Ethanol Production from Hydrolyzed Kraft Pulp by Mono- and Co-Cultures of Yeasts: The Challenge of C6 and C5 Sugars Consumption


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Kraft Pulp
2.2. Enzymatic Hydrolysis
2.3. Microorganisms
2.4. Culture Media
2.5. Pre-Inocula and Inocula
2.6. Fermentation
2.6.1. Erlenmeyer Flask Assays
2.6.2. Bioreactor Assays
2.7. Analytical Methods
2.8. Calculations
3. Results
3.1. Enzymatic Hydrolysis
3.2. Erlenmeyer Flask Mono-Culture Assays
3.3. Erlenmeyer Flask Co-Culture Assays
3.4. Bioreactor Assays
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- REN21. Renewables 2018 Global Status Report; REN21 Secretariat: Paris, France, 2018. [Google Scholar]
- Liew, W.H.; Hassim, M.H.; Ng, D.K.S. Review of evolution, technology and sustainability assessments of biofuel production. J. Clean. Prod. 2014, 71, 11–29. [Google Scholar] [CrossRef]
- Berni, M.D.; Dorileo, I.L.; Prado, J.M.; Forster-Carneiro, T.; Meireles, M.A.A. Advances in biofuel production. In Biofuels Production; Babu, V., Thapliyal, A., Patel, G.K., Eds.; Scrivener Publishing: Boston, MA, USA, 2014. [Google Scholar]
- Dutta, K.; Daverey, A.; Lin, J.-G. Evolution retrospective for alternative fuels: First to fourth generation. Renew. Energy 2014, 69, 114–122. [Google Scholar] [CrossRef]
- Manochio, C.; Andrade, B.R.; Rodriguez, R.P.; Moraes, B.S. Ethanol from biomass: A comparative overview. Renew. Sustain. Energy Rev. 2017, 80, 743–755. [Google Scholar] [CrossRef]
- Zabed, H.; Sahu, J.N.; Suely, A.; Boyce, A.N.; Faruq, G. Bioethanol production from renewable sources: Current perspectives and technological progress. Renew. Sustain. Energy Rev. 2017, 71, 475–501. [Google Scholar] [CrossRef]
- Zabed, H.; Sahu, J.N.; Boyce, A.N.; Faruq, G. Fuel ethanol production from lignocellulosic biomass: An overview on feedstocks and technological approaches. Renew. Sustain. Energy Rev. 2016, 66, 751–774. [Google Scholar] [CrossRef]
- Yamakawa, C.K.; Qin, F.; Mussatto, S.I. Advances and opportunities in biomass conversion technologies and biorefineries for the development of a bio-based economy. Biomass Bioenergy 2018, 119, 54–60. [Google Scholar] [CrossRef]
- Bhutto, A.W.; Qureshi, K.; Harijan, K.; Abro, R.; Abbas, T.; Bazmi, A.A.; Karim, S.; Yu, G. Insight into progress in pre-treatment of lignocellulosic biomass. Energy 2017, 122, 724–745. [Google Scholar] [CrossRef]
