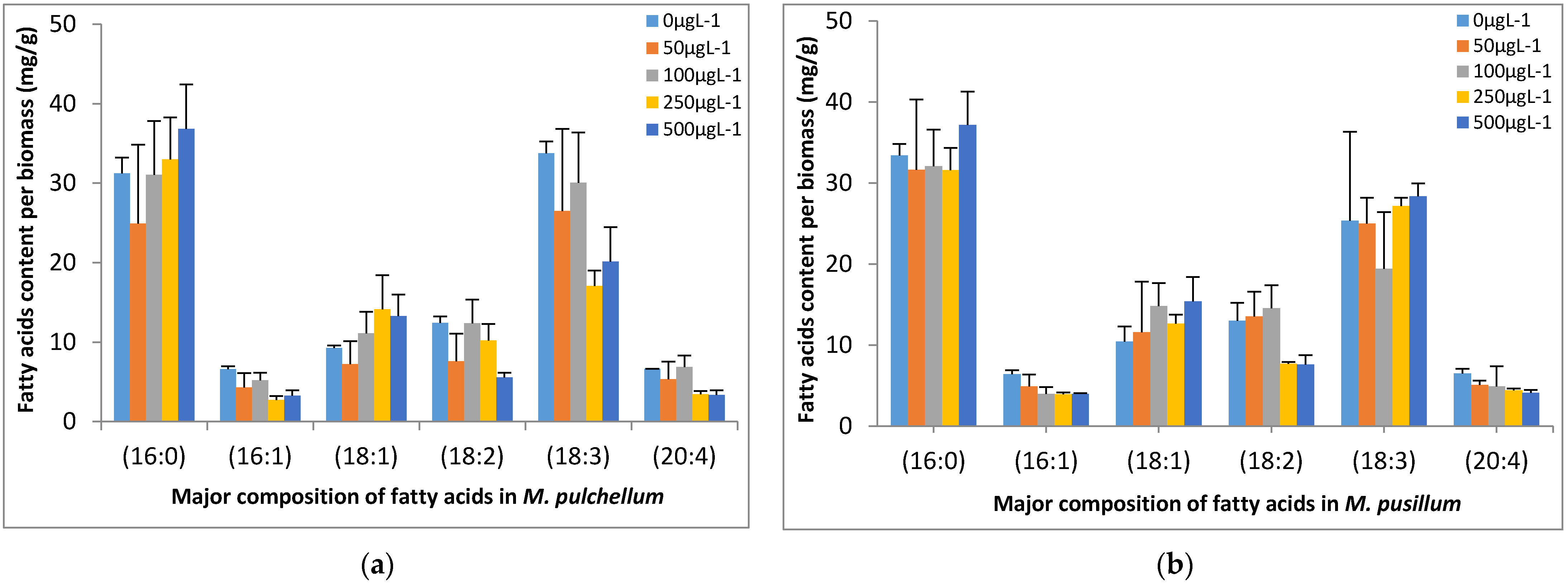
The fatty acids were analyzed by GC, and the major fatty acid composition of both tested microalgae species included palmitic acid (16:0; hexadecanoic acid), palmitoleic acid (C16:1; ω-7), vaccenic acid (C18:1; ω-7), linoleic acid (C18:2; ω-6), α-linoleic acid (C18:3; ω-3) and arqachidonic acid (C20:4; ω-6) as shown in Figure 8. Higher levels of palmitic acid and α-linoleic acid were observed in both microalgae cultures at all the tested concentrations. Higher Cr(VI) concentrations in M. pulchellum cultures resulted in slightly higher content of palmitic acid among the major fatty acids. However, all other fatty acids showed a slightly decreasing tendency at higher concentrations. The increasing Cr(VI) concentrations in M. pusillum cultures showed a slightly higher content of vaccenic acid and α-linoleic acid. However, a slightly decreasing tendency was observed for palmitoleic acid and arachidonic acid. Furthermore, higher tested Cr(VI) concentrations (250 µg L−1 and 500 µg L−1) significantly (p < 0.05) reduced the linoleic acid content.
4. Discussion
Chromium exists in the environment as trivalent, Cr(III) and hexavalent form, Cr(VI), where Cr(VI) is the highly toxic, carcinogenic and mutagenic [29,30]. The discharge of chromium from anthropogenic sources such as household, industry, transport, mining and agriculture increases its concentration several times above normal levels [31]. The aquatic ecosystems are seriously affected by Cr(VI) toxicity, which depends on its physiochemical, oxidative and structural properties [32]. The Cr(VI) constituents are generally soluble and mobile in the environment [33]. They can easily pass cell membrane due to the structural similarity to inorganic anions which makes Cr(VI) an alternative substrate in the sulfate transport system [34,35]. The cytotoxic effects of Cr(VI) on living organisms including plants, animals and humans are well reported, and they are also a source of a variety of human cancers [32,36].