- Stoklosa, R.J.; Hodge, D.B. Integration of (hemi)-cellulosic biofuels technologies with chemical pulp production. In Biorefineries: Integrated Biochemical Processes for Liquid Biofuels; Qureshi, N., Hodge, D., Vertès, A.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2014. [Google Scholar]
- Branco, R.; Serafim, L.S.; Xavier, A.M.R.B. Second generation bioethanol production: On the use of pulp and paper industry wastes as feedstock. Fermentation 2018, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Jönsson, L.J.; Martín, C. Pretreatment of lignocellulose: Formation of inhibitory by-products and strategies for minimizing their effects. Bioresour. Technol. 2016, 199, 103–112. [Google Scholar] [CrossRef] [Green Version]
- FAO. Yearbook of Forest Products 2015; Food and Agriculture Organization of the United Nations: Rome, Italy, 2017. [Google Scholar]
- Lachenal, D. Kraft pulping. In Lignocellulosic Fibers and Wood Handbook; Belgacem, M.N., Pizzi, A., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2016. [Google Scholar]
- Monrroy, M.; García, J.R.; Mendonça, R.T.; Baeza, J.; Freer, J. Kraft pulping of Eucalyptus globulus as a pretreatment for bioethanol production by simultaneous saccharification and fermentation. J. Chil. Chem. Soc. 2012, 57, 1113–1117. [Google Scholar] [CrossRef] [Green Version]
- Buzała, K.; Przybysz, P.; Rosicka-Kaczmarek, J.; Kalinowska, H. Production of glucose-rich enzymatic hydrolysates from cellulosic pulps. Cellulose 2015, 22, 663–674. [Google Scholar] [CrossRef] [Green Version]
- Solihat, N.N.; Fajriutami, T.; Adi, D.T.N.; Fatriasari, W.; Hermiati, E. Reducing sugar production of sweet sorghum bagasse kraft pulp. In Proceedings of the AIP Conference Proceedings, Tangerang Banten, Indonesia, 3–5 October 2016. [Google Scholar]
- Buzała, K.P.; Kalinowska, H.; Przybysz, P.; Małachowska, E. Conversion of various types of lignocellulosic biomass to fermentable sugars using kraft pulping and enzymatic hydrolysis. Wood Sci. Technol. 2017, 51, 873–885. [Google Scholar] [CrossRef] [Green Version]
- Troncoso, E.; Castillo, R.; Valenzuela, R.; Reyes, P.; Freer, J.; Norambuena, M.; RodríGuez, J.; Parra, C. Chemical and microstructural changes in Eucalyptus globulus fibers subjected to four different pretreatments and their influence on the enzymatic hydrolysis. J. Chil. Chem. Soc. 2017, 62, 3442–3446. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; He, J.; Min, D.; Lai, C.; Yong, Q. Understanding the nonproductive enzyme adsorption and physicochemical properties of residual lignins in moso bamboo pretreated with sulfuric acid and Kraft pulping. Appl. Biochem. Biotech. 2016, 180, 1508–1523. [Google Scholar] [CrossRef]
- Novozhilov, E.V.; Sinel’nikov, I.G.; Aksenov, A.S.; Chukhchin, D.G.; Tyshkunova, I.V.; Rozhkova, A.M.; Osipov, D.O.; Zorov, I.N.; Sinitsyn, A.P. Biocatalytic conversion of kraft pulp using cellulase complex of Penicillium verruculosum. Catal. Ind. 2016, 8, 95–100. [Google Scholar] [CrossRef]
- Buzała, K.P.; Kalinowska, H.; Małachowska, E.; Przybysz, P. The utility of selected kraft hardwood and softwood pulps for fuel ethanol production. Ind. Crops Prod. 2017, 108, 824–830. [Google Scholar] [CrossRef]
- Huang, C.; He, J.; Chang, H.; Jameel, H.; Yong, Q. Coproduction of ethanol and lignosulfonate from moso bamboo residues by fermentation and sulfomethylation. Waste Biomass Valorization 2017, 8, 965–974. [Google Scholar] [CrossRef]
- Ko, C.-H.; Wang, Y.-N.; Chang, F.-C.; Chen, J.-J.; Chen, W.-H.; Hwang, W.-S. Potentials of lignocellulosic bioethanols produced from hardwood in Taiwan. Energy 2012, 44, 329–334. [Google Scholar] [CrossRef]
- Wistara, N.J.; Pelawi, R.; Fatriasari, W. The effect of lignin content and freeness of pulp on the bioethanol productivity of Jabon wood. Waste Biomass Valorization 2016, 7, 1141–1146. [Google Scholar] [CrossRef]
- Edgardo, A.; Carolina, P.; Manuel, R.; Juanita, F.; Baeza, J. Selection of thermotolerant yeast strains Saccharomyces cerevisiae for bioethanol production. Enzyme Microb. Technol. 2008, 43, 120–123. [Google Scholar] [CrossRef]
- Bauer, N.A.; Gibbons, W.R. Saccharification versus simultaneous saccharification and fermentation of kraft pulp. Int. J. Agric. Biol. Eng. 2012, 5, 48–55. [Google Scholar]
- Amoah, J.; Ishizue, N.; Ishizaki, M.; Yasuda, M.; Takahashi, K.; Ninomiya, K.; Yamada, R.; Kondo, A.; Ogino, C. Development and evaluation of consolidated bioprocessing yeast for ethanol production from ionic liquid-pretreated bagasse. Bioresour. Technol. 2017, 245, 1413–1420. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, L.V.; de Barros Grassi, M.C.; Gallardo, J.C.M.; Pirolla, R.A.S.; Calderón, L.L.; de Carvalho-Netto, O.V.; Parreiras, L.S.; Camargo, E.L.O.; Drezza, A.L.; Missawa, S.K.; et al. Second-generation ethanol: The need is becoming a reality. Ind. Biotechnol. 2016, 12, 40–57. [Google Scholar] [CrossRef]