Heavy metal toxicity and the effect of heavy metals on microalgae have been extensively reported. However, there is a limited or no literature available on the heavy metal toxicity of economically promising freshwater eukaryotic microalgae species M. pulchellum and M. pusillum. The freshwater eukaryotic non-model microalgae species M. pulchellum and M. pusillum were chosen for this study due to their economic potential as they can grow at low light intensity as well as in CO2-deficient conditions [37]. They are diverse green phytoplankton species which occasionally inhabit freshwater lakes. Furthermore, the genetic transformation of M. pulchellum (formerly D. pulchellum) for higher erythropoietin protein accumulation was reported by our group [38], which shows the broad scope of this microalgae species. During this study, the toxicity of hexavalent chromium (0–500 µg L−1) on M. pulchellum and M. pusillum was investigated by comparing variations in cell density, modulated fluorescence yield, relative photosynthetic inhibition and lipid accumulation. Both microalgae species exhibited significant sensitivities to the tested Cr(VI) concentrations as indicated by the fluorescence kinetics and lipid content.
Both microalgae species showed decreased growth at any Cr(VI) tested concentration when compared to the chromium free control cultivation. This effect has been previously attributed to the PSII inhibition via electron transport inhibition between QA_ and QB/QB_ by the hexavalent chromium ion [39]. Additionally, higher tested concentrations showed a decline in growth after day 7, likely via the well elucidated genotoxic route, where chromium ions form complexes with nucleic acids, causing strand breaks and the formation of mutagenic Cr-DNA fragments. Similarly, significant increases in minimal (F0) and maximal (Fm) fluorescence values were observed till 100 μg L−1; however, a decline in F0 and Fm values was observed at concentrations exceeding 100 μg L−1. Furthermore, 50 μg L−1 chromium treatment showed higher maximal fluorescence in M. pulchellum as compared to M. pusillum, which shows higher sensitivity of M. pusillum to chromium even at low concentration. All tested Cr(VI) concentrations showed a significant decline in quantum yield and photosynthetic inhibition as compared to the control. A maximum photosynthetic inhibition of up to 67% was observed in M. pulchellum at 500 μg L−1 whereas M. pusillum showed a maximum photosynthetic inhibition of up to 66%. This shows that Cr(VI) has a negative effect on normal growth of both microalgae species by interrupting photosynthesis.
Interestingly, both tested microalgae species showed quit similar trends for growth and fluorescence yields; this could be because both microalgae are freshwater eukaryotic microalgae species and belong to a same taxon—Chlorophyta—and thus showed similar Cr(VI) uptake and a similar PSII structure. Contrary to the present study, a 50% inhibition in growth was observed by Hörcsik et al. [40] when analyzing Cr(VI) toxicity using chlorophyll composition of Auxenochlorella pyrenoidosa (Chlorella pyrenoidosa) for 72 h in media supplemented with 2 mg L−1 of Cr(VI) [40]. Similarly, in another study, a very minor amount of Cr(VI), 5 µmol L−1, showed up to 40% inhibition in the maximal quantum yield of PSII of Chlorella vulgaris when treated for 96 h [12]. The variation in the present study could be due to different microalgae species and different experimental conditions.
The lipid accumulation results were quite interesting; both tested microalgae showed increases in lipid content. A lipid content of up to 36% was observed in M. pulchellum at the maximum tested Cr(VI) concentration in this study (500 µg L−1), which was approximately 10 times higher than the control. Whereas a maximum lipid content of up to 30% was observed in M. pusillum. The fatty acids composition analysis showed higher levels of polyunsaturated fatty acids (hexadecenoic acid) and saturated fatty acids (ω3, ω6, and ω7). However, chromium exposure significantly affected the saturated fatty acids content. In this study, linoleic acid (C18:2; ω6) and α-linoleic (C18:3; ω3) were among the mostly affected fatty acids by Cr(VI). This agrees with the previously reported studies of Barsanti et al. [41] and Rochhetta et al. [42], which states that chloroplast structure related lipids such as linoleic acid and α-linoleic are mostly affected by chromium. Furthermore, Cr(VI) treated cultures showed no significant differences for the non-photosynthetic structure related fatty acids such as arachidonic acid (C20:4; ω6), which agrees with Rochhetta et al. [42]. This suggests that chloroplasts would be the main target organelle of Cr(VI) toxicity in M. pulchellum and M. pusillum.