- Henriques, T.; Pereira, S.; Serafim, L.; Xavier, A. Two-stage aeration fermentation strategy to improve bioethanol production by Scheffersomyces stipitis. Fermentation 2018, 4, 97. [Google Scholar] [CrossRef] [Green Version]
- Pereira, S.R.; Ivanuša, Š.; Evtuguin, D.V.; Serafim, L.S.; Xavier, A.M.R.B. Biological treatment of eucalypt spent sulphite liquors: A way to boost the production of second generation bioethanol. Bioresour. Technol. 2012, 103, 131–135. [Google Scholar] [CrossRef]
- Pereira, S.R.; Sànchez i Nogué, V.; Frazão, C.J.R.; Serafim, L.S.; Gorwa-Grauslund, M.F.; Xavier, A.M.R.B. Adaptation of Scheffersomyces stipitis to hardwood spent sulfite liquor by evolutionary engineering. Biotechnol. Biofuels 2015, 8, 50. [Google Scholar] [CrossRef] [Green Version]
- Mendes, J.A.S.; Xavier, A.M.R.B.; Evtuguin, D.V.; Lopes, L.P.C. Integrated utilization of grape skins from white grape pomaces. Ind. Crops Prod. 2013, 49, 286–291. [Google Scholar] [CrossRef]
- Jiang, C.-X.; He, Y.-C.; Chong, G.-G.; Di, J.-H.; Tang, Y.-J.; Ma, C.-L. Enzymatic in situ saccharification of sugarcane bagasse pretreated with low loading of alkalic salts Na2SO3/Na3PO4 by autoclaving. J. Biotechnol. 2017, 259, 73–82. [Google Scholar] [CrossRef]
- Chandel, A.K.; Antunes, F.A.; Silva, M.B.; da Silva, S. Unraveling the structure of sugarcane bagasse after soaking in concentrated aqueous ammonia (SCAA) and ethanol production by Scheffersomyces (Pichia) stipitis. Biotechnol. Biofuels 2013, 6, 102. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, M.C.; Ferro, M.D.; Paulino, A.F.C.; Chaves, H.T.; Evtuguin, D.V.; Xavier, A.M.R.B. Comparative study on hydrolysis and bioethanol production from cardoon and rockrose pretreated by dilute acid hydrolysis. Ind. Crops Prod. 2018, 111, 633–641. [Google Scholar] [CrossRef]
- Chen, Y. Development and application of co-culture for ethanol production by co-fermentation of glucose and xylose: A systematic review. J. Ind. Microbiol. Biotechnol. 2011, 38, 581–597. [Google Scholar] [CrossRef] [PubMed]
- Rice, E.W.; Baird, R.B.; Eaton, A.D. Standard Methods for Examination of Water and Wastewater, 23rd ed.; AWWA: City and County of Denve, CO, USA, 2017. [Google Scholar]
- Kang, Q.; Appels, L.; Tan, T.; Dewil, R. Bioethanol from lignocellulosic biomass: Current findings determine research priorities. Sci. World J. 2014, 2014, 298153. [Google Scholar] [CrossRef]
- Arévalo, C.; Freer, J.; Naulin, P.A.; Barrera, N.P.; Troncoso, E.; Araya, J.; Peña-Farfal, C.; del P Castillo, R. Study of the ultrastructure of Eucalyptus globulus wood substrates subjected to auto-hydrolysis and diluted acid hydrolysis pre-treatments and its influence on enzymatic hydrolysis. Bioenergy Res. 2017, 10, 714–727. [Google Scholar] [CrossRef]
- Snoek, T.; Verstrepen, K.J.; Voordeckers, K. How do yeast cells become tolerant to high ethanol concentrations? Curr. Genet. 2016, 62, 475–480. [Google Scholar] [CrossRef]
- Hahn-Hägerdal, B.; Karhumaa, K.; Fonseca, C.; Spencer-Martins, I.; Gorwa-Grauslund, M.F. Towards industrial pentose-fermenting yeast strains. Appl. Microbiol. Biotechnol. 2007, 74, 937–953. [Google Scholar] [CrossRef]
- Agbogbo, F.K.; Coward-Kelly, G.; Torry-Smith, M.; Wenger, K.S. Fermentation of glucose/xylose mixtures using Pichia stipitis. Process Biochem. 2006, 41, 2333–2336. [Google Scholar] [CrossRef]
- Agbogbo, F.K.; Coward-Kelly, G. Cellulosic ethanol production using the naturally occurring xylose-fermenting yeast, Pichia stipitis. Biotechnol. Lett. 2008, 30, 1515–1524. [Google Scholar] [CrossRef]
- Pereira, S.R.; Portugal-Nunes, D.J.; Evtuguin, D.V.; Serafim, L.S.; Xavier, A.M.R.B. Advances in ethanol production from hardwood spent sulphite liquors. Process Biochem. 2013, 48, 272–282. [Google Scholar] [CrossRef]
- Slininger, P.J.; Gorsich, S.W.; Liu, Z.L. Culture nutrition and physiology impact the inhibitor tolerance of the yeast Pichia stipitis NRRL Y-7124. Biotechnol. Bioeng. 2009, 102, 778–790. [Google Scholar] [CrossRef]
- Geng, A.; Wang, Z.; Lai, K.S.; Tan, M.W.Y. Bioenergy II: Comparison of laboratory and industrial Saccharomyces cerevisiae strains for their stress tolerance. Int. J. Chem. React. Eng. 2010, 8, A87. [Google Scholar] [CrossRef]
- Silva, J.P.A.; Mussatto, S.I.; Roberto, I.C.; Teixeira, J.A. Fermentation medium and oxygen transfer conditions that maximize the xylose conversion to ethanol by Pichia stipitis. Renew. Energy 2012, 37, 259–265. [Google Scholar] [CrossRef] [Green Version]
- Silva, D.D.V.; Dussán, K.J.; Hernández, V.; Silva, S.S.d.; Cardona, C.A.; Felipe, M.d.G.d.A. Effect of volumetric oxygen transfer coefficient (kLa) on ethanol production performance by Scheffersomyces stipitis on hemicellulosic sugarcane bagasse hydrolysate. Biochem. Eng. J. 2016, 112, 249–257. [Google Scholar] [CrossRef]