Despite significant decreases in observed saturated fatty acids content at higher Cr(VI) concentration (especially ω3 and ω6), total lipid content showed a significant increase. This could be a microalgal defense mechanism to counteract oxidative damage [42]. Both microalgae species showed higher lipid accumulation at higher tested Cr(VI) concentration than the control; this shows that the addition of Cr(VI) could significantly enhance the lipid accumulation in M. pulchellum and M. pusillum.
The results of this study indicate that Cr(VI) can affect total lipids and fatty acids content, especially affecting the fatty acids related to photosynthetic activity. Changes in fatty acids composition in the treated cells could be due to their defense mechanism to reduce cellular damage caused by Cr(VI) and its route outlined above. Additionally, intracellular Cr(VI) reduction consumes intracellular antioxidants which could induce synthesis of simple and poly-unsaturated fatty acids as a defense mechanism. However, further analytical and biochemical analyses are necessary to assist the findings. The present study could aid in aquaculture industry, in maintenance of microalgae stock cultures and in estimating the possible side effects of using hexavalent chromium in microalgae cultures. It can further aid in the design and construction of biomarkers using eukaryotic freshwater microalgae species.
Author Contributions
Conceptualization, K.M.I.B.; methodology, K.M.I.B. and H.-J.L.; software, K.M.I.B. and H.-J.L.; validation, K.M.I.B., S.M. and A.J.; formal analysis, K.M.I.B. and H.-J.L.; resources, M.-G.C.; data curation, K.M.I.B. and H.-J.L.; writing—original draft preparation, K.M.I.B. and S.M.; writing—review and editing, S.M., A.J. and M.-G.C.; visualization, K.M.I.B. and H.-J.L.; supervision, M.-G.C.; project administration, M.-G.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Busan Metropolitan City, BB21+ 2020 grant. The APC was funded by BB21+ 2020 grant.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data generated or analyzed during this study are included in this article, and the primary data could be provided by the corresponding author upon request.
Acknowledgments
This work was supported by the BB21+ Project in 2020; authors are thankful for their support.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses or interpretation of data; in the writing of the manuscript or in the decision to publish the results.
References
- Bashir, K.M.I.; Mansoor, S.; Kim, N.-R.; Grohmann, F.R.; Shah, A.A.; Cho, M.-G. Effect of organic carbon sources and environmental factors on cell growth and lipid content of Pavlova lutheri. Ann. Microbiol. 2019, 69, 353–368. [Google Scholar] [CrossRef]
- Lu, C.M.; Chau, C.W.; Zhang, J.H. Acute toxicity of excess mercury on the photosynthetic performance of cyanobacterium, Spirulina platensis—Assessment by chlorophyll fluorescence analysis. Chemosphere 2000, 41, 191–196. [Google Scholar] [CrossRef]
- Juneau, P.; Dewez, D.; Matsui, S.; Kim, S.G.; Popovic, R. Evaluation of different algal species sensitivity to mercury and metolachlor by PAM-fluorometry. Chemosphere 2001, 45, 589–598. [Google Scholar] [CrossRef]
- Pena-Vazquez, E.; Perez-Conde, C. Development of a microalgal PAM test method for Cu (II) in water: Comparison of using spectrofluorometry. Ecotoxicology 2010, 19, 1059–1065. [Google Scholar] [CrossRef] [PubMed]
- Bashir, K.M.I.; Cho, M.-G. The effect of kanamycin and tetracycline on growth and photosynthetic activity of two Chlorophyte algae. BioMed Res. Int. 2016, 2016, 5656304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miazek, K.; Iwanek, W.; Remacle, C.; Richel, A.; Goffin, D. Effect of metals, metalloids and metallic nanoparticles on microalgae growth and industrial product biosynthesis: A review. Int. J. Mol. Sci. 2015, 16, 23929–23969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Rodas, V.; Agrelo, M.; Carrillo, E.; Ferrero, L.; Larrauri, A.; Martín-Otero, L.; Costas, E. Resistance of microalgae to modern water contaminants as the result of rare spontaneous mutations. Eur. J. Phycol. 2001, 36, 179–190. [Google Scholar] [CrossRef]
- Hawrot-Paw, M.; Koniuszy, A.; Gałczyńska, M. Sustainable production of Monoraphidium microalgae biomass as a source of bioenergy. Energies 2020, 13, 5975. [Google Scholar] [CrossRef]
- Bengtson Nash, S.M.; Quayle, P.A.; Schreiber, U.; Müller, J.F. The selection of a model microalgal species as biomaterial for a novel aquatic phytotoxicity assay. Aquat. Toxicol. 2005, 72, 315–326. [Google Scholar] [CrossRef]
- Herlory, O.; Bonzom, J.M.; Gilbin, R. Sensitivity evaluation of green alga Chlamydomonas reinhardtii to uranium by pulse amplitude modulated (PAM) fluorometry. Aquat. Toxicol. 2013, 140–141, 288–294. [Google Scholar] [CrossRef]
- Kumar, K.S.; Dahms, H.-U.; Lee, J.-S.; Kim, H.C.; Lee, W.C.; Shin, K.-H. Algal photosynthetic responses to toxic metals and herbicides assessed by chlorophyll a fluorescence. Ecotoxicol. Environ. Saf. 2014, 104, 51–71. [Google Scholar] [CrossRef]
- Ouyang, H.L.; Kong, X.Z.; He, W.; Qin, N.; He, Q.S.; Wang, Y.; Wang, R.; Xu, F.L. Effects of five heavy metals at sub-lethal concentrations on the growth and photosynthesis of Chlorella vulgaris. Chin. Sci. Bull. 2012, 57, 3363–3370. [Google Scholar] [CrossRef] [Green Version]
- Fu, F.; Wang, Q. Removal of heavy metal ions from wastewaters: A review. J. Environ. Manag. 2011, 92, 407–418. [Google Scholar] [CrossRef] [PubMed]
- Siddiquee, S.; Rovina, K.; Azad, S.A.; Naher, L.; Suryani, S.; Chaikaew, P. Heavy metal contaminants removal from wastewater using the potential filamentous fungi biomass: A review. J. Microb. Biochem. Technol. 2015, 7, 384–393. [Google Scholar] [CrossRef]
- Ng, C.C.; Motior Rahman, M.; Boyce, A.N.; Abas, M.R. Heavy metals phyto-assessment in commonly grown vegetables: Water spinach (I. aquatica) and okra (A. esculentus). SpringerPlus 2016, 5, 469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vignati, D.A.L.; Dominik, J.; Beye, M.L.; Pettine, M.; Ferrari, B.J.D. Chromium (VI) is more toxic than chromium (III) to freshwater algae: A paradigm to revise? Ecotoxicol. Environ. Saf. 2010, 73, 743–749. [Google Scholar] [CrossRef]
- WHO. Chromium in Drinking-Water Background Document for Development of WHO Guidelines for Drinking-Water Quality; World Health Organization: Geneva, Switzerland, 2003; Available online: https://www.who.int/water_sanitation_health/dwq/chemicals/chromium.pdf (accessed on 2 March 2021).
- Yang, L.; Chen, P. Biosorption of hexavalent chromium onto raw and chemically modified Sargassum sp. Bioresour. Technol. 2008, 99, 297–307. [Google Scholar] [CrossRef] [PubMed]
- An, M.; Gao, L.; Zhao, W.; Chen, W.; Li, M. Effects of nitrogen forms and supply mode on lipid production of microalga Scenedesmus obliquus. Energies 2020, 13, 697. [Google Scholar] [CrossRef] [Green Version]
- Ismail, M.; Phang, S.-M.; Tong, S.-L.; Brown, M.T. A modified toxicity testing method using tropical marine microalgae. Environ. Monit. Assess 2002, 75, 145–154. [Google Scholar] [CrossRef]
- Baumann, H.A.; Morrison, L.; Stenge, D.B. Metal accumulation and toxicity measured by PAM-chlorophyll fluorescence in seven species of marine macroalgae. Ecotoxicol. Environ. Saf. 2009, 72, 1063–1075. [Google Scholar] [CrossRef] [PubMed]
- Bashir, K.M.I.; Lee, J.-H.; Petermann, M.J.; Shah, A.A.; Jeong, S.-J.; Kim, M.-S.; Park, N.G.; Cho, M.-G. Estimation of antibacterial properties of Chlorophyta, Rhodophyta and Haptophyta microalgae species. Microbiol. Biotechnol. Lett. 2018, 46, 225–233. [Google Scholar] [CrossRef]
- Watanabe, M.M.; Kawachi, M.; Hiroki, M.; Kasai, F. NIES Collection List of Strains. In Microalgae and Protozoa. Microbial Culture Collections, 6th ed.; National Institute for Environmental Studies: Tsukuba, Japan, 2000; p. 159. [Google Scholar]