- Gutiérrez-Rivera, B.; Ortiz-Muñiz, B.; Gómez-Rodríguez, J.; Cárdenas-Cágal, A.; Domínguez González, J.M.; Aguilar-Uscanga, M.G. Bioethanol production from hydrolyzed sugarcane bagasse supplemented with molasses “B” in a mixed yeast culture. Renew. Energy 2015, 74, 399–405. [Google Scholar] [CrossRef]
- Papini, M.; Nookaew, I.; Uhlén, M.; Nielsen, J. Scheffersomyces stipitis: A comparative systems biology study with the Crabtree positive yeast Saccharomyces cerevisiae. Microb. Cell Fact. 2012, 11, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santosh, I.; Ashtavinayak, P.; Amol, D.; Sanjay, P. Enhanced bioethanol production from different sugarcane bagasse cultivars using co-culture of Saccharomyces cerevisiae and Scheffersomyces (Pichia) stipitis. J. Environ. Chem. Eng. 2017, 5, 2861–2868. [Google Scholar] [CrossRef]
- Sreemahadevan, S.; Singh, V.; Roychoudhury, P.K.; Ahammad, S.Z. Mathematical modeling, simulation and validation for co-fermentation of glucose and xylose by Saccharomyces cerevisiae and Scheffersomyces stipitis. Biomass Bioenergy 2018, 110, 17–24. [Google Scholar] [CrossRef]
- Beck, M.J.; Johnson, R.D.; Baker, C.S. Ethanol production from glucose/xylose mixes by incorporating microbes in selected fermentation schemes. Appl. Biochem. Biotechnol. 1990, 24, 415–424. [Google Scholar] [CrossRef]
- Zambare, V.P.; Bhalla, A.; Muthukumarappan, K.; Sani, R.K.; Christopher, L.P. Bioprocessing of agricultural residues to ethanol utilizing a cellulolytic extremophile. Extremophiles 2011, 15, 611–618. [Google Scholar] [CrossRef]
- Zambare, V.P.; Christopher, L.P. Optimization of enzymatic hydrolysis of corn stover for improved ethanol production. Energy Explor. Exploit. 2012, 30, 193–205. [Google Scholar] [CrossRef] [Green Version]
- Kelbert, M.; Romaní, A.; Coelho, E.; Pereira, F.B.; Teixeira, J.A.; Domingues, L. Simultaneous saccharification and fermentation of hydrothermal pretreated lignocellulosic biomass: Evaluation of process performance under multiple stress conditions. Bioenergy Res. 2016, 9, 750–762. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
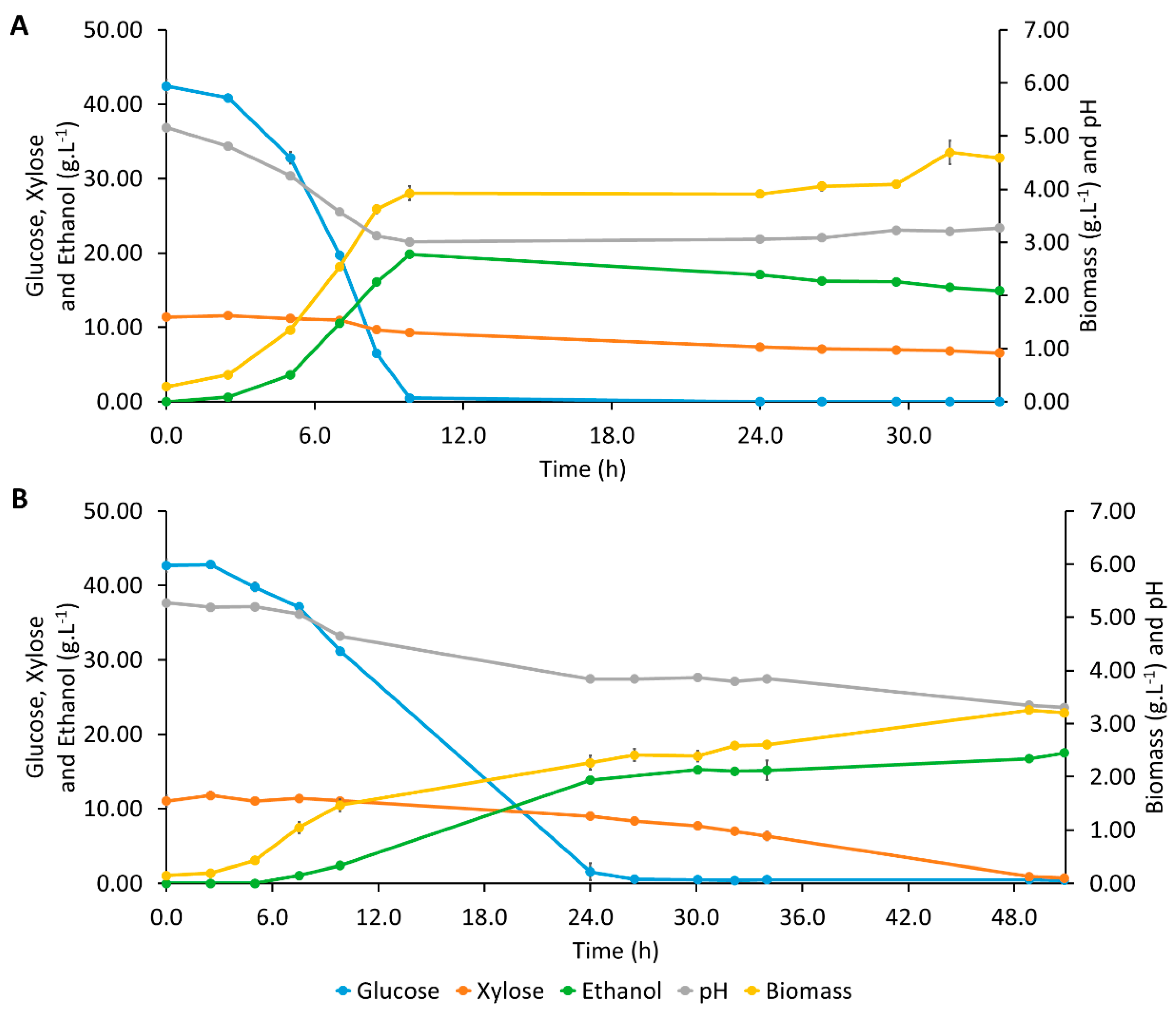{kind=link}
{kind=link}
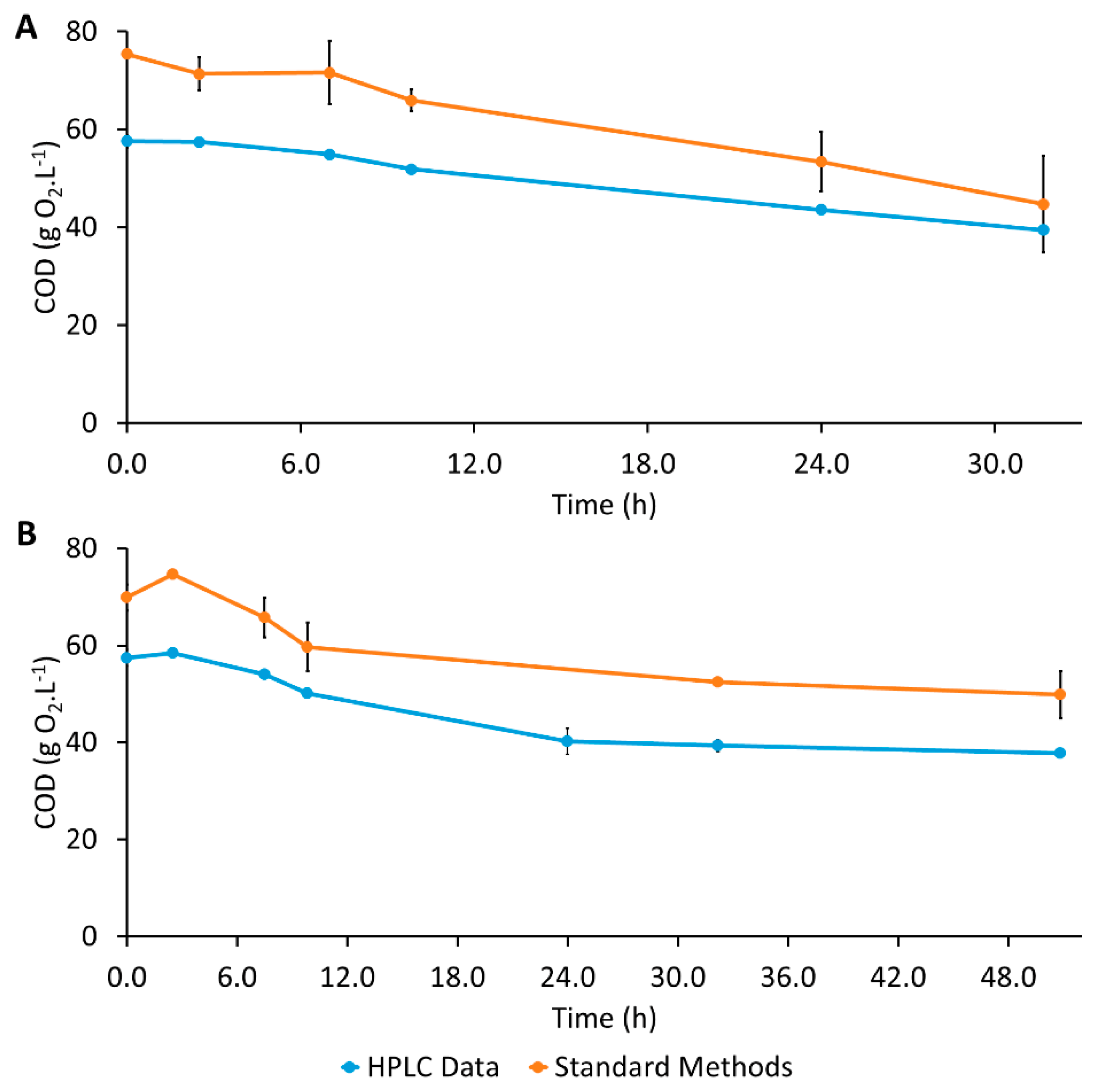{kind=link}
{kind=link}
| Parameter | Erlenmeyer Assays | Bioreactor Assays | ||||
|---|---|---|---|---|---|---|
| S. cerevisiae | S. stipitis | Sequential Co-Culture | Simultaneous Co-Culture | S. cerevisiae | Sequential Co-Culture | |