- Schreiber, U. Detection of rapid induction kinetics with a new type of high-frequency modulated chlorophyll fluorometer. Photosynth. Res. 1986, 9, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Genty, B.; Briantais, J.-M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ichihara, K.; Fukubayashi, Y. Preparation of fatty acid methyl esters for gas-liquid chromatography. J. Lipid Res. 2010, 51, 635–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balan, R.; Suraishkumar, G.K. Simultaneous increases in specific growth rate and specific lipid content of Chlorella vulgaris through UV-induced reactive species. Biotechnol. Prog. 2014, 30, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Rai, R.; Eary, L.E.; Zachara, J.M. Environmental chemistry of chromium. Sci. Total Environ. 1989, 86, 15–23. [Google Scholar] [CrossRef]
- Gomez, V.; Callao, M.P. Chromium determination and speciation since 2000. TRAC-Trend Anal. Chem. 2006, 25, 1006–1015. [Google Scholar] [CrossRef]
- Nogales, B.; Lanfranconi, M.P.; Pina-Villalonga, J.M.; Bosch, R. Anthropogenic perturbations in marine microbial communities. FEMS Microbiol. Rev. 2011, 35, 275–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagchi, D.; Stohs, S.J.; Downs, B.W.; Bagchi, M.; Preuss, H.G. Cytotoxicity and oxidative mechanisms of different forms of chromium. Toxicology 2002, 180, 5–22. [Google Scholar] [CrossRef]
- Ertani, A.; Mietto, A.; Borin, M.; Nardi, S. Chromium in agricultural soils and crops: A review. Water Air Soil Pollut. 2017, 228, 190. [Google Scholar] [CrossRef]
- Cieslak-Golonka, M. Toxic and mutagenic effects of chromium (VI). In Polyhedron Rep; Elsevier Science Ltd: Amsterdam, The Netherlands, 1996; Volume 61, p. 3667. [Google Scholar]
- Haglund, K. The use of algae in aquatic toxicity assessment. In Progress in Phycological Research; Biopress: Bristol, UK, 1997; Volume 12, pp. 181–212. [Google Scholar]
- Vajpayee, P.; Rai, U.N.; Ali, M.B.; Tripathi, V.; Yadav, V.; Sinha, S.; Singh, S.N. Chromium-induced physiologic changes in Vallisneria spiralis L. and its role in phytoremediation of tannery effluent. Bull. Environ. Contam. Toxicol. 2001, 67, 246–256. [Google Scholar] [CrossRef] [PubMed]
- Irfanullah, H.M.D.; Moss, B. Ecology of Dictyosphaerium pulchellum Wood (Chlorophyta, Chlorococcales) in a shallow, acid, forest lake. Aquat. Ecol. 2006, 40, 1–12. [Google Scholar] [CrossRef]
- Bashir, K.M.I.; Kim, M.-S.; Stahl, U.; Cho, M.-G. Agrobacterium-mediated genetic transformation of Dictyosphaerium pulchellum for the expression of erythropoietin. J. Appl. Phycol. 2018, 30, 3503–3518. [Google Scholar] [CrossRef]
- Pan, X.; Chen, X.; Zhang, D.; Wang, J.; Deng, C.; Mu, G.; Zhu, H. Effect of chromium(VI) on photosystem II activity and heterogeneity of Synechocystis sp. (Cyanophyta): Studied with in vivo chlorophyll fluorescence tests. J. Phycol. 2009, 45, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Hörcsik, Z.; Oláh, V.; Balogh, A.; Mészáros, I.; Simon, L.; Lakatos, G. Effect of Chromium(VI) on growth, element, and photosynthetic pigment composition of Chlorella pyrenoidosa. Acta. Biol. Szeged. 2006, 50, 19–23. [Google Scholar]
- Barsanti, L.; Bastianini, A.; Passarelli, V.; Tredici, M.R.; Gualtieri, P. Fatty acid content in wild type and WZSL mutant of Euglena gracilis. J. Appl. Phycol. 2000, 12, 515–520. [Google Scholar] [CrossRef]
- Rocchetta, I.; Mazzuca, M.; Conforti, V.; Ruiz, L.; Balzaretti, V.; Ríos de Molina, M.d.C. Effect of chromium on the fatty acid composition of two strains of Euglena gracilis. Environ. Pollut. 2006, 141, 353–358. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effect of Cr(VI) on cell density of (a) M. pulchellum and (b) M. pusillum. X-axis represents culturing days and the cell densities (×106 mL−1) are shown along the y-axis. Data are means ± SE, n = 3.
Figure 1.
Effect of Cr(VI) on cell density of (a) M. pulchellum and (b) M. pusillum. X-axis represents culturing days and the cell densities (×106 mL−1) are shown along the y-axis. Data are means ± SE, n = 3.

Figure 2.
Effect of Cr(VI) on optical density of (a) M. pulchellum and (b) M. pusillum. X-axis represents culturing days and the optical density values at 750 nm (OD750) are shown along the y-axis. Data are means ± SE, n = 3.
Figure 2.
Effect of Cr(VI) on optical density of (a) M. pulchellum and (b) M. pusillum. X-axis represents culturing days and the optical density values at 750 nm (OD750) are shown along the y-axis. Data are means ± SE, n = 3.

Figure 3.
Effect of Cr(VI) on minimal fluorescence of (a) M. pulchellum and (b) M. pusillum. X-axis represents the culturing days, and the minimum fluorescence (F0) values are shown along the y-axis. Data are means ± SE, n = 3.
Figure 3.
Effect of Cr(VI) on minimal fluorescence of (a) M. pulchellum and (b) M. pusillum. X-axis represents the culturing days, and the minimum fluorescence (F0) values are shown along the y-axis. Data are means ± SE, n = 3.

Figure 4.
Effect of Cr(VI) on maximal fluorescence of (a) M. pulchellum and (b) M. pusillum. X-axis represents the culturing days, and the maximal fluorescence (Fm) values are shown along the y-axis. Data are means ± SE, n = 3.
Figure 4.
Effect of Cr(VI) on maximal fluorescence of (a) M. pulchellum and (b) M. pusillum. X-axis represents the culturing days, and the maximal fluorescence (Fm) values are shown along the y-axis. Data are means ± SE, n = 3.
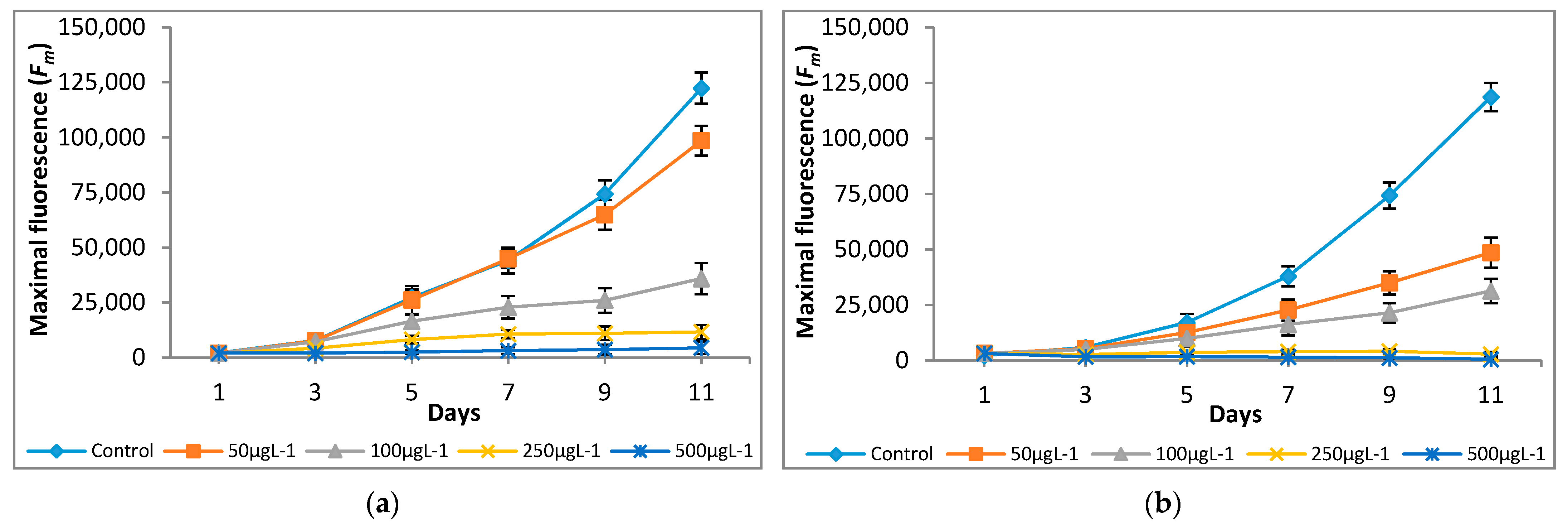
Figure 5.
Effect of Cr(VI) on quantum yield of (a) M. pulchellum and (b) M. pusillum. X-axis represents the culturing days, and the quantum yield (Y) values are shown along the y-axis. Data are means ± SE, n = 3.
Figure 5.
Effect of Cr(VI) on quantum yield of (a) M. pulchellum and (b) M. pusillum. X-axis represents the culturing days, and the quantum yield (Y) values are shown along the y-axis. Data are means ± SE, n = 3.