| µ (h−1) | 0.360 ± 0.008 | 0.284 ± 0.017 | 0.251 | - | ||
| [Ethanol]max (g·L−1) | 19.81 ± 0.15 | 17.50 ± 0.09 | 20.03 ± 0.61 | 20.25 ± 0.29 | 19.24 | 20.13 |
| rglucose (g·L−1·h−1) | 6.91 ± 0.20 | 1.98 ± 0.03 | 5.13 ± 0.19 | 3.83 ± 0.36 | 4.66 | 5.24 |
| rxylose (g·L−1·h−1) | 0.193 ± 0.003 | 0.349 ± 0.017 | 0.309 ± 0.003 | 0.410 ± 0.003 | 0.385 | 0.388 |
| Prodvol (g·L−1·h−1) | 2.01 ± 0.01 | 0.344 ± 0.002 | 1.42 ± 0.05 (a) | 0.754 ± 0.021 | 0.733 | 0.668 |
| Yethanol/substrate (g·g−1) | 0.450 ± 0.009 | 0.333 ± 0.011 | 0.464 ± 0.009 | 0.426 ± 0.016 | 0.433 | 0.391 |
| Ybiomass/substrate (g·g−1) | 0.081 ± 0.004 | 0.058 ± 0.001 | 0.074 ± 0.001 | 0.086 ± 0.002 | 0.091 | 0.060 |
| Conversion efficiency (%) | 88.3 ± 1.7 | 65.3 ± 2.2 | 91.0 ± 1.8 | 83.6 ± 3.1 | 84.8 | 76.6 |
| Consumed sugars (%)(b) | 81.8 ± 0.4 | 97.9 ± 0.2 | 84.3 ± 0.3 | 90.8 ± 0.3 | 84.1 | 86.0 |
| Feedstock | Pretreatment | Config | Microbial Strain | [Ethanol]max (g·L−1) | Prodvol (g·L−1·h−1) | Yethanol/substrate | Ref. |
|---|---|---|---|---|---|---|---|
| E. globulus | Kraft pulping | SHF | PYCC 5246 | 19.24 | 0.733 | 0.433 g·g−1 0.529 g·g d.w. pulp−1 0.275 g·g d.w. wood−1 | This work |
| Corn stover | Extrusion | SHF | ATCC 24860 | 6.2 | 0.086 | 0.47 g·g−1 | [55] |
| S. pectinata | Extrusion | SHF | ATCC 24860 | 7.2 | 0.10 | 0.426 g·g−1 | [56] |
| E. globulus | Kraft pulping | SSF | D5A | 5.67 | 0.032 | 0.042 g·g d.w. wood−1 | [24] |
| E. globulus | Kraft pulping | SSF | IR2T9 (a) | NA | NA | 0.469 g·g d.w. pulp−1 | [15] |
| E. globulus | Autohydrolysis | SHF | PE-2 | 39.7 | 0.551 | 0.38 g·g−1 | [57] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Branco, R.H.R.; Amândio, M.S.T.; Serafim, L.S.; Xavier, A.M.R.B. Ethanol Production from Hydrolyzed Kraft Pulp by Mono- and Co-Cultures of Yeasts: The Challenge of C6 and C5 Sugars Consumption. Energies 2020, 13, 744. https://doi.org/10.3390/en13030744
Branco RHR, Amândio MST, Serafim LS, Xavier AMRB. Ethanol Production from Hydrolyzed Kraft Pulp by Mono- and Co-Cultures of Yeasts: The Challenge of C6 and C5 Sugars Consumption. Energies. 2020; 13(3):744. https://doi.org/10.3390/en13030744
Chicago/Turabian StyleBranco, Rita H. R., Mariana S. T. Amândio, Luísa S. Serafim, and Ana M. R. B. Xavier. 2020. "Ethanol Production from Hydrolyzed Kraft Pulp by Mono- and Co-Cultures of Yeasts: The Challenge of C6 and C5 Sugars Consumption" Energies 13, no. 3: 744. https://doi.org/10.3390/en13030744