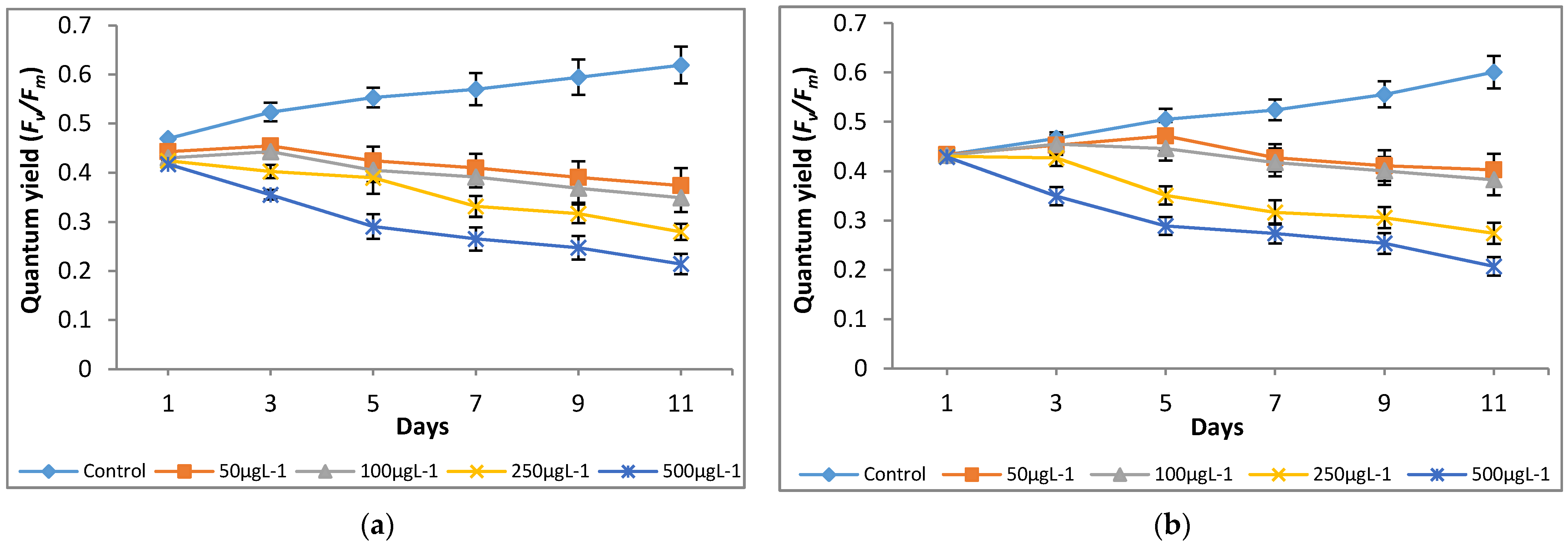
Figure 6.
Effect of Cr(VI) on photosynthetic inhibition of (a) M. pulchellum and (b) M. pusillum. X-axis represents the culturing days, and the inhibition (%) values are shown along the y-axis. Data are means ± SE, n = 3.
Figure 6.
Effect of Cr(VI) on photosynthetic inhibition of (a) M. pulchellum and (b) M. pusillum. X-axis represents the culturing days, and the inhibition (%) values are shown along the y-axis. Data are means ± SE, n = 3.

Figure 7.
Effect of Cr(VI) on lipid content in M. pulchellum (■) and M. pusillum (■). X-axis represents the chromium concentrations (µg L−1), and the lipid content (%, g of lipid/g of dry weight biomass) is shown along the y-axis. Data are means ± SE, n = 3.
Figure 7.
Effect of Cr(VI) on lipid content in M. pulchellum (■) and M. pusillum (■). X-axis represents the chromium concentrations (µg L−1), and the lipid content (%, g of lipid/g of dry weight biomass) is shown along the y-axis. Data are means ± SE, n = 3.
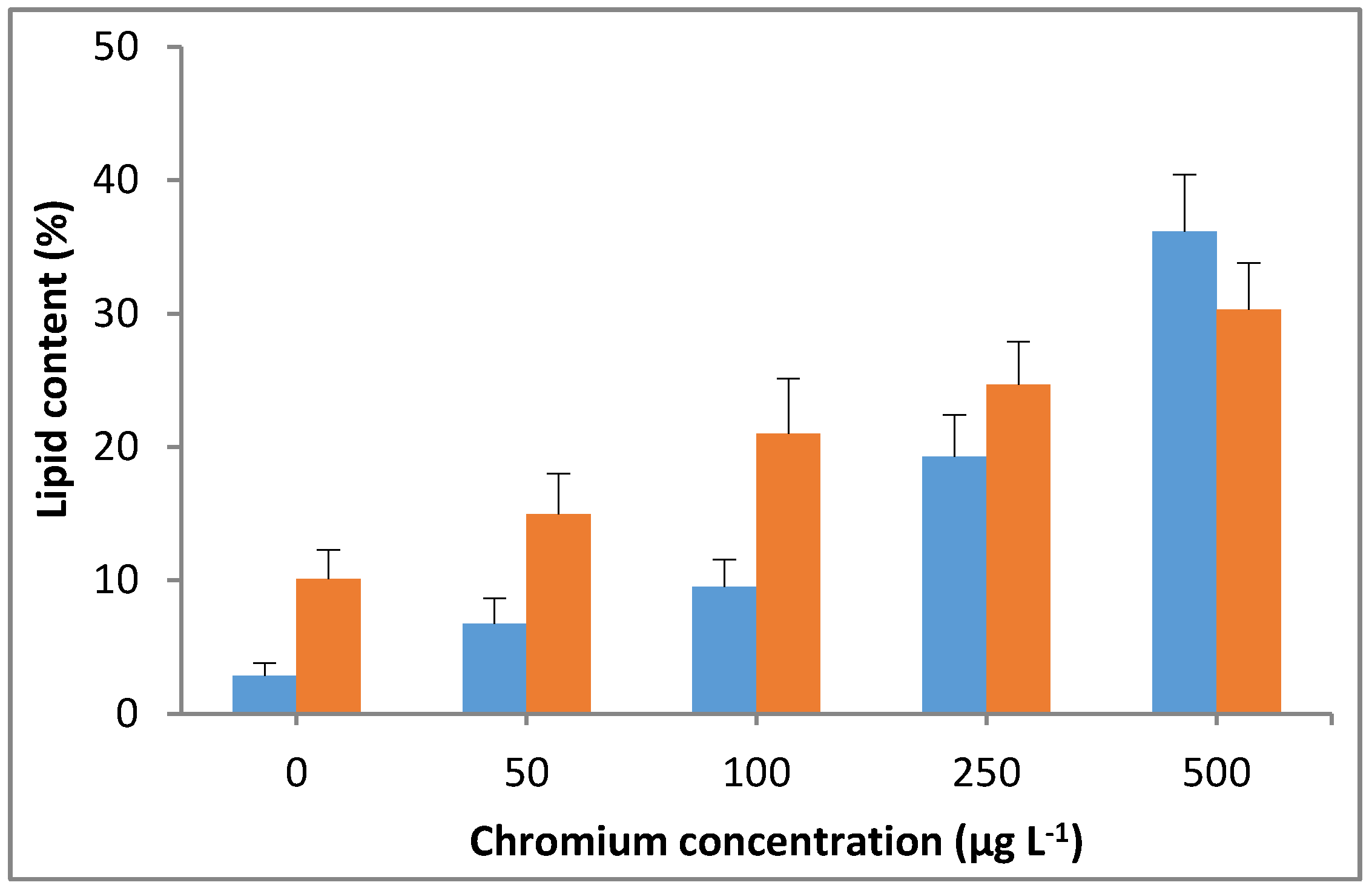
Figure 8.
Effect of Cr(VI) on fatty acids composition in (a) M. pulchellum and (b) M. pusillum. X-axis represents the fatty acids, and fatty acid contents per biomass (mg/g) are shown along the y-axis. Data are means ± SE, n = 3.
Figure 8.
Effect of Cr(VI) on fatty acids composition in (a) M. pulchellum and (b) M. pusillum. X-axis represents the fatty acids, and fatty acid contents per biomass (mg/g) are shown along the y-axis. Data are means ± SE, n = 3.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Effect of Cr(VI) on growth rates (cells/day).
| Species | Cr(VI) Concentration | ||||
|---|---|---|---|---|---|
| 0 μg L−1 | 50 μg L−1 | 100 μg L−1 | 250 μg L−1 | 500 μg L−1 | |
| M. pulchellum | μ = 0.1912 | μ = 0.1605 | 0.3688 × 106 * | 0.2139 × 106 * | 0.1311 × 106 * |
| M. pusillum | 0.6007 × 106 * | 0.4706 × 106 * | 0.3937 × 106 * | 0.1678 × 106 * | 0.1110 × 106 * |
* indicates linear growth.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bashir, K.M.I.; Lee, H.-J.; Mansoor, S.; Jahn, A.; Cho, M.-G. The Effect of Chromium on Photosynthesis and Lipid Accumulation in Two Chlorophyte Microalgae. Energies 2021, 14, 2260. https://doi.org/10.3390/en14082260
AMA Style
Bashir KMI, Lee H-J, Mansoor S, Jahn A, Cho M-G. The Effect of Chromium on Photosynthesis and Lipid Accumulation in Two Chlorophyte Microalgae. Energies. 2021; 14(8):2260. https://doi.org/10.3390/en14082260
Chicago/Turabian StyleBashir, Khawaja Muhammad Imran, Hyeon-Jun Lee, Sana Mansoor, Alexander Jahn, and Man-Gi Cho. 2021. "The Effect of Chromium on Photosynthesis and Lipid Accumulation in Two Chlorophyte Microalgae" Energies 14, no. 8: 2260. https://doi.org/10.3390/en14082260
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